
Introduction
Ornamental plants are an important part of horticulture and our culture as a source of beauty and emotional enjoyment, while also contributing to the economy. As living standards continue to improve, the demand for these plants has grown consistently1. In order to meet the growing demands of the market, micropropagation, i.e. the reproduction of plants in vitro, is gaining interest among commercial plant producers2. According to Podwyszyńska et al. (2022), the global annual production of ornamental plants through in vitro cultures has increased from 800 million to 2 billion in the last decade3. Unfortunately, like any other technique, micropropagation also faces several challenges, e.g. contamination, plantlet browning, in vitro rooting difficulty, somaclonal variations, hyperhydricity, shoot tip necrosis, problems with acclimatization4. Some of these problems can be overcome, by the application of nanotechnology.
Nanoparticles (NPs) are a promising tool useful in horticulture, particularly for the production of ornamental plants in vitro. This is due to their unique physiochemical properties, versatility, and the possibility of direct interaction with the plant cell5. For example, silver nanoparticles (Ag NPs), applied at 20–100 ppm to non-meristematic explants (leaves and internodes), increased genetic variation and mutation occurrence in chrysanthemum [Chrysanthemum × morifolium (Ramat.) Hemsl.] plants regenerated through adventitious organogenesis. By those means, it was possible to create novel cultivars of this species with distinctive flower colours and shapes6,7. Kumar et al. (2024) highlight that various nanomaterials modulate plant hormone pathways (including auxin, cytokinin, gibberellin, ethylene, salicylic acid, jasmonic acid, and brassinosteroids), influencing growth, plasticity, and nutrient uptake8. Gold nanoparticles (50–100 ppm Au NPs) significantly improved micropropagation rates and post-acclimatization plant quality in bleeding heart [Lamprocapnos spectabilis (L.) Fukuhara] compared to the control and traditional plant growth regulators (PGRs)9,10. Zinc oxide nanoparticles (25–75 ppm ZnO NPs), on the other hand, elevated phenolic content (p-coumaric, chlorogenic, and caffeic acid) in the bulblets of lilium (Lilium candidum L.)11. Likewise, ZnO NPs treatment increased the levels of anthocyanins, chlorophyll, and flavonoids, as well as root length, leaf length, leaf number, and bulb count in the explants of Lilium ledebourii Bioss12. The results obtained by Krzepiłko et al. indicated that high concentrations of ZnO NPs (30–40 ppm) stimulated the propagation of stevia (Stevia rebaudiana Bertoni) shoots13. On the other hand, NPs negatively affected shoot length, root number and length, as well as the fresh weight of the plantlets13. Nanomaterials also play a significant role in improving plants’ resistance to environmental stress, such as pathogens or drought, by influencing the synthesis of plant hormones and nutrient absorption14. In contrast with the previous reports, Ag NPs inhibited rhizogenesis in chrysanthemum and gerbera (Gerbera × jamesonii H. Bol), as well as adventitious organogenesis in chrysanthemum. Gold nanoparticles (Au NPs) were less toxic and improved the efficiency of Cape Primrose (Streptocarpus × hybridus Voss.) micropropagation15. A newly designed nanohybrid technology that conjugates indole acetic acid (IAA) with nanoparticles of iron (Fe) and manganese (Mn) micronutrients was developed to combat heavy metal stress in strawberry (Fragaria spp. ‘Fertona’)16. Despite their undeniable promise, further studies are needed to fully explore the potential of NPs and their application in the large-scale production of ornamental plants. Notably, little is known about the use of NPs in the production of synthetic seeds.
The synthetic seed technology, developed primarily by Murashige in 1977, is a highly useful technique in the propagation of seedless species, such as chrysanthemum17. In this approach, somatic embryos (usually) or other vegetative parts; such as shoot tips, cell aggregates, auxiliary buds, or any other micropropagules; are artificially encapsulated, sown, and converted into a plant under in vitro or in vivo conditions18. Sodium alginate is the most common type of hydrogel coating used in the synthetic seed technology. It is water-soluble; however, when sodium ions are replaced by calcium ions, ionic cross-binding occurs between calcium and the alginate polymer chains, leading to the formation of an insoluble gel19. This artificial coating mimics natural endosperm, by supplying carbon sources, minerals, and vitamins to support the early development of the encapsulated explants. Moreover, sodium alginate is considered a potential elicitor that improves plants’ tolerance to environmental stress, such as drought or salinity20. Despite the growing interest in synthetic seed technology for mass propagation and conservation of plant genetic resources, the role of nutritional composition within the encapsulation matrix is still poorly explored. Optimization of alginate matrix composition is essential for synthetic seed technology, as inadequate or imbalanced nutrition can reduce germination rate or plant vigour21. Most studies focus on the use of basal Murashige and Skoog (MS) medium22 with traditional plant growth regulators (PGRs) or carbon sources23. Moreover, there is little research on nutrient interactions with novel additives, such as nanoparticles (NPs), particularly in a time-dependent manner24. Supplementation of the artificial alginate matrix with NPs alone or as the carriers of PGRs can improve the performance of synthetic seeds. Iron oxide nanoparticles (Fe3O4 NPs) are particularly useful as iron is an essential micronutrient for plants, significantly affecting crop yield25. As a central atom in iron–sulphur (FeS) proteins, it plays a crucial role in electron transport during respiration and photosynthesis. Iron is a key component of cytochromes and is essential for chlorophyll biosynthesis by activating enzymes, such as cis‑aconitase and glutamyl‑tRNA reductase26. Mitochondria are also abundant in iron-dependent enzymes, with respiratory complexes I and II containing multiple FeS clusters. Iron-containing proteins can also be found in other organelles, i.e. peroxisomes and the endoplasmic reticulum house haem proteins, whereas mono- and di-iron enzymes are found in all cell compartments27. Finally, in response to pathogenic fungal attacks, plants redistribute iron to the apoplast, where its controlled accumulation affects both intracellular and extracellular defence mechanisms, supporting the overall health of plants28. Its deficiency can severely affect plant growth and productivity29. Plants take up iron oxide nanoparticles primarily via root tips through diffusion and membrane-bound carriers, then transport them through apoplastic and symplastic routes to the shoot. From there, it is redistributed to actively growing tissues, with the xylem and phloem serving as the main systems for long-distance iron transport30. In foliar applications, iron oxide NPs can also enter leaves through stomata or cuticle pores and adhere to epidermal surfaces, although this process is less effective than in roots31. Fe3O4 NPs are the most efficient NPs type, compared to γ-Fe2O3 and α-Fe2O3 and bulk Fe3O4. Once inside the cells, Fe3O4 NPs may dissociate, releasing Fe ions that enhance photosynthetic gene expression and antioxidant enzyme activity, resulting in improved plant growth, as observed in field-grown maize31. Kulus et al. reported that the addition of Fe3O4 NPs into the alginate matrix can increase the germination rate of synthetic seeds and the acclimatization efficiency of chrysanthemum plants, however, their effect on the content of metabolites, particularly in the inflorescences, and the development of generative organs remains unknown24. The aims of this study included (1) to investigate the time-dependent effects of Fe3O4 NPs and IAA on the metabolism of germinated synthetic seeds; (2) to verify the genetic stability of chrysanthemum plants under nanoparticle treatment; (3) to optimize the regulatory conditions for ornamental traits (inflorescence diameter).
Results
Morphology studies of NPs
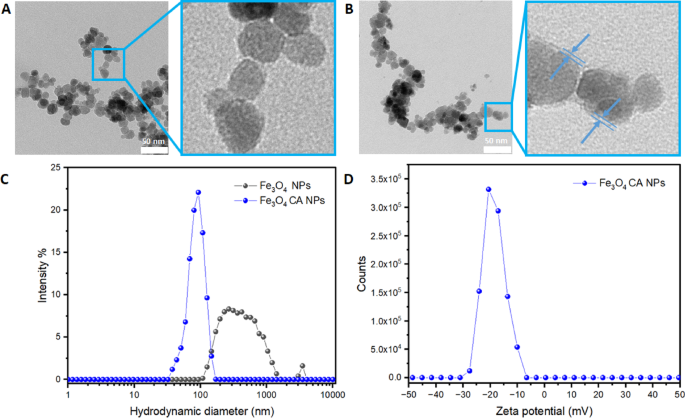
The morphology results obtained with Transmission Electron Microscope (TEM) confirmed the spherical shape of bare Fe3O4 NPs sized below 20 nm (Fig. 1A). As for the Fe3O4CA NPs presented in Fig. 1B, a thin organic layer of citrate on the surface of NPs can be distinguished. The Dynamic Light Scattering (DLS) analysis shown in Fig. 1C revealed that stabilized Fe3O4CA NPs exhibit a monodisperse distribution with a lower hydrodynamic diameter of about 90 nm. The presence of a citrate coating effectively improved dispersion in aqueous media, whereas bare NPs Fe3O4 had a broader hydrodynamic diameter due to the lack of citrates, resulting in agglomeration. The Zeta potential of Fe3O4CA NPs showed a narrow peak centred around −20 mV, indicating a negatively charged surface in the measured conditions (pH 7, 0.01 M NaCl at room temperature).

TEM images of Fe3O4 NPs (A), Fe3O4CA NPs (B), hydrodynamic size distribution for both NPs (C), and Zeta potential for Fe3O4CA NPs (D).
Morphometric analysis of inflorescences
It was found that after 30 days of in vitro culture, plants treated with iron oxide nanoparticles (both bare and stabilized) and a combination of Fe3O4CA NPs and IAA produced inflorescences of greater diameter (7.6–8.5 cm) than the untreated control (6.6 cm) (Fig. 2). In contrast, after 60 days, plants treated with Fe3O4 NPs alone or in combination with IAA had smaller inflorescences (6.1 cm) than the control (7.6 cm). Representative inflorescences of chrysanthemum ‘Richmond’ after various treatments are shown in Fig. 3. The inflorescences maintain a generally stable colour range across all treatments and time points, consistent with the typical purple colour for this cultivar. Minor variations in petal intensity are observed but remain within the expected phenotypic range.

Effect of indole-3-acetic acid (IAA) and iron oxide nanoparticles in bare (Fe3O4 NPs) or citrate-stabilized form (Fe3O4CA NPs), applied singularly or in combination, on the inflorescence diameter of Chrysanthemum × morifolium ‘Richmond’ after 30 and 60 days of treatment. Box-and-whisker plots with the same letters (a, b) are not significantly different (p < 0.05) according to one-way ANOVA followed by Duncan’s multiple range test (n = 98).

Example inflorescences of Chrysanthemum × morifolium ‘Richmond’: control after 30 days of in vitro culture (A), treatment with Fe3O4 NPs for 30 days (B), control after 60 days of in vitro culture (C), treatment with Fe3O4 NPs for 60 days (D). Scale bar: 1 cm.
Biochemical array of leaves and inflorescences
The repeatability of the biochemical methods was assessed using quality control samples analysed in triplicate. Precision was evaluated based on intra-day repeatability and expressed as %RSD (relative standard deviation). The mean RSD values were around 4–11%, indicating acceptable precision of the protocols.
Most IAA- and NP-treated plants had an increased TPC value compared to the control, regardless of treatment duration (Fig. 4). The highest content of polyphenols in the leaves of chrysanthemum plants grown for 30 days was found after incorporating a combination of IAA and Fe3O4CA NPs. As for the plants grown for 60 days, the highest TPC value was detected after the sole application of IAA. In contrast, the lowest TPC value was found in the control and Fe3O4CA NPs treatment.

Effect of indole-3-acetic acid (IAA) and iron oxide nanoparticles in bare (Fe3O4 NPs) or citrate-stabilized form (Fe3O4CA NPs), applied singularly or in combination, on the content of polyphenols in the leaves of Chrysanthemum × morifolium ‘Richmond’ after 30 and 60 days of treatment. Box-and-whisker plots with the same letters (a–c) are not significantly different (p < 0.05) according to one-way ANOVA followed by Duncan’s multiple range test (three biological replicates).
As for the inflorescences of plants treated for 30 days, the highest concentration of polyphenols was reported after applying Fe3O4 NPs and IAA + Fe3O4CA NPs. No significant differences were found between the other treatments and the control (p > 0.05) (Fig. 5). After 60 days, inflorescences of plants treated with IAA + Fe3O4CA NPs and the control had the highest TPC, whereas the Fe3O4 NPs treatment—the lowest.

Effect of indole-3-acetic acid (IAA) and iron oxide nanoparticles in bare (Fe3O4 NPs) or citrate-stabilized form (Fe3O4CA NPs), applied singularly or in combination, on the content of polyphenols in the inflorescences of Chrysanthemum × morifolium ‘Richmond’ after 30 and 60 days of treatment. Box-and-whisker plots with the same letters (a–c) are not significantly different (p < 0.05) according to one-way ANOVA followed by Duncan’s multiple range test (nine biological replicates).
Plants treated for 30 days with Fe3O4 NPs and IAA + Fe3O4CA NPs had a higher concentration of anthocyanins in their inflorescences than the chrysanthemums from other treatments (Fig. 6). The results were somewhat different after 60 days of exposure, i.e. the highest content was found in the untreated control, whereas the lowest – after the application of Fe3O4 NPs.

Effect of indole-3-acetic acid (IAA) and iron oxide nanoparticles in bare (Fe3O4 NPs) or citrate-stabilized form (Fe3O4CA NPs), applied singularly or in combination, on the content of anthocyanins in the inflorescences of Chrysanthemum × morifolium ‘Richmond’ after 30 and 60 days of treatment. Box-and-whisker plots with the same letters (a–d) are not significantly different (p < 0.05) according to one-way ANOVA followed by Duncan’s multiple range test (nine biological replicates).
Genetic stability of plants
A total of 2160 scorable bands were detected by eight RAPD primers in 72 ‘Richmond’ plants (Table 1). Primer R3 generated the highest number of bands (7 per sample), while primer R5 produced only one amplicon per sample. The band profiles of individual plants did not differ among each other, i.e. no polymorphic genotypes were detected by any of the primers (Fig. 7).

Example RAPD band profiles of Chrysanthemum × morifolium ‘Richmond’ (cultured for 30 days) received with primer R1 (upper line) and R2 (bottom line) as a result of auxin and/or nanoparticle treatments. The outermost lanes (wm) serve as DNA base pair (bp) weight markers, while inner lines represent control plants, plants treated with indole-3acetic acid (IAA), iron oxide nanoparticles (Fe3O4 NPs), and/or iron oxide nanoparticles stabilized with citrate (Fe3O4CA NPs).
Discussion
The integration of nanoparticles (NPs) and plant growth regulators has emerged as a promising strategy in plant biotechnology32. Our study confirmed the application of engineered NPs as PGR carriers in the management of synthetic seed practices. Nonetheless, the observed time-dependent effects of iron oxide nanoparticles on inflorescence diameter in chrysanthemum plants highlight complex interactions between NPs treatments and plant developmental stages.
After 30 days of in vitro culture, iron oxide nanoparticles and their combination with IAA significantly increased inflorescence diameter (7.6–8.5 cm vs. 6.6 cm in the control). A similar phenomenon was reported by Srivastava et al. (2022) in calendula (Calendula officinalis L.)33. This is likely due to enhanced nutrient uptake, increased photosynthesis and respiration efficiencies, as well as improved water utilization, stimulated by nanoparticles, described by other researchers34. Similar to our study, Fe3O4 NPs enhanced plant growth of wheat (Triticum aestivum L.) through improved iron and phosphorous uptake and enhanced content of photosynthetic pigments in the leaves. This effect, however, depended on the light intensity during plant growth and the age of the plants35. Au NPs (10 mg L− 1) promoted the antioxidant activities of ascorbate peroxidase, catalase, and guaiacol peroxidase, which in turn resulted in higher fresh and dry biomass, as well as leaf area in wheat36. Silicon NPs stimulated crop development in tomato (Solanum lycopersicum L.) through increased availability and accumulation of macronutrients (nitrogen, potassium, calcium, and sulphur) and micronutrients (iron and manganese), as well as improved drought resistance by optimized water usage efficiency and reduced water loss during transpiration37. Zinc oxide NPs, on the other hand, enhanced growth and nutrient uptake in maize38.
As in the case of biometrical analysis, most of the treatments in the present study generally increased the content of total polyphenols in the leaves and inflorescences of chrysanthemum, with effects depending on the duration of exposure. After 30 days, most treatments stimulated polyphenol and anthocyanin accumulation, especially Fe3O4 NPs and IAA combined with Fe3O4CA NPs. These compounds play crucial roles in plant defense mechanisms but are also appreciated by the phytopharmaceutical industry, therefore their increase is highly beneficial39,40. Similarly, Fe2O3 NPs in combination with endophytes boosted the levels of phenolic compounds, flavonoids, and essential oils in thyme (Thymus vulgaris L.) and Dracocephalum kotschyi Boiss41,42. However, in the present experiment, after a longer exposition of 60 days, a decline in these compounds was observed in most cases. This negative effect of prolonged treatments on both the development of plants and their phytochemical composition could be related to NPs accumulation; iron toxicity or oxidative stress, disrupting metabolic pathways43. Excessive iron content can catalyse the Fenton reaction, producing highly reactive hydroxyl radicals (•OH) that induce oxidative stress. These reactive oxygen species (ROS) can damage lipids, proteins, and DNA, including enzymes involved in phenylpropanoid biosynthesis – the primary pathway responsible for polyphenol production44. At the same time, iron overload may disturb IAA homeostasis, affecting auxin signalling and gene expression, including members of the auxin/induced-3-acetic acid (Aux/IAA) family45. These genes are essential for auxin-regulated transcriptional repression that affects plant growth and resistance to abiotic stresses46. Altered Aux/IAA expression under iron-induced stress may shift metabolic priorities toward cell survival and stress mitigation, rather than secondary metabolism. Moreover, IAA-NPs interactions might overstimulate early growth, reducing resources for later reproductive stages47. The observed differences in the effect of Fe3O4 NPs and Fe3O4CA NPs, such as sustained polyphenol content after 60 days in plants treated with Fe3O4CA NPs, can be explained by surface modification (citrate coating) of nanoparticles. Citrate-coated NPs have increased colloidal stability and higher aqueous solubility, which enhances their bioavailability and facilitates more controlled cellular uptake48,49. The negatively charged citrate layer reduces aggregation and may limit uncontrolled ROS generation by minimizing rapid iron ion release, consequently reducing oxidative stress as observed in soybean (Glycine max L.) or alfalfa (Medicago sativa L.) plants50. Răcuciu et al. (2022) confirmed a positive influence of Fe3O4 NPs coated with aspartic acid on the performance of maize (Zea mays L.) seedlings51. On the other hand, no effect of Fe3O4 NPs on the primary photochemical processes was observed in wheat, regardless of the experimental factors studied; NPs concentration and light intensity35. This highlights the species-dependent reaction of plants to similar treatments.
Stability verification of the in vitro-derived material is of paramount importance. RAPD markers are a valuable tool for identifying genetic variation within chrysanthemum, due to their simplicity, low cost, and high efficiency corroborated by numerous studies6,52,53. The performed here analyses confirmed the genetic and phenotypic homogeneity of plants, regardless of IAA or NPs treatment. Our results correspond with Sun et al.54, who claimed that Fe3O4 NPs had no evident phytotoxicity even at the level of 1000 mg L− 1 in mung bean (Vigna radiata (L.) R. Wilczek). This is extremely important since nanoparticles are known to induce genotoxic effects on plants if used at suboptimal concentrations55. Research indicates that nanoparticles, such as Ag NPs and ZnO NPs, can penetrate plant cells and generate reactive oxygen species that contribute to oxidative stress and subsequent DNA damage. For example, exposure to Ag NPs resulted in increased DNA fragmentation and chromosomal aberrations in faba bean (Vicia faba L.) and tobacco (Nicotiana tabacum L.)56,57. In a study by Fouda et al. (2021), Fe3O4 NPs showed the lowest genetic toxicity in Salvadora persica L. callus cultures, causing only minimal changes in genomic template stability (GTS) compared to ZnO and SiO2 nanoparticles58. The extent of genotoxicity, however, is influenced by several factors, i.e. nanoparticle size, concentration, and exposure duration. Apparently, iron oxide nanoparticles applied at low concentrations (7.7 mg L− 1) do not cause adverse effects on the DNA and can be safely used in synthetic seed technology. Likewise, plant growth regulators (e.g. auxins) may stimulate the development of callus and occurrence of somaclonal variation, reported also in chrysanthemum59. In the present study, no indirect regeneration happened, therefore stability was maintained. This is crucial for the commercial propagation of plants. Nevertheless, it should be highlighted that RAPDs are dominant markers, which may miss recessive mutations. In future studies it is recommended to supplement SSR or SNP marker verification. Propagating genetically and phenotypically stable ornamental plants ensures predictable, high-quality products, efficient production, intellectual property protection, and meeting consumer demands60.
In summary, this study provides a promising nano-regulation strategy for the large-scale production of synthetic seeds in ornamental plants. By using iron oxide nanoparticles in tissue culture systems of chrysanthemum, it is possible to achieve sustained-release effects that enhance PGR stability and bioavailability, reduce the frequency of media supplementation, and lower overall chemical reagent consumption. Our findings open new pathways in commercial plant biotechnology.
Materials and methods
Nanoparticles utilized in the experiment were synthesized (using the chemical co-precipitation method), modified (citrate-coated), and characterized at the Institute of Fundamental Technological Research, Polish Academy of Sciences, as described by Kulus et al.24. The experiment on plants was performed in the Laboratory of Horticulture and Landscape Architecture, and the Department of Agronomy and Food Technology, Bydgoszcz University of Science and Technology, Poland.
Characterization of nanoparticles
The morphology of nanoparticles was studied using Transmission Electron Microscopy Libra 120 (Carl Zeiss, Stuttgart, Germany). The aqueous suspensions of NPs were placed onto the copper mesh with a Formvar layer and dried overnight. The Zeta potential was studied using Zetasizer Nano Sizer (Malvern Instruments, Malvern, UK).
Preparation, storage, and conversion of synthetic seeds
Nodal segments (3–4 mm long) with a single axillary bud of chrysanthemum [Chrysanthemum × morifolium (Ramat.) Hemsl.] ‘Richmond’ were used in the experiment. The plants were obtained from our gene bank. Prof. Marek Jerzy undertook the formal identification of the plant material used. A voucher specimen (no. 1594) was deposited in the Herbarium of the Department of Botany, Ecology and Landscape Architecture, Bydgoszcz University of Science and Technology, Poland. No approvals were needed to conduct research with this plant material. Explants were immersed for 10 min in 3% sodium alginate. The alginate solution was prepared on MS medium salts22, without CaCl2, with the addition of 3% sucrose, iron oxide nanoparticles (bare Fe3O4 NPs or stabilized with citrate Fe3O4CA NPs; 7.7 mg L− 1) and/or indole-3-acetic acid (IAA; 1 mg L− 1) obtained from Sigma-Aldrich, Darmstadt, Germany. The concentrations of NPs and IAA were chosen based on our previous studies24. Alginate solutions with NPs were placed for 30 min in the Elmasonic S80(H) Ultrasonic Cleaner (37 kHz, 150 W; Elma Schmidbauer GmbH, Singen, Germany) for proper nanoparticle dispersion. Next, the beads were hardened in a 0.1 M CaCl2 solution for 30 min and rinsed thrice with distilled sterile water. A control group without NPs or IAA was also included. The obtained synthetic seeds were inoculated onto a water-agar medium in a 90-mm Petri dish sealed with parafilm. The cultures were stored in a refrigerator at 4 °C in the dark for two months and then transferred to the growth room for 30 or 60 days (16-h photoperiod, 22 °C).
Acclimatization and biochemical array of leaves and inflorescences
The initially germinated synthetic seeds were sown in a greenhouse, in a mixture of peat and perlite (2:1), provided by Hartmann (Poznań, Poland), in multi-pot trays. Next, the plants were transferred to pots (16 cm diameter) filled with the same substrate and brought to full flowering. At that time, the diameter of 98 inflorescences was measured with a ruler.
The determination of total polyphenol content (TPC) in the leaves was carried out in triplicates according to the method by Podsędek et al.61 using a Dionex UltiMate 3000 set (4-component pump, autosampler, with sample chamber thermostated, diode array UV detector incl. 3D spectrum collection option, RI detector). The separation was carried out on a Kinetex C18 column (5 μm, 150 × 4.6 mm), using a gradient separation. A mixture of 6% acetic acid in 2 mmol L− 1 acetate and 100% acetonitrile (Sigma Aldrich) was used as a solvent. The qualitative assessment was carried out based on retention times and the spectra of pure standards. For polyphenolic compounds for which external standards were not available, the content is presented as the equivalent of related substances; chlorogenic acid (Sigma Aldrich). The results were presented in mg g− 1 fresh weight (FW).
Anthocyanins were extracted from 200 mg fresh weight (FW) of inflorescence samples using methanol containing 1% HCl, according to the Harborne (1984) method62. The same extract was used to analyze the total polyphenol content (TPC) with the Folin-Ciocalteau reagent, according to Waterhouse (2001)63. Absorbance was measured using a NanoPhotometer NP80 (Implen, München, Germany) at a wavelength of 530 nm (anthocyanins) and 765 nm (TPC). Nine replicates from each experimental treatment were included. Results were calculated based on a calibration curve prepared for gallic acid equivalent (GAE) and the molar extinction coefficient (ε) of cyanidin-3-O-glucoside (C3G), and expressed in mg g− 1 FW.
Genetic stability analysis
The genetic fidelity of 72 ex vitro-grown shoots (six plants from each of the six experimental treatments and two treatment durations) was assessed using randomly amplified polymorphic DNA (RAPD). Total genomic DNA was isolated from fresh leaves using a Genomic Mini AX Plant Spin kit (A&A Biotechnology, Gdańsk, Poland) according to the manufacturer’s protocol. The isolated DNA was stored at − 80 °C in a laboratory deep-freezer U101 Innova (Eppendorf, Hamburg, Germany).
Eight primers were used for the PCR reaction. A ready-to-use mix for PCR for GC-rich templates was used, containing Taq DNA polymerase, stabilizers, and PCR anti-inhibitors (A&A Biotechnology). The amplification was carried out in a C1000 Touch thermal cycler (Bio-Rad Laboratories, Hercules, CA, USA) with the program described by Miler et al.52.
The amplified DNA fragments were separated on a 1.5% (w/v) DN- and RNase-free agarose gel in TBE buffer at 110 V for 60 min (Biometra P25, Jena, Germany) and visualized by staining with ethidium bromide (BrEt). Gel images were captured using a GelDoc XR + Gel Documentation System (Bio-Rad) UV transilluminator with Image Lab 4.1 software (Bio-Rad). The molecular weights of the fragments were estimated using a GeneRuler Express DNA Ladder (100–5000 bp) from Thermo Scientific, Waltham, MA, USA.
The banding patterns were documented as a binary matrix, with “1” indicating the presence and “0” the absence of a specific fragment. For each primer tested, the total number of bands, monomorphic, polymorphic (present in the electrophoretic profile of more than one individual), and specific loci (present in the electrophoretic profile of a single individual) were counted.
Statistical analysis
A total of six experimental combinations were included, i.e. alginate + IAA, alginate + Fe3O4 NPs, alginate + Fe3O4CA NPs, alginate + IAA + Fe3O4 NPs, alginate + IAA + Fe3O4CA NPs, alginate only (control). The study was set in a completely randomized design.
The obtained results were statistically analysed through one-way ANOVA, and means were compared with Duncan’s multiple range test (p ≤ 0.05), using Statistica 12.0 software (StatSoft, Tulsa, OK, USA).
Conclusions
This study demonstrates that iron oxide nanoparticles and IAA can affect the metabolic activity and flower quality of Chrysanthemum × morifolium plants derived from synthetic seeds. The size of inflorescences was higher following shorter exposure to nanoparticles but decreased with extended treatment. Likewise, NPs and IAA generally increased the content of polyphenols and anthocyanins in the plants, especially after shorter application. Importantly, the RAPD analysis confirmed the genetic uniformity of plants, indicating that the studied factors did not cause mutations. Among the studied nanoparticle forms, citrate-stabilized iron oxide NPs in combination with IAA most effectively enhance phytochemical content and flower quality in chrysanthemum, presenting a promising nano-regulation approach. However, the duration of application is critical, as prolonged exposure may reduce these beneficial effects. Since only one chrysanthemum cultivar was tested and the transport pathway of nanoparticles in plants was not explored, further research is needed to better understand the mechanisms of NPs (inter)action and optimize treatment protocols for practical use in plant biotechnology. These should include comparative studies of multiple cultivars, transcriptomic analysis to reveal metabolic regulatory pathways, and development of nanoparticle sustained-release systems.
Data availability
Data that support the findings of this study are available from the public repository: https://doi.org/10.18150/TAS3FB.
References
-
Zeng, Z. et al. Nanomaterials: Cross-disciplinary applications in ornamental plants. Nanotechnol. Rev. 13 (1), 20240049 https://doi.org/10.1515/ntrev-2024-0049 (2024).
-
Yasemin, S. & Beruto, M. A review on flower bulb micropropagation: challenges and opportunities. Horticulturae. 10 (3), 284 https://doi.org/10.3390/horticulturae10030284 (2024).
-
Podwyszyńska, M., Orlikowska, T., Trojak-Goluch, A. & Wojtania, A. Application and improvement of in vitro culture systems for commercial production of ornamental, fruit, and industrial plants in Poland. Acta Soc. Bot. Pol. 91, 914 https://doi.org/10.5586/asbp.914 (2022).
-
Abdalla, N. et al. An academic and technical overview on plant micropropagation challenges. Horticulturae. 8 (8), 677 https://doi.org/10.3390/horticulturae8080677 (2022).
-
Mgadi, K., Ndaba, B., Roopnarain, A., Rama, H. & Adeleke, R. Nanoparticle applications in agriculture: overview and response of plant-associated microorganisms. Front. Microbiol. 15, 1354440 https://doi.org/10.3389/fmicb.2024.1354440 (2024).
-
Tymoszuk, A. & Kulus, D. Silver nanoparticles induce genetic, biochemical, and phenotype variation in chrysanthemum. Plant. Cell. Tiss. Organ. Cult. 143, 331–344 https://doi.org/10.1007/s11240-020-01920-4 (2020).
-
Tymoszuk, A. & Kulus, D. Effect of silver nanoparticles on the in vitro regeneration, biochemical, genetic, and phenotype variation in adventitious shoots produced from leaf explants in chrysanthemum. Int. J. Mol. Sci. 23 (13), 7406 https://doi.org/10.3390/ijms23137406 (2022).
-
Kumar, D. et al. Review on interactions between nanomaterials and phytohormones: novel perspectives and opportunities for mitigating environmental challenges. Plant. Sci. 340, 111964 https://doi.org/10.1016/j.plantsci.2023.111964 (2024).
-
Kulus, D., Tymoszuk, A., Jedrzejczyk, I. & Winiecki, J. Gold nanoparticles and electromagnetic irradiation in tissue culture systems of bleeding heart: biochemical, physiological, and (cyto)genetic effects. Plant. Cell. Tiss Organ. Cult. 149, 715–734 https://doi.org/10.1007/s11240-022-02236-1 (2022).
-
Kulus, D. Influence of growth regulators on the development, quality, and physiological state of in vitro-propagated Lamprocapnos spectabilis (L.) Fukuhara. Vitro Cell. Dev. Biol. Plant. 56, 447–457 https://doi.org/10.1007/s11627-020-10064-1 (2020).
-
Pałka, P., Muszyńska, B., Szewczyk, A. & Pawłowska, B. Elicitation and enhancement of phenolics synthesis with zinc oxide nanoparticles and led light in Lilium candidum L. cultures in vitro. Agronomy. 13 (6), 1437 https://doi.org/10.3390/agronomy13061437 (2023).
-
Chamani, E., Ghalehtaki, K., Mohebodini, S., Ghanbari, A. & M. & The effect of zinc oxide nano particles and humic acid on morphological characters and secondary metabolite production in Lilium ledebourii Bioss. Iran. J. Genet. Plant. Breed. 4 (2), 11–19 (2015).
-
Krzepiłko, A., Prażak, R., Matyszczuk, K. & Dyduch-Siemińska, M. The effect of zinc oxide nanoparticles on the growth and development of Stevia plants cultured in vitro. Acta Sci. Pol. Hortorum Cultus. 23 (3), 43–56 https://doi.org/10.24326/asphc.2024.5354 (2024).
-
Zhou, X. et al. Nanoparticles: a promising tool against environmental stress in plants. Front. Plant. Sci. 15, 1509047 https://doi.org/10.3389/fpls.2024.1509047 (2025).
-
Tymoszuk, A. & Miler, N. Silver and gold nanoparticles impact on in vitro adventitious organogenesis in chrysanthemum, Gerbera and cape primrose. Sci. Hortic. 257, 108766 https://doi.org/10.1016/j.scienta.2019.108766 (2019).
-
Hamed, H. A., Abeed, A. H. A., Geioushy, R. A., Fouad, O. A. & El-Mahdy, M. T. Innovative auxin-micronutrient based nanocomposites (IAA-Fe2O3NPs and IAA-Mn2O3NPs) shield strawberry plants from lead toxicity. Plant. Physiol. Biochem. 219, 109429 https://doi.org/10.1016/j.plaphy.2024.109429 (2025).
-
Murashige, T. Plant cell and organ cultures as horticultural practices. Acta Hortic. 78, 17–30 https://doi.org/10.17660/ActaHortic.1977.78.1 (1977).
-
Rihan, H., Kareem, F., El-Mahrouk, M. & Fuller, M. Artificial seeds (principle, aspects and applications). Agronomy. 7 (4), 71 https://doi.org/10.3390/agronomy7040071 (2017).
-
Škoro, N., Živković, S., Jevremović, S. & Puač, N. Treatment of chrysanthemum synthetic seeds by air SDBD plasma. Plants. 11, 907 https://doi.org/10.3390/plants11070907 (2022).
-
Golkar, P., Taghizadeh, M. & Noormohammadi, A. Effects of sodium alginate elicitation on secondary metabolites and antioxidant activity of safflower genotypes under in vitro salinity stress. In Vitro Cell. Dev. Biol. Plant. 55, 527–538 https://doi.org/10.1007/s11627-019-10008-4 (2019).
-
Salachna, P., Grzeszczuk, M., Meller, E. & Soból, M. Oligo-alginate with low molecular mass improves growth and physiological activity of Eucomis autumnalis under salinity stress. Molecules. 23 (4), 812 https://doi.org/10.3390/molecules23040812 (2018).
-
Murashige, T. & Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15, 473–497 https://doi.org/10.1111/j.1399-3054.1962.tb08052.x (1962).
-
Kulus, D. Application of synthetic seeds in propagation, storage, and preservation of Asteraceae plant species. In Synthetic Seeds (eds Faisal, M. & Alatar, A.) 155–179 https://doi.org/10.1007/978-3-030-24631-0_6 (Springer, 2019).
-
Kulus, D., Tymoszuk, A., Kulpińska, A. & Osial, M. Synergistic effects of iron oxide nanoparticles and indole-3-acetic acid on the germination and development of cold-stored chrysanthemum synthetic seeds. Plant. Cell. Tiss Organ. Cult. 160, 18 https://doi.org/10.1007/s11240-024-02955-7 (2025).
-
Briat, J. F., Curie, C. & Gaymard, F. Iron utilization and metabolism in plants. Curr. Opin. Plant. Biol. 10 (3), 276–282 https://doi.org/10.1016/j.pbi.2007.04.003 (2007).
-
Tripathi, D. K. et al. Acquisition and homeostasis of iron in higher plants and their probable role in abiotic stress tolerance. Front. Environ. Sci. 5, 86 https://doi.org/10.3389/fenvs.2017.00086 (2018).
-
Connorton, J. M., Balk, J. & Rodríguez-Celma, J. Iron homeostasis in plants—a brief overview. Metallomics 9 (7), 813–823 https://doi.org/10.1039/c7mt00136c (2017).
-
Greenshields, D. L., Liu, G. & Wei, Y. Roles of iron in plant defence and fungal virulence. Plant. Signal. Behav. 2 (4), 300–302 https://doi.org/10.4161/psb.2.4.4042 (2007).
-
Celletti, S. et al. Evaluation of a legume-derived protein hydrolysate to mitigate iron deficiency in plants. Agronomy. 10 (12), 1942 https://doi.org/10.3390/agronomy10121942 (2020).
-
Ning, X. et al. Research progress on iron absorption, transport, and molecular regulation strategy in plants. Front. Plant. Sci. 14, 1190768 https://doi.org/10.3389/fpls.2023.1190768 (2023).
-
Yousaf, N. et al. Characterization of root and foliar-applied iron oxide nanoparticles (α-Fe₂O₃, γ-Fe₂O₃, Fe₃O₄, and bulk Fe₃O₄) in improving maize (Zea mays L.) performance. Nanomaterials. 13 (23), 3036 https://doi.org/10.3390/nano13233036 (2023).
-
Mosqueda-Frómeta, O. et al. Mini-review: application of nanoparticles in plant biotechnology and agriculture. Vitro Cell. Dev. Biol. Plant. https://doi.org/10.1007/s11627-025-10539-z (2025).
-
Srivastava, R. K., Tarakeshwari, K. R., Bhandari, N. S. & Chand, S. Effect of nano and macro iron sprays on growth, flowering, seed and oil yielding attributes in calendula (Calendula officinalis L). J. Hortic. Sci. 17 (2), 353–362 https://doi.org/10.24154/jhs.v17i2.1132 (2022).
-
Satya, S. et al. The vital role of nanoparticles in enhancing plant growth and development. Eng. Proc. 67(1), 48 https://doi.org/10.3390/engproc2024067048 (2024).
-
Feng, Y. et al. Effects of iron oxide nanoparticles (Fe3O4) on growth, photosynthesis, antioxidant activity and distribution of mineral elements in wheat (Triticum aestivum) plants. Plants. 11 (14), 1894 https://doi.org/10.3390/plants11141894 (2022).
-
Manaf, A. et al. Antioxidant enzyme activities correlated with growth parameters of wheat sprayed with silver and gold nanoparticle suspensions. Agronomy. 11 (8), 1494 https://doi.org/10.3390/agronomy11081494 (2021).
-
Rea, R. S., Islam, M. R., Rahman, M. M., Nath, B. & Mix, K. Growth, nutrient accumulation, and drought tolerance in crop plants with silicon application: A review. Sustainability. 14 (8), 4525 https://doi.org/10.3390/su14084525 (2022).
-
Seleiman, M. F., Ahmad, A., Alhammad, B. A. & Tola, E. Exogenous application of zinc oxide nanoparticles improved antioxidants, photosynthetic, and yield traits in salt-stressed maize. Agronomy. 13 (10), 2645 https://doi.org/10.3390/agronomy13102645 (2023).
-
Rana, A., Samtiya, M., Dhewa, T., Mishra, V. & Aluko, R. E. Health benefits of polyphenols: A concise review. J. Food Biochem. 46 (10), e14264 https://doi.org/10.1111/jfbc.14264 (2022).
-
Stiller, A. et al. From fighting critters to saving lives: polyphenols in plant defense and human health. Int. J. Mol. Sci. 22 (16), 8995 https://doi.org/10.3390/ijms22168995 (2021).
-
Khanizadeh, P., Mumivand, H., Morshedloo, M. R. & Maggi, F. Application of Fe2O3 nanoparticles improves the growth, antioxidant power, flavonoid content, and essential oil yield and composition of Dracocephalum kotschyi Boiss. Front. Plant. Sci. 15, 1475284 https://doi.org/10.3389/fpls.2024.1475284 (2024).
-
Kamyab, A., Samsampour, D., Ahmadinasab, N. & Bagheri, A. The combination of nanoparticles and endophytes boosts thyme (Thymus vulgaris L.) resistance to drought stress by elevating levels of phenolic compounds, flavonoids, and essential oils. Chem. Biol. Technol. Agric. 11, 163 https://doi.org/10.1186/s40538-024-00682-x (2024).
-
Mathur, P., Chakraborty, R., Aftab, T. & Roy, S. Engineered nanoparticles in plant growth: phytotoxicity concerns and the strategies for their attenuation. Plant. Physiol. Biochem. 199, 107721 https://doi.org/10.1016/j.plaphy.2023.107721 (2023).
-
Connolly, E. L. & Guerinot, M. L. Iron stress in plants. Genome Biol. 3, reviews10241 https://doi.org/10.1186/gb-2002-3-8-reviews1024 (2002).
-
Wu, T. et al. Induction of root Fe(III) reductase activity and proton extrusion by iron deficiency is mediated by auxin-based systemic signalling in Malus xiaojinensis. J. Exp. Bot. 63 (2), 859–870 https://doi.org/10.1093/jxb/err314 (2012).
-
Zhang, J., Li, S., Gao, X., Liu, Y. & Fu, B. Genome-wide identification and expression pattern analysis of the Aux/IAA (auxin/indole-3-acetic acid) gene family in alfalfa (Medicago sativa) and the potential functions under drought stress. BMC Genom. 25, 382 https://doi.org/10.1186/s12864-024-10313-2 (2024).
-
Kulus, D., Tymoszuk, A., Gościnna, K. & Osial, M. Enhancing germination and growth of chrysanthemum synthetic seeds through iron oxide nanoparticles and indole-3-acetic acid: impact of treatment duration on metabolic activity and genetic stability. Nanotechnol. Sci. Appl. 18, 139–155 https://doi.org/10.2147/nsa.s503868 (2025).
-
Valdeperez, D. et al. Colloidal stability of polymer coated zwitterionic Au nanoparticles in biological media. Inorg. Chim. Acta. 534, 120820 https://doi.org/10.1016/j.ica.2022.120820 (2022).
-
Sun, X. D. et al. Magnetite nanoparticle coating chemistry regulates root uptake pathways and iron chlorosis in plants. Proc. Natl. Acad. Sci. U.S.A. 120 (27) https://doi.org/10.1073/pnas.2304306120 (2023).
-
Iannone, M. F. et al. Magnetite nanoparticles coated with citric acid are not phytotoxic and stimulate soybean and alfalfa growth. Ecotoxicol. Environ. Saf. 211, 111942 https://doi.org/10.1016/j.ecoenv.2021.111942 (2021).
-
Răcuciu, M., Tecucianu, A. & Oancea, S. Impact of magnetite nanoparticles coated with aspartic acid on the growth, antioxidant enzymes activity and chlorophyll content of maize. Antioxidants. 11 (6), 1193 https://doi.org/10.3390/antiox11061193 (2022).
-
Miler, N., Jędrzejczyk, I., Trafara, P. & Winiecki, J. Effect of high-energy ionizing radiation on the DNA content and genetic variation in chrysanthemum plants regenerated from irradiated ovaries. Acta Sci. Pol. Hortorum Cultus. 22 (4), 117–134 https://doi.org/10.24326/asphc.2023.4912 (2023).
-
Sehrawat, S. K., Kumar, R., Dahiya, D. S., Boora, K. S. & Yadav, R. DNA fingerprinting of Chrysanthemum cultivars using RAPDs. Acta Hortic. 624, 479–485 https://doi.org/10.17660/ActaHortic.2003.624.66 (2003).
-
Sun, Y. et al. Phytotoxicity of iron-based materials in mung bean: seed germination tests. Chemosphere. 251, 126432 https://doi.org/10.1016/j.chemosphere.2020.126432 (2020).
-
Marmiroli, M., Marmiroli, N. & Pagano, L. Nanomaterials induced genotoxicity in plant: methods and strategies. Nanomaterials. 12 (10), 1658 https://doi.org/10.3390/nano12101658 (2022).
-
Patlolla, A. K., Berry, A., May, L. & Tchounwou, P. B. Genotoxicity of silver nanoparticles in Vicia faba: A pilot study on the environmental monitoring of nanoparticles. Int. J. Environ. Res. Public. Health. 9 (5), 1649–1662 https://doi.org/10.3390/ijerph9051649 (2012).
-
Lovecká, P. et al. Genomic damage induced in Nicotiana tabacum L. plants by colloidal solution with silver and gold nanoparticles. Plants. 10 (6), 1260 https://doi.org/10.3390/plants10061260 (2021).
-
Fouda, M. S. et al. Nanoparticles induce genetic, biochemical, and ultrastructure variations in Salvadora persica callus. J. Genet. Eng. Biotechnol. 19 (1), 27 https://doi.org/10.1186/s43141-021-00124-3 (2021).
-
Miler, N. & Zalewska, M. Somaclonal variation of chrysanthemum propagated in vitro from different explant types. Acta Sci. Pol. Hortorum Cultus. 13 (2), 69–82 (2014).
-
Roberto, S. R. & Colombo, R. C. Innovation in propagation of fruit, vegetable and ornamental plants. Horticulturae. 6 (2), 23 https://doi.org/10.3390/horticulturae6020023 (2020).
-
Podsędek, A., Sosnowska, D., Redzynia, M. & Anders, B. Antioxidant capacity and content of Brassica oleracea dietary antioxidants. Int. J. Food Sci. Technol. 41 (1), 49–58 https://doi.org/10.1111/j.1365-2621.2006.01260.x (2006).
-
Harborne, J. B. Phytochemical Methods (Springer, 1984). https://doi.org/10.1007/978-94-009-5570-7
-
Waterhouse, A. L. Determination of Total Phenolics. In Current Protocols in Food Analytical Chemistry (ed. Wrolstad, R. E.) I1.1.1–I1.1.8. (Wiley, 2001).
Acknowledgements
The authors wish to thank M.Sc. Alicja Kulpińska for her help in cultivating the plants in the glasshouse.
Funding
This research received no external funding.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kulus, D., Tymoszuk, A., Cichorek, M. et al. Fe3O4 nanoparticles and IAA auxin affect secondary metabolism over time without altering genetic stability in chrysanthemum. Sci Rep 15, 27934 (2025). https://doi.org/10.1038/s41598-025-13279-7
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-13279-7