Introduction
Immunoglobulin E (IgE) is one of the five classes of human immunoglobulins—IgM, IgG, IgD, IgA, and IgE—and plays an essential role in the immune defense, particularly against parasitic infections and in triggering allergic reactions such as atopic dermatitis and food allergies1. Measuring IgE levels is essential for diagnosing and managing allergic diseases. Elevated IgE levels are strong indicators of allergic conditions such as asthma, allergic rhinitis, and atopic dermatitis. Conversely, abnormally low IgE levels may suggest underlying immune deficiencies or other medical conditions2,3. By assessing IgE levels, healthcare providers can identify patients at risk for allergic diseases, monitor disease progression, and tailor treatment plans accordingly. Therefore, blood tests for IgE are valuable tools in diagnosis and management.
For serum IgE testing, two general types of blood tests are commonly used: (1) total IgE tests, which measure the overall concentration of IgE antibodies in the blood, and (2) specific IgE (sIgE) tests, which measure IgE antibodies produced in response to a particular allergen4. Several in vitro methods are available for detecting human IgE, including enzyme-linked immunosorbent assay (ELISA)5, radioallergosorbent test (RAST)6, and fluorescence enzyme immunoassay (FEIA)7. These assays rely on antibodies specific to human IgE, which are essential for achieving high specificity and sensitivity. However, traditional antibody production involves animal immunization and complex purification procedure, presenting ethical and technical challenges. As a result, alternative in vitro methods for antibody production are needed.
One such alternative involves the use of single-chain variable fragment (scFv) antibodies specific to anti-human IgE, which have been constructed and expression in Escherichia coli8. Although these scFvs exhibit high affinity and specificity, their production remains complex, requiring advanced antibody engineering. More recently, plant-based expression systems have been explored for producing anti-human IgE antibodies. While these plant-derived antibodies have shown IgE-binding activity, improvements in yield and purification are still necessary9.
Another promising approach for developing IgE-binding molecules involves the use of aptamers—short single-stranded oligonucleotides selected to bind specific targets. Aptamers have been used in various IgE detection platforms, including colorimetric assays, label-free techniques such as surface plasmon resonance, fluorescence assays, and electrochemical detection10. Despite their high affinity and selectivity, aptamers lack standardized protocols; a method optimized for one aptamer cannot be readily applied to others11.
Phage display technology offers a powerful strategy for generating specific binding molecules12. Through the genetic modification of bacteriophages, this technology enables the expression of diverse ligands—including antibody fragments, proteins, and peptides—on the phage surface13. Phage display has been widely used in applications such as cancer-targeted therapy14,15,16,17, vaccine development18,19,20, epitope mapping21,22, mimotope identifications23,24, molecular recognition25,26,27,28,29,30,31,32,33, and the detection of hazardous substances in foodstuff34,35. For example, human IgE-specific scFv phage libraries have been developed to detect hevein—a lectin-like protein and a major allergen found in the latex of rubber trees36. Two scFvs were combined to form a Fab fragment antibody capable of binding human IgE serum in a competitive assay for hevein detection36. In our previous study, we screened a phage-displayed human antibody fragment library against Salmonella spp. and successfully identified phages that specifically bound Salmonella Enteritidis. These phages could differentiate between live and dead cells, and we subsequently developed a rapid test using phage-antibodies as molecular binders29.
When comparing phage-displayed antibody fragments and peptides, antibody fragments are more susceptible to changes in pH, salt composition, and temperature, which can lead to denaturation and loss of structure37. In contrast, peptides derived from phage display can be chemically synthesized and are generally more stable under diverse conditions—a key advantage for point-of-care diagnostics. Additionally, synthetic peptides can be chemically modified to further improve their stability38.
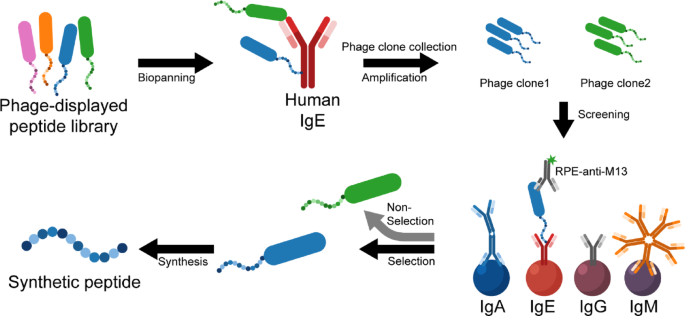
In our previous publications, we demonstrated the utility of phage-derived peptides as binders for detecting foodborne pathogens30,31. Furthermore, numerous studies have explored peptide-based strategies for immunotherapy39,40,41,42,43,44. Owing to their advantages—such as ease of synthesis, rapid development, high efficiency, strong stability, and cost-effectiveness45—phage-displayed peptides represent a robust platform for developing tools to detect human IgE. To the best of our knowledge, synthetic peptides have not yet been reported for binding to human IgE. Therefore, in this study, we aimed to identify peptides with specific affinity for human IgE using a phage-displayed 12-mer peptide library. Phages were biopanned against human IgE, and clones were screened using a bead array technique that enables simultaneous testing against different antibody isotypes. Clones that showed specific binding to IgE were selected for peptide synthesis and further characterization (Fig. 1). We hope that our findings will contribute to advancements in allergy diagnostics and therapeutics development.

Schematic of phage-displayed peptide screening. The phage library was biopanned against the human IgE antibody. Individual phage clones were collected and amplified before being screened using the antibody-coated bead array method. Each human antibody isotype was conjugated to MagPlex beads. Phage clones specific to IgE were selected, followed by DNA sequencing. The peptides identified from the sequencing data were synthesized and characterized.
Materials and methods
Chemicals and reagents
The Phage Display Peptide Library (Ph.D.-12 Dodecapeptide Library, #E8110S) was purchased from New England Biolabs. Native human IgE protein (IgE, #ab65866) and rabbit IgG anti-ovalbumin (Rabbit IgG, #ab306592) were purchased from Abcam. Human isotype controls for IgA (#31148), IgG (#31154), and IgM (#31146); a biotin-labeled monoclonal antibody specific to the anti-M13 phage coat protein (anti-M13-biotin, #MA5-30020), 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide hydrochloride (EDC, #22980), and N-hydroxysulfosuccinimide (sulfo-NHS, #24510); and biotin-labeled goat anti-human IgE (#A18797) were purchased from Thermo Fisher Scientific. R-phycoerythrin-conjugated streptavidin (SAPE, #S866) and CaptureSelect IgE affinity beads (#1TFS-BP-2943542005) were obtained from Invitrogen. Mouse IgG anti-mite Group 1 (Mouse IgG, MA-4C1-1) was purchased from Indoor Biotechnologies. Goat anti-mouse IgA + IgG + IgM (H + L) (Goat IgG #5210-0187) was purchased from KPL (SeraCare). Peptides conjugated with l-propargylglycine (Pra) were synthesized and purchased from GenScript, Singapore.
Suspension biopanning against native human IgE protein
Biopanning was performed following the instructions provided in the Ph.D.™ Phage Display Libraries kit manual, with some modifications. Each round of biopanning included four main steps: binding, washing, elution, and amplification.
Briefly, 200 µL of the Ph.D.-12 library (1011 pfu/mL) was mixed with 5 µg of native human IgE protein and incubated for 20 min at room temperature (RT). The mixture was then added to 25 µL of Capture Select IgE affinity beads and incubated for an additional 15 min. After incubation, unbound phages were removed by washing the beads ten times with Tris-buffered saline containing 0.1% Tween 20 (TBST; 50 mM Tris–HCl pH 7.5, 150 mM NaCl, and 0.1% Tween 20). Elution was performed by adding 1 mL of elution buffer (1 mg/mL bovine serum albumin [BSA, #A7906, Sigma-Aldrich] in 0.2 M glycine–HCl, pH 2.2), followed by a 10-min incubation. The eluted phages were neutralized with 1 M Tris–HCl, pH 9.1, and stored at 4 °C for subsequent rounds of biopanning.
For amplification, the eluted phages were added to 20 mL of an E. coli ER2738 culture (OD600 at 0.05) or to a 100-fold dilution of an overnight culture E. coli ER2738 (without a helper phage), and incubated at 37 °C with shaking at 250 rpm for 4.5 h. After centrifugation at 12,000×g for 10 min at 4 °C, the supernatant was collected and precipitated overnight at 4 °C using 1/6 volume of 20% polyethylene glycol (PEG) and 2.5 M NaCl solution. The precipitated phages were collected by centrifugation and resuspended in 200 µL of TBS. After the third round of biopanning, 208 individual phage clones were amplified in 1 mL of E. coli ER2738 in a 48-well plate. The collected supernatants were screened using a multiplex assay.
For phage DNA sequencing, clones that yielded positive results in the multiplex assay were selected. Each selected clone was amplified and purified as described above. Phage DNA was extracted and purified using iodide buffer and ethanol precipitation. DNA quality and concentration were assessed by 1.5% (w/v) agarose gel electrophoresis and OD260/280 measurement. Purified phage DNA samples were then submitted to First BASE Laboratories for Sanger sequencing. The corresponding 12-mer peptide sequences were translated from 36-nucleotide sequences using the ExPAsy translate tool. To determine potential matches with known target peptides, the sequences were analyzed using the UniProt peptide search tool (https://www.uniprot.org/peptide-search)46.
Antibody and peptide coupling MagPlex beads for a multiplex assay
Bead array technology utilizes various sets of microspheres coded with fluorescent dyes, each set containing a distinct ratio of red and infrared dyes47. This technique was employed to simultaneously detect and characterize phage clones. Antibody-coupled beads were used to screen phage clones, while peptide-coupled beads were used for the characterization of synthesized peptides.
For antibody-bead conjugation, four human antibody isotypes—IgA, IgE, IgG, and IgM—were conjugated to separate MagPlex bead sets using a carbodiimide reaction. Briefly, a bead set (1 × 106 beads) was activated with 10 µL of 50 mg/mL sulfo-NHS and 10 µL of 50 mg/mL EDC in activation buffer (0.1 M NaH2PO4, pH 6) for 20 min on a rotator at RT. The beads were then washed three times with activation buffer and incubated with 5 µg of antibody for 2 h on a rotator at RT. Unbound antibodies were removed by washing with phosphate-buffered saline pH 7.4 (PBS, pH 7.4) containing 0.05% Tween 20 (PBST; 1 mM KH2PO4, 0.15 mM Na2HPO4, 3 mM NaCl, and 0.05% Tween 20). The beads were then resuspended in 500 µL of PBST and stored at 4 °C until use. Prior to use, the antibody-conjugated beads were subjected to a blocking step.
For peptide-bead conjugation, the peptides derived from the phage display screening were synthesized with l-propargylglycine (Pra) at the C-terminus to enable potential conjugation via ‘click’ chemistry, while the N-terminus remained unmodified. To determine the most suitable bead conjugation method, Peptide 5 was used as a model and tested for conjugation to MagPlex beads using both carbodiimide chemistry (as used for antibody conjugation) and copper-catalyzed azide-alkyne cycloaddition (‘click’ chemistry).
For peptide-bead conjugation via carbodiimide chemistry, the amine group at the N-terminus of the synthetic peptide was coupled to carboxyl groups on the surface of MagPlex beads. This procedure followed the same protocol described above for antibody-bead conjugation.
For peptide-bead conjugation via ‘click’ chemistry, carboxyl groups on the surface of the beads were first coupled to the amine group of amine-PEG-azide using EDC. The l-propargylglycine moiety of the peptide was then conjugated to the azide-modified beads via copper-catalyzed azide-alkyne cycloaddition (‘click’ chemistry). The conjugation protocol was adapted, with modifications, from Coppock and Stratis-Cullum48. Briefly, 10 µL of beads (1.25 × 107 beads/mL) were washed with 250 µL of deionized water (DI) using a magnetic tube separator, followed by washing with 250 µL of 0.1 M MES buffer (pH 6.0). The beads were resuspended in 0.1 M MES buffer, then 15 µL of 100 mg/mL azido-PEG23-amine and 25 µL of 0.8 mg/mL EDC were added and incubated for 2 h. After incubation, the beads were washed twice with 250 µL of DI water. For the click reaction, a mixture containing 0.4 µL of 6.4 mg/mL peptide, 0.9 µL of “click” solution (prepared from 1.25 µL of 3.2 mg/mL CuSO₄, 2.5 µL of 22 mg/mL tris-hydroxypropyltriazolylmethylamine [THPTA]), 3.1 µL of 40 mg/mL sodium ascorbate, and 27 µL of DI water was added to the beads and incubated for 1 h. After the reaction, the mixture was removed, and 250 µL of 5% sodium diethyldithiocarbamate trihydrate was added to chelate any free residual copper, followed by a 2-min incubation. The beads were washed twice with 250 µL DI water, then once with 250 µL of protein-free PBST. The peptide-conjugated beads were resuspended in 250 µL of PBST and stored at 4 °C until use. A blocking step was performed prior to use.
Biotin–poly-l-lysine–peptide conjugation
Synthetic peptides were conjugated to biotin and used as detection molecules. Poly-l-lysine (PLL) served as a linker between biotin and peptides. The conjugation process involved three main steps: (1) conjugation of biotin to PLL via carbodiimide chemistry, (2) attachment of azido-PEG-amine using glutaraldehyde as a crosslinker, and (3) coupling of alkyne-functionalized peptides to the azide-modified biotin–PLL via click chemistry.
First, biotin was conjugated to PLL using the carbodiimide chemistry. Biotin (#14400, Sigma-Aldrich); 50 µL of 2.5 mg/mL in 25 mM MES, pH 5.0) was mixed with 20 µL of 50 mg/mL EDC and 20 µL of 50 mg/mL sulfo-NHS in a total volume of 400 µL of 25 mM MES, pH 5.0 to activate the carboxyl group of biotin. The mixture was incubated for 20 min at RT. Then, PLL (#P2658, Sigma-Aldrich; 100 µL of 1 mg/mL) was added and incubated for 2 h at RT. Excess biotin was removed using a 10 kDa molecular weight cut-off filter of (Amicon #UFC501096, Merck). The resulting biotin–conjugated PLL (biotin–PLL) was reconstituted in 100 µL of deionized water (DI). The remaining free amine groups were quantified using the 2,4,6-trinitrobenzene sulfonate (TNBS) assay. For peptide conjugation, azido-PEG23-amine was coupled to biotin–PLL using glutaraldehyde to crosslink the amine groups. Biotin–PLL from the previous step (50 µL) was mixed 4 µL of 25% glutaraldehyde (#G5882, Sigma-Aldrich) and 10 µL of 5 mg/mL azido-PEG23-amine (#QBD10525, Sigma-Aldrich) in a total volume of 500 µL of 0.1 M NaCl, 0.1 M NaHCO3 buffer, pH 8.5. The mixture was incubated on ice for 3 h. After incubation, excess azido-PEG23-amine was removed by filtration, and the solution was reconstituted in 50 µL of DI water. Peptides were conjugated to the biotin–PLL–azide using a click chemistry reaction. Briefly, peptide (5 µL of 5 mg/mL) was mixed with 25 µL of biotin–PLL–azide, 2 µL of click solution (prepared from 5 µL of 0.8 mg/mL CuSO4 and 10 µL of 5.5 mg/mL tris-hydroxypropyltriazolylmethylamine [THPTA]), and 6.2 µL of 10 mg/mL sodium ascorbate. The mixture was incubated at RT for 1 h in the dark. After incubation, the solution was purified by filtration, and reconstituted in 50 µL of PBS.
2,4,6-Trinitrobenzene sulfonate (TNBS) assay
To quantify free amino groups, 2,4,6-Trinitrobenzene Sulfonic Acid (TNBS) was performed49. Briefly, samples (50 µL) were prepared in PBS and mixed with 25 µL of 0.01% TNBS in 0.05 M NaHCO3 buffer (pH 9.6). The reaction mixture was incubated at 37 °C for 1 h. Absorbance was measured at 335 nm using a SpectraMax M5 microplate reader (Molecular Devices). The concentration of l-lysine on PLL was calculated from a standard curve generated using l-lysine.
Enzyme-linked immunosorbent assay (ELISA) method
To verify the presence of biotin after peptide conjugation, an ELISA was performed. Biotin–PLL–peptide conjugates (100 µL of 0.5 µg/mL) were prepared in 0.05 M NaHCO3 buffer (pH 9.6), and coated onto a microtiter plate (#3590, Corning) overnight at 4 °C. The plate was then washed three times with 300 µL/well of PBST using an automatic plate washer (Bio-Rad). After washing, each well was blocked with 300 µL of 2% (w/v) BSA in PBST at RT for 1 h. Following incubation, the wells were washed again before adding 100 µL of streptavidin–horseradish peroxidase (SA–HRP, #434323, Invitrogen) at a 1:2500 dilution. The plate was incubated at RT for 1 h. Subsequently, 100 µL of 3,3′,5,5′-Tetramethylbenzidine (TMB, #002023, Invitrogen) substrate solution was added to each well to develop the signal. After 3 min, the reaction was stopped by adding 50 µL/well of 0.5 M H2SO4. Absorbance was measured at 450 nm using a SpectraMax M5 microplate reader (Molecular Devices). Biotin–PLL and biotin-labeled goat anti-mouse antibodies were used as positive controls, while NaHCO3 buffer served as the negative control. Signals at least twice the background level were considered positive.
Screening of phage clones using a multiplex assay
To screen phage clones, a mixture of 50 µL of the antibody-coated beads (1 × 105 beads/mL for each bead region) was prepared in 1% (w/v) BSA in PBST and blocked for 1 h at RT. After incubation, the blocking solution was removed, and the beads were washed with PBST using an automatic magnetic microplate washer (Bio-Rad). Each individual phage clone (100 µL) was then added to the antibody-coated beads and incubated for 1 h at RT. The washing step was repeated before adding 100 µL of 2 µg/mL anti-M13-biotin antibody, followed by a 1-h incubation. Another washing step was performed to remove unbound components. Next, 100 µL of 2 µg/mL R-phycoerythrin-labeled streptavidin (SAPE) was added and incubated for 1 h. After a final wash, 100 µL of PBST was added to each well. Fluorescent signals were measured using a Luminex MagPix system. PBST and biotin-labeled goat anti-human IgE (0.5 µg/mL) were used as the negative and positive controls, respectively. Fluorescent signals greater than three times that of the negative control were considered as a positive result50.
Specificity test of synthetic peptides
To evaluate the specificity of the selected synthetic peptides, two multiplex assay formats—direct and sandwich—were performed. The direct multiplex system was similar to the phage clone screening method, in which peptides bound to antibody-coated bead; however, biotin–PLL–peptides were used instead of whole phages. In the sandwich multiplex system, synthetic peptides were used as capture molecules, and biotin-labeled goat anti-human IgE served as the detection reagent.
For the direct multiplex assay, the procedure followed the method described in the ‘Screening of phage clones using multiplex assay’ section, except that biotin–PLL–peptides (50 µL of 1.25 µg/mL) and 2 µg/mL of SAPE were used instead of phages and biotin-labeled anti-M13 antibody.
For the sandwich multiplex assay, a mixture of 50 µL of the peptide-coated beads (1 × 105 beads/mL of each bead region) and 50 µL of 2 µg/mL sample (human antibody isotypes or antibodies from other species such as rabbit, mouse, goat) was incubated for 1 h at RT. Unbound components were removed by washing with PBST before adding 100 µL of 4 µg/mL biotin-labeled goat anti-human IgE. After a 1-h incubation at RT, the beads were washed again, followed by the addition of 100 µL of 2 µg/mL SAPE and a further 1-h incubation. After a final wash, 100 µL of PBST was added to each well. Fluorescent signals were measured using a Luminex MagPix system.
Sensitivity test and limit of detection of synthetic peptides
To evaluate the sensitivity of the synthetic peptides, five concentrations of human IgE (two-fold dilutions ranging from 0.15 to 2.5 µg/mL) were tested using the multiplex assay method. A linear calibration curve was generated by plotting the normalized signal-to-background values against the corresponding human IgE concentrations. The limit of detection (LOD) was calculated from the linear regression equation, using a normalized signal-to-background value of 3-representing three times of background signal51,52.
Results and discussions
Screening of phage-displayed peptide against human IgE
To select phage clones specific to IgE, clones that bound to IgE but not to other isotypes were used as the selection criterion. After the third round of biopanning, a total of 208 individual phage clones were randomly picked and screened against four human immunoglobulin isotypes using the antibody-coated bead array method (Fig. 2A). Unfortunately, only one phage-peptide clone was found to be specific for human IgE.

Heat map of phage clone screening using an antibody-bead array method. (A) A total of 208 individual phage clones (rows) were tested against human IgA, IgE, IgG, and IgM (columns). The fluorescent intensity values were categorized into four levels: Negative (0–50), Low (50.1–250), Medium (250.1–500), and High (> 500.1). An assay buffer (PBST) and a biotin-labeled goat-anti-human IgE were used as negative and positive controls, respectively. (B) Sixteen phage clones were selected and sequenced for DNA analysis. (C) Peptide sequences were decoded from DNA sequence analysis. (D) The peptides were synthesized with l-propargylglycine (Pra) at the C-terminus. The heat map was created using GraphPad Prism 10, Version 10.5.0 (774).
Surprisingly, most phage clones exhibited reactivity toward human IgG, even though human IgG was not included during the biopanning step. The human IgE used in this study was produced from a monoclonal hybridoma with a kappa light chain. In contrast, the human IgG used in the screening was a polyclonal antibody, which may contain both kappa and lambda light chains. The observed similarity between IgE and IgG might be due to their shared light chains, which are encoded by the IGLC (constant) gene, while the variable domain is encoded by two types of genes, IGLV (variable) and IGLJ (joining)53. To confirm the phage’s ability to bind to IgG, one clone was selected for further analysis.
Since only one clone was initially found to be specific to IgE, additional selection criteria were applied to improve the likelihood of identifying IgE-binding phage clones. Clones exhibiting strong binding to IgE and minimal cross-reactivity with other isotypes were selected. Based on these criteria, fourteen additional clones were chosen. Thus, a total of sixteen clones—one clone that bound only to IgE (Clone #1), fourteen that bound strongly to IgE with low cross-reactivity (Clone #2–13 and #15–16), and one that bound to IgG (Clone #14)—were amplified, purified, and tested using the multiplex assay. The results showed that Clone #1 remained highly reactive to IgE (Fig. 2B). Six clones—#1, #2, #5, #9, #10, and #13—produced high signal intensities specific to the human IgE antibody (Fig. 2B). However, among the fourteen clones selected for strong IgE binding with low cross-reactivity, not all yielded results consistent with the initial screening. These inconsistencies may have results from mutations introduced during the amplification process54. The clone that bound to IgG remained consistent with the initial screening results.
To identify the peptides of all sixteen clones, the DNA was sequenced (Table S1), and the corresponding 12-mer peptides were translated from the 36-nucleotide inserts (Fig. 2C). Among the sixteen clones, thirteen unique peptide sequences were identified. Two peptide sequences—“NTSRLPNYELLH” and “LHAANSSRNLGA”—were found in multiple clones. The peptide “NTSRLPNYELLH” appeared in three clones: Clone #1, which showed high binding to IgE, and Clones #3 and #4, which exhibited low binding to both IgE and IgG. This redundancy might be due to insufficient control over the number of phages during the screening process. The specificity of the peptide “NTSRLPNYELLH” requires further investigation. Although the sequence “LHAANSSRNLGA” was present in two clones, it did not yield a detectable signal for IgE binding and was excluded from further analysis.
Based on the multiplex assay results and peptide sequences, seven peptides showing high signal intensity—six peptides for IgE and one peptide for IgG—were selected for further characterization. To assess the novelty of these peptides, a BLAST search was performed against the UniProt peptide database (https://www.uniprot.org/peptide-search)46. The absence of significant matches suggests that these sequences are potentially novel IgE- or IgG-binding peptides. Since this study focused on IgE, six peptides were synthesized with a GGGS spacer and l-propargylglycine (Pra) at the C-terminus to enable site-specific conjugation (Fig. 2D).
Specificity of synthetic peptides
Two detection systems, direct and sandwich multiplex assays, were employed to evaluate the specificity of the synthetic peptides.
In the direct multiplex assay, four human antibody isotypes were immobilized on beads, and biotin–PLL–peptides were tested against these antibodies. This approach resembled the phage screening method, in which a biotin-labeled anti-M13 antibody was used to detect interactions between antibodies and peptides displayed multivalently on phages. Since no commercially available antibodies were specific to the synthetic peptides, the peptides were biotinylated using poly-l-Lysine (PLL) as a linker. PLL is widely used in diagnostic assay development, particularly for surface functionalization and signal amplification in biosensor applications55,56,57,58,59. The resulting biotin–PLL–peptide conjugates could be efficiently purified using molecular weight cut-off (MWCO) filters.
Biotin was first conjugated to PLL via carbodiimide chemistry. The remaining free amine groups on the biotin–PLL conjugate were then quantified using the TNBS method, with a standard curve based on l-lysine (Fig. S1). At a PLL concentration of 50 µg/mL, the amine content corresponded to 395.2 ± 3.1 µg/mL of l-lysine. After biotin conjugation, the amine content decreased to 306.2 ± 10.1 µg/mL, indicating that free amine groups were still available for peptide conjugation via click chemistry. The peptides were subsequently conjugated to the biotin–PLL using click chemistry. However, because TNBS quantifies free amine groups, it could not be used to accurately assess the biotin–PLL–peptide conjugates, as the peptides themselves contain amine groups that interfere with the measurement. These biotin–PLL–peptide conjugates were then used to evaluate specificity in the multiplex assay. The results showed that Peptides 1, 3, and 4 specifically bound to IgE without binding to IgA, IgG, or IgM, whereas Peptides 2, 5, and 6 showed no binding to any of the antibody isotypes (Fig. 3A). Peptide 2 appeared to have some ability to bind IgE, but the signal was weak and remained below the cut-off value, defined as three times the background signal. For Peptides 5 and 6, the lack of binding may have resulted from issues during the biotin–PLL–peptide conjugation and/or purification steps. To investigate this, all biotin–PLL–peptide conjugates were tested using a direct ELISA to determine the presence of biotin in each sample. The results showed that all conjugates contained biotin, except for those corresponding to Peptides 5 and 6, which yielded negative ELISA results (Table S2). This suggests that these two conjugates may have aggregated after peptide conjugation, resulting in their loss during the purification process. Although the biotin–PLL–Peptide 2 conjugate tested positive in the ELISA, it did not produce a detectable signal in the multiplex assay, likely due to a low degree of conjugation between Peptide 2 and biotin–PLL. In summary, three peptides exhibited specific binding to IgE, while the remaining peptides may require improved conjugation strategies prior to testing.

Specificity of synthetic peptides. The synthetic peptides were tested against four human isotype antibodies using a direct multiplex assay (A) and a sandwich multiplex assay (B). The dotted lines represent the cut-off value, defined as three times the background value (PBST). Error bars indicate the standard deviation (SD) (n = 3). Peptides binding to the human IgE Fc region were predicted and analyzed using the pepATTRACT web server61.
In the sandwich multiplex assay, peptides were coupled to the beads. In this study, the peptides were synthesized with l-propargylglycine (Pra) at the C-terminus, while the N-terminus was left unmodified. Two conjugation strategies were evaluated to determine the most effective method using Peptide 5 as a model. The first strategy utilized copper-catalyzed azide–alkyne cycloaddition (CuAAC or “click” chemistry), in which the alkyne group of the peptide (from Pra) was conjugated to azido-PEG-amine-functionalized beads48. The second strategy involved carbodiimide chemistry, where the amine group(s) of the peptide were conjugated to the carboxyl groups on the surface of beads using EDC/NHS coupling.
Between the two strategies, in our case, carbodiimide chemistry demonstrated better conjugation performance that the click chemistry method (Fig. S2). Thus, carbodiimide chemistry was applied for all peptides to conjugate the beads. The peptide-conjugated beads were tested with four human immunoglobulin isotypes. The results showed that all peptides could specifically bind to human IgE antibodies with varying normalized signal-to-background values (Fig. 3B). The immunoglobulin isotypes are distinguished by the structure of their constant regions, suggesting that the peptides might target these regions.
To further analyze peptide binding, the binding of synthetic peptides to the human IgE Fc region structure60 was predicted using the pepATTRACT web server61. The performance of pepATTRACT has been previously evaluated using 31 protein–peptide complexes from the peptiDB benchmark62, all featuring unbound receptor structures and 1:1 stoichiometry. In that benchmark, at least one of the 50 predicted models achieved an interface RMSD (iRMSD) below 2 Å for 13 out of 31 complexes, and this accuracy was observed in the top 10 ranked models for 11 of those cases. The full pepATTRACT protocol reached this threshold for 16 cases. According to the interaction prediction model, all peptides displayed wavy conformations, except for Peptides 5 and 6, which exhibited spiral shapes (Fig. 3C). Given that peptides function as molecular binders, their conformations could affect binding to the Fc region of the antibody. According to the binding predictions, Peptide 6 interacted within the Fc structure and also had a spiral shape, leading to a low binding signal in the multiplex assay. In contrast, Peptide 5, despite its spiral conformation, was predicted to interact around the Fc antibody. Furthermore, Peptide 5 contains two l-arginine (R) residues, which play important biological roles due to their positive charges, resulting in the formation of ionic interaction analyte complexes63. Spiral-shaped peptides may exhibit low binding affinity and limited interaction with Fc regions due to several factors. First, their inherent structural rigidity can restrict conformational flexibility necessary for optimal binding. Second, entropic penalties upon binding, as ordered structures lose conformational entropy when interacting with targets. Third, they offer limited interaction surface area, reducing the potential for multivalent or extensive contact with the protein interface64,65,66.
To assess peptide specificity, the peptides were tested against four human immunoglobulin isotypes as well as immunoglobulins from other species, including rabbit, mouse, and goat. All peptides exhibited high specificity for human IgE only (Fig. 4).

Specificity of synthetic peptides against goat, mouse, and rabbit antibodies. The synthetic peptides were tested using a sandwich multiplex assay. The human IgE antibody was used as a positive control. Fluorescent intensity values from peptide binding were normalized to the background (PBST). The dotted lines represent the cut-off value, defined as three times the background value (PBST). Error bars indicate the standard deviation (SD) (n = 3).
With their high specificity, these peptides have the potential to be developed into a test for human IgE detection. The sandwich format is an easy-to-use method, and the use of a sandwich peptide–peptide approach could be beneficial for test kit development. However, the current biotin–PLL–peptide conjugation requires further optimization. The successful development of biotin–PLL–peptide conjugates would enable the creation of a cost-effective and long-term stable test kit. To the best of our knowledge, there have been no prior reports of peptides that specifically bind to human IgE. However, one publication reported novel peptides for the detection of spike protein of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) antibodies. These peptides were able to detect SARS-CoV-2 antibodies in clinical samples and could potentially be developed for use in various methods67.
Sensitivity of synthetic peptides
To determine the sensitivity of peptide detection, the sandwich multiplex assay method was performed. Peptides were used to capture human IgE antibodies, while biotin-labeled goat anti-human IgE antibodies were employed for detection. Five concentrations of human IgE (two-fold serial dilutions ranging from 0.15 to 2.50 µg/mL) were tested (Fig. 5). The limit of detection (LOD) was calculated using three times the background signal (without the addition of IgE) based on linear regression equations. The LOD values for Peptides 1, 2, 3, 4, and 5 were 270, 200, 161, 138, and 69 ng/mL, respectively. However, the LOD for Peptide 6 could not be determined due to significant fluctuations in fluorescence intensity. Additionally, Peptide 6 showed a weak correlation between IgE concentration and signal response (R2 = 0.894), which may be attributed to its conformational structure and binding site characteristics. Specifically, Peptide 6 adopts a spiral conformation and may engages in intramolecular interactions with the IgE molecule (Fig. 3C). Other studies have reported higher-sensitivity methods for IgE detection. For example, ImmunoCAP Fluorescence Enzyme Immunoassay (FEIA) reported an LOD of 0.24 ng/mL4. Other biosensor platforms have demonstrated LODs of 7.2 ng/mL for an electrochemical immunosensor68, 4.8 ng/mL for a quantum dot-based immunochromatographic assay69, and as low as 8.19 fg/mL for an aptamer-based electrochemical biosensor70. The normal range of human serum IgE levels is 50–300 ng/mL (21–125 IU/mL, based on 1 IU/mL = 2.4 ng/mL)71. In allergic patients, total IgE levels are typically > 240 ng/mL (> 100 IU/mL)72. Accordingly, Peptides 2, 3, 4, and 5 demonstrate potential for application in diagnostic tests for allergic patients, whereas Peptide 1 exhibited a higher LOD, limiting its suitability for this purpose. To achieve high-sensitivity detection, we propose further development of a biosensor incorporating these peptides. Furthermore, validation using clinical samples from allergic patients is essential, given the presence of various interfering components in the complex biological matrices.

Limit of detection (LOD) of synthetic peptides. Synthetic peptides were tested against five concentrations of human IgE using the sandwich multiplex assay. Linear regression lines were plotted between concentrations and normalized signal-to-background values. The LOD value was calculated from the linear equation, with a signal-to-background value of 3.
Conclusion
This study successfully identified and characterized novel human IgE-binding peptides using phage display technology. Six unique peptides exhibited high specificity for human IgE, highlighting their potential for use in the development of diagnostic test for allergic diseases. The peptides demonstrated promising performance in multiplex assay, indicating their suitability for point-of-care applications. Nevertheless, further optimization and validation using clinical patient samples are necessary to establish a reliable and efficient diagnostic tool for routine clinical use.
Data availability
The authors declare that the data supporting the findings of this study are available in the paper and its Supplementary Information file.
References
-
Vitte, J., Vibhushan, S., Bratti, M., Montero-Hernandez, J. E. & Blank, U. Allergy, anaphylaxis, and nonallergic hypersensitivity: IgE, mast cells, and beyond. Med. Princ. Pract. Int. J. Kuwait Univ. Health Sci. Cent. 31, 501–515. https://doi.org/10.1159/000527481 (2022).
-
Al, S. & Asilsoy, S. Is there a clinical significance of very low serum immunoglobulin E level?. J. Clin. Immunol. 41, 1893–1901. https://doi.org/10.1007/s10875-021-01127-y (2021).
-
Ünsal, H. et al. Characteristics of patients with low serum IgE levels and selective IgE deficiency: Data from an immunodeficiency referral center. Clin. Immunol. 270, 110403. https://doi.org/10.1016/j.clim.2024.110403 (2025).
-
Hamilton, R. G. Clinical laboratory assessment of immediate-type hypersensitivity. J. Allergy Clin. Immunol. 125, S284–S296. https://doi.org/10.1016/j.jaci.2009.09.055 (2010).
-
Karsonova, A. et al. Highly sensitive ELISA-based assay for quantification of allergen-specific IgE antibody levels. Allergy 75, 2668–2670. https://doi.org/10.1111/all.14325 (2020).
-
Nair, B. et al. Proteomic approach to evaluate mechanisms that contribute to food allergenicity: Comparative 2D-DIGE analysis of radioallergosorbent test positive and negative patients. Int. J. Proteom. 2011, 673618. https://doi.org/10.1155/2011/673618 (2011).
-
Wassmann-Otto, A., Heratizadeh, A., Wichmann, K. & Werfel, T. Microarray analysis confirms ImmunoCAP-fluorescence enzyme immunoassay results on specific IgE in patients with atopic dermatitis and suspected birch pollen-related food allergy. Int. Arch. Allergy Immunol. 183, 814–823. https://doi.org/10.1159/000522525 (2022).
-
Wang, M. et al. Bacterial expression and characterization of a novel human anti-IgE scFv fragment. MAbs 3, 495–499. https://doi.org/10.4161/mabs.3.5.16087 (2011).
-
Hanittinan, O. et al. Feasibility of plant-expression system for production of recombinant anti-human IgE: An alternative production platform for therapeutic monoclonal antibodies. Front. Plant Sci. 13, 1012583. https://doi.org/10.3389/fpls.2022.1012583 (2022).
-
Bognár, Z. & Gyurcsányi, R. E. Aptamers against immunoglobulins: Design, selection and bioanalytical applications. Int. J. Mol. Sci. 21, 5748. https://doi.org/10.3390/ijms21165748 (2020).
-
Lakhin, A. V., Tarantul, V. Z. & Gening, L. V. Aptamers: Problems, solutions and prospects. Acta Nat. 5, 34–43 (2013).
-
Barderas, R. & Benito-Peña, E. The 2018 nobel prize in chemistry: Phage display of peptides and antibodies. Anal. Bioanal. Chem. 411, 2475–2479. https://doi.org/10.1007/s00216-019-01714-4 (2019).
-
Hoogenboom, H. R. et al. Antibody phage display technology and its applications. Immunotechnology 4, 1–20. https://doi.org/10.1016/S1380-2933(98)00007-4 (1998).
-
Ferreira, D. et al. Rational identification of a colorectal cancer targeting peptide through phage display. Sci. Rep. 9, 3958. https://doi.org/10.1038/s41598-019-40562-1 (2019).
-
Saw, P. E. & Song, E. W. Phage display screening of therapeutic peptide for cancer targeting and therapy. Protein Cell 10, 787–807. https://doi.org/10.1007/s13238-019-0639-7 (2019).
-
Xing, X. et al. Target-specific peptides for BK virus agnoprotein identified through phage display screening: Advancing antiviral therapeutics. Sci. Rep. 15, 2718. https://doi.org/10.1038/s41598-025-86439-4 (2025).
-
Ivanova, A. et al. In vivo phage display identifies novel peptides for cardiac targeting. Sci. Rep. 14, 12177. https://doi.org/10.1038/s41598-024-62953-9 (2024).
-
Aghebati-Maleki, L. et al. Phage display as a promising approach for vaccine development. J. Biomed. Sci. 23, 66. https://doi.org/10.1186/s12929-016-0285-9 (2016).
-
Hess, K. L. & Jewell, C. M. Phage display as a tool for vaccine and immunotherapy development. Bioeng. Transl. Med. 5, e10142. https://doi.org/10.1002/btm2.10142 (2020).
-
Palma, M. Epitopes and mimotopes identification using phage display for vaccine development against infectious pathogens. Vaccines 11, 1176 (2023).
-
Moreira, G., Fühner, V. & Hust, M. Epitope mapping by phage display. Methods Mol. Biol. 1701, 497–518. https://doi.org/10.1007/978-1-4939-7447-4_28 (2018).
-
Pérez-Martínez, D., Infante, Y. C., Ramírez, B. S. & Rojas, G. Domain-level epitope mapping of polyclonal antibodies against HER-1 and HER-2 receptors using phage display technology. Sci. Rep. 12, 12268. https://doi.org/10.1038/s41598-022-16411-z (2022).
-
Zahirovic, A. et al. Identification of bee venom Api m 1 IgE epitopes and characterization of corresponding mimotopes. J. Allergy Clin. Immunol. 143, 791–794. https://doi.org/10.1016/j.jaci.2018.10.003 (2019).
-
Davies, J. M., O’Hehir, R. E. & Suphioglu, C. Use of phage display technology to investigate allergen–antibody interactions. J. Allergy Clin. Immunol. 105, 1085–1092. https://doi.org/10.1067/mai.2000.107040 (2000).
-
Malmborg, A.-C., Dueñas, M., Ohlin, M., Söderlind, E. & Borrebaeck, C. A. K. Selection of binders from phage displayed antibody libraries using the BIAcore™ biosensor. J. Immunol. Methods 198, 51–57. https://doi.org/10.1016/0022-1759(96)00159-7 (1996).
-
Peltomaa, R., Benito-Peña, E., Barderas, R. & Moreno-Bondi, M. C. Phage display in the quest for new selective recognition elements for biosensors. ACS Omega 4, 11569–11580. https://doi.org/10.1021/acsomega.9b01206 (2019).
-
Takakusagi, Y., Takakusagi, K., Sakaguchi, K. & Sugawara, F. Phage display technology for target determination of small-molecule therapeutics: An update. Expert Opin. Drug Discov. 15, 1199–1211. https://doi.org/10.1080/17460441.2020.1790523 (2020).
-
Burton, K., Ghadami, S., Dellinger, K., Wang, B. & Dong, M. Screening peptide-binding partners for GenX via phage display. Int. J. Mol. Sci. 25, 2686 (2024).
-
Charlermroj, R., Makornwattana, M., Phuengwas, S. & Karoonuthaisiri, N. A rapid colorimetric lateral flow test strip for detection of live Salmonella Enteritidis using whole phage as a specific binder. Front. Microbiol. 13, 1–11. https://doi.org/10.3389/fmicb.2022.1008817 (2022).
-
Morton, J. et al. Phage display-derived binders able to distinguish Listeria monocytogenes from other Listeria species. PLoS ONE 8, e74312. https://doi.org/10.1371/journal.pone.0074312 (2013).
-
Karoonuthaisiri, N. et al. Development of a M13 bacteriophage-based SPR detection using Salmonella as a case study. Sens. Actuators B Chem. 190, 214–220. https://doi.org/10.1016/j.snb.2013.08.068 (2014).
-
Niyomdecha, S. et al. Phage-based capacitive biosensor for Salmonella detection. Talanta 188, 658–664. https://doi.org/10.1016/j.talanta.2018.06.033 (2018).
-
Kim, T. et al. Screening of novel peptides that specifically interact with vitamin D bound biocomplex proteins. Sci. Rep. 13, 2116. https://doi.org/10.1038/s41598-023-28881-w (2023).
-
Garcia-Calvo, E. et al. Construction of a fab library merging chains from semisynthetic and immune origin, suitable for developing new tools for gluten immunodetection in food. Foods 12, 149. https://doi.org/10.3390/foods12010149 (2022).
-
Garcia-Calvo, E. et al. Development of a new recombinant antibody, selected by phage-display technology from a celiac patient library, for detection of gluten in foods. Curr. Res. Food Sci. 7, 100578. https://doi.org/10.1016/j.crfs.2023.100578 (2023).
-
Laukkanen, M.-L. et al. Hevein-specific recombinant IgE antibodies from human single-chain antibody phage display libraries. J. Immunol. Methods 278, 271–281. https://doi.org/10.1016/S0022-1759(03)00070-X (2003).
-
Dhar, P., Samarasinghe, R. M. & Shigdar, S. Antibodies, nanobodies, or aptamers—Which is best for deciphering the proteomes of non-model species?. Int. J. Mol. Sci. 21, 2485. https://doi.org/10.3390/ijms21072485 (2020).
-
Al Musaimi, O. & Lombardi, L. Strategies for improving peptide stability and delivery. Pharmaceuticals 15, 1283. https://doi.org/10.3390/ph15101283 (2022).
-
Prickett, S. R. et al. Safety and tolerability of a novel peptide-based immunotherapy for peanut allergy. J. Allergy Clin. Immunol. 143, AB431. https://doi.org/10.1016/j.jaci.2018.12.975 (2019).
-
Larché, M. Update on the current status of peptide immunotherapy. J. Allergy Clin. Immunol. 119, 906–909. https://doi.org/10.1016/j.jaci.2007.02.015 (2007).
-
Kinnunen, T. et al. Potential of an altered peptide ligand of lipocalin allergen Bos d 2 for peptide immunotherapy. J. Allergy Clin. Immunol. 119, 965–972. https://doi.org/10.1016/j.jaci.2007.01.011 (2007).
-
Worm, M. et al. Development and preliminary clinical evaluation of a peptide immunotherapy vaccine for cat allergy. J. Allergy Clin. Immunol. 127, 89-97.e14. https://doi.org/10.1016/j.jaci.2010.11.029 (2011).
-
Down, G., Evans, K. L., Larché, M. & Laidler, P. Field study of a novel synthetic peptide immunotherapy in cat allergic patients. J. Allergy Clin. Immunol. 143, AB432. https://doi.org/10.1016/j.jaci.2018.12.979 (2019).
-
Hafner, R. P., Salapatek, A., Patel, D., Larché, M. & Laidler, P. Validation of peptide immunotherapy as a new approach in the treatment of allergic rhinoconjunctivitis: The clinical benefits of treatment with Amb a 1 derived T cell epitopes. J. Allergy Clin. Immunol. 129, AB368. https://doi.org/10.1016/j.jaci.2012.01.017 (2012).
-
Anand, T. & Virmani, N. Phage display technique as a tool for diagnosis and antibody selection for coronaviruses. Curr. Microbiol. 78, 1124–1134. https://doi.org/10.1007/s00284-021-02398-9 (2021).
-
The UniProt Consortium. UniProt: The universal protein knowledgebase in 2023. Nucleic Acids Res. 51, D523–D531. https://doi.org/10.1093/nar/gkac1052 (2022).
-
Dunbar, S. A. Applications of Luminex® xMAP™ technology for rapid, high-throughput multiplexed nucleic acid detection. Clin. Chim. Acta 363, 71–82. https://doi.org/10.1016/j.cccn.2005.06.023 (2006).
-
Coppock, M. B. & Stratis-Cullum, D. N. A universal method for the functionalization of dyed magnetic microspheres with peptides. Methods 158, 12–16. https://doi.org/10.1016/j.ymeth.2019.01.014 (2019).
-
Habeeb, A. F. S. A. Determination of free amino groups in proteins by trinitrobenzenesulfonic acid. Anal. Biochem. 14, 328–336. https://doi.org/10.1016/0003-2697(66)90275-2 (1966).
-
Dunbar, S. A., Vander Zee, C. A., Oliver, K. G., Karem, K. L. & Jacobson, J. W. Quantitative, multiplexed detection of bacterial pathogens: DNA and protein applications of the Luminex LabMAP system. J. Microbiol. Methods 53, 245–252 (2003).
-
Yu, S., Zhu, B., Lv, F., Li, S. & Huang, W. Rapid analysis of cyclamate in foods and beverages by gas chromatography-electron capture detector (GC-ECD). Food Chem. 134, 2424–2429. https://doi.org/10.1016/j.foodchem.2012.04.028 (2012).
-
Lawaniya, S. D., Pandey, G., Yu, Y. & Awasthi, K. Efficient detection of p-nitrophenol via a polypyrrole flower-decorated nickel foam-based electrochemical sensor. Nanoscale 16, 13915–13924. https://doi.org/10.1039/d4nr01580k (2024).
-
Das, S., Nikolaidis, N., Klein, J. & Nei, M. Evolutionary redefinition of immunoglobulin light chain isotypes in tetrapods using molecular markers. Proc. Natl. Acad. Sci. U.S.A. 105, 16647–16652. https://doi.org/10.1073/pnas.0808800105 (2008).
-
Sinkjaer, A. W. et al. A comparative analysis of sequence composition in different lots of a phage display peptide library during amplification. Virol. J. 22, 24. https://doi.org/10.1186/s12985-024-02600-x (2025).
-
Movilli, J., Rozzi, A., Ricciardi, R., Corradini, R. & Huskens, J. Control of probe density at DNA biosensor surfaces using poly(l-lysine) with appended reactive groups. Bioconjug. Chem. 29, 4110–4118. https://doi.org/10.1021/acs.bioconjchem.8b00733 (2018).
-
Estrada-Osorio, D. V., Escalona-Villalpando, R. A., Gutiérrez, A., Arriaga, L. G. & Ledesma-García, J. Poly-l-lysine-modified with ferrocene to obtain a redox polymer for mediated glucose biosensor application. Bioelectrochemistry 146, 108147. https://doi.org/10.1016/j.bioelechem.2022.108147 (2022).
-
Gao, J. et al. Poly-l-lysine-modified graphene field-effect transistor biosensors for ultrasensitive breast cancer miRNAs and SARS-CoV-2 RNA detection. Anal. Chem. 94, 1626–1636. https://doi.org/10.1021/acs.analchem.1c03786 (2022).
-
Gong, P. et al. Probe type optical fiber DNA hybridization biosensor based on polyethylenimine and poly-l-lysine surface functionalization method. Opt. Laser Technol. 168, 109978. https://doi.org/10.1016/j.optlastec.2023.109978 (2024).
-
Wang, C., Chen, L., Wang, P., Li, M. & Liu, D. A novel ultrasensitive electrochemiluminescence biosensor for glutathione detection based on poly-l-lysine as co-reactant and graphene-based poly(luminol/aniline) as nanoprobes. Biosens. Bioelectron. 133, 154–159. https://doi.org/10.1016/j.bios.2019.03.016 (2019).
-
Doré, K. A. et al. Thermal sensitivity and flexibility of the Cε3 domains in immunoglobulin E. Biochim. Biophys. Acta Proteins Proteom. 1336–1347, 2017. https://doi.org/10.1016/j.bbapap.2017.08.005 (1865).
-
de Vries, S. J., Rey, J., Schindler, C. E. M., Zacharias, M. & Tuffery, P. The pepATTRACT web server for blind, large-scale peptide-protein docking. Nucleic Acids Res. 45, W361-w364. https://doi.org/10.1093/nar/gkx335 (2017).
-
Schindler, C. E. M., de Vries, S. J. & Zacharias, M. Fully blind peptide-protein docking with pepATTRACT. Structure 23, 1507–1515. https://doi.org/10.1016/j.str.2015.05.021 (2015).
-
Martins, J. N., Lima, J. C. & Basilio, N. Selective recognition of amino acids and peptides by small supramolecular receptors. Molecules 26, 106. https://doi.org/10.3390/molecules26010106 (2020).
-
Diderich, P. et al. Phage selection of chemically stabilized α-helical peptide ligands. ACS Chem. Biol. 11, 1422–1427. https://doi.org/10.1021/acschembio.5b00963 (2016).
-
Miki, T. et al. Pattern enrichment analysis for phage selection of stapled peptide ligands. Chem. Sci. 13, 12634–12642. https://doi.org/10.1039/d2sc04058a (2022).
-
Dudak, F. C., Boyaci, I. H. & Orner, B. P. The discovery of small-molecule mimicking peptides through phage display. Molecules 16, 774–789 (2011).
-
Bulut, A., Temur, B. Z. & Kirimli, C. E. A novel peptide-based detection of SARS-CoV-2 antibodies. Biomimetics 8, 89. https://doi.org/10.3390/biomimetics8010089 (2023).
-
Park, M. et al. Electrochemical immunosensor for human IgE using ferrocene self-assembled monolayers modified ITO electrode. Biosensors 10, 38 (2020).
-
Berlina, A. N. et al. Quantum-dot-based immunochromatographic assay for total IgE in human serum. PLoS ONE 8, e77485. https://doi.org/10.1371/journal.pone.0077485 (2013).
-
Lin, X. et al. Self-assembly of functional nucleic acid-based colorimetric competition assay for the detection of immunoglobulin E. Sensors 19, 2224 (2019).
-
Platts-Mills, T. A., Snajdr, M. J., Ishizaka, K. & Frankland, A. W. Measurement of IgE antibody by an antigen-binding assay: Correlation with PK activity and IgG and IgA antibodies to allergens. J. Immunol. 120, 1201–1210 (1978).
-
Li, Y. et al. Combination of magnetic beads extraction and ultraperformance liquid chromatography tandem mass spectrometry detection for the clinical diagnosis of allergies. Anal. Chim. Acta 1221, 340157. https://doi.org/10.1016/j.aca.2022.340157 (2022).
Acknowledgements
This project was financially supported by the National Science and Technology Development Agency (NSTDA) under Grant Agreement No. P2350623.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kaew-amdee, S., Makornwattana, M. & Charlermroj, R. Identification of novel human IgE-binding peptides from a phage display library for total IgE detection. Sci Rep 15, 27986 (2025). https://doi.org/10.1038/s41598-025-12574-7
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12574-7