- Review
- Open access
- Published:
Biotechnology for Biofuels and Bioproducts volume 18, Article number: 64 (2025) Cite this article
Abstract
Isoprenoids constitute a large and various number of bio-compounds, with many profitable applications in pharmaceutical, nutraceutical, and industrial fields. The complexity of isoprenoid molecules leads to a challenging, expensive, and environmentally unfriendly chemical synthesis of these metabolites. In addition, the awareness and desire of many consumers for products generated by natural microbial processes has increased recently. Metabolic engineering tools and synthetic biology strategies have been used as a means for the enhancement and optimization of the natural isoprenoid biosynthetic pathways of wild strains. Microalgae as production organisms have been manipulated for the bioproduction of diverse isoprenoids. Particularly when cultivated in unsuitable conditions (such as wastewater, unbalanced nutritional sources, and distinct environmental conditions), microalgae can adjust their metabolic pathways and generate compounds with significant technological potential. Several metabolic engineering approaches have been developed, modifying the metabolic pathways in microalgae to redirect the flow of carbon toward isoprenoid biosynthesis, including pathway engineering, strain improvement, and synthetic biology. In this review, some beneficial features of these high-value metabolites are summarized. Besides, recent advancements in metabolic engineering approaches for the biosynthesis of isoprenoids are discussed in detail. At last, the viewpoints and challenges for the biosynthesis of novel compositions with isoprene units in the microalgae are also included.
Introduction
Isoprenoids (terpenes/terpenoids) are one of the most abundant and structurally highly complex natural products, which are utilized in various industries such as pharmaceutical, medical, nutraceutical, agricultural, fragrance, rubber, and advanced biofuels [78, 129]. Isoprenoids are hydrocarbons made up of two or more isoprene units, each consisting of five carbon atoms organized in a special pattern [13]. Several reactions, e.g., oxidation, reduction, rearrangements, cleavage of the rings, or cyclization, lead to the conversion of isoprenoid structures to cyclic or acyclic structures [29]. Hence, they are categorized based on the number of isoprene units appearing in their composition (e.g., hemiterpenoids (1 unit), monoterpenoids (2 unit), sesquiterpenoids (3 units), diterpenoids (4 units), sesterterpenoids (5 units), triterpenoids (6 units), tetraterpenoids (8 units), and polyterpenoids (> 8 units); [43]. In the configuration of all isoprenoid structures, isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (DMAPP) are the two key precursor molecules [14, 27, 128]. The 2-C-methyl-d-erythritol-4-phosphate (MEP) pathway, in bacteria and plastids of plant cells, and the mevalonate (MVA) pathway, in most eukaryotes, archaea, and certain bacteria, are responsible for the production of IPP and DMAPP [27]. In addition, IPP and DMAPP come from acetyl-CoA or pyruvate/glyceraldehyde-3-phosphate in the MVA or MEP pathway, respectively [146]. Currently, plants, bacteria, and animal tissues are the major resources in the production of terpenoids for industrial applications [15, 82, 89]. Previous publications have dealt with cultivation, harvesting, recovery, purification, extraction, and conversion, and have the ultimate intention to develop an economically feasible commercial process. Nevertheless, the high costs associated with the process and low overall productivity make the industrial terpenoids production from microbial sources unsuitable. As a way to address the limitations of unspecific isoprenoid production, which limits the specific yields of isoprenoids, metabolic engineering approaches have been applied as an efficient way to synthesize isoprenoids from microbial processes [58, 89]. Among microbial sources, microalgae have several clear benefits for isoprenoid production via metabolic engineering approaches over conventional microbial hosts such as Escherichia coli and Saccharomyces cerevisiae. First, a large number of microalgae have native MVA or MEP pathways and naturally create a variety of isoprenoids, which reduces the requirement for complex heterologous pathway construction [80]. Additionally, they can fix atmospheric CO₂ through photosynthesis, which minimizes the need for costly organic feedstocks and offers a sustainable, carbon–neutral pathway to isoprenoid biosynthesis [49]. Furthermore, subcellular compartmentalization in microalgae, as present in chloroplasts, allows pathways to be physically separated and allows for focused metabolic engineering to achieve higher yields of target compounds [133]. In addition, advancements in genetic tools, including Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) or CRISPR-associated (Cas) systems and organelle-specific promoters, have improved the metabolic flexibility and engineering potential of microalgae [30]. Microalgae also facilitate the co-production of isoprenoids with other valuable bioproducts, such as proteins and lipids; hence, promoting integrated biorefinery methods [95, 130]. Besides, as compared to traditional microorganisms as hosts, microalgae cultivation in selected conditions, like high salinity or light-dependent systems, lowers the risk of contamination. Finally, microalgae are a resource-efficient and environmentally friendly platform for industrial isoprenoid synthesis since they can be cultivated on non-arable land with wastewater and less freshwater [80, 86].
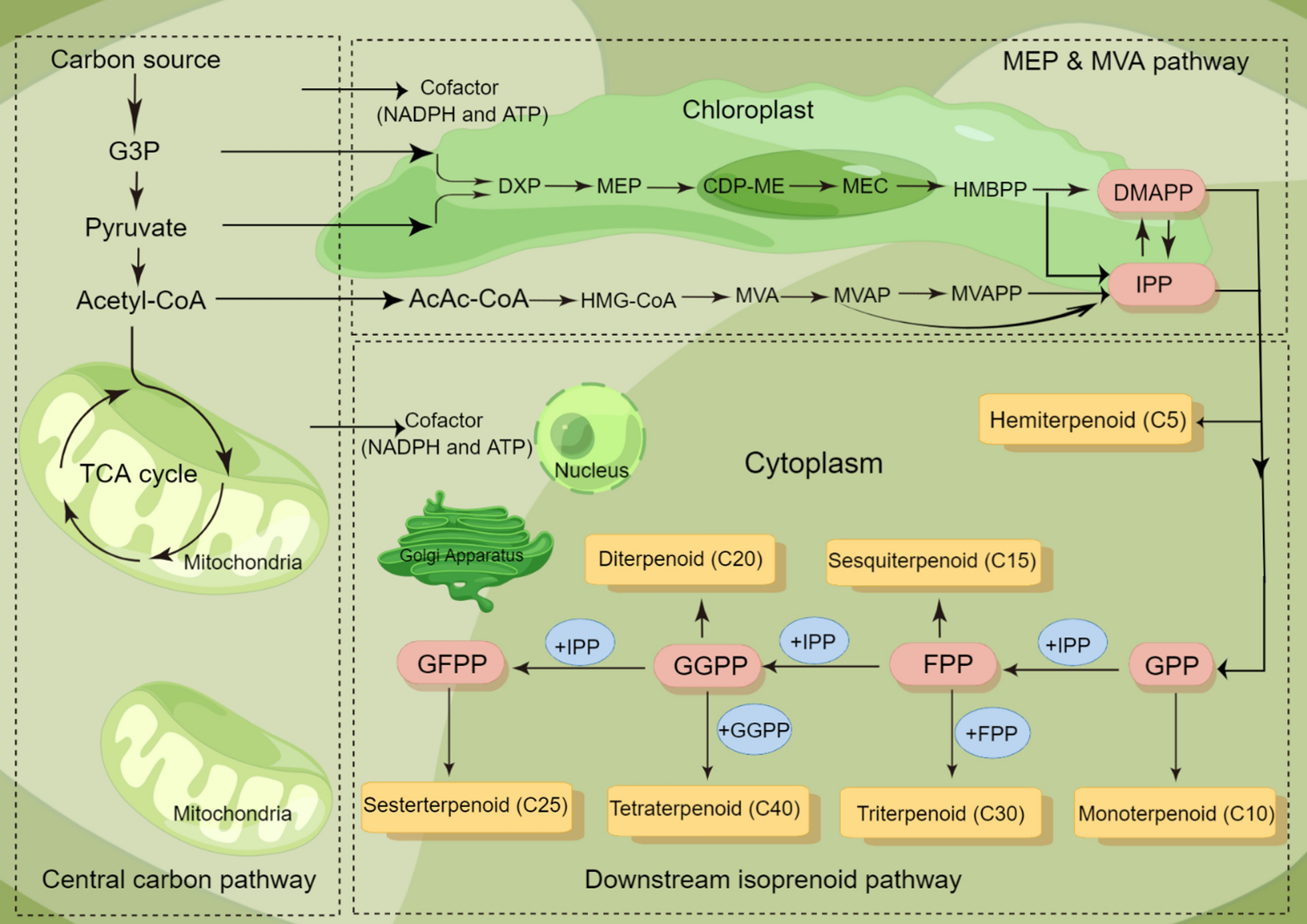
Table 1 presents numerous publications on microalgal bioprocess and biorefinery for sustainable isoprenoid synthesis at lower production costs, which have been conducted mainly in Chlamydomonas reinhardtii (C. reinhardtii) and Phaeodactylum tricornutum (P. tricornutum). Photosynthesis of microalgae requires the presence of chlorophyll and carotenoid pigments, which are derived from isoprenoid metabolism. Therefore, to ensure effective photosynthesis, a directed carbon flux towards the isoprenoid pathway in microalgae is needed. Isoprenoids synthesis pathway in microalgae is subdivided into three steps (Fig. 1). At the first stage, precursors and cofactors, e.g., glyceraldehyde-3-phosphate (G3P), pyruvate, acetyl-CoA, NADPH, and ATP, are produced either through glycolysis pathway (organic carbon sources) or Calvin cycle (inorganic carbon sources). In the second step, IPP and DMAPP are derived from MEP and MVA pathways and are sequentially condensed to each other through isoprenyl diphosphate synthases [27]. It is worth noticing that diatoms maintain both MEP and MVA pathways for the generation of the precursors, unlike other algae species that contain only one of the biosynthetic pathways (MEP and MVA) [7, 85]. For example, most green algae only maintain the MEP pathway [47, 100, 113]. Among red algae, the MVA pathway has been absent in the lineages of Cyanidioschyzon, Porphyridium, and Chondrus, but is present in Galdieria. In the third step, different kinds of prenyl diphosphates, including geranyl diphosphate (GPP), farnesyl diphosphate (FPP), geranylgeranyl diphosphate (GGPP), and geranylfarnesyl diphosphate (GFPP), are synthesized [78]. These are the trigger molecules for the hemiterpenes (C5), monoterpenes (C10), sesquiterpenes (C15), diterpenes (C20), sesterterpenes (C25), triterpenes (C30), and tetraterpenes (C40) [67]. Actually, this stage involves the linear condensation of isoprene units to produce prenyl diphosphates with varying chain lengths. Typically, prenyltransferases catalyze these reactions and lead to a wide range of complicated compounds that have been addressed [63, 69]. The cytochrome P450 superfamily of enzymes (P450s or CYPs) is involved in the subsequent modification steps, such as oxidation and hydroxylation, of these terpenoid compounds. Ultimately, the prenyl diphosphate molecules enter different pathways and turn out several products such as sterols, alkaloids, carotenoids, etc. [82].

The metabolic pathway of isoprenoid biosynthesis is divided into three distinct parts. The carbon flux to the isoprenoids pathway supplies the necessary precursors and cofactors for the MVA/MEP pathways, which then produce the common precursors DMAPP and IPP. These two precursors are then condensed to generate GPP, FPP, GGPP, and GFPP, which are catalyzed to form various isoprenoids. 2-C-methyl-d-erythritol-4-phosphate (MEP), mevalonate (MVA). Isopentenyl PyroPhosphate (IPP). DiMethylallyl PyroPhosphate (DMAPP). MeVAlonate-5-diphosphate (MVAPP). MeVAlonate-5-Phosphate (MVAP). 3-Hydroxy-3-MethylGlutaryl-CoA (HMG-CoA). AcetoAcetyl-CoA (AcAcCoA). (E)−4-Hydroxy-3-Methyl-But-2-enyl diphosphate (HMBPP). 2-C-Methyl-D-Erythritol 2,4-cyclodiphosphate (ME-cPP). 4-(cytidine-5ʹ-diphospho)−2-C-methyl-D-Erythritol (CDP-ME). 1-Deoxy-d Xylulose 5-Phosphate (DXP). Glyceraldehyde-3-Phosphate (G3P). Geranyl diphosphate (GPP). Farnesyl diphosphate (FPP). GeranylGeranyl diphosphate (GGPP). GeranylFarnesyl diphosphate (GFPP)
There are many studies to promote the production of carotenoids, phytosterols, triacyglycerol, and provitamin A over MVA and/or MEP ways via genome modification in plants [66]. Perpetuated structures, species-dependent molecules, and prominent metabolic and physiological characteristics, proposed microalgae as an excellent origin of isoprenoids among all natural sources [7]. Nowadays, growing information and data availability from genomic and transcriptomic studies the isoprenoids biosynthesis opens for possibility for genetic manipulation of microalgal metabolism [62]. Figure 2 presents the current state of the art concerning the approaches for promoting and enhancing the bioproduction of high-value metabolites in microalgae, including natural diversity, growth, and productivity optimization combined with omics technologies and genetic engineering tools, with a focus on application in a variety of industrial sectors.

An overview of microalgae strain development using various tools/approaches
The main strategy is supplying more precursors, mainly IPP and DMAPP, through metabolic engineering strategies.
The main five strategies that have been extracted from the studies and experiments to overcome precursor limitations are [16, 48, 146]:
- 1.
Overproduction of the enzymes in the rate-limited reactions in the biosynthetic pathway of isoprenoids.
- 2.
Knockout of some deficient genes and regulators in the biosynthetic pathway.
- 3.
Precursor production by engineered host cells through new metabolic pathways.
- 4.
Adequate presence of cofactors in biosynthetic routes.
- 5.
Diminishing the rate of reactions that lead to the production of unfavorable metabolites, thus increasing the production of desirable products and essential intermediates from the metabolic pathways.
Overcoming the availability of precursors for the wide range production of isoprenoids is one of the major issues facing researchers. On the other hand, formation of certain isoprenoids (species-dependent) in commercial scale in a heterogeneous host is also of great importance.
This review initially describes some applications of well-known isoprenoids in food, energy, pharmaceuticals, and cosmetics industries. Then, the metabolic engineering strategies progression to produce isoprenoids through the biosynthetic pathway of microalgae is provided. To overcome the limitations in the sustainable production of microalgae-associated isoprenoids, enormous efforts and techniques have been developed to enhance their commercial-level production, which has also been discussed. In the last section, some of the challenges and the potential developments for advancing the production of isoprenoids are identified.
Isoprenoids: healthful and therapeutic biocompounds
Isoprenoids or terpenoids have several important roles as protective agents, accessory and photoprotective pigments, toxins (neurotoxin), and sterols (Table 1) [7, 19, 145]. In addition there are some highly branched isoprenoid (HBI) that have several health effect such as anticancer, anti-HIV, and cytostatic [7, 93]. However, the production of terpenoids that have medical and health properties is very low. The majority of terpene compounds with health properties are nowadays taken directly from plants. However, the amount of terpene compounds in plants is modest, and their regulating mechanism is unknown [37]. Furthermore, there are many challenges with the extraction and purification of these valuable substances. Thus, there is an emerging need for biotechnological production of these valuable compounds from microbial cell factories. A new technology based on bacteria or yeasts as hosts for non-native production of isoprenoid metabolites is under development [74]. In addition, microalgae can be another suitable host for sustainable heterologous production of isoprenoids due to their easy cultivation by using light and CO2 as energy and carbon source [20, 74]. However, inadequate knowledge of the genomes of algae has led to slow improvement of the biosynthesis processes by genetic engineering methods (e.g., Chlamydomonas reinhardtii or Phaeodactylum tricornutum [74]. The green alga Botryococcus braunii synthesizes some triterpenes, e.g., botryococcene and squalene oils, that are used in the petrochemical industry [101, 131]. According to heterologous gene expression in microalgae from various organisms, e.g., plants, animals, fungi, or bacteria [3], and with respect to this point that most of the microalgae have not been chemically specified, novel isoprenoid structures may emerge through future studies. In the following sections, some of the fundamental isoprenoids derived from microalgae and their applications are introduced.
Diterpenes
These compounds are consisted from two terpene parts, comprises of 20 carbon atoms and is one of the largest category of natural compounds with more than 18,000 products [114]. This group of isoprenoids have several structures with diverse bioactivities. Generally, diterpenoids may contain linear, monocyclic, bicyclic, tricyclic, or tetracyclic molecules [8]. They are produced from plants, animals, microalgae, some diatoms, and fungi through the mevalonate pathway, with geranylgeranyl pyrophosphate as precursor. Important diterpenoids are retinol, retinal, and phytol, which have antimicrobial and anti-inflammatory [11]. Taxadiene, ingenol-3-angelate, and forskohlin are also valuable diterpenoids with medicinal or nutraceutical properties [4, 74, 106]. Forskolin, the anticancer and antiobesity metabolite, is derived from 13R(+) manoyl oxide. Lauersen et al. [74] over-expressed diterpene synthases (diTPSs) and enzymes associated with the MEP pathway and produced heterologous diterpenoids, casbene, taxadiene, and 13R(+) manoyl oxide from the model microalgae Chlamydomonas reinhardtii. Taxadiene is the first committed intermediate in the synthesis of paclitaxel (Taxol). Amination of the natural tetracyclic diterpenes leads to the formation of diterpenoid alkaloids (DAs), which have complex structural properties with many stereocenters [114]. DAs form a vast group of natural metabolites with remarkably complicated structures and biological activities, which are also notoriously toxic. They have several pharmacological properties, including analgesic, antiarrhythmic, anti-inflammatory, local anesthetic, anti-arrhythmic, muscle relaxant, hypotensive, antimicrobial, antitumor, spasmolytic, psychotropic, and neurotropic [114, 135]. Although microalgae have great potential for the production of geranylgeranyl pyrophosphate (GGPP), a common precursor of pigments and diterpenoids, the production of heterologous diterpenoids in eukaryotic microalgae is still in its beginning stages.
Sesterterpenes
Sesterterpenes (C25) which contain 25 carbon atoms are the rare found compounds among terpene. Geranylfarnesyl diphosphate (GFPP) is the linear precursor for the polycyclic structures of sesterterpenoids. These compounds have several biological activities such as anticancer, anti-inflammatory, antiprotozoal, antitubercular, antimicrobial, and modulation of receptor signaling [93].
Sesterterpenoids can be extracted from variety of natural resources including fungi, bacteria, lichens, higher plants, insects, various marine invertebrates, especially sponges, and some diatoms [77, 93]. Massé et al. [93] reported that the diatom Rhizosolenia setigera produces highly branched isoprenoid (HBI) sesterterpenes (haslenes). In addition, Rowland et al. [110] manifested in vitro anticancer activity of the haslenes obtained from the diatom Haslea ostrearia.
Triterpenes
The structure of triterpenoids (C30) comprises of 30 carbon atoms, which make up six isoprenoid units. Over 100 triterpenoid compositions have been recognized until now, including α-amyrin, β-amyrin, dammarene-diol, and lupeol [25]. These are the precursors for the compounds used for anticancer or HIV treatment. Several metabolites under the triterpenes family can be synthesized through primary or secondary metabolism. Sterols, as the only primary substance, play an important role in adjusting the cell membrane dynamics. In addition, sterols produce secondary compounds such as steroid hormones that control human growth and development [22]. Other secondary triterpenes are phytosterols and ergosterol, applied for decreasing the blood low-density lipoprotein (LDL) cholesterol and producing vitamin D2 (ergocalciferol), respectively [45, 55]. Squalene, high-value added compound with numerous applications in pharmaceutical, medical, cosmetic, and food industries, convert to (S)−2,3-epoxysqualene, which is the precursor in triterpenoids biosynthesis [22]. Lupette et al. [90] identified brassicasterol, structurally same as cholesterol, as an element of lipid droplets (LDs) surface in nitrogen starved Phaeodactylum tricornotum. Other than free sterols, there are several microalga and diatom species that synthesize conjugated types of these compounds. For instance, Thalassiosira pseudonana, Chaetoceros calcitrans, and Haslea ostrearia contain esterified form of sterols, glycoconjugated (steryl glucosides and acylated steryl glucosides), and 23,23-dimethylcholest-5-en-3β-ol, respectively [7]. Ergosterol, usually produced in fungi, is observed in P. tricornotum sterol biosynthetic pathway [36]. Cylindrotheca fusiformis and Nitzschia closterium contain cholesterol that is generated by animals in most cases [109].
Tetraterpenes (carotenoids)
Essential roles of carotenoids, composed of 40 carbon atoms (C40), in human life attract the attention of researchers for discovering the new strategies in their biosynthesis. Eight isoprene units are the building block of these pigments [99, 142]. Carotenoids are divided into (a) carotenes (containing carbon and hydrogen) and (b) xanthophylls (oxygenated derivatives of carotene). Color and antioxidant properties of carotenoids are owing to the conjugated double bonds in their structures. These compounds constitute a vast category of tetraterpenoids that contain several applications in industrial fields [96] such as anticancer, antioxidant, antiangiogenic, antidiabetic, antiobese, anti-inflammatory, etc. Carotenes, e.g. lycopene and β-carotene, are applied as food additive in food industry and play an important role as an antioxidant, antimalarial, and anticancer agents [42, 54, 140]. Fucoxanthin is another tetraterpenoids that plays as an accessory light harvesting xanthophyll, originated from β-carotene and transfer the light energy to the photosynthetic system [70]. Fucoxanthinol, deacetylated form of fucoxanthin, has manifested great potential in the prevention and treatment of many types of cancer [92]. Regardless of its diverse applications, the extraction of fucoxanthin from its natural resources such as alga, specially macroalga, is low and its chemical production is troublesome [38]. Manfellotto et al. [91] by exploitation genetic transformation enhanced the expression of the genes attends in the carotenoid biosynthetic pathway of P. tricornotum. Some of the other well-known tetraterpenes are astaxanthin, diatoxanthin, and diadinoxanthin that can be extracted from microalgae and have many biological activities [96, 111].
Metabolic engineering approaches enhance the biosynthesis of isoprenoids
Reconstruction and optimization of the metabolic pathways to enhance the bio-production of the isoprenoid molecules through metabolic engineering and synthetic biology approaches has attracted the attention of many researchers in recent years. Even though the majority of studies has centered around C. reinhardtii and P. tricornutum, other microalgae such as H. pluvialis, D. salina, Nannochloropsis oceanica, Chlorella vulgaris, Scenedesmus sp., Thalassiosira pseudonana, and Porphyridium purpureum have also been explored due to their economic value [60]. The planet is host to approximately ten million algal species, representing an abundant potential bioengineering asset. However, taking advantage of these will likely necessitate further knowledge of their genetics combined with the latest advances in omic approaches such as transcriptomics, proteomics, metabolomics, and phonemics to select the desired gene. Recent reviews provide an all-inclusive look at these resources, with information on the species, mutants, omics datasets, and genetic tools at our disposal [72, 118]. Despite this, low levels of isoprenoid production in microalgae are the principal impediments currently preventing commercial development. Nevertheless, terpene synthases have been successfully expressed heterologously in microalgae, indicating their potential as a production platform.
Currently, methods to augment isoprenoid engineering are mainly limited to overexpressing rate-reducing enzymes. So, the technical progressions in gene editing tools and novel design tactics are imperative. Table 2 provides a summary of the metabolic engineering investigations for the production of isoprenoids in microalgae that have been reported in the last five years.
Precursors availability
The native glycolysis pathway in microbial hosts generates the essential precursors for isoprenoid production, namely G3P, pyruvate, and acetyl-CoA (Fig. 1). To ensure sufficient precursor availability for isoprenoids production, implementation of genetic engineering strategies on host cells is necessary. When the MVA pathway is modified for isoprenoid production, strategies should be implemented to maximize acetyl-CoA accumulation. Deleting or downregulating genes associated with byproduct formation presents a viable option [78]. Down-regulation of the genes aldose reductase (AR), galactose dehydrogenase (GALDH), lactate dehydrogenase (LDH), acetaldehyde dehydrogenase (ADH), pyruvate oxidase (POX), and acetyl-CoA carboxylases (ACCs) can result in more carbon being available for enhanced isoprene synthesis [61].
There are also some reports about introducing the new precursor synthesis pathway/carbon source, e.g., heterologous acetyl-CoA synthetic pathway introduced into S. cerevisiae to boost the levels of acetyl-CoA. The dual utilization of mitochondrial and cytoplasmic acetyl-CoA was applied in S. cerevisiae, leading to an increase in isoprene production of approximately 2.1- and 1.6-fold, respectively, greater than those strains generated by mitochondrial or cytoplasmic engineering alone [81, 94]. When the MEP pathway is implemented for isoprenoid production, a major limitation is an unequal supply of G3P and pyruvate, as G3P gets converted to pyruvate through multiple steps in glycolysis (Fig. 1). Hence, G3P needs to be balanced toward the biosynthesis of isoprenoids in microalgae. In addition to G3P, pyruvate, and acetyl-CoA in the biosynthesis of isoprenoids, the limited availability of the two main intermediate precursors, IPP and DMAPP generated via MEP or MVA pathway, also diminishes the productivity of the desired isoprenoid products. The synergistic effect of two modifications was investigated to increase isoprene production in Synechocystis from 0.2 to 12.3 mg g−1. The first modification increased intermediate precursor (DMAPP) availability by overexpressing fni from Streptococcus pneumonia, and the second increased cellular IspS protein concentration by expressing IspS as a fusion construct with the cyanobacterial cpcB gene [61]. Eukaryotes, fungi, and some plants function through the MVA pathway in order to produce IPP and DMAPP, while bacteria and cyanobacteria possess the MEP pathway. Unlike these species, diatoms retain both MEP and MVA pathways and convert the obtained precursors to end-products [78]. According to transcriptomic sequencing of numerous species and isotope tracer analysis, it was verified that diatoms generate IPP and DMAPP through both MEP and MVA routes [7, 85]. This causes higher metabolic flux and greater storage of available precursors for the production of isoprenoids [124]. Cvejić et al. [24] presented P. tricornutum and Nitzschia ovalis produced epibrassicasterol and cholestadienol (sterols) through the MVA pathway by consuming acetate as a carbon source under mixotrophic conditions. Furthermore, they converted the IPP/DMAPP obtained precursors from CO2 to phytol via the MEP route [24]. In another study, the human Gult1 gene was expressed in P. tricornutum to create a glucose-using heterotrophic transformant. The Embden–Meyerhof–Parnas (EMP) route was frequently embolized, and excessive G6P accumulation inhibits the Gult1 transformant’s heterotrophic development. In the Gult1 transformant, the 6-phosphofructokinase (Pfk), glucose-6-phosphate dehydrogenase (G6pd), and 6-phosphogluconolactonase (Pgl) genes were expressed to boost the EMP and Pentose Phosphate Pathway (PPP) for accelerating G6P metabolic rate. Under heterotrophy, Pfks expressed transformants have 14–33% more biomass than Gult1 transformants and 4 times more biomass than autotrophy. Under heterotrophy, the G6pds and Pgls expressed transformants have 40% more biomass than the Gult1 transformant and 4.5 times more biomass than autotrophy condition [143]. Skeletonema marinoi and Cyclotella cryptica formed sterols via cyclization of squalene, the first intermediate in the generation of sterols, from mevalonate pathway [41]. Rhizosolenia setigera is another diatom that uses MVA pathway in order to produce sterols and HBIs. While, H. ostrearia supplies HBIs and β-sitosterol through MEP rout [93].
Engineering associated enzymes in the biosynthetic pathway of isoprenoids
Constructing a multigene pathway according to the strategies outlined may lead to several enzymes in the MEP and MVA pathways limiting metabolic flux, requiring the optimization of rate-limiting enzymatic conversions. Depending on the engineered strain, these restrictive points may vary. To identify target enzymes for optimization, kinetic flux profiling has been used to identify bottlenecks in an engineered isoprene production strain. After identifying rate-limiting enzymes, various engineering methods have been developed to optimize key enzymes. Screening of enzymes from different species has been utilized to choose the most appropriate enzyme [78].
There are diverse enzymes that might limit the metabolic flux of the MEP and MVA pathways, thereby diminishing precursors supply and finally isoprenoid production. Isopentenyl phosphate kinase (IPK), iso-pentenyl diphosphate isomerase (IDI), 1-deoxy-d-xylulose 5-phosphate synthase (DXS), 1-deoxy-d-xylulose 5-phosphate reductoisomerase (DXR), 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase (MDS), 4-hydroxy-3-methylbut-2-en-1-yl diphosphate reductase (HDR) are some of the main rate-limiting enzymes in pathway under various conditions and organisms [7, 10, 23, 78]. Henry et al. [51, 52] represented the regulatory role of IPK and two nudix hydrolases in contributing of isopentenyl phosphate (IP) and IPP production through active phosphorylation/dephosphorylation reactions. In fact, IP to IPP ratio influences the productivity of isoprenoids obtained from MEP and MVA and this indicates interference in the crosstalk mechanism between the two paths. The regulatory roles of these enzymes were observed in several diatoms [6]. One of the outstanding features of most diatoms is that they contain the enzyme IDI, which is integrated into the squalene synthase (SQS), that accelerates the first stage of sterol biosynthesis [28]. In addition, P. tricornotum, R. setigera, and H. ostrearia catalyze the sterol biosynthesis wherein R. setigera and H. ostrearia FPP synthase and IDI have located in the endoplasmic reticulum (ER) membrane together and this helps the effective conversion of precursors to sterols [28]. In the MEP pathway, the step catalyzed by 1-deoxy-D-xylulose 5-phosphate synthase (DXS) is one of the major limitations. For example, the DXS mRNA expression improved when dark-adapted P. tricornotum cells transferred to light and this issue promoted the production of carotenoids through enhancing the precursors flow. Overexpression of DXS in P. tricornotum increased the production of fucoxanthin in a 2.4 fold in comparison with the natural strains to with more accumulation of other intermediates such as diadinoxanthin and β-carotene [31]. According to this research, it was confirmed that DXS is a rate-limiting enzyme in diatoms and leads to more metabolic engineering experiments to improve the isoprenoid biosynthesis in P. tricornotum. As mentioned previously, most of the diatoms have the enzyme that isomerizes IPP to DMAPP with integration of squalene synthase (IDI-SQS). The reductive dehydroxylation of (E)−4-hydroxy-3-methylbut-2-enyl diphosphate (HMBPP) to IPP and DMAPP is carried out by the enzyme HDR. Diatoms have cytosolic IDI, the enzyme that isomerizes IPP to DMAPP, in a fusion with squalene synthase, as was mentioned earlier (IDI-SQS). While some species have an additional IDI that is oriented at the plastids, other species do not have a second IDI [39]. If these data are accurate, HDR must be an essential component of the plastids’ isoprenoid synthesis, modulating the ratio of IPP to DMAPP for subsequent reactions. Although catalytic residue (F212) at the enzyme’s active site is essential to produce the final product in diatoms, more in-depth research on HDR enzymes from various species is required to determine the precise IPP:DMAPP ratio formed. Based on Shin et al. [115], before the product is released, this residue mediates the proton transfer to the intermediate allyl anion [115]. Species that lack an independent IDI must have developed unique mechanisms for the isomerization of IPP to DMAPP in plastids if this is inadequate to support the synthesis of plastidic isoprenoids. Examples of such mechanisms include the dual targeting of IDI-SQS or the alternative splicing of the IDI-SQS transcript. Another metabolic engineering approach, which is utilized to enhance the production of fucoxanthin and its precursors, is introducing phytoene synthase, a regulator of the metabolic fluxes towards carotenoid synthesis, into the P. tricornutum genome [7]. To metabolically engineer cells to produce particular isoprenoids, an ideal IPP:DMAPP ratio must be achieved because the desired molecule depends on this ratio. For instance, sesquiterpenoids or sterols need a 2:1 ratio, but diterpenoids or carotenoids need a 3:1 ratio.
In MVA-containing eukaryotic cells, hydroxy methyl glutaryl reductase (HMGR) is known as the key regulator and rate-limiting enzyme in the early production of sterol and nonsterol isoprenoids, and it is extensively controlled at the transcriptional, translational, and post-translational levels [56]. Jaramillo et al. [56] found that the overexpression of two versions of the native HMGR and a conventional, heterologous squalene epoxidase (SQE) gene in the diatoms Phaeodactylum tricornutum resulted in significant differential accumulation of downstream sterol pathway intermediates. Squalene, cycloartenol, and obtusifoliol accumulated in response to HMGR-mVenus overexpression, whereas cycloartenol and obtusifoliol accumulated in response to heterologous NoSQE-mVenus overexpression. Furthermore, all P. tricornutum overexpression lines accumulated the end-point sterol 24-methylenecholesta-5,24(24ʹ)-dien-3-ol, and campesterol increased threefold in P. tricornutum lines expressing NoSQE-mVenus [56]. HMGR and IDI SQS were two of the genes that were discovered to be continuously increased during the heterologous expression of plant triterpenoid biosynthesis genes in P. tricornutum [25]. They are, therefore, excellent candidates for future pathway optimization research.
In a recent study, Fabris et al. [35] investigated the heterologous production of the monoterpenoid geraniol in P. tricornutum, which is known for its use in perfumes and insect repellents (Table 2). Despite the presence of a GPP (C10) in this diatom, a precursor of geraniol, it was unable to produce geraniol on its own. To overcome this obstacle, the researchers expressed Catharanthus roseus geraniol synthase (CrGES), previously verified as the most effective enzyme for geraniol synthesis in S. cerevisiae, into the cytosol of P. tricornutum using an episome vector under the control of both an inducible alkaline phosphatase gene promoter (AP1p) and an endogenous constitutive promoter (Phatr3_J492020p). This method ultimately led to high yields of 48.60 μg L−1 and 309 μg L−1 of geraniol production, respectively, thereby providing proof-of-concept that extrachromosomal expression through plasmids can be used as a useful tool for future metabolic engineering endeavors in P. tricornutum.
Impediment of the competitive pathway
The isoprenoid synthetic pathway in microalgae has implications that influence cell growth, resulting in the formation of alternative and competitive pathways for the synthesis of target isoprenoids. To maximize isoprenoid production, it is necessary to redirect carbon flux away from these bypass and competitive pathways. However, when the bypass pathway is essential for cell viability, complete knockout of the enzyme can be lethal [125, 129, 137]. Hence, for a successful engineering plan utilizing carbon redirection the physiological effects on cell growth and reproduction must be taken into account by analyzing a variety of output metrics. Comprehending the functioning of homeostasis in engineered cells necessitates a thorough understanding of central carbon metabolism. To address this issue, downregulation strategies at the transcriptional level have been proposed including the selection of a weaker promoter or the use of RNA interference technology [78]. For sesquiterpenoid production from FPP, squalene production from FPP catalyzed by SQS (ERG9) is considered a bypass pathway that needs to be inhibited. Wichmann et al. [132] managed to produce (E)-α-bisabolene, a sesquiterpene biodiesel precursor, in the C. reinhardtii cytosol. This was accomplished through threefold overexpression of Abies grandis bisabolene synthase (AgBS), which converts FPP to (E)-α-bisabolene, as well as knockdown of the ADP-glucose pyrophosphorylase small subunit (AGPP), farnesyltransferase (PFT), and squalene synthase (SQS). Through this strategy, the engineered strain achieved a 15-fold increase in titer at 10.3 ± 0.7 mg g−1 DCW as compared to single expression of AgBS. Furthermore, under light/dark cycle conditions, the titer further increased to 11 mg L−1 due to the increased availability of FPP for ubiquinone synthesis in the mitochondria during dark cultivation for respiration. Similarly, Kajikawa et al. [59] attempted to overexpress endogenous squalene synthase and downregulate squalene epoxidase in order to increase squalene production.
Introducing novel pathway for accumulating precursors
The engineering strategies outlined above are implemented using the native MEP and MVA pathways. To minimize the metabolic load associated with multi-gene expression and the buildup of toxic intermediates, new pathways have been developed. Multiple studies leveraging the novel MVA pathway to generate isoprene have been reported in the literature, wherein fatty acid decarboxylase and oleate hydratase were incorporated to create isoprene from mevalonate in two steps [136]. In novel pathway I, mevalonate-5-pyrophosphate decarboxylase (MDD) catalyzes the isoprenol production reaction from mevalonate. In novel pathway II, MDD catalyzes mevalonate-5-phosphate (MVAP) to produce IP, and phosphatase subsequently catalyzes a reaction which produces isoprenol. The low activity and affinity of fatty acid decarboxylase, oleate hydratase, and MDD resulted in a decreased production of isoprene or isoprenol. Hence, further studies to identify enzymes with better activity and higher affinity are essential. In comparison to the novel MVA pathway, there has been limited research on the novel MEP pathway. Hence, the lack of efficient enzymes in these pathway continues to hinder its application, necessitating enzyme screening and engineering to improve metabolic flux [44].
Novel engineered pathway for hemiterpenoid production has been largely confined to the production of isoprene and isopentenol, simple five-carbon molecules that do not involve IPP/DMAPP condensation [78]. Consequently, examining a novel pathway which eliminates the long pathway but does not bypass IPP and DMAPP synthesis would be more valuable and could be applied to all types of isoprenoid production. In two studies, a two-step pathway for IPP and DMAPP generation from prenol and isoprenol was discovered. Two kinases were identified that catalyze the transformation of prenol and isoprenol to DMAP and IP, respectively. This separate metabolic route from the main carbon metabolism has potential for high metabolic flux, with Stephanopoulos’group’s pathway showing adequate metabolic flux for nearly all downstream pathways tested. Nonetheless, further research is needed to supply inexpensive and sustainable supplies of prenol and isoprenol substrates in order to enable utilization of this novel pathway for additional isoprenoid synthesis [15, 88].
In addition to the native pathway, another novel pathway for the synthesis of 1-deoxy-d-xylulose-5-phosphate (DXP) of the MEP pathway from ribulose 5-phosphate (Ru5P) through a reaction catalyzed by YajO, a putative xylose reductase, and mutant RibB was investigated by Kirby et al. [68]. This novel pathway does not rely on the central carbon system and bypasses the rate-limiting step, which catalyzes the conversion of G3P and pyruvate to DXP. Consequently, it is promising for high metabolic flux towards the isoprenoid pathway without diminishing normal cell growth [68]. Thus, precise control and regulation of biosynthetic pathways have been made possible by the employment of synthetic biology technologies, such as dynamic gene circuits and synthetic promoters, particularly in the synthesis of high-value isoprenoids. In this regard, there is notable research by Einhaus et al. [33], which focused on rational promoter engineering in C. reinhardtii. They created novel synthetic expression elements by systematically evaluating existing expression elements and integrating them with rational promoter engineering. This approach revealed a synergy between the PSAD 5′ untranslated region (UTR) and its corresponding chloroplast targeting peptide, and it also enhanced the uniform application of synthetic biology techniques. The developed synthetic algal promoter enhanced the production of the sesquiterpene (E)-α-bisabolene by four times when compared to commonly used expression elements and eighteen times when compared to its native form [33].
These techniques greatly increase yield and stability by enabling real-time adaptation to metabolic or environmental signals. Algal biotechnology is undergoing a paradigm shift as a result of these developments, shifting toward more reliable, scalable, and effective engineering techniques.
In the end, proper selection of a microalgal host for biotechnological applications critically depends on its isoprenoid biosynthesis pathways. Microalgae such as diatoms possessing both MVA and MEP pathways offer distinct advantages over MEP-only species. For the synthesis of high-value isoprenoids, such as terpenoids, carotenoids, and sterols, the dual-pathway architecture offers improved precursor availability and compartmentalized flux distribution, resulting in increased metabolic flexibility [6]. In addition, when compared with the plastid-confined MEP system, the cytosolic location of the MVA pathway enables more accessible and effective metabolic engineering approaches. On the other hand, the MEP pathway is preferred for CO2-based isoprenoids production because it is stoichiometrically more efficient, requiring just six molecules of CO2 for the synthesis of either IPP or DMAPP, in comparison to the nine molecules needed for the MVA [32]. These advantages must be balanced, though, with the technical drawbacks of diatoms, which include complex regulatory systems and less advanced genetic tools than those of model green algae [18]. Nevertheless, microalgae with the MVA route have greater potential for producing isoprenoids, their efficient use necessitates ongoing developments in systems biology and genetic engineering technologies. The precise biosynthetic objectives and the organism’s engineering capabilities should therefore serve as guidance when choosing the best host.
Emerging metabolic engineering tools
In the last 10 years, significant progress has been achieved in expanding the range of tools available for genetically modifying microalgae, increasing their potential as model organisms for use in both industrial and scientific applications. These include Zinc-Finger Nucleases (ZFNs), Transcription Activator-Like Effector Nucleases (TALENs), and (CRISPR/Cas) tools [1, 2]. These techniques mostly cause DNA double-strand breaks, which in turn initiate the cell’s natural DNA repair mechanisms. By applying this method, donor DNA can be added for transgenic integration, or targeted genes can be disabled.
ZFNs are artificial proteins that bind DNA and have two domains connected by a linker sequence. Through a zinc-finger DNA binding domain and a restriction endonuclease FokI-mediated DNA cleaving domain, they offer sequence specificity [5]. However, because of their complexity, difficulties, and expense, their application has been restricted, leading scientists to create a more straightforward, efficient, and economical technique for precise genome modification.
TALENs are specially designed tools that merge DNA-binding domains from TALE proteins with the FokI cleavage domain. Each repeat domain of these transcription activator-like effectors (TALE) proteins can recognize a single base pair. In order to allow precise genome editing, TALENs induce specific double-strand breaks (DSBs) in DNA, which initiate repair processes. The TALENs technology functions essentially as a specially designed restriction enzyme that can target particular genomic sites. Meganucleases and TALE nucleases were both employed in a recent study to change the diatom P. tricornutum. The transformed strain produced more oil as a result of their effective editing of seven genes necessary for lipid synthesis [26, 40, 71]. This highlights how TALENs can be used to precisely alter genes and improve desirable characteristics in microalgae.
Although all of these genetic modification tools have created new opportunities for the bioproduction of economically significant compounds using microalgae, CRISPR is the most promising technology for the future and is preferred in microalgae genetic engineering. Scientists are able to identify the roles of several genes in metabolism, growth, and other cellular processes of microalgae by employing CRISPR technology, which allows for specific gene modification or knockout, specifically in diatoms like P. tricornutum. Furthermore, CRISPR-based editing has been utilized to produce microalgal strains with superior characteristics for a range of applications, including higher biomass productivity, improved stress tolerance, and increased lipid and pigment production. Despite some challenges, the CRISPR/Cas technique has been effectively used for developing microalgal strains for several kinds of applications in industry. In biofuel production, researchers enhanced overall lipid productivity in C. reinhardtii by 64.25% without noticeably altering cell growth when they knocked out the phospholipase A2 gene, leading to a greater diacylglycerol pool and increased triacylglycerol accumulation [123]. In another study, the genes involved in acyl-CoA use, including fatty acid synthesis, β-oxidation, and autophagy, were impacted by functional knockout of the acyl-CoA binding protein (ACBP), especially when cells were dependent on endogenous carbon deposits. Triacylglycerols (TAGs) lipolysis was also inhibited by knocking out this gene, and this was linked to the inhibition of lipid droplets/TAGs degradation observed in previous studies [76]. CRISPR is also used for enhancing photosynthetic efficiency and pigment production. Recently, the CpSRP54 gene, which is essential for P. tricornutum’s indirect carbon fixation pathways, has been targeted using a CRISPR-based strategy. This alteration highlights the potential of CRISPR/Cas9 to fine-tune metabolic pathways that are essential for optimizing carbon utilization and storage, hence increasing the organism’s applicability in industrial applications [75]. However, industrial applications of microalgae need to achieve the same growth rate as bacteria and yeast. One significant development is the one-step transformation of C. reinhardtii using a DNA-free CRISPR/Cas9 technique, which allowed the CpFTSY and ZEP genes to be sequentially knocked out. In dense cultures, light penetration was improved by the reduction in chlorophyll antenna size caused by the CpFTSY gene deletion. Moreover, a strain with a markedly elevated zeaxanthin production without compromising photosynthesis was produced by altering the ZEP gene. This method is crucial for optimizing biomass production in systems of large-scale cultivation. Targeting the photosynthetic system in microalgae not only improves light usage but also increases pigment yields for commercial applications. Since there are many pigments associated with the photosynthetic system, such as carotenoids, astaxanthin, lutein, phycoerythrin, and phycobiliproteins, which are useful for human consumption [57].
Techno-economic and life-cycle assessment of microalgae-based isoprenoids
Techno-economic analysis (TEA) and life-cycle assessment (LCA) are essential tools for the evaluation of the industrial viability of microalgae as a platform for isoprenoid production. Cost-effectiveness of the recent TEAs involves several factors such as biomass productivity, downstream processing efficiency, capital investment, and operating expenses. Based on the recent findings, it suggests that while microalgae-based systems are currently more expensive than conventional microbial platforms, their costs can be significantly reduced through advances in metabolic engineering strategies and process integration for co-product recovery (e.g., lipids or proteins) as mentioned earlier [65, 116]. In parallel, LCAs evaluate the environmental sustainability of microalgal isoprenoid production, accounting for energy input, greenhouse gas emissions, water use, and land footprint. Several LCA studies have demonstrated that microalgal production systems, particularly those using CO₂ capture and wastewater as inputs, offer lower environmental impacts compared to fossil-based or sugar-fed microbial processes. However, the energy-intensive nature of harvesting and drying algae remains a key environmental hotspot. Integrating renewable energy sources and optimizing bioprocesses via engineered strains can further enhance the sustainability profile. Together, TEA and LCA highlight that while microalgae-based isoprenoid production is not yet cost-competitive at commercial scale, it holds strong long-term potential given its renewability, co-product synergy, and carbon neutrality [79].
Conclusions and future perspectives
Microalgae have the potential to be employed as a sustainable source of biochemical and bioactive compounds using organic and inorganic carbon sources. The recent advancements in genetic engineering have enabled the engineering of microalgal strains to increase both biomass productivity and the yield of high-value products. Various studies have evaluated the metabolic engineering strategies adopted to optimize the MEP and MVA pathways for isoprenoid production in microalgae. This review summarizes findings published in the recent five years. Engineering of isoprenoid biosynthesis in heterologous hosts offers the potential to produce adequate quantities and various structures of these compounds to make them industrially and commercially applicable. Despite considerable progress, microalgal metabolic engineering has encountered various challenges. To date, random integration has been the primary method of microalgal genetic engineering as episomal plasmid expression and homologous recombination are not available to most microalgae. As a result, transgene integration sites have led to unexpected and inconsistent phenotypes. In addition, unstable expression and silencing of transgenes have impeded the development of microalgae metabolic engineering. Fortunately, innovative gene editing tools such as RNAi, CRISPR/Cas9, ZNFs, and TALENs or intron-mediated gene expression are promising for producing more sophisticated and stable transformants in the future [103, 119]. Besides, advancements in metabolic engineering of microalgae could be facilitated by gaining a comprehensive understanding of the location and purpose of specific metabolic pathways, as well as having the ability to manipulate them. However, compared to simple organisms like bacteria, it remains a challenge to manipulate the genome of microalgae. Finally, difficulties arise when attempting to find low-cost and eco-friendly techniques for extracting all bioactive components from microalgal cells. Furthermore, some economic and environmental constraints hinder large-scale applications.
Implementation of metabolic engineering approaches in microalgae offers novel platforms for the generation of terpenoids requiring less expensive growth conditions and possessing a background metabolism that may be more suitable in comparison to the existing microbial production platforms. Despite the considerable progress made in the past decade in elucidating the biosynthesis of isoprenoids within the microalgal group, there are still certain biosynthetic pathways that have to be investigated. Exploring these unknown pathways could further enhance our comprehension of the entire process and distinguish between microalgae-associated products.
Availability of data and materials
No datasets were generated or analysed during the current study.
References
-
Ahmad Kamal AH, Mohd Hamidi NF, Zakaria MF, Ahmad A, Harun MR, Chandra Segaran T, Jusoh M. Genetically engineered microalgae for enhanced bioactive compounds. Discov Appl Sci. 2024;6:482.
-
Ambily B, Limna Mol VP, Sini H, Nevin KG. CRISPR-based microalgal genome editing and the potential for sustainable aquaculture: a comprehensive review. J Appl Phycol. 2024;37:1–21.
-
Amin SA, Parker MS, Armbrust EV. Interactions between diatoms and bacteria. Microbiol Mol Biol Rev. 2012;76:667–84.
-
Andersen-Ranberg J, Kongstad KT, Nielsen MT, Jensen NB, Pateraki I, Bach SS, Hamberger B, Zerbe P, Staerk D, Bohlmann J. Expanding the landscape of diterpene structural diversity through stereochemically controlled combinatorial biosynthesis. Angew Chem Int Ed. 2016;55:2142–6.
-
Arshad S, Qadir ML, Hussain N, Ali Q, Han S, Ali D. Advances in CRISPR/Cas9 technology: shaping the future of photosynthetic microorganisms for biofuel production. Funct Plant Biol. 2025;52:FP24255.
-
Athanasakoglou A, Grypioti E, Michailidou S, Ignea C, Makris AM, Kalantidis K, Massé G, Argiriou A, Verret F, Kampranis SC. Isoprenoid biosynthesis in the diatom Haslea ostrearia. New Phytol. 2019;222:230–43.
-
Athanasakoglou A, Kampranis SC. Diatom isoprenoids: advances and biotechnological potential. Biotechnol Adv. 2019;37: 107417.
-
Avila C. Terpenoids in marine heterobranch molluscs. Mar Drugs. 2020;18:162.
-
Bommareddy A, Brozena S, Steigerwalt J, Landis T, Hughes S, Mabry E, Knopp A, VanWert AL, Dwivedi C. Medicinal properties of alpha-santalol, a naturally occurring constituent of sandalwood oil: review. Nat Prod Res. 2019;33:527–43.
-
Botella-Pavía P, Besumbes Ó, Phillips MA, Carretero-Paulet L, Boronat A, Rodríguez-Concepción M. Regulation of carotenoid biosynthesis in plants: evidence for a key role of hydroxymethylbutenyl diphosphate reductase in controlling the supply of plastidial isoprenoid precursors. Plant J. 2004;40:188–99.
-
Breitmaier E. Terpenes: flavors, fragrances, pharmaca, pheromones. John Wiley & Sons; 2006.
-
Brunson JK, McKinnie SMK, Chekan JR, McCrow JP, Miles ZD, Bertrand EM, Bielinski VA, Luhavaya H, Oborník M, Smith GJ. Biosynthesis of the neurotoxin domoic acid in a bloom-forming diatom. Science. 2018;361:1356–8.
-
Buch P, Airao V, Sharma T, Parmar S. Monoterpenoids: a neuropharmacological approach. Am J Pharm Health Res. 2020;8:7–27.
-
Chang W-C, Song H, Liu H-W, Liu P. Current development in isoprenoid precursor biosynthesis and regulation. Curr Opin Chem Biol. 2013;17:571–9.
-
Chatzivasileiou AO, Ward V, Edgar SM, Stephanopoulos G. Two-step pathway for isoprenoid synthesis. Proc Natl Acad Sci. 2019;116:506–11.
-
Chaves JE, Melis A. Biotechnology of cyanobacterial isoprene production. Appl Microbiol Biotechnol. 2018;102:6451–8.
-
Chen CY, Kao AL, Tsai ZC, Shen YM, Kao PH, Ng IS, Chang JS. Expression of synthetic phytoene synthase gene to enhance β-carotene production in Scenedesmus sp. CPC2. Biotechnol J. 2017;12: 1700204.
-
Chen J, Huang Y, Shu Y. Recent progress on systems and synthetic biology of diatoms for improving algal productivity. Front Bioeng Biotechnol. 2022;10: 908804.
-
Chen Q, Li J, Ma Y, Yuan W, Zhang P, Wang G. Occurrence and biosynthesis of plant sesterterpenes (C25), a new addition to terpene diversity. Plant Commun. 2021;2: 100184.
-
Christensen U, Vazquez-Albacete D, Søgaard K, Hobel T, Nielsen M, Harrison S, Hansen A, Møller B, Seppälä S, Nørholm M. De-bugging and maximizing plant cytochrome P450 production in Escherichia coli with C-terminal GFP fusions. Appl Microbiol Biotechnol. 2017;101:4103–13.
-
Chu LL. CRISPR-Cas system in microbial hosts for terpenoid production. Crit Rev Biotechnol. 2022;42:1116–33.
-
Commault AS, Kuzhiumparambil U, Herdean A, Fabris M, Jaramillo-Madrid AC, Abbriano RM, Ralph PJ, Pernice M. Methyl jasmonate and methyl-β-cyclodextrin individually boost triterpenoid biosynthesis in Chlamydomonas Reinhardtii UVM4. Pharmaceuticals. 2021;14:125.
-
Cordoba E, Salmi M, León P. Unravelling the regulatory mechanisms that modulate the MEP pathway in higher plants. J Exp Bot. 2009;60:2933–43.
-
Cvejić JH, Rohmer M. CO2 as main carbon source for isoprenoid biosynthesis via the mevalonate-independent methylerythritol 4-phosphate route in the marine diatoms Phaeodactylum tricornutum and Nitzschia ovalis. Phytochemistry. 2000;53:21–8.
-
D’Adamo S, Schiano G, di Visconte G, Lowe J-N, Beacham T, Landels A, Allen MJ, Spicer A, Matthijs M. Engineering the unicellular alga Phaeodactylum tricornutum for high-value plant triterpenoid production. Plant Biotechnol J. 2019;17:75–87.
-
Daboussi F, Leduc S, Maréchal A, Dubois G, Guyot V, Perez-Michaut C, Amato A, Falciatore A, Juillerat A, Beurdeley M. Genome engineering empowers the diatom Phaeodactylum tricornutum for biotechnology. Nat Commun. 2014;5:3831.
-
Daletos G, Katsimpouras C, Stephanopoulos G. Novel strategies and platforms for industrial isoprenoid engineering. Trends Biotechnol. 2020;38:811–22.
-
Davies FK, Jinkerson RE, Posewitz MC. Toward a photosynthetic microbial platform for terpenoid engineering. Photosynth Res. 2015;123:265–84.
-
Degenhardt J, Köllner TG, Gershenzon J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry. 2009;70:1621–37.
-
Dhokane D, Shaikh A, Yadav A, Giri N, Bandyopadhyay A, Dasgupta S, Bhadra B. CRISPR-based bioengineering in microalgae for production of industrially important biomolecules. Front Bioeng Biotechnol. 2023;11:1267826.
-
Eilers U, Bikoulis A, Breitenbach J, Büchel C, Sandmann G. Limitations in the biosynthesis of fucoxanthin as targets for genetic engineering in Phaeodactylum tricornutum. J Appl Phycol. 2016;28:123–9.
-
Einhaus A, Baier T, Kruse O. Molecular design of microalgae as sustainable cell factories. Trends Biotechnol. 2024;42:728–38.
-
Einhaus A, Baier T, Rosenstengel M, Freudenberg RA, Kruse O. Rational promoter engineering enables robust terpene production in microalgae. ACS Synth Biol. 2021;10:847–56.
-
Einhaus A, Steube J, Freudenberg RA, Barczyk J, Baier T, Kruse O. Engineering a powerful green cell factory for robust photoautotrophic diterpenoid production. Metab Eng. 2022;73:82–90.
-
Fabris M, George J, Kuzhiumparambil U, Lawson CA, Jaramillo-Madrid AC, Abbriano RM, Vickers CE, Ralph P. Extrachromosomal genetic engineering of the marine diatom Phaeodactylum tricornutum enables the heterologous production of monoterpenoids. ACS Synth Biol. 2020;9:598–612.
-
Fabris M, Matthijs M, Carbonelle S, Moses T, Pollier J, Dasseville R, Baart GJE, Vyverman W, Goossens A. Tracking the sterol biosynthesis pathway of the diatom P. haeodactylum tricornutum. New Phytol. 2014;204:521–35.
-
Fan M, Yuan S, Li L, Zheng J, Zhao D, Wang C, Wang H, Liu X, Liu J. Application of terpenoid compounds in food and pharmaceutical products. Fermentation. 2023;9:119.
-
Fan Y, Ding X-T, Wang L-J, Jiang E-Y, Van PN, Li F-L. Rapid sorting of fucoxanthin-producing Phaeodactylum tricornutum mutants by flow cytometry. Mar Drugs. 2021;19:228.
-
Ferriols VM, Emmanuel N, Yaginuma-Suzuki R, Fukunaga K, Kadono T, Adachi M, Matsunaga S, Okada S. An exception among diatoms: unique organization of genes involved in isoprenoid biosynthesis in Rhizosolenia setigera CCMP 1694. Plant J. 2017;92:822–33.
-
Gaj T, Gersbach CA, Barbas CF. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013;31:397–405.
-
Gallo C, Landi S, d’Ippolito G, Nuzzo G, Manzo E, Sardo A, Fontana A. Diatoms synthesize sterols by inclusion of animal and fungal genes in the plant pathway. Sci Rep. 2020;10:1–13.
-
Gao F, Sun Z, Kong F, Xiao J. Artemisinin-derived hybrids and their anticancer activity. Eur J Med Chem. 2020;188: 112044.
-
Garcia R, Alves ESS, Santos MP, Aquije GMF, Alberto A, Fernandes R, dos Santos RB, Ventura JA, Fernandes P. Antimicrobial activity and potential use of monoterpenes as tropical fruits preservatives. Braz J Microbiol. 2008;39:163–8.
-
Ge D, Xue Y, Ma Y. Two unexpected promiscuous activities of the iron–sulfur protein IspH in production of isoprene and isoamylene. Microb Cell Fact. 2016;15:1–11.
-
Genser B, Silbernagel G, De Backer G, Bruckert E, Rafael Carmena M, Chapman J, Deanfield J, Descamps OS, Rietzschel ER, Dias KC. Plant sterols and cardiovascular disease: a systematic review and meta-analysis. Eur Heart J. 2012;33:444–51.
-
Ghasemi Y, Mohagheghzadeh A, Moshavash M, Ostovan Z, Rasoul-Amini S, Morowvat MH, Ghoshoon MB, Raee MJ, Mosavi-Azam SB. Biotransformation of monoterpenes by Oocystis pusilla. World J Microbiol Biotechnol. 2009;25:1301–4.
-
Grauvogel C, Petersen J. Isoprenoid biosynthesis authenticates the classification of the green alga Mesostigma viride as an ancient streptophyte. Gene. 2007;396:125–33.
-
Gutiérrez MS, Campusano S, González AM, Gómez M, Barahona S, Sepúlveda D, Espenshade PJ, Fernández-Lobato M, Baeza M, Cifuentes V. Sterol regulatory element-binding protein (Sre1) promotes the synthesis of carotenoids and sterols in Xanthophyllomyces dendrorhous. Front Microbiol. 2019;10:586.
-
Gutiérrez S, Overmans S, Wellman GB, Lauersen KJ. Compartmentalized sesquiterpenoid biosynthesis and functionalization in the Chlamydomonas reinhardtii plastid. ChemBioChem. 2025;26: e202400902.
-
Gutiérrez S, Overmans S, Wellman G, Lauersen KJ. Pathway engineering and synthetic transgene design to produce diverse fragrant sesquiterpenoids from the green microalga Chlamydomonas reinhardtii; 2023. KAUST Res Repository. https://doi.org/10.25781/KAUST-F804N
-
Henry LK, Gutensohn M, Thomas ST, Noel JP, Dudareva N. Orthologs of the archaeal isopentenyl phosphate kinase regulate terpenoid production in plants. Proc Natl Acad Sci. 2015;112:10050–5.
-
Henry LK, Thomas ST, Widhalm JR, Lynch JH, Davis TC, Kessler SA, Bohlmann J, Noel JP, Dudareva N. Contribution of isopentenyl phosphate to plant terpenoid metabolism. Nature Plants. 2018;4:721–9.
-
Hu J, Wang D, Chen H, Wang Q. Advances in genetic engineering in improving photosynthesis and microalgal productivity. Int J Mol Sci. 2023;24:1898.
-
Huang PW, Wang LR, Geng SS, Ye C, Sun XM, Huang H. Strategies for enhancing terpenoids accumulation in microalgae. Appl Microbiol Biotechnol. 2021;105:1–12.
-
Jäpelt RB, Jakobsen J. Vitamin D in plants: a review of occurrence, analysis, and biosynthesis. Front Plant Sci. 2013;4:136.
-
Jaramillo-Madrid AC, Abbriano R, Ashworth J, Fabris M, Pernice M, Ralph PJ. Overexpression of key sterol pathway enzymes in two model marine diatoms alters sterol profiles in Phaeodactylum tricornutum. Pharmaceuticals. 2020;13:481.
-
Jeon MS, Han S-I, Jeon M, Choi Y-E. Enhancement of phycoerythrin productivity in Porphyridium purpureum using the clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9 ribonucleoprotein system. Biores Technol. 2021;330: 124974.
-
Jiang Z, Kempinski C, Chappell J. Extraction and analysis of terpenes/terpenoids. Curr Protoc Plant Biol. 2016;1:345–58.
-
Kajikawa M, Kinohira S, Ando A, Shimoyama M, Kato M, Fukuzawa H. Accumulation of squalene in a microalga Chlamydomonas reinhardtii by genetic modification of squalene synthase and squalene epoxidase genes. PLoS ONE. 2015;10: e0120446.
-
Kang NK, Baek K, Koh HG, Atkinson CA, Ort DR, Jin Y-S. Microalgal metabolic engineering strategies for the production of fuels and chemicals. Biores Technol. 2022;345: 126529.
-
Kant G, Pandey A, Shekhar H, Srivastava S. Enhanced bio-synthesis of isoprene via modifying mevalonate and methylerythritol phosphate pathways for industrial application: a review. Process Biochem. 2023;130:256–71.
-
Keeling PJ, Burki F, Wilcox HM, Allam B, Allen EE, Amaral-Zettler LA, Virginia Armbrust E, Archibald JM, Bharti AK, Bell CJ. The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): illuminating the functional diversity of eukaryotic life in the oceans through transcriptome sequencing. PLoS Biol. 2014;12: e1001889.
-
Kemper K, Hirte M, Reinbold M, Fuchs M, Brück T. Opportunities and challenges for the sustainable production of structurally complex diterpenoids in recombinant microbial systems. Beilstein J Org Chem. 2017;13:845–54.
-
Keramati A, Shariati FP, Tavakoli O, Akbari Z, Rezaei M. The effect of audible sound frequency on the growth and beta-carotene production of Dunaliella salina. S Afr J Bot. 2021;141:373–82.
-
Khanum F, Giwa A, Nour M, Al-Zuhair S, Taher H. Improving the economic feasibility of biodiesel production from microalgal biomass via high-value products coproduction. Int J Energy Res. 2020;44:11453–72.
-
Kim Y-J, Lee OR, Ji Yeon Oh, Jang M-G, Yang D-C. Functional analysis of 3-hydroxy-3-methylglutaryl coenzyme a reductase encoding genes in triterpene saponin-producing ginseng. Plant Physiol. 2014;165:373–87.
-
Kirby J, Keasling JD. Biosynthesis of plant isoprenoids: perspectives for microbial engineering. Annu Rev Plant Biol. 2009;60:335–55.
-
Kirby J, Nishimoto M, Chow RWN, Baidoo EEK, Wang G, Martin J, Schackwitz W, Chan R, Fortman JL, Keasling JD. Enhancing terpene yield from sugars via novel routes to 1-deoxy-d-xylulose 5-phosphate. Appl Environ Microbiol. 2015;81:130–8.
-
Köksal M, Jin Y, Coates RM, Croteau R, Christianson DW. Taxadiene synthase structure and evolution of modular architecture in terpene biosynthesis. Nature. 2011;469:116–20.
-
Kuczynska P, Jemiola-Rzeminska M, Strzalka K. Photosynthetic pigments in diatoms. Mar Drugs. 2015;13:5847–81.
-
Kumar A, Perrine Z, Stroff C, Postier BL, Coury DA, Sayre RT, Allnutt FCT. Molecular tools for bioengineering eukaryotic microalgae. Curr Biotechnol. 2016;5:93–108.
-
Kumar G, Shekh A, Jakhu S, Sharma Y, Kapoor R, Sharma TR. Bioengineering of microalgae: recent advances, perspectives, and regulatory challenges for industrial application. Front Bioeng Biotechnol. 2020;8:914.
-
Lauersen KJ, Baier T, Wichmann J, Wördenweber R, Mussgnug JH, Hübner W, Huser T, Kruse O. Efficient phototrophic production of a high-value sesquiterpenoid from the eukaryotic microalga Chlamydomonas reinhardtii. Metab Eng. 2016;38:331–43.
-
Lauersen KJ, Wichmann J, Baier T, Kampranis SC, Pateraki I, Møller BL, Kruse O. Phototrophic production of heterologous diterpenoids and a hydroxy-functionalized derivative from Chlamydomonas reinhardtii. Metab Eng. 2018;49:116–27.
-
Lee TM, LinJY TTH, Yang RY, Ng IS. Clustered regularly interspaced short palindromic repeats (CRISPR) technology and genetic engineering strategies for microalgae towards carbon neutrality: a critical review. Bioresour Technol. 2023;368: 128350.
-
Leyland B, Novichkova E, Dolui AK, Jallet D, Daboussi F, Legeret B, Li Z, Li-Beisson Y, Boussiba S, Khozin-Goldberg I. Acyl-CoA binding protein is required for lipid droplet degradation in the diatom Phaeodactylum tricornutum. Plant Physiol. 2024;194:958–81.
-
Li K, Gustafson KR. Sesterterpenoids: chemistry, biology, and biosynthesis. Nat Prod Rep. 2021;38:1251–81.
-
Li M, Hou F, Tong Wu, Jiang X, Li F, Liu H, Xian Mo, Zhang H. Recent advances of metabolic engineering strategies in natural isoprenoid production using cell factories. Nat Prod Rep. 2020;37:80–99.
-
Li S, Chang H, Zhang S, Ho S-H. Production of sustainable biofuels from microalgae with CO2 bio-sequestration and life cycle assessment. Environ Res. 2023;227: 115730.
-
Li X, Lan C, Li X, Hu Z, Jia B. A review on design-build-test-learn cycle to potentiate progress in isoprenoid engineering of photosynthetic microalgae. Biores Technol. 2022;363: 127981.
-
Lian J, Si T, Nair NU, Zhao H. Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains. Metab Eng. 2014;24:139–49.
-
Liao P, Hemmerlin A, Bach TJ, Chye M-L. The potential of the mevalonate pathway for enhanced isoprenoid production. Biotechnol Adv. 2016;34:697–713.
-
Lin B, Cui Y, Yan M, Wang Y, Gao Z, Meng C, Qin S. Construction of astaxanthin metabolic pathway in the green microalga Dunaliella viridis. Algal Res. 2019;44: 101697.
-
Liu J, Sun Z, Gerken H, Huang J, Jiang Y, Chen F. Genetic engineering of the green alga Chlorella zofingiensis: a modified norflurazon-resistant phytoene desaturase gene as a dominant selectable marker. Appl Microbiol Biotechnol. 2014;98:5069–79.
-
Lohr M, Schwender J, Polle JEW. Isoprenoid biosynthesis in eukaryotic phototrophs: a spotlight on algae. Plant Sci. 2012;185:9–22.
-
Lu L, Shen X, Sun X, Yan Y, Wang J, Yuan Q. CRISPR-based metabolic engineering in non-model microorganisms. Curr Opin Biotechnol. 2022;75: 102698.
-
Lu Y, Tarkowská D, Turečková V, Luo T, Xin Yi, Li J, Wang Q, Jiao N, Strnad M, Jian Xu. Antagonistic roles of abscisic acid and cytokinin during response to nitrogen depletion in oleaginous microalga Nannochloropsis oceanica expand the evolutionary breadth of phytohormone function. Plant J. 2014;80:52–68.
-
Lund S, Hall R, Williams GJ. An artificial pathway for isoprenoid biosynthesis decoupled from native hemiterpene metabolism. ACS Synth Biol. 2019;8:232–8.
-
Luo Z, Liu N, Lazar Z, Chatzivasileiou A, Ward V, Chen J, Zhou J, Stephanopoulos G. Enhancing isoprenoid synthesis in Yarrowia lipolytica by expressing the isopentenol utilization pathway and modulating intracellular hydrophobicity. Metab Eng. 2020;61:344–51.
-
Lupette J, Jaussaud A, Seddiki K, Morabito C, Brugière S, Schaller H, Kuntz M, Putaux J-L, Jouneau P-H, Rébeillé F. The architecture of lipid droplets in the diatom Phaeodactylum tricornutum. Algal Res. 2019;38: 101415.
-
Manfellotto F, Stella GR, Falciatore A, Brunet C, Ferrante MI. Engineering the unicellular alga Phaeodactylum tricornutum for enhancing carotenoid production. Antioxidants. 2020;9:757.
-
Martin LJ. Fucoxanthin and its metabolite fucoxanthinol in cancer prevention and treatment. Mar Drugs. 2015;13:4784–98.
-
Massé G, Belt ST, Rowland SJ, Rohmer M. Isoprenoid biosynthesis in the diatoms Rhizosolenia setigera (Brightwell) and Haslea ostrearia (Simonsen). Proc Natl Acad Sci. 2004;101:4413–8.
-
Meadows AL, Hawkins KM, Tsegaye Y, Antipov E, Kim Y, Raetz L, Dahl RH, Tai A, Mahatdejkul-Meadows T, Lan Xu. Rewriting yeast central carbon metabolism for industrial isoprenoid production. Nature. 2016;537:694–7.
-
Mohamadnia S, Tavakoli O, Faramarzi MA. Enhancing production of fucoxanthin by the optimization of culture media of the microalga Tisochrysis lutea. Aquaculture. 2021;533: 736074.
-
Mohamadnia S, Tavakoli O, Faramarzi MA, Shamsollahi Z. Production of fucoxanthin by the microalga Tisochrysis lutea: a review of recent developments. Aquaculture. 2020;516: 734637.
-
Morikawa T, Uraguchi Y, Sanda S, Nakagawa S, Sawayama S. Overexpression of DnaJ-like chaperone enhances carotenoid synthesis in Chlamydomonas reinhardtii. Appl Biochem Biotechnol. 2018;184:80–91.
-
Mund NK, Liu Y, Chen S. Advances in metabolic engineering of cyanobacteria for production of biofuels. Fuel. 2022;322: 124117.
-
Naik PS, Chanemougasoundharam A, Paul Khurana SM, Kalloo G. Genetic manipulation of carotenoid pathway in higher plants. Curr Sci. 2003;85:1423–30.
-
Narang PK, Dey J, Mahapatra SR, Roy R, Kushwaha GS, Misra N, Suar M, Raina V. Genome-based identification and comparative analysis of enzymes for carotenoid biosynthesis in microalgae. World J Microbiol Biotechnol. 2022;38:1–22.
-
Niehaus TD, Okada S, Devarenne TP, Watt DS, Sviripa V, Chappell J. Identification of unique mechanisms for triterpene biosynthesis in Botryococcus braunii. Proc Natl Acad Sci. 2011;108:12260–5.
-
Okada S, Devarenne TP, Murakami M, Abe H, Chappell J. ‘Characterization of botryococcene synthase enzyme activity, a squalene synthase-like activity from the green microalga Botryococcus braunii, Race B. Arch Biochem Biophys. 2004;422:110–8.
-
Onn SM, Koh GJ, Yap WH, Teoh ML, Low CF, Goh BH. Recent advances in genetic engineering of microalgae: bioengineering strategies, regulatory challenges and future perspectives. J Appl Phycol. 2024;37:1–18.
-
Oslan SN, Hazwani NF, Shoparwe AH, Yusoff AA, Rahim CS, Chang JS, Tan SN, Oslan KA, Ariff AB, Sulaiman AZ. A Review on Haematococcus pluvialis bioprocess optimization of green and red stage culture conditions for the production of natural astaxanthin. Biomolecules. 2021;11:256.
-
Papaefthimiou D, Diretto G, Demurtas OC, Mini P, Ferrante P, Giuliano G, Kanellis AK. Heterologous production of labdane-type diterpenes in the green alga Chlamydomonas reinhardtii. Phytochemistry. 2019;167: 112082.
-
Pateraki I, Andersen-Ranberg J, Jensen NB, Wubshet SG, Heskes AM, Forman V, Hallström B, Hamberger B, Motawia MS, Olsen CE. Total biosynthesis of the cyclic AMP booster forskolin from Coleus forskohlii. Elife. 2017;6: e23001.
-
Perozeni F, Cazzaniga S, Baier T, Zanoni F, Zoccatelli G, Lauersen KJ, Wobbe L, Ballottari M. Turning a green alga red: engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. Plant Biotechnol J. 2020;18:2053–67.
-
Polzin J, Rorrer GL. Selective production of the acyclic monoterpene β-myrcene by microplantlet suspension cultures of the macrophytic marine red alga Ochtodes secundiramea under nutrient perfusion cultivation with bromide-free medium. Algal Res. 2018;36:159–66.
-
Rampen SW, Abbas BA, Schouten S, Damste JSS. A comprehensive study of sterols in marine diatoms (Bacillariophyta): implications for their use as tracers for diatom productivity. Limnol Oceanogr. 2010;55:91–105.
-
Rowland SJ, Belt ST, Wraige EJ, Massé G, Roussakis C, Robert J-M. Effects of temperature on polyunsaturation in cytostatic lipids of Haslea ostrearia. Phytochemistry. 2001;56:597–602.
-
Sathasivam R, Ki J-S. A review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Mar Drugs. 2018;16:26.
-
Savakis P, Hellingwerf KJ. Engineering cyanobacteria for direct biofuel production from CO2. Curr Opin Biotechnol. 2015;33:8–14.
-
Schwender J, Gemünden C, Lichtenthaler HK. Chlorophyta exclusively use the 1-deoxyxylulose 5-phosphate/2-C-methylerythritol 4-phosphate pathway for the biosynthesis of isoprenoids. Planta. 2001;212:416–23.
-
Shen Y, Liang W-J, Shi Y-N, Kennelly EJ, Zhao D-K. Structural diversity, bioactivities, and biosynthesis of natural diterpenoid alkaloids. Nat Prod Rep. 2020;37:763–96.
-
Shin B-K, Kim M, Han J. Exceptionally high percentage of IPP synthesis by Ginkgo biloba IspH is mainly due to Phe residue in the active site. Phytochemistry. 2017;136:9–14.
-
Singh AK, Srivastava RK, Pal P, Mandal S, Sahoo UK, Prakash A, Sridhar K, Sharma M, Sarangi PK, Inbaraj BS. Microalgal biorefinery as a sustainable and cost-effective platform for co-production of high-value-added products/metabolites: an insight into emerging trends, challenges, and opportunities. Biocatal Agric Biotechnol. 2024;58: 103192.
-
Song J, Zhao H, Zhang L, Li Z, Han J, Zhou C, Jilin Xu, Li X, Yan X. The heat shock transcription factor PtHSF1 mediates triacylglycerol and fucoxanthin synthesis by regulating the expression of GPAT3 and DXS in Phaeodactylum tricornutum. Plant Cell Physiol. 2023;64:622–36.
-
Sproles AE, Fields FJ, Smalley TN, Le CH, Badary A, Mayfield SP. Recent advancements in the genetic engineering of microalgae. Algal Res. 2021;53: 102158.
-
Stavridou E, Karapetsi L, Nteve GM, Tsintzou G, Chatzikonstantinou M, Tsaousi M, Martinez A, Flores P, Merino M, Dobrovic L. Landscape of microalgae omics and metabolic engineering research for strain improvement: an overview. Aquaculture. 2024;587: 740803.
-
Stirk WA, van Staden J. Potential of phytohormones as a strategy to improve microalgae productivity for biotechnological applications. Biotechnol Adv. 2020;44: 107612.
-
Syrenne RD. Autotrophic production of limonene in engineered microalgae Chlamydomonas reinhardtii; 2016.
-
Tetali SD. Terpenes and isoprenoids: a wealth of compounds for global use. Planta. 2019;249:1–8.
-
Tran Q-G, Le TT, Choi DY, Cho DH, Yun J-H, Choi HI, Kim H-S, Lee YJ. Progress and challenges in CRISPR/Cas applications in microalgae. J Microbiol. 2025;63:e2501028–128.
-
Vavitsas K, Fabris M, Vickers CE. Terpenoid metabolic engineering in photosynthetic microorganisms. Genes. 2018;9:520.
-
Vickers CE, Williams TC, Peng B, Cherry J. Recent advances in synthetic biology for engineering isoprenoid production in yeast. Curr Opin Chem Biol. 2017;40:47–56.
-
Vieira KR, Pinheiro PN, Santos AB, Cichoski AJ, de Menezes CR, Wagner R, Zepka LQ, Jacob-Lopes E. The role of microalgae-based systems in the dynamics of odors compounds in the meat processing industry. Desalin Water Treat. 2019;150:282–92.
-
Volkman JK, Barrett SM, Dunstan GA. C25 and C30 highly branched isoprenoid alkenes in laboratory cultures of two marine diatoms. Org Geochem. 1994;21:407–14.
-
Vranová E, Coman D, Gruissem W. Structure and dynamics of the isoprenoid pathway network. Mol Plant. 2012;5:318–33.
-
Wang C, Zada B, Wei G, Kim S-W. Metabolic engineering and synthetic biology approaches driving isoprenoid production in Escherichia coli. Biores Technol. 2017;241:430–8.
-
Wang Xi, Baidoo EEK, Kakumanu R, Xie S, Mukhopadhyay A, Lee TS. Engineering isoprenoids production in metabolically versatile microbial host Pseudomonas putida. Biotechnol Biofuels Bioprod. 2022;15:137.
-
Weiss TL, Chun HJ, Okada S, Vitha S, Holzenburg A, Laane J, Devarenne TP. Raman spectroscopy analysis of botryococcene hydrocarbons from the green microalga Botryococcus braunii. J Biol Chem. 2010;285:32458–66.
-
Wichmann J, Baier T, Wentnagel E, Lauersen KJ, Kruse O. Tailored carbon partitioning for phototrophic production of (E)-α-bisabolene from the green microalga Chlamydomonas reinhardtii. Metab Eng. 2018;45:211–22.
-
Wichmann J, Eggert A, Elbourne LDH, Paulsen IT, Lauersen KJ, Kruse O. Farnesyl pyrophosphate compartmentalization in the green microalga Chlamydomonas reinhardtii during heterologous (E)-α-bisabolene production. Microb Cell Fact. 2022;21:190.
-
Widiyanto S, Sarto M, Fitria L, Yudo R, Suyono EA. Biochemical compounds and sub-chronic toxicity test of Chlorella sp. and Spirulina sp. isolated from Glagah Coastal Water. Berkala Penelitian Hayati J Biol Res. 2018;24:58–64.
-
Yan S, Tian-Peng Y, Jia-Peng W, Dong G, Zhang QY, Le C, Zhong-Tao D. Three new diterpenoid alkaloids isolated from Aconitum brevicalcaratum. Chin J Nat Med. 2018;16:866–70.
-
Yang J, Nie Q, Liu H, Xian Mo, Liu H. A novel MVA-mediated pathway for isoprene production in engineered E. coli. BMC Biotechnol. 2016;16:1–9.
-
Yang J, Nie Q, Ren M, Feng H, Jiang X, Zheng Y, Liu M, Zhang H, Xian Mo. Metabolic engineering of Escherichia coli for the biosynthesis of alpha-pinene. Biotechnol Biofuels. 2013;6:1–10.
-
Yang Q, Xie Z, Zheng X, Li K, Tao Lu, Yinghua Lu, Chen C, Ling X. Genetic regulation and fermentation strategy for squalene production in Schizochytrium sp. Appl Microbiol Biotechnol. 2022;106:2415–31.
-
Yang R, Zhu L, Li T, Zhu LY, Ye Z, Zhang D. Photosynthetic conversion of CO2 into pinene using engineered Synechococcus sp. PCC 7002. Front Bioeng Biotechnol. 2021;9:779437.
-
Yazdani M, Tavakoli O, Khoobi M, Wu YS, Faramarzi MA, Gholibegloo E, Farkhondeh S. Beta-carotene/cyclodextrin-based inclusion complex: improved loading, solubility, stability, and cytotoxicity. J Incl Phenomena Macrocycl Chem. 2021;102:1–10.
-
Yazdani M, Croen MG, Fish TL, Thannhauser TW, Ahner BA. Overexpression of native ORANGE (OR) and OR mutant protein in Chlamydomonas reinhardtii enhances carotenoid and ABA accumulation and increases resistance to abiotic stress. Metab Eng. 2021;68:94–105.
-
Ye Z-W, Jiang J-G, Guang-Hong Wu. Biosynthesis and regulation of carotenoids in Dunaliella: progresses and prospects. Biotechnol Adv. 2008;26:352–60.
-
Zhang W, Zhou W, Jiang Si, Wang Y, Chen L, Yang G, Liu T. Heterotrophic modification of Phaeodactylum tricornutum Bohlin. Algal Res. 2023;72: 103137.
-
Zhao M-L, Cai W-S, Zheng S-Q, Zhao J-L, Zhang J-L, Huang Y, Zhang-Li Hu, Jia B. Metabolic engineering of the isopentenol utilization pathway enhanced the production of terpenoids in Chlamydomonas reinhardtii. Mar Drugs. 2022;20:577.
-
Zhou F, Pichersky E. More is better: the diversity of terpene metabolism in plants. Curr Opin Plant Biol. 2020;55:1–10.
-
Zu Y, Prather KLJ, Stephanopoulos G. Metabolic engineering strategies to overcome precursor limitations in isoprenoid biosynthesis. Curr Opin Biotechnol. 2020;66:171–8.
Acknowledgements
Not applicable.
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mohamadnia, S., Valverde-Pérez, B., Tavakoli, O. et al. Progress and prospects in metabolic engineering approaches for isoprenoid biosynthesis in microalgae. Biotechnol. Biofuels Bioprod. 18, 64 (2025). https://doi.org/10.1186/s13068-025-02665-y
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1186/s13068-025-02665-y