Introduction
Potato (Solanum tuberosum L.) is an annual herb, belongs to Solanaceae family. The major color of tuber-skin and -flesh are yellow, red, pink, and purple in colored potato, which rich in anthocyanin. Anthocyanins are a class of water-soluble pigments widely present in plants, which determines the color of flowers, leaves, fruits, stems and seed coats. A large number of studies have shown that anthocyanins had the property of anti-mutagenesis and antioxidant1, and played important roles in antibacterial and anti-inflammatory, prevention of cardiovascular diseases and inhibition of tumor cytogenesis2,3. The anthocyanin biosynthesis is mainly regulated by two types of genes, including structural genes, which encode enzymes in anthocyanin pathway, such as Phenylalanine synthetase (PAL), Chalcone synthetase (CHS), Anthocyanin synthetase (ANS), and regulatory genes, which encode transcription factors (TFs) that regulate anthocyanin biosynthesis by regulating structural gene expression. Previous studies have shown that three kinds of transcription factors: MYB transcription factor, bHLH transcription factor and WD40 repeat protein, played essential roles in anthocyanin biosynthesis4, they usually function independently or forming a MBW complex.
The MYB proteins are characterized by a highly conserved DNA-binding domain (MYB) at the N-terminus and a variable regulatory region at the C-terminus. Each MYB repeat consists of 50–53 amino acid residues encoding three α-helices and special DNA motifs5. According to the number of adjacent MYB repeats in DNA-binding domains, the MYB family can be classified into four major groups, namely 1R-MYB (MYB-related), 2R-MYB (R2R3-MYB), 3R-MYB (R1R2R3-MYB) and 4R-MYB6. V-myb myeloblastosis viral oncogene homolog (avian) is the firstly identified MYB transcription factor7. Subsequently, MYB genes were identified in animals, plants, and fungi. So far, the MYB TFs have been comprehensively identified in a number of plant genome based on substantial genome sequence data, such as Arabidopsis, rice and Gossypium hirsutum8,9,10. A total of 158, 233 and 217 StMYB genes were identified in DM potato genome with the earliest version of whole genome sequencing data (DM v4.03, The PGSC 2011)11,12,13. However, MYB family genes in DM potato genome with the latest version of whole genome sequencing data (DM v6.1, The PGSC 2020) have not yet been comprehensively identified and systematically analyzed.
In higher plants studied to date, a large amount of TFs involved in anthocyanin biosynthesis have been identified, especially MYB regulators. The first plant transcription factor involved in the flavonoid pigment pathway, colorless 1 (C1), belong to 2R-MYB family, was cloned from Zea mays14,15. In Arabidopsis, the overexpression of PAP1 (AtMYB75) and PAP2 (AtMYB90) significantly promote flavonol and anthocyanin accumulation16. In petunia, AN2 and AN4 responsible for the pigmentation patterns of petal limb, and petal tube and anthers, respectively17. The over-expression of DPL and PHZ significantly enhanced the anthocyanin accumulation in the leaves and flower tube18. In apple, overexpressed MdMYB10a gene in tobacco plants results in substantial anthocyanin accumulation19. Several activators have been also identified in tomato, such as SlAN2, SlAN2-like, SlANT1, SlANT1-like, and SlMYB7520,21. In addition to the activators mentioned above, several MYB TFs have an antagonistic inhibitory effect on the regulation of anthocyanin metabolism22. The first anthocyanin inhibitor, AmMYB308, was isolated from snapdragon, and its overexpression reduced the pigmentation of tobacco corollas23. Subsequently, the inhibitors of anthocyanin synthesis were identified in many other species. For example, Arabidopsis AtMYB6024, strawberry FaMYB125, tomato SlMYB-ATV26 and SlTRY27, petunia PhMYB2728, and apple MdMYB1629 were also found to be inhibitors of anthocyanin synthesis.
At present, anthocyanin biosynthesis in potato has received extensive attention. Multiple structural genes related to anthocyanin biosynthesis have been cloned successively in potato, including CHS, F3H, F3′5′H, DFR, ANS, and 5GT. The expression levels of these structural genes in purple tubers were higher than those in white tubers, and F3′5′H gene was highly expressed in purple potato, while its expression significantly decreased in the red mutant30. It was found that the color of tuber skin of the StF3′5′H overexpression lines changed from red to purple, suggesting the StF3′5′H is a key gene for tuber pigmentation31. In contrast, the tuber skin and flesh changed from purple to red when the StF3′5′H gene was knocked out by using CRISPR/Cas9 technology. Overexpression of StANS gene contributed to the anthocyanin biosynthesis of potato tubers32. In addition, 2R-MYB transcription factors play essential roles in the regulation of anthocyanin synthesis in potato. So far, six St2R-MYB genes have been reported to be involved in regulating anthocyanin biosynthesis in potato. Among them, StMtf133, Stan234/StAN135,36/StMYB21037, StMYB11336,38, and StMYBA136,39,40/StMYB21037 significantly enhanced the anthocyanin accumulation in tobacco and potato, while StMYB4441 and StMYB342,43 had negative regulatory functions. However, the molecular mechanisms for anthocyanin biosynthesis and whether there are more potential 2R-MYB genes regulating anthocyanin metabolic pathway need to be further studied.
In this study, a total of 294 StMYB genes were identified from the latest version of DM potato genome data (DM, v6.1). The syntenic analysis, phylogenetic tree, conserved motifs, gene structures, and cis-elements of the StMYB genes were systematically analyzed. Meanwhile, a total of twelve candidate St2R-MYB genes were screened out according to the phylogenetic relationships between all St2R-MYB genes in potato and the anthocyanin-related 2R-MYB genes in other seven species. Subsequently, regulatory functions in anthocyanin synthesis for the candidate St2R-MYB genes were validated by transient transformation assays in tobacco. These results laid a foundation for further research on biological functions and molecular mechanisms of the St2R-MYB gene regulating anthocyanin biosynthesis in potato, and provided theoretical basis and new gene resources for the breeding of colorful potato varieties.
Materials and methods
Plant materials and growth conditions
The doubled monoploid potato clone DM1-3 516 R44 (DM, a homozygous line, 2n = 2x = 24) with whole genome sequencing information was used in this study. The DM seedlings preserved in our laboratory were cultured in test tube containing MS solid medium and grown in the light incubator. The growth condition was set to 16 h/8 h and 22 °C/18 °C for day/night, and light intensity was 150 µmol/m2/s (white fluorescent lamps). The plant leaves were used for RNA extraction, cDNA reverse transcription, and cloning of candidate St2R-MYB genes.
Nicotiana benthamiana and Nicotiana tabacum seeds preserved in our laboratory were sown in plastic bowl containing nutrient soil (vermiculite: peat soil = 1:1) and placed in an artificial climate chamber. The growth conditions were set to 16 h/8 h and 24 °C/18 °C for day/night, and light intensities were 200 µmol/m2/s (white fluorescent lamps). The seedlings were transplanted into small square pot after emerging 2–3 cotyledon, and cultured for 4–5 weeks (Nicotiana benthamiana) and 6–8 weeks (Nicotiana tabacum) for subcellular localization and transient over-expression assays, respectively.
Identification and classification of StMYB genes
The corresponding protein sequences of MYB gene family of A. thaliana were obtained from TAIR (https://www.arabidopsis.org/). The potato genomic data file was retrieved from the latest PGSC database (https://spuddb.uga.edu/dm_v6_1_download.shtml). Firstly, the potential MYB genes in the potato were identified using a local BLASTP search, the MYB protein sequences of Arabidopsis were used as queries with a threshold E-value < 1e−544. Meanwhile, the Hidden Markov Model (HMM) sequence files of the Myb-DNA-binding signature domain (PF00249) were downloaded from the Pfam (https://pfam.janelia.org/), and then used to search all MYB-containing sequences against the potato genome by HMMER software with E-value < 1e−545. The longest transcript of each candidate gene was applied for further analyses. In order to verify whether these candidate StMYB genes have MYB domain and analyze the number of MYB domain, the candidate StMYB genes were further validated using SMART (https://smart.emblheidelberg.de/), NCBI conserved domains (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi), and InterProScan (https://www.ebi.ac.uk/interpro/), and the sequences without MYB domain were removed.
Chromosome localization of StMYB genes
The position information of 294 StMYB genes on the 12 potato chromosomes were analyzed based on the general feature format (GFF) annotation file. The StMYB genes were named in chromosome order (Chr1 to Chr12) and in short-arm to long-arm direction. The location map of the StMYB genes was drawn using TBtools46. Meanwhile, the tandem duplication events of StMYB genes were identified.
Analysis of physicochemical properties and prediction of subcellular localization
The physicochemical parameters of StMYB proteins, including molecular weight (MW), theoretical isoelectric point (PIs), instability index (II), grand average of hydropathicity (GRAVY), and aliphatic index, were analyzed using the ExPASy (https://www.expasy.ch/tools/protparam.html) online website. In addition, the subcellular localization of StMYB proteins was predicted by using the WoLF PSORT (https://wolfpsort.hgc.jp/) online website.
Gene duplication and collinear analysis
The genome data of tobacco, eggplant, pepper, and tomato were downloaded from the Sol Genomics Network (SGN, https://solgenomics.net/). The grape genome was download from grape database (https://data.jgi.doe.gov/refine-download/phytozome?organism=Vvinifera). The collinearity of intraspecific and interspecific MYB genes was illustrated by the Multiple Collinearity Scan toolkit (MCScanX), with parameters set to: gap_penalty “− 1”, E-value “1e−10”47. The final syntenic maps were visualized using the TBtools software. To further evaluate duplication events, the non-synonymous rate (Ka), synonymous rate (Ks), and evolutionary constraint (Ka/Ks) between the duplicated pairs of StMYBs were computed using KaKs_Calculator2.048.
Gene structure, motif composition, and promoter cis-elements analysis
The protein sequences of St2R-MYB genes were extracted from potato DM v6.1 database, and the phylogenetic tree was constructed using the neighbor-joining (NJ) method with default parameter (pairwise deletion, 1000 bootstrap) by MEGA11.0 software49. The intron/exon structure was analyzed online using the Gene Structure Display Server (GSDS: https://gsds.gao-lab.org/). The conserved motifs of St2R-MYB proteins were determined using the MEME Suite (https://meme-suite.org/meme/tools/meme) with the parameters of highest motif number of 20, and motif occurrences distribution of zero or one per sequence50. The promoter sequences of 2000-bp regions upstream of the transcription start site (ATG) of St2R-MYB genes were extracted using the genomic sequences, and were used to analyze the cis-acting regulatory elements using the online PlantCARE database (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/). The results of gene structures, conserved motifs, and promoter elements were displayed by the TBtools software46.
Identification of candidate St2R-MYB genes associated with anthocyanin synthesis
A total of 38 2R-MYB genes involved in anthocyanin synthesis were searched from seven species, including tomato, eggplant, pepper, tobacco, grape, Arabidopsis thaliana and petunia51,52,53,54,55. The protein sequences of 2R-MYB genes in different species were downloaded from the National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/) (Table S1). The phylogenetic tree of 2R-MYB proteins clustered from potato and other species were constructed to screen the candidate St2R-MYB genes, which may be involved in regulating anthocyanin biosynthesis, using MEGA 11 by the neighbor-joining (NJ) algorithm method with 1000 replicates of bootstrap value49. Additionally, the phylogenetic tree was visualized and beautified by ChiPlot software (https://www.chiplot.online/).
Gene cloning and sequence analysis
The total RNA of DM leaves was extracted by OmniPlant RNA Kit (DNase I) (CWBIO, China) in terms of the instructions. The cDNA was synthesized by the reverse transcription of 1 µg RNA using PrimeScript™ first strand cDNA synthesis kit (TaKaRa, China). Full-length CDSs of the candidate St2R-MYB genes were retrieved from the potato genome database. PCR was carried out with the specific primers designed by primer 5.0 (Table S2). The PCR products were cloned into pMD19-T vector (Takara, China) and were sequenced by the Sangon Biotech (Shanghai, China) to gain the coding sequences. Multiple sequence alignment was performed by DNAMAN software.
Construct development and subcellular localization
The CDS of the candidate genes without termination codon was inserted into the pRI101 vector containing the green fluorescent protein (GFP) reporter gene and driven by the CaMV35S promoter to generate the recombinant construct 35S::St2R-MYB-GFP. The empty vector (35S::GFP) was used as the control. The empty vector and fusion constructs were introduced into Agrobacterium tumefaciens strain GV3101, respectively, and then transformed into 5-week-old tobacco (N. benthamiana) leaves by agroinfiltration56. The cells of tobacco leaves after 2–3 days infiltration were observed and photographed using a laser scanning confocal microscope (Leica TCS SP5-II, Wetzlar, Germany). The excitation wavelength with 488 nm was used for GFP observation.
Transient transformation assays in N. tabacum leaves
Transient transformation assays were performed in tobacco with reference to the previous method57. The full-length CDS of the candidate genes was fused into pRI101 vector and transferred into Agrobacterium tumefaciens strain GV3101. The transformed Agrobacterium was cultured at 28 °C until the optical density (OD) 0.6–0.8, and the cells were centrifuged. The suspension buffer (10 mmol/L MES, 10 mmol/L MgCl2, 0.15 mmol/L acetosyringone, pH 5.8) was used to dilute the cells and then cultured at room temperature for 4 h for further infiltration. For validation of the negative regulators, the Agrobacterium cultures containing the candidate gene were transformed into the tobacco (N. tabacum) leaves separately. For validation of the positive regulators, two Agrobacterium cultures containing the candidate gene and the StMYB_243 gene were thoroughly mixed in a volume of 1:1 and were then co-transformed into the tobacco leaves. The phenotype and anthocyanin content of tobacco leaves was observed and measured after 7 days injection, respectively. Three biological replicates were performed, and three replicates per trial.
Determination of anthocyanins content
The measurement of total anthocyanin content in tobacco leaves was described as previous study58. In brief, 0.1 g of sample were taken from the infected tobacco leaves, and 3 mL of hydrochloric acid ethanol solution was added into test tube. The anthocyanin was extracted overnight at room temperature. Add another 3 mL of extract solution to 6 mL, stand for 1 h. The optical density of the extracts at 530 nm, 620 nm, and 650 nm was determined by microplate reader (BioTek Epoch, Vermont, America). The content of anthocyanin was calculated using the following formula:
$${text{OD}}_{{uplambda }} = left( {{text{OD}_{530}} – {text{OD}_{620}}} right) – left( {{text{0.1* (OD}_{650}} – {text{OD}_{620}}} right)$$
(1)
$${text{Anthocyanidin}};{text{content}};left( {{text{nmol}}/{text{g}}} right) = {text{OD}}_{lambda} *{text{V}}*{10}^{6}/varepsilon /{text{m}}$$
(2)
ODλ represents optical density, ɛ represents the molar extinction coefficient of anthocyanidin (4.62 × 104), V represents the total volume of the extraction solution (mL), m represents the weight of the sample (g), and 106 is calculated as a multiple of nmol. Three biological replicates and three technical replicates were carried out for each sample. Statistical significance was analyzed by the Student’s t-test or by one-way ANOVA followed by the Duncan’s post-hoc test using SPSS software.
Results
Identification and classification of MYB family genes in potato
To comprehensively identify MYB family genes in DM potato genome with the latest version of whole genome sequencing data (DM v6.1, The PGSC 2020), the MYB protein sequences of Arabidopsis and the HMM profiles of the Myb-DNA-binding domain (PF00249) were provided simultaneously as queries to search against the potato genome, 349 and 288 candidate StMYB genes were screened out, respectively. After removing redundant genes, a total of 364 StMYB genes were obtained by the two methods. Subsequently, the SMART, NCBI conserved domains, and InterProScan online database were used to validate whether the obtained genes belong to the MYB family. As a result, a total of 294 StMYB genes were identified from the latest potato genome data (DM v6.1).
The 294 StMYB genes were further classified into three different gene subfamilies according to the number of MYB domain repeats, which consists of 149 St1R-MYBs (50.68%), 142 St2R-MYBs (48.30%), and 3 St3R-MYBs (1.02%) (Table S3).
Chromosome distribution of the StMYB genes
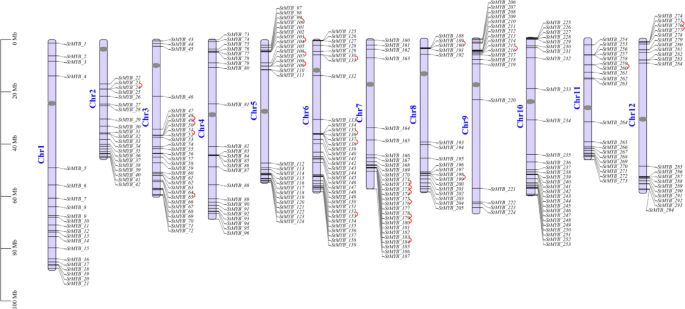
All of the 294 newly identified StMYB genes were mapped to the 12 chromosomes of potato according to their annotation information from the latest potato genome data (DM v6.1) (Fig. 1, Table S3). We named them StMYB_1 to StMYB_294 in the order based on their physical positions from chromosome 1 (Chr1) to chromosome 12 (Chr12) and from short arm (top) to long arm (bottom) of each chromosome. As shown in Fig. 1, the 294 StMYB genes were unevenly distributed on the 12 chromosomes, and the number of StMYB genes on each chromosome ranged from 18 to 35. Chromosome 6 had the largest number (35) of StMYB genes, followed by chromosomes 3 and 10, which contained 30 and 29 StMYBs, respectively. Both chromosomes 5 and 7 contained 28 StMYBs. In contrast, chromosomes 8 and 9 only had 18 and 19 StMYBs, respectively. Interestingly, we found that the majority of StMYB genes were mapped to opposite ends of the chromosomes, and were distant from the centromeres. However, the StMYB_132 gene was strikingly close to the centromere of chromosome 6 (Chr6), indicating that it may have some specific functions.

Distribution of 294 StMYB genes on chromosomes of DM potato. The chromosome name is given to the left of each chromosome. Gray oval on the chromosome bar represents centromere location for each chromosome, information of centromeres was collected according to the PGSC 2020. The tandem duplication pair of two StMYB genes was marked by red line.
Physicochemical properties and subcellular localization of MYB proteins in potato
To understand the characteristics of the 294 MYB proteins in potato, we systematically analyzed their physicochemical properties as shown in Table S4. The assessed number of amino acids encoded by StMYBs ranged greatly from 77 (StMYB_225) to 1677 (StMYB_82). The molecular weights (MWs) of these StMYB proteins ranged from 8.629 kDa (StMYB_100) to 183.739 kDa (StMYB_82), with an average of 38.075 kDa per protein. Their instability indexs (II) ranged from 1.02 (StMYB_282) to 80.46 (StMYB_20), with only eighteen implicated stable proteins (II < 40). The aliphatic indexs of these StMYB proteins were between 44.24 (StMYB_131) and 97.15 (StMYB_198), with an average of 69.3. And their theoretical isoelectric points (PIs) ranged from 4.35 (StMYB_121) to 10.27 (StMYB_225), with 58.5% of them lower than 7. The hydrophilicities of these StMYB proteins ranged from − 8.52 (StMYB_170) to 0.936 (StMYB_28), and only three StMYB proteins (StMYB_28, StMYB_36, and StMYB_244) whose hydrophilicities greater than zero, suggesting that most of these StMYB proteins are hydrophilic.
Prediction of subcellular localization of the 294 StMYB proteins showed that the majority (275, 93.5%) of them were mainly located in the nucleus, only a few of them in chloroplasts (9, 3.1%), cytoplasms (4, 1.4%), mitochondria (4, 1.4%), and endoplasmic reticulums (ER) (2, 0.7%; StMYB_22 and StMYB_209), respectively (Table S4). To validate the prediction results of subcellular localization, three representative proteins StMYB_99, StMYB_150, and StMYB_243 predicted mainly in the nucleus were selected for validation. Plasmid DNAs of three fusion constructs (35S::St2R-MYB-GFP) and the control (35S::GFP) were infiltrated into the tobacco (N. benthamiana) leaves, respectively. The results exhibited that the GFP fluorescence signals generated from the three constructs were all only distributed in the nucleus, while the control had an obvious fluorescence signals throughout the whole cells (Fig. 2), which were consistent with the prediction results mentioned above.

The subcellular localization of StMYB_99, StMYB_150, and StMYB_243 in tobacco epidermal cells. The panels from left to right correspond to fluorescence (GFP), bright field (Bright) and merged fluorescence (Merged) images, respectively. Scale bars: 20 µm (red) and 50 µm (white).
Evolutionary analysis of MYB genes
In order to comprehend the evolutionary relationship of StMYB genes in potato, homology analysis was performed between adjacent StMYB genes on each chromosome and between StMYB genes on two different chromosomes. Two adjacent StMYB genes sharing more than 70% similarity were considered as one pair of tandem-duplicated genes on each chromosome. A total of 28 tandem-duplicated gene pairs consisting of 51 StMYB genes were identified on 10 chromosomes with the exception of chromosomes 1 and 4 (Fig. 1, Table S5). Among them, seven tandem-duplicated gene pairs with 11 StMYB genes were distributed on chromosome 7 (Fig. 1); and there were 4 tandem-duplicated gene pairs on chromosomes 3, 5, and 6; two tandem-duplicated gene pairs on chromosomes 8, 10, and 12; and only one tandem-duplicated gene pair on chromosomes 2, 9, and 11 (Fig. 1). In addition to tandem-duplicated gene pairs, 72 segmental-duplicated gene pairs consisting of 105 StMYB genes were also identified on 12 chromosomes (Fig. 3, Table S6). These results indicated that StMYB genes undergone gene duplication and expansion events during the evolutionary process of potato.

Interchromosomal relationships of StMYB genes from DM potato. The 72 putative segmental duplication pairs of the StMYB genes were linked by the red lines.
To further explore the evolutionary relationship between StMYB genes in potato and MYB genes in six other plant species, including A. thaliana (Arabidopsis), V. vinifera (grape), C. annuum (pepper), N. tabacum (tobacco), S. lycopersicum (tomato), and S. melongena (eggplant), synteny analysis was performed. The results demonstrated that syntenic relationship between 225 of the StMYB genes with MYB genes from tomato, followed by 209 StMYB genes with eggplant, 174 StMYB genes with pepper, 165 StMYB genes with tobacco, 140 and 126 StMYB genes with MYB genes in grape and Arabidopsis, respectively (Fig. 4, Table S7). Additionally, a total of 199 StMYB genes were identified to form collinear pairs with MYB genes from eggplant and tomato, 136 collinear pairs of StMYB genes from pepper and tobacco, and 109 collinear pairs of StMYB genes from grape and Arabidopsis (Fig. 4, Table S7). Notably, the number of collinear pairs of StMYB genes localized at highly conserved syntenic regions from eggplant and tomato, both belonging to the Solanum family, were much more than those from pepper and tobacco representing Solanaceae family, and from grape and Arabidopsis, both belonging to the dicot species. These results were consistent with the evolutionary relationship between potato and the other six plant species. In addition, a total of 72 StMYB genes were found to form collinear pairs with MYB genes from all of the other six plant species, indicating that these genes may have been presented at the initial differentiation stage of dicot species. Ka/Ks analysis showed that the Ka/Ks values of 111 MYB gene pairs were less than 0.5 (Table S8), indicating that these genes undergone strong purify selection, and were relatively conservative in evolution process and gene function.

Synteny analyses of MYB genes between DM potato and six representative plant species. The synteny analysis was performed and visualized using the MCScanX and TBtools software, respectively. Gray lines in the background represented collinear blocks within DM potato and other representative plant species, while colored lines exhibited syntenic MYB gene pairs. Chromosomes of six representative plant species were showed by different colored rectangles.
Gene structure, motif composition, and promoter cis-acting element analysis of St2R-MYB genes
To further understand the evolutionary relationships among 142 St2R-MYB genes, a phylogenetic tree was built (Fig. 5A). In addition to StMYB_77, StMYB_82, and StMYB_109, all St2R-MYB genes were classified into 20 subgroups (S1 to S20), with an average of 6.95 genes per subgroup (Fig. 5A). S16 was the largest subgroup comprising 17 St2R-MYB genes. Subgroups 1, 2, 10, 11, and 18 containing 10–13 St2R-MYB genes; Subgroups 3, 6, 8, 9, 14, 15, 17, and 20 containing 5–7 St2R-MYB genes; Subgroups 4, 5, 7, 12, 13, and 19 containing 2–4 St2R-MYB genes.

Analyses of phylogenetic relationships, gene structures, conserved motifs, and cis-acting elements of St2R-MYB genes. (A) Phylogenetic tree of the 142 St2R-MYB proteins. (B) Exon–intron structures of the St2R-MYB genes. Yellow boxes indicated 5′-UTR and 3′-UTR regions, and red boxes and gray lines represented exons and introns, respectively. (C) Motif compositions of the St2R-MYB proteins. Twenty different types of motifs were exhibited in different colored boxes. The scale at the bottom can be used for estimation of the protein lengths. (D) Distribution of cis-acting elements of St2R-MYB genes promoters. Twenty-eight different types of responsiveness elements were displayed in different colored blocks.
The layout of the exon–intron structures showed that 137 St2R-MYB genes contained 1–10 introns with the exception of four genes StMYB_93, StMYB_36, StMYB_92, and StMYB_86 located in S18, and the StMYB_109 gene not located in any subgroup, which had no intron (Fig. 5B). Meanwhile, there were differences in gene structures among different subgroups. The gene structure of StMYB_77 with 11 exons and 10 introns was the most complicated, followed by StMYB_258 (5 introns) located in S20. Seven St2R-MYB genes located in five subgroups S9 (1), S13 (1), S16 (2), S17 (2), and S20 (1), respectively, had 4 exons and 3 introns. Remarkably, most of the St2R-MYB genes (105, 73.94%) contained similar gene structures with 3 exons and 2 introns, followed by 23 (16.20%) St2R-MYB genes having 2 exons and 1 intron (Fig. 5B).
Diversity of conserved motifs may reflect the functional divergence within gene families. Therefore, we analyzed the conserved motifs of the St2R-MYB genes (Fig. 5C). We identified 20 conserved motifs (named motif 1–20) from 142 St2R-MYB proteins and extracted their corresponding sequences logos via online MEME program. As shown in Fig. 5C, the motifs 1, 2, 3, and 4 were generally appeared in most of the St2R-MYB genes, which indicated that these four motifs were relatively conservative. Intriguingly, we found that the motif distribution and composition were similar among members within the same subgroup, such as motif 9 was specific to S16, and motifs 7 and 8 were special to S20. Strikingly, the number and type of motifs were similar in the same subgroup, demonstrating that these genes may have parallel functions.
Promoter cis-elements play essential roles in the activation of gene expression. Thus, we further analyzed the conserved cis-acting elements of the 2000-bp upstream regions of the 142 St2R-MYB genes for better understanding their biological functions. In this study, a total of 28 cis-acting elements were identified in the promoter regions of the St2R-MYB genes, which could be divided into four categories, including light-responsive elements (G-Box and Box 4), environmental stress elements (Low-temperature responsiveness element (LTR), Drought-responsive element (DRE) core, MBS), phytohormones elements (TCA-element, Estrogen responsive element (ERE), Abscisci acid (ABA), Absicsic acid responsive element (ABRE), and growth and development elements (CAT-box, TATA-box) (Fig. 5D, Table S9). Except StMYB_178 and StMYB_275, the 140 St2R-MYB genes contained 3–12 cis-acting elements. Among them, 140 genes contained light-responsive elements, 111 genes had abscisic acid (ABA) responsiveness elements, 91 genes contained Methyl jasmonate (MeJA)-responsive elements, 59 genes contained salicylic acid (SA) responsiveness elements, 55 genes contained gibberellin-responsive elements, 55 genes contained auxin-responsive elements, 52 genes contained drought-responsive elements (DREs), 39 genes contained low-temperature responsiveness elements (LTRs), and 8 genes contained mechanical damage responsiveness elements (Fig. 5D, Table S9). These results revealed that St2R-MYB genes might play important regulatory roles in the processes of hormone regulations, stress and light responses.
Screening of candidate St2R-MYB genes associated with anthocyanin biosynthesis
A mass of researches have shown that 2R-MYB genes played crucial regulatory roles in the anthocyanin biosynthesis. Meanwhile, the analysis of phylogenetic relationships based on protein sequences could provide valuable evidence to functionality. Thus, to comprehensively identify candidate St2R-MYB genes related to anthocyanin biosynthesis in potato, we constructed a phylogenetic tree with 142 St2R-MYB proteins from potato and 38 St2R-MYB proteins from other seven species comprising A. thaliana (Arabidopsis), V. vinifera (grape), P. hybrida (petunia), C. annuum (pepper), N. tabacum (tobacco), S. lycopersicum (tomato), and S. melongena (eggplant) (Fig. 6). Strikingly, the phylogenetic tree exhibited that six genes, including StMYB_1, StMYB_239, StMYB_240, StMYB_241, StMYB_242, and StMYB_243, were clustered simultaneously together with multiple 2R-MYB genes reported to promote anthocyanin biosynthesis in other species, while StMYB_99 was close to four 2R-MYB genes that have been reported to inhibit anthocyanin biosynthesis in grape and petunia (Fig. 6). In addition, genes StMYB_15 and StMYB_52 were closest to genes SmMYB35 and NtMYB4a known as positive regulators of anthocyanin biosynthesis in eggplant and tobacco, respectively, whereas three genes, StMYB_66, StMYB_150, and StMYB_248, were clustered together with CaMYB306, SmMYB86, and AtMYB60 verified as negative regulators of anthocyanin biosynthesis, respectively (Fig. 6). Finally, a total of 12 St2R-MYB genes, including 8 possible positive regulators (StMYB_1, StMYB_15, StMYB_52, StMYB_239, StMYB_240, StMYB_241, StMYB_242, and StMYB_243) and 4 possible negative regulators (StMYB_66, StMYB_99, StMYB_150, and StMYB_248), were screened as candidate regulatory genes for regulating anthocyanin biosynthesis.

Unrooted phylogenetic treebetween St2R-MYB genes and 2R-MYB genes from other species. The 2R-MYB genes highlighted in orange, pink, red, dark blue, dark green, yellow, blue, light green, and light yellow were from DM potato, tomato, eggplant, pepper, tobacco, grape, Arabidopsis, and petunia, respectively. “ + ” represents anthocyanin related promoter genes, while “-” represents anthocyanin related inhibitor genes. The phylogenetic tree was constructed using MEGA 11 by the neighbor-joining (NJ) algorithm method with 1,000 replicates of bootstrap value, and visualized using the ChiPlot software.
Heterologous overexpression of the candidate St2R-MYB genes in N. tabacum leaves
To further validate regulatory functions of the 12 candidate St2R-MYB genes screened above in anthocyanin biosynthesis, the transient expression assay of tobacco leaves was carried out. For validation of 8 possible positive regulators of anthocyanin biosynthesis, the Agrobacterium tumefaciens cultures containing the overexpression construct with the target gene were injected separately into tobacco leaves, the phenotypes of leaf colors were observed and the anthocyanin contents were measured. As shown in Fig. 7, the tobacco leaves overexpressed by the genes StMYB_240, StMYB_241, StMYB_242, and StMYB_243, respectively, appeared purplish red, and in which the anthocyanin contents were also significantly increased compared with the negative control (EV, empty vector) (P < 0.01) (Fig. 7A–D), demonstrating that genes StMYB_240, StMYB_241, StMYB_242, and StMYB_243 could significantly promote anthocyanin biosynthesis in tobacco. However, no evident color changes were observed in tobacco leaves overexpressed by the genes StMYB_15, StMYB_52, and StMYB_239, respectively, and in which the differences of anthocyanin contents were also not significant (P < 0.05, data not shown) when compared to their negative controls, indicating that these three genes may not directly participated in regulating anthocyanin biosynthesis in tobacco leaves. The regulatory function of StMYB_1 gene in tobacco leaves is unknown due to the failure of PCR amplification for gene cloning which may be resulted from the complex genomic locus.

Transient assays of the candidate St2R-MYB genes. Phenotype and anthocyanin content in tobacco leaves collected 7 days after infiltration with Agrobacterium harboring StMYB_240, StMYB_241, StMYB_242, StMYB_243, StMYB_66, StMYB_99, StMYB_150, and StMYB_248, an empty vector (EV) as a control. The error bars means the standard deviation of three biological replicates. Statistical significance was analyzed by the Student’s t-test: ** P < 0.01 (A–D) or by one-way ANOVA followed by the Duncan’s post-hoc test using SPSS software, different lowercase letters indicate significant differences (P < 0.01) (E–H). Degrees of freedom and F-values were showed in Table S10.
For validation of 4 possible negative regulators of anthocyanin biosynthesis, Agrobacterium tumefaciens cultures containing the overexpression constructs with the target gene and the StMYB_243 gene, respectively, were thoroughly mixed in a volume of 1:1 and were then injected into tobacco leaves to observe leaf colors and measure anthocyanin contents. Tobacco leaves injected by Agrobacterium tumefaciens cultures containing the empty vector (EV) or the overexpression construct with the target gene were considered to be negative controls, whereas tobacco leaves injected by Agrobacterium tumefaciens cultures containing both of the empty vector (EV) and the overexpression construct with the StMYB_243 gene were used as positive controls. As we expected, the colors of tobacco leaves simultaneously expressed both of the target gene and the StMYB_243 gene were lighter than the positive controls, but darker than the two negative controls (Fig. 7E–H). Consistently, the anthocyanin contents were also significantly decreased (P < 0.01) (Fig. 7E–H, Table S10). These results demonstrated that these four genes could significantly inhibit anthocyanin biosynthesis in tobacco leaves.
Discussion
The MYB transcription factors played important roles in plant growth, development, metabolism, biotic and abiotic stresses, hormone signal transduction, and protecting against pathogens. With the rapid development of whole genome sequencing (WGS), the genome-wide identifications of MYB TF superfamily have been carried out in numerous species, such as Arabidopsis, rice, maize, and tomato9,59,60. In previous studies, a total of 158, 233, and 217 MYB genes were identified in DM potato genome with the earliest version of WGS data (DM v4.03, The PGSC 2011)11,12,13. However, the MYB family genes have not yet been comprehensively identified and systematically analyzed in DM potato genome with the latest version of WGS data (DM v6.1, The PGSC 2020), and the data contained greatly comprehensive and accurate genome information. A total of 294 StMYB genes, comprising of 149 St1R-MYBs, 142 St2R-MYBs, and three St3R-MYBs, were identified in this study, and the number of StMYB genes was obviously more than those identified from previous studies (Fig. 1, Table S3). In plants, 2R-MYB genes played central roles in controlling plant-specific processes such as plant development, secondary metabolism, and biotic and abiotic stress responses6,29,61. Remarkably, 20 novel St2R-MYB genes were firstly discovered in this study (Table S11), which have a potential positive regulatory effect on potato anthocyanin biosynthesis, which was definitely benefited from the more comprehensive and accurate WGS data assembled by the improved long-read chromosome-scale genome assembly technology62. Sequence analysis of the 142 St2R-MYB genes exhibited that most of them shared similar motif compositions and exon–intron structures in the same subgroup (Fig. 5), suggesting these genes may have similar functions. Furthermore, 28 pairs of tandem-duplicated genes and 72 pairs of segmental-duplicated genes were found among the 294 StMYB genes (Figs. 1 and 3), which was more than that identified from the previous study containing 8 and 31 pairs of the genes, respectively12. A large number of tandem duplication and segmental duplication events were the main cause of the expansion of StMYB genes in potato genome. Additionally, a total of 72 StMYB genes were found to form collinear pairs with MYB genes from all of the other six plant species (Fig. 4, Table S7), which indicated that gene duplication events might lead to the expansion of MYB family genes in different plant species and that these genes might have been presented at the initial differentiation stage of dicot species.
Anthocyanin is widely distributed in plants, and leads to colored roots, tubers, stems, leaves, and flowers. The anthocyanin biosynthesis is mainly regulated by the expression patterns of structural genes and regulatory genes63. MYB TFs, one of the largest gene families, played important roles in anthocyanin biosynthesis in potato64. In order to comprehensively investigate St2R-MYB genes associated with anthocyanin biosynthesis, 38 St2R-MYB genes previously reported to involve with anthocyanin biosynthesis in seven plant species were merged with 142 St2R-MYB genes identified in this study to construct a phylogenetic tree. Finally, a total of 12 candidate St2R-MYB genes were screened out for regulating anthocyanin biosynthesis (Fig. 6). The transient transformation assays validated that the candidate genes StMYB_240, StMYB_241, StMYB_242, and StMYB_243 could significantly promoted anthocyanin biosynthesis in tobacco leaves (Fig. 6). Interestingly, the sequence analysis showed that StMYB_240, StMYB_241, StMYB_242, and StMYB_243 homologous to the previously reported StMYB genes (Table S12), and the sequence identities were 89% to 100% (Table S12). Strikingly, four novel genes StMYB_66, StMYB_99 (homologous to previously reported StMYB3), StMYB_150, and StMYB248 were validated as negative regulators to repress anthocyanin biosynthesis in tobacco leaves (Fig. 6), and they were clustered together with CaMYB30665, PhMYB2728, SmMYB8666, and AtMYB6024, respectively, which have been reported as repressors.
The promoter cis-acting elements analysis of these 8 potentially St2R-MYB regulatory genes exhibited that they all had multiple light-responsive elements and anaerobic induction elements, and also contained ABA-responsive elements, demonstrating that these genes may be involved in inhibiting plant growth and promoting tuber formation of potato. All promoters of StMYB_66, StMYB_240, and StMYB_248 contained gibberellin-responsive elements, indicating that these three genes may be participated in plant growth and development, and have potential effects on improving yield and quality. As we understand, the potato perceives the short-day photoperiod through the above-ground parts, then generates tuber induction signals, and transmits the signals to the underground stolons, which receive the signals for tuber formation. During the induction process, gibberellin plays a role in inhibiting the conversion of stolons to tubers67. Both StMYB_241 and StMYB_242 gene promoters contained low-temperature responsiveness (LTRs) elements, suggesting potential roles in enhancing the activities of pigment synthetases under low temperature conditions, so as to promote the anthocyanins biosynthesis. All promoters of genes StMYB_66, StMYB_150, StMYB_242, and StMYB_243 contained MeJA-responsive elements, which may be involved in the regulation of secondary metabolic pathways including anthocyanin synthesis. However, these need further study.
Conclusions
In this study, a total of 294 StMYB genes were identified and their chromosomal distribution, protein properties, and duplication event were evaluated. Remarkably, 20 novel St2R-MYB genes were firstly discovered at present study. The phylogenetic tree, conserved motifs, gene structures, and cis-elements of the St2R-MYB genes were also analyzed to get a better understanding the characterizations and roles of the St2R-MYB genes in potato. An unrooted phylogenetic tree between potato and other seven species were constructed to screeen the candidate St2R-MYB genes for regulating anthocyanin biosynthesis. As we expected, heterologous expression in tobacco validated that the candidate genes StMYB_240, StMYB_241, StMYB_242, and StMYB_243 functioned as positive regulators for anthocyanin biosynthesis, while StMYB_66, StMYB_99, StMYB_150, and StMYB_248 acted as negative regulators. Our study laid the foundation for further research on biological functions and molecular mechanisms of the candidate St2R-MYB genes regulating anthocyanin biosynthesis in potato, and provided theoretical basis and new gene resources for the breeding of colorful potato varieties.
Data availability
All data generated or analysed during this study are included in this published article.
References
-
Han, K. H. et al. Anthocyanin-rich purple potato flake extract has antioxidant capacity and improves antioxidant potential in rats. Br. J. Nutr. 96(6), 1125–1133. https://doi.org/10.1017/bjn20061928 (2006).
-
Han, K. H. et al. Purple potato flake reduces serum lipid profile in rats fed a cholesterol-rich diet. J. Funct. Foods 5(2), 974–980. https://doi.org/10.1016/j.jff.2013.01.007 (2013).
-
Madiwale, G. P., Reddivari, L., Holm, D. G. & Vanamala, J. Storage elevates phenolic content and antioxidant activity but suppresses antiproliferative and pro-apoptotic properties of colored-flesh potatoes against human colon cancer cell lines. J. Agric. Food Chem. 59(15), 8155–8166. https://doi.org/10.1021/jf201073g (2011).
-
Ramsay, N. A. & Glover, B. J. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 10(2), 63–70. https://doi.org/10.1016/j.tplants.2004.12.011 (2005).
-
Jia, L., Clegg, M. T. & Jiang, T. Evolutionary dynamics of the DNA-binding domains in putative R2R3-MYB genes identified from rice subspecies indica and japonica genomes. Plant Physiol. 134(2), 575–585. https://doi.org/10.1104/pp.103.027201 (2004).
-
Dubos, C. et al. MYB transcription factors in Arabidopsis. Trends Plant Sci. 15(10), 573–581. https://doi.org/10.1016/j.tplants.2010.06.005 (2010).
-
Klempnauer, K. H., Gonda, T. J. & Bishop, J. M. Nucleotide sequence of the retroviral leukemia gene v-myb and its cellular progenitor c-myb: The architecture of a transduced oncogene. Cell 31(2 Pt 1), 453–463. https://doi.org/10.1016/0092-8674(82)90138-6 (1982).
-
Stracke, R., Werber, M. & Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 4(5), 447–456. https://doi.org/10.1016/s1369-5266(00)00199-0 (2001).
-
Katiyar, A. et al. Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis. BMC Genomics 13, 544. https://doi.org/10.1186/1471-2164-13-544 (2012).
-
Wang, N. H. et al. A comparative genome-wide analysis of the R2R3-MYB gene family among four gossypium species and their sequence variation and association with fiber quality traits in an interspecific G. hirsutum × G. barbadense population. Front. Genet. 10, 741. https://doi.org/10.3389/fgene.2019.00741 (2019).
-
Sun, W. J., Ma, Z. T., Chen, H. & Liu, M. Y. MYB gene family in potato (Solanum tuberosum L.): Genome-wide identification of hormone-responsive reveals their potential functions in growth and development. Int. J. Mol. Sci. 20(19), 4847. https://doi.org/10.3390/ijms20194847 (2019).
-
Li, X. X. et al. Systematic analysis of MYB family genes in potato and their multiple roles in development and stress responses. Biomolecules 9(8), 317. https://doi.org/10.3390/biom9080317 (2019).
-
Li, Y. M. et al. Genome-wide analysis of MYB gene family in potato provides insights into tissue-specific regulation of anthocyanin biosynthesis. Hortic. Plant J. 7(2), 129–141. https://doi.org/10.1016/j.hpj.2020.12.001 (2021).
-
Cone, K. C., Burr, F. A. & Burr, B. Molecular analysis of the maize anthocyanin regulatory locus C1. Proc. Natl. Acad. Sci. U. S. A. 83(24), 9631–9635. https://doi.org/10.1073/pnas.83.24.9631 (1986).
-
Paz-Ares, J., Ghosal, D., Wienand, U., Peterson, P. A. & Saedler, H. The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators. EMBO J. 6(12), 3553–3558. https://doi.org/10.1002/j.1460-2075.1987.tb02684.x (1987).
-
Borevitz, J. O., Xia, Y. J., Blount, J., Dixon, R. A. & Lamb, C. Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant Cell 12(12), 2383–2393. https://doi.org/10.1105/tpc.12.12.2383 (2000).
-
Li, G., Serek, M. & Gehl, C. Physiological changes besides the enhancement of pigmentation in Petunia hybrida caused by overexpression of PhAN2, an R2R3-MYB transcription factor. Plant Cell Rep. 42(3), 609–627. https://doi.org/10.1007/s00299-023-02983-1 (2023).
-
Albert, N. W. et al. Members of an R2R3-MYB transcription factor family in Petunia are developmentally and environmentally regulated to control complex floral and vegetative pigmentation patterning. Plant J. 65(5), 771–784. https://doi.org/10.1111/j.1365-313X.2010.04465.x (2011).
-
Espley, R. V. et al. Red coloration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 49(3), 414–427. https://doi.org/10.1111/j.1365-313X.2006.02964.x (2007).
-
Menconi, J., Perata, P. & Gonzali, S. Novel R2R3 MYB transcription factors regulate anthocyanin synthesis in Aubergine tomato plants. BMC Plant Biol. 23(1), 148. https://doi.org/10.1186/s12870-023-04153-7 (2023).
-
Jian, W. et al. SlMYB75, an MYB-type transcription factor, promotes anthocyanin accumulation and enhances volatile aroma production in tomato fruits. Hortic. Res. 6, 22. https://doi.org/10.1038/s41438-018-0098-y (2019).
-
Jun, J. H., Liu, C. G., Xiao, X. R. & Dixon, R. A. The transcriptional repressor MYB2 regulates both spatial and temporal patterns of proanthocyandin and anthocyanin pigmentation in Medicago truncatula. Plant Cell 27(10), 2860–2879. https://doi.org/10.1105/tpc.15.00476 (2015).
-
Tamagnone, L. et al. The AmMYB308 and AmMYB330 transcription factors from antirrhinum regulate phenylpropanoid and lignin biosynthesis in transgenic tobacco. Plant Cell 10(2), 135–154. https://doi.org/10.1105/tpc.10.2.135 (1998).
-
Park, J. S. et al. Arabidopsis R2R3-MYB transcription factor AtMYB60 functions as a transcriptional repressor of anthocyanin biosynthesis in lettuce (Lactuca sativa). Plant Cell Rep. 27(6), 985–994. https://doi.org/10.1007/s00299-008-0521-1 (2008).
-
Aharoni, A. et al. The strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in transgenic tobacco. Plant J. 28(3), 319–332. https://doi.org/10.1046/j.1365-313X.2001.01154.x (2001).
-
Colanero, S., Perata, P. & Gonzali, S. The atroviolacea gene encodes an R3-MYB protein repressing anthocyanin synthesis in tomato plants. Front. Plant Sci. 9, 830. https://doi.org/10.3389/fpls.2018.00830 (2018).
-
Tominaga-Wada, R., Nukumizu, Y. & Wada, T. Tomato (Solanum lycopersicum) homologs of TRIPTYCHON (SlTRY) and GLABRA3 (SlGL3) are involved in anthocyanin accumulation. Plant Signal. Behav. 8(7), e24575. https://doi.org/10.4161/psb.24575 (2013).
-
Albert, N. W. et al. A conserved network of transcriptional activators and repressors regulates anthocyanin pigmentation in eudicots. Plant Cell 26(3), 962–980. https://doi.org/10.1105/tpc.113.122069 (2014).
-
Allan, A. C., Hellens, R. P. & Laing, W. A. MYB transcription factors that colour our fruit. Trends Plant Sci. 13(3), 99–102. https://doi.org/10.1016/j.tplants.2007.11.012 (2008).
-
Liu, F., Yang, Y. J., Gao, J. W., Ma, C. L. & Bi, Y. P. A comparative transcriptome analysis of a wild purple potato and its red mutant provides insight into the mechanism of anthocyanin transformation. PLoS ONE 13(1), e0191406. https://doi.org/10.1371/journal.pone.0191406 (2018).
-
Jung, C. S. et al. The potato P locus codes for flavonoid 3′,5′-hydroxylase. Theor. Appl. Genet. 110(2), 269–275. https://doi.org/10.1007/s00122-004-1829-z (2005).
-
Zhang, H. L. et al. Functional analysis of an anthocyanin synthase gene StANS in potato. Sci. Hortic. 272, 109569. https://doi.org/10.1016/j.scienta.2020.109569 (2020).
-
Rommens, C. M. et al. Engineered native pathways for high kaempferol and caffeoylquinate production in potato. Plant Biotechnol. J. 6(9), 870–886. https://doi.org/10.1111/j.1467-7652.2008.00362.x (2008).
-
Jung, C. S. et al. The potato developer (D) locus encodes an R2R3 MYB transcription factor that regulates expression of multiple anthocyanin structural genes in tuber skin. Theor. Appl. Genet. 120(1), 45–57. https://doi.org/10.1007/s00122-009-1158-3 (2009).
-
D’Amelia, V. et al. High AN1 variability and interaction with basic helix-loop-helix co-factors related to anthocyanin biosynthesis in potato leaves. Plant J. 80(3), 527–540. https://doi.org/10.1111/tpj.12653 (2014).
-
Liu, Y. H. et al. Functional diversification of the potato R2R3 MYB anthocyanin activators AN1, MYBA1, and MYB113 and their interaction with basic helix-loop-helix cofactors. J. Exp. Bot. 67(8), 2159–2176. https://doi.org/10.1093/jxb/erw014 (2016).
-
Du, H. et al. Two tandem R2R3 MYB transcription factor genes cooperatively regulate anthocyanin accumulation in potato tuber flesh. Plant Biotechnol. J. 23(5), 1521–1534. https://doi.org/10.1111/pbi.14602 (2025).
-
Zhang, Z. H. et al. StMYB113 promotes anthocyanin biosynthesis in potato (Solanum tuberosum L.) Désirée tubers. Potato Res. 67(1), 307–324. https://doi.org/10.1007/s11540-023-09639-3 (2024).
-
Liu, Y. H. et al. The MYB transcription factor StMYBA1 from potato requires light to activate anthocyanin biosynthesis in transgenic tobacco. J. Plant Biol. 60(1), 93–101. https://doi.org/10.1007/s12374-016-0199-9 (2017).
-
Zhao, X. J. et al. Transcriptome analysis provides StMYBA1 gene that regulates potato anthocyanin biosynthesis by activating structural genes. Front. Plant Sci. 14, 1087121. https://doi.org/10.3389/fpls.2023.1087121 (2023).
-
Liu, Y. H. et al. StMYB44 negatively regulates anthocyanin biosynthesis at high temperatures in tuber flesh of potato. J. Exp. Bot. 70(15), 3809–3824. https://doi.org/10.1093/jxb/erz194 (2019).
-
Liu, Y. H. et al. Integrative analysis of metabolome and transcriptome reveals a dynamic regulatory network of potato tuber pigmentation. iScience 26(2), 105903. https://doi.org/10.1016/j.isci.2022.105903 (2023).
-
Liu, Y. H. et al. Integrated transcriptomic and metabolomic analysis revealed altitude-related regulatory mechanisms on flavonoid accumulation in potato tubers. Food Res. Int. 170, 112997. https://doi.org/10.1016/j.foodres.2023.112997 (2023).
-
Camacho, C. et al. BLAST+: Architecture and applications. BMC Bioinform. 10, 421. https://doi.org/10.1186/1471-2105-10-421 (2009).
-
Mistry, J., Finn, R. D., Eddy, S. R., Bateman, A. & Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 41(12), e121. https://doi.org/10.1093/nar/gkt263 (2013).
-
Chen, C. J. et al. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 13(8), 1194–1202. https://doi.org/10.1016/j.molp.2020.06.009 (2020).
-
Wang, Y. P. et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 40(7), e49. https://doi.org/10.1093/nar/gkr1293 (2012).
-
Wang, D. P., Zhang, Y. B., Zhang, Z., Zhu, J. & Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinf. 8(1), 77–80. https://doi.org/10.1016/S1672-0229(10)60008-3 (2010).
-
Tamura, K., Stecher, G. & Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38(7), 3022–3027. https://doi.org/10.1093/molbev/msab120 (2021).
-
Bailey, T. L. et al. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 37(suppl_2), W202–W208. https://doi.org/10.1093/nar/gkp335 (2009).
-
Kang, S. I. et al. Expression of anthocyanin biosynthesis-related genes reflects the peel color in purple tomato. Hortic. Environ. Biotechnol. 59(3), 435–445. https://doi.org/10.1007/s13580-018-0046-7 (2018).
-
Zhang, Y. J. et al. Anthocyanin accumulation and molecular analysis of anthocyanin biosynthesis-associated genes in eggplant (Solanum melongena L.). J. Agric. Food Chem. 62(13), 2906–2912. https://doi.org/10.1021/jf404574c (2014).
-
Luo, Q. et al. Isolation and molecular characterization of NtMYB4a, a putative transcription activation factor involved in anthocyanin synthesis in tobacco. Gene 760, 144990. https://doi.org/10.1016/j.gene.2020.144990 (2020).
-
Li, N. et al. The heterologous expression of Arabidopsis PAP2 induces anthocyanin accumulation and inhibits plant growth in tomato. Funct. Integr. Genomics 18(3), 341–353. https://doi.org/10.1007/s10142-018-0590-3 (2018).
-
Fu, Z. Z. et al. Three paralogous R2R3-MYB genes contribute to delphinidin-related anthocyanins synthesis in Petunia hybrida. J. Plant Growth Regul. 40(4), 1687–1700. https://doi.org/10.1007/s00344-020-10224-y (2021).
-
Ma, Y. et al. Genome-wide characterization of Solanum tuberosum UGT gene family and functional analysis of StUGT178 in salt tolerance. BMC Genomics 25(1), 1206. https://doi.org/10.1186/s12864-024-11140-1 (2024).
-
Li, J. et al. Combined transcriptomic and proteomic analysis constructs a new model for light-induced anthocyanin biosynthesis in eggplant (Solanum melongena L.). Plant Cell Environ. 40(12), 3069–3087. https://doi.org/10.1111/pce.13074 (2017).
-
Zhang, C., Ma, Y., Zhao, X. Y. & Mu, J. Influence of copigmentation on stability of anthocyanins from purple potato peel in both liquid state and solid state. J. Agric. Food Chem. 57(20), 9503–9508. https://doi.org/10.1021/jf901550u (2009).
-
Du, H., Feng, B. R., Yang, S. S., Huang, Y. B. & Tang, Y. X. The R2R3-MYB transcription factor gene family in maize. PLoS ONE 7(6), e37463. https://doi.org/10.1371/journal.pone.0037463 (2012).
-
Zhao, P. P., Li, Q., Li, J., Wang, L. & Ren, Z. H. Genome-wide identification and characterization of R2R3MYB family in Solanum lycopersicum. Mol. Genet. Genomics 289(6), 1183–1207. https://doi.org/10.1007/s00438-014-0879-4 (2014).
-
Roy, S. Function of MYB domain transcription factors in abiotic stress and epigenetic control of stress response in plant genome. Plant Signal. Behav. 11(1), e1117723. https://doi.org/10.1080/15592324.2015.1117723 (2016).
-
Pham, G. M. et al. Construction of a chromosome-scale long-read reference genome assembly for potato. Gigascience 9(9), giaa100. https://doi.org/10.1093/gigascience/giaa100 (2020).
-
Tang, B. Y. et al. Anthocyanin accumulation and transcriptional regulation of anthocyanin biosynthesis in purple pepper. J. Agric. Food Chem. 68(43), 12152–12163. https://doi.org/10.1021/acs.jafc.0c02460 (2020).
-
Ye, G. J. et al. The MYB transcription factor LrAN2, from Lycium ruthenicum, led to enhanced accumulation of anthocyanins and modified profile of the total glycoalkaloids in potato. Plant Cell Tissue Organ Cult. 147(3), 519–528. https://doi.org/10.1007/s11240-021-02144-w (2021).
-
Ma, X., Yu, Y. N., Jia, J. H., Li, Q. H. & Gong, Z. H. The pepper MYB transcription factor CaMYB306 accelerates fruit coloration and negatively regulates cold resistance. Sci. Hortic. 295, 110892. https://doi.org/10.1016/j.scienta.2022.110892 (2022).
-
Li, L. Z., He, Y. J., Ge, H. Y., Liu, Y. & Chen, H. Y. Functional characterization of SmMYB86, a negative regulator of anthocyanin biosynthesis in eggplant (Solanum melongena L.). Plant Sci. 302, 110696. https://doi.org/10.1016/j.plantsci.2020.110696 (2021).
-
Martínez-García, J. F., García-Martínez, J. L., Bou, J. & Prat, S. The interaction of gibberellins and photoperiod in the control of potato tuberization. J. Plant Growth Regul. 20(4), 377–386. https://doi.org/10.1007/s003440010036 (2001).
Funding
This research was funded by the National Natural Science Foundation of China (32201853), the Anhui Provincial Natural Science Foundation (2308085MC85, 2108085QC122), the Natural Science Research Projects of Anhui Province University (KJ2021ZD0013, KJ2020A0098), the Scientific Research Fund for Candidates of Anhui Provincial Academic and Technical Leaders (2021H276), and the Anhui Province Vegetable Industry Technology System Fund to X.Z. (24532009).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hou, H., Wang, L., Wang, N. et al. Systematic analysis of potato MYB family genes and their potential functions in anthocyanin biosynthesis. Sci Rep 15, 28113 (2025). https://doi.org/10.1038/s41598-025-13284-w
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-13284-w