Introduction
In an era of increasing health consciousness, functional foods, particularly those enriched with probiotic microorganisms, reflect a heightened consumer awareness of the intricate link between diet and health, simultaneously driving vigorous scientific inquiry in this domain1. Probiotics are defined as “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host”2. These beneficial microbes have garnered substantial scientific and commercial interest due to their diverse positive impacts on the host, encompassing immunomodulatory3, antibacterial4, anticancer5, and hypolipidemic effects6. Their recognized capacity to favorably alter the immune system and reshape the intestinal microbiota has positioned them as valuable preventatives and therapeutics in both human and veterinary medicine7. In animal nutrition, probiotics have been extensively investigated for their role in controlling harmful gut pathogens and promoting a balanced gut microbiome (eubiosis)8. This leads to a cascade of benefits, including enhanced gastrointestinal tract functionality, improved gut and systemic immunity, and overall improved health status in both ruminant and non-ruminant livestock9. Specifically in ruminants, probiotic administration primarily aims to augment ruminal fermentation efficiency through mechanisms such as pH stabilization, improved fiber digestion, and reduced rumen methane generation, ultimately enhancing production output10.
Lactic acid bacteria (LAB) are pivotal in food production and preservation, and increasingly recognized for their role in enhancing livestock productivity11. Historically, LAB have served as natural fermenters and integral components in various industries, including food, pharmaceutical, and feed, as they are widely recognized as food-grade, non-toxic microorganisms safe for human consumption12. Important LAB members include the genera Lactobacillus13, Lactococcus14, Leuconostoc15, Pediococcus16, and Streptococcus17.
Pediococcus spp. are Gram-positive, catalase-negative, facultatively anaerobic lactic acid bacteria16. They typically exhibit a coccus morphology, often arranged in tetrads. These microorganisms are non-motile, do not form spores, and are frequently referred to as probiotic bacteria16. Diverse Pediococcus species, such as P. acidilactici M7618, P. parvulus 19, P. damnosus20 and P. pentosaceus M4121, exhibit varied industrial applications. For instance, P. pentosaceus is widely utilized as a starter culture in the food industry, particularly for fermenting meat and vegetable products22. One Pediococcus probiotic strain that has been extensively studied is P. lolii23, a strain sharing a high degree of similarity (97.7%) with P. acidilactici24. Despite the growing accessibility of comprehensive genomic data on Pediococcus species and expanding research into diverse strains, novel strains continue to emerge from untapped sources, potentially harboring unique characteristics. Whole-genome sequencing is rapidly expanding our comprehension of Pediococcus diversity and functionality25. Recent investigations have illuminated the metabolic and probiotic potential of distinct Pediococcus lineages. For instance, the identification of novel exopolysaccharide synthesis clusters by Lu et al.26 and the characterization of probiotic P. acidilactici by Zhao et al.27 exemplify the power of genomics in discerning strain-specific attributes. Furthermore, Li et al.28 have advanced our understanding of bacteriocin production and core genome architecture within the Pediococcus genus. These collective efforts highlight the crucial role of genomics in unlocking the biotechnological potential of Pediococcus strains.
In this study, strain P10 was isolated from the gut microbiota of native broiler chickens. Our investigation employed comprehensive in vitro analyses to assess its key probiotic characteristics, including acid and bile tolerance, absence of hemolytic activity, and adherence to intestinal epithelial cells. Complementing these phenotypic assays, whole-genome sequencing was performed to further elucidate the strain’s probiotic potential and identify unique genomic features.
Results
Examination of probiotic properties
The 16S rRNA gene of the selected strain was sequenced via the Sanger method, and the resulting sequence was deposited in the GenBank database (accession number MG255949.1). To assess safety, hemolytic activity of strain P10 was evaluated on blood agar plates, which revealed no lysis of red blood cells, classifying the strain as gamma-hemolytic. This finding was further corroborated by bioinformatics analyses, which confirmed the absence of genes typically associated with hemolytic activity.
Strain P10’s survivability was assessed under simulated gastrointestinal conditions. It showed high survival rates at both pH 3 (95% viability) and pH 4 (99% viability), with no statistically significant difference (p > 0.05) (Table 1). The isolate also maintained a significant survival rate of 55% in the presence of 0.3% bile salts after 8 h of incubation (p > 0.05) (Table 1).
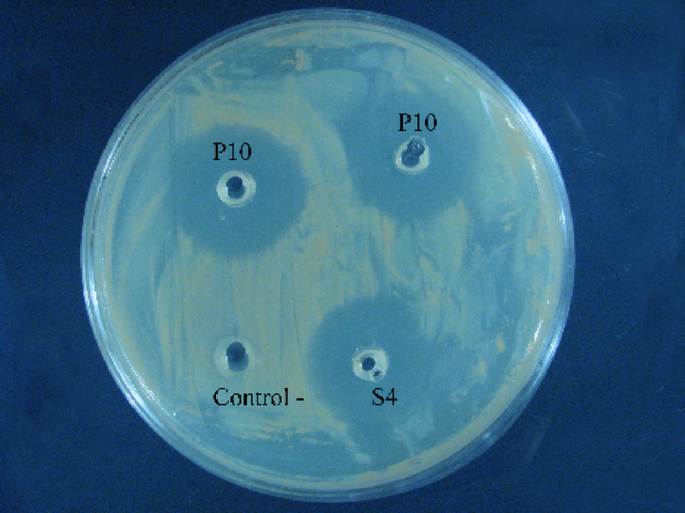
We investigated strain P10’s in vitro antagonistic effect against a panel of indicator pathogens using the well diffusion method. P10 showed robust antimicrobial activity against various pathogenic bacteria, including E. coli ATCC 25922, S. enterica serovar Typhimurium ATCC 14028, S. aureus ATCC 25923, L. monocytogenes ATCC 119115, and P. aeruginosa ATCC 27853 (Table 2). The observed inhibition zones consistently exceeded 2 mm in diameter (Fig. 1).

Agar plate containing pathogenic bacteria showing the antagonistic effect of strain P10 and controls. Wells containing the cell-free supernatant of strain P10 (P10), negative control (Control -), and comparative strain L. salivarius (S4, Control +) are indicated on the plate. The clear inhibition zones around the P10 wells indicate the antimicrobial activity of this strain.
Antibiotic susceptibility testing was performed on the isolate against a panel of commonly used antibiotics. Results, summarized in Table 3, revealed resistance to ciprofloxacin, fosfomycin, enrofloxacin, colistin, sulfamethoxazole/trimethoprim (SXT), and flumequine. The isolate demonstrated sensitivity to all other tested antibiotics.
Strain P10’s adherence to Caco-2 cells was evaluated, with microscopic analysis revealing a notably greater tendency for the strain to adhere compared to the control across different fields of observation (Fig. 2).

Examining the adhesion level of a: P. acidilactici P10 b: E. coli strain ATCC 25,922 to Caco-2 cell line.
Genome assembly and annotation
Next-generation shotgun sequencing of strain P10 yielded 10,705,328 raw reads (150 bp each), totaling 1.6 Gigabases. Following quality control and trimming, 10,509,008 clean reads were used for downstream analysis. De novo assembly of the P10 genome resulted in a bacterial genome with a length of 1,847,305 bp (Fig. 3).

With the use of the CGView tool, a circular map of the P10 genome could be shown. Circles 1 and 2 show Prokka annotated forward and reverse CDS (coding sequence), respectively, with tRNA and rRNA; circles 3 and 4 indicate GC content and GC skew; circles 5 represents genome size.
Genome annotation using Prokka predicted a total of 1,889 genes, comprising 1,832 protein-coding sequences (CDS), 53 tRNAs, 3 rRNAs, and 1 transfer-messenger RNA (tmRNA) (Table 4, Supplementary File S1). Of these, 1,207 genes (64.23%) were functionally annotated, while 672 genes (35.76%) were classified as hypothetical/unknown. The 44 identified tRNA sequences corresponded to 21 natural amino acids, with varying counts: Leu (6), Arg and Leu (5), Ser (4), Gly and Met (3), Gln, Asn, Glu, Thr, Lys, Pro, and Asp (2), and Ala, Phe, Ile, Met, Cys, His, Trp, Val, and Tyr (1).
Phylogenetic analysis
Using Hmmalign for genome similarity alignment, strain P10 showed 99.57% similarity with Pediococcus species (MG255949). This analysis confirmed that P10 is most closely related to P. acidilactici (Fig. 4).

Taxonomic assignment of P. acidilactici P10. The tree was inferred using TYGS webserver of P10 genes comparing to 19225 genomes of microbial collection available at the TYGS database. The tree was inferred with FASTME 2.1.6.1. from GBDP distances calculated from 16S rDNA gene sequences.
Identification and characterization of a type II-A CRISPR-Cas system
Bioinformatic analysis of the P. acidilactici P10 genome revealed the presence of a complete and highly conserved CRISPR-Cas system. Using CRISPRDetect, a prominent CRISPR array (designated Array 1 or P7741_1) was identified spanning positions 316,209 and 317,565 in the forward direction. This array comprises 20 unique spacer sequences, each 30 bp in length, interspersed by highly conserved 36 bp repeat units. The consensus repeat sequence was determined as GTTTCAGAAGGATGTTAAATCAATAAGGTTAAGATC. The array’s high integrity and perfect repeat identity (100%), coupled with a high confidence score from CRISPRDetect, strongly suggest a functional and actively maintained locus (Supplementary File S2). Further analysis with CRISPRCasFinder classified this system as a Type II-A CRISPR-Cas system. Crucially, this system harbored the core cas genes characteristic of this type, including cas9, cas1, cas2, and csn2. The presence of cas1 and cas2 indicates the capacity for new spacer acquisition, implying an active adaptive immune response. The identified cas9 gene is particularly notable as it encodes the effector nuclease commonly utilized in genome-editing applications (Supplementary File S3).
Spacer analysis for phage resistance
To investigate the potential role of this CRISPR-Cas system in phage defense, we queried the 20 identified spacer sequences against known bacteriophage genomes using BLASTn. Of these, 17 spacers showed 100% identity to sequences within the genomes of various bacteriophages, confirming prior encounters with these viral elements. These matching protospacer sequences primarily belonged to unclassified bacteriophages identified as Metagenome-Assembled Genomes (MAGs) from diverse environments (e.g., isolate 3676_35671, isolate 2563_4092, isolate ct0Ms11, isolate ctgiD18). While specific host ranges for these phages aren’t always definitively assigned in public databases, their “Bacteriophage sp.” designation implies their potential to interact with bacterial hosts. The remaining 3 spacers didn’t show significant homology to any currently available phage genomes in public databases, suggesting they may target novel or uncharacterized phages, or other mobile genetic elements (Supplementary File S4).
Pan-genome analysis of genome
To assess the genomic diversity of P. acidilactici strains, we conducted a pan-genome analysis on 38 available complete genomes of P. acidilactici (Supplementary File S5). Our results revealed a dynamic genomic landscape, where the pan-genome comprised 5,233 orthologous gene clusters. This included a core genome of 1,248 gene clusters shared by all strains, a diverse accessory genome with 1,261 shell gene clusters (15–95% prevalence), and 2,724 cloud gene clusters (less than 15% prevalence) (Fig. 5a, Supplementary File S6). A comprehensive gene presence/absence matrix was generated for the analyzed strains, which revealed striking similarities in gene cluster distribution among certain strains. Notably, genomes with accession numbers GCA_13127755.1, GCA_019448175.1, and GCA_011604585.1 exhibited highly conserved gene clusters. The analysis also revealed a significant degree of genetic diversity among strains, with the number of strain-specific genes varying substantially from a single unique gene in some strains to a remarkable 162 in others. On average, each strain carried approximately 40 unique genes, representing about 2% of its genome, highlighting the substantial impact of strain-specific variation (Fig. 5c). Crucially, our newly discovered strain P10 harbored the most strain-specific genes, totaling 59 unique genes (Fig. 5b). COG classification of the proteins encoded by these unique genes was performed using EggNOG-mapper, assigning them to 10 functional groups. The majority (9 genes, COG Class S) were classified as unknown function. Additionally, 7 genes were categorized under Carbohydrate transport and metabolism (COG Class G), and 5 genes were assigned to Replication, recombination, and repair (COG Class L). This unique genetic repertoire in P10 highlights its potential for novel biological functions.

Pan-genome analysis of 37 P. acidilactici available complete genomes at NCBI database strains and the P10 revealed a total of 5233 genes, highlighting the extensive genetic diversity within this species. a) The pie chart showing how genes are distributed among genotypes b) Heatmap showing the presence and absence of genes based on pan-genome results c) Bar chart showing the number of genes plotted based on the distribution of genomes, with most genes present in only one strain.
In addition to a core genome of 1,248 genes shared among P. acidilactici strains, our pan-genome analysis of P. acidilactici P10 uniquely identified 59 strain-specific genes not present in other analyzed genomes. KEGG29 pathway analysis of these unique genes (Supplementary File S7) revealed their potential functional significance, particularly in diverse metabolic pathways, membrane transport, and environmental adaptation.
Specifically, these unique genes were enriched in pathways related to:
-
Global and Overview Maps: including Metabolic pathways, Biosynthesis of secondary metabolites, Carbon metabolism, Biosynthesis of amino acids, and Biosynthesis of cofactors.
-
Carbohydrate Metabolism: such as Glycolysis/Gluconeogenesis, the Pentose phosphate pathway, and Starch and sucrose metabolism.
-
Lipid Metabolism: encompassing Fatty acid biosynthesis and Glycerolipid metabolism.
-
Amino Acid Metabolism: specifically Cysteine and methionine metabolism.
-
Metabolism of Cofactors and Vitamins: including Biotin metabolism.
-
Membrane Transport: notably, the Phosphotransferase system (PTS).
Intriguingly, a unique gene associated with “Plant-pathogen interaction” was also identified, suggesting specific environmental response mechanisms. These findings collectively indicate that P. acidilactici P10 possesses distinct metabolic capabilities and adaptive advantages, differentiating it from other strains within the species.
Intra-species phylogenetic tree analysis
The phylogenetic relationships of P10 and 37 other P. acidilactici strains were elucidated based on pan-genome analysis. As shown in Fig. 6, P10 clustered within the same clade as SRCM103367 (GCA_004102605.1), demonstrating close genetic proximity to the ZY271 (GCA_019844055.1) genome.

Phylogeny of the P. acidilactici strains based on the analysis of pan-genomes.
P. acidilactici P10 comparative genome analysis
Based on the pan-genome phylogenetic tree, we selected seven closely related reference genomes for a detailed genome assembly comparison. Using progressive Mauve, we aligned the ordered genome assembly of strain P10 against strain PMC65 (GenBank: GCA_013127755.1), chosen as the best reference genome for P. acidilactici in NCBI. This alignment revealed 27 collinear blocks (Fig. 7), indicating nearly identical chromosomal arrangements between these strains, with large segments of high similarity across most portions of their chromosomes.

Genome alignment showing syntenic blocks between P. acidilactici strains GCA_013127755.1 (top) and P10 genomes (bottom).
We also analyzed additional genomic features of P10 and these seven closely related genomes, including sequence similarity, GC content distribution, and the number of genes (tRNA and rRNA) (Fig. 8, Table 5). P10’s genome size, total gene count, and predicted CDS number were similar to, but slightly smaller than, those of SRCM103367 and ZY271. Notably, SRCM103367 appeared to have a larger genome size of approximately 1.8 Mb.

Genome comparisons of P10 strain and other P. acidilactici strains against reference genome (strain PMC65) generating by BRIG 0.95, the circular map illustrates the whole genome comparison of strain PMC65 against the other 7 sequenced P. acidilactici. The inner cycle (back) represents the complete genome of the reference strain PMC65 and the shade of each colors show the similarities between each strain with strain PMC65.
Functional annotation
Analysis of the Clusters of Orthologous Groups (COG) functional categories using EggNOG-mapper v2 assigned 1,684 genes (94.1%) into 21 clusters (Fig. 9, Supplementary File S8). The largest proportion of genes fell under “Function unknown” (S: 320 genes). Other prominent functional categories included:
-
Carbohydrate transport and metabolism (G: 160 genes)
-
Translation, ribosomal structure, and biogenesis (J: 158 genes)
-
Transcription (K: 149 genes)
-
Replication, recombination, and repair (L: 119 genes)
-
Cell wall/membrane/envelope biogenesis (M: 107 genes)
-
Nucleotide transport and metabolism (F: 101 genes), among others.

Distribution of Cluster of Orthologous group (COG) functional categories to the proteins of P10.
Collectively, the functional annotations from both COG and KEGG29 searches indicate P10’s high potential metabolic capacity. The abundance of genes related to genetic information processing (COG categories J, K, L), followed by transport and metabolism (COG categories E, P, G, F, H), underscores the significance of these genes for P10’s conserved cellular mechanisms, enabling it to thrive in a wide range of environments (Supplementary File S9).
Probiotic properties
To genomically ascertain the probiotic properties of strain P10, we examined its genome for several genes associated with key probiotic traits, including stress resistance, bile salt hydrolase activity, adhesion ability, and immunomodulatory activities, based on a comprehensive literature review. As summarized in Table 6, numerous stress-related genes were detected within the P10 genome.
Based on BlastKOALA results, seven heat shock-related genes, including regulators (hrcA, htpX, and ctsR) and molecular chaperones (dnaK, dnaJ, grpE, and hslO), were identified as encoded by the P10 genome30. P10 also encodes two cold shock protein genes (cspC and cspLA), essential for survival at sub-freezing temperatures31. The P10 genome consequently contained nine genes encoding low pH resistance32 and two genes for bile resistance30. The cell surface proteins of probiotic strains are crucial for their capacity to adhere to the host epithelium. Strain P10 possesses seven putative adhesion-related genes, including those encoding triosephosphate isomerase (tpiA), L-glyceraldehyde 3-phosphate reductase (gpr), elongation factor Tu (tuf), and lipoprotein signal peptidase II (lspA), all of which may contribute to a high adhesion capacity. Additionally, the genome contained the xylA gene, which is important for intestinal persistence32. Six of P10’s nine oxidative stress-related genes (tpx, trxA, trxB, ndh, npr, and nox) encode the complete thioredoxin and NADH antioxidant systems, which are involved in reactive oxygen species (ROS) scavenging33,34.
Antibiotic resistance genes
Two antibiotic resistance genes such as ermB, tetM were found in Resfinder 4.5 database (90% threshold and 60% minimum length). While only five gene coding vanT, vanG, tetM, qacG and sdrM, was detected in the CARD database search under default parameters (perfect and strict hits, only). The BlastKOALA results revealed that, there are 16 genes in the P10 genome which are associated with antibiotic resistance (Table 7). The identified genes were related to Lincomycin (lmrB), tetracycline (tetM, tetO), beta-lactame (penP), Vancomycin (vanX), Ciprofloxacin (nisG, spaG and cprC) and Colistin (nisE, spaE, cprB, epiE,nisF, spaF, cprA, epiF).
In line with the findings from bioinformatics analysis, the in vitro antibiotic susceptibility testing (Table 3) also confirmed phenotypic resistance. Out of 17 antibiotics tested, the strain showed resistance to 7: ciprofloxacin (CP), streptomycin (S), fosfomycin (FO), enrofloxacin (NFX), colistin (CL), sulfamethoxazole/trimethoprim (SXT), and flumequine (FM). Conversely, it demonstrated sensitivity to the remaining 10 antibiotics: chloramphenicol (CC), tetracycline (TE), clindamycin (C), penicillin (P), erythromycin (E), ampicillin (AM), rifampicin (RA), gentamicin (GM), tylosin (TY), and florfenicol (FF).
Antimicrobial activity
P. acidilactici exhibits broad-spectrum antimicrobial properties, primarily through the production of bacteriocins like pediocin, which target and disrupt bacterial cell membranes. This multifaceted activity makes it a promising candidate for enhancing food safety and potentially promoting gut health by suppressing pathogenic microorganisms within the gastrointestinal tract. Furthermore, its lactic acid production effectively lowers environmental pH, creating an acidic milieu that inhibits the growth of many undesirable microorganisms. Some strains also produce hydrogen peroxide and diacetyl, further augmenting their antimicrobial effects. The overall effectiveness of these mechanisms, however, varies depending on factors such as bacteriocin production levels, lactic acid concentration, and specific strain characteristics.
BaPres analysis predicted 57 proteins with potential bacteriocin activity, with the majority exhibiting high probabilities (> 0.9), strongly suggesting antimicrobial potential. Further bioinformatic analysis revealed the presence of genes encoding specific putative bacteriocins, including plaA (prebacteriocin) and a putative entrocin P bacteriocin. These types of bacteriocins have been previously reported in Pediococcus species, such as P. acidilactici35 and P. pentosaceus36, as evidenced by BLAST results against known bacteriocin sequences in UniProt.
Furthermore, genomic analysis identified several additional genes with putative antimicrobial activity in strain P10, such as bhtA, Col, and truE3, among others (Supplementary File S10). The presence of these genes, including bhtA which is known to be involved in bacteriocin production, collectively suggests that strain P10 possesses a broad spectrum of antimicrobial properties.
Immune system
Based on bioinformatic analyses, several key genes including TrxA, cas9, cas1, cas2, MPRF, and CatA were identified within the P10 genome. These genes are predicted to play crucial roles in enhancing bacterial immune defense mechanisms and host interaction. Specifically, TrxA contributes to redox homeostasis; the cas genes are involved in CRISPR-Cas-mediated phage resistance; and MPRF confers resistance to cationic antimicrobial peptides (AMPs) produced by the host immune system. Furthermore, CatA protects against oxidative stress.
Notably, BlastKOALA analysis also identified genes in P10 implicated in modulating the host immune response or interacting with a range of bacterial infections, including those caused by E. coli, Salmonella, S. aureus, Legionella and M. tuberculosis.
Discussion
Our study successfully sequenced and annotated the genome of P. acidilactici strain P10, providing a foundational understanding of its genetic makeup. The 1.84 Mb circular chromosome, characteristic of the genus, revealed a compact genome size consistent with other Pediococcus species such as P. acidilactici JKY1828, P. parvulus37, and P. pentosaceus MR00122. The presence of a comprehensive set of tRNA genes, along with rRNA and tmRNA, suggests a robust translational machinery vital for its metabolic activities. Furthermore, the GC content aligned with established ranges for Pediococcus strains, a factor linked to microbial energy expenditure and genetic stability38,39.
Phylogenetic analysis based on the pan-genome clustered strain P10 closely with P. acidilactici SRCM103367 and ZY271, suggesting a shared evolutionary lineage and potentially similar ecological niches or functional adaptations. The functional annotation of P10’s genes highlights its strong metabolic capabilities, with a significant abundance of genes involved in carbohydrate metabolism, translation, ribosomal structure, and transcription. The high number of genes dedicated to genetic information processing and various transport and metabolism pathways underscores P10’s inherent metabolic versatility, enabling it to thrive across diverse environments40.
Strain P10’s genomic profile strongly supports its potential as a robust probiotic. Its notable stress tolerance is evident through the identification of genes involved in managing various environmental challenges. Seven heat shock-related genes, including crucial molecular chaperones and regulators (e.g., dnaK, hrcA), align with their established roles in maintaining protein homeostasis and membrane stability under elevated temperatures in Pediococcus strains11. Similarly, the presence of cold shock protein genes (cspC, cspLA) suggests P10’s adaptability to low temperatures, consistent with observations in other Pediococcus strains that synthesize CSP family proteins to counteract cold stress41. P10’s acid tolerance is further supported by eight genes contributing to low pH survival, including F0F1 ATP synthases (e.g., F0F1 ATP synthase subunit A-C cluster) and Na+/H+ antiporters (Nhac), critical for cytoplasmic pH regulation42. Its resilience to bile stress is indicated by genes such as ppaC (inorganic pyrophosphatase) and cfa (cyclopropane-fatty-acyl phospholipid synthase), mechanisms also noted in other bile-resistant Pediococcus species43,44.
P10’s adhesion capacity, a critical probiotic trait for gut colonization and host interaction45, is supported by seven putative adhesion-related genes (e.g., gpr, tuf, lspA). These findings are consistent with robust adhesion abilities observed in other Pediococcus strains46,47. Furthermore, P10 exhibits a strong defense against oxidative stress. Six of its nine oxidative stress-related genes encode components of the thioredoxin (tpx, trxA, trxB) and NADH antioxidant systems, crucial for reactive oxygen species (ROS) scavenging48. The presence of glutaredoxin (nrdH) and methionine sulfoxide reductase genes (msrA, msrB, msrC), coupled with its compensation for superoxide dismutase deficiency via manganese transport systems (mntH), further enhances its antioxidant capabilities, aligning with findings in other protective Pediococcus strains48,49,50. These comprehensive stress-response mechanisms underscore P10’s adaptive traits for the demanding gastrointestinal environment.
Regarding antibiotic resistance, P10’s profile aligns with the general susceptibility of P. acidilactici while exhibiting intrinsic resistance to glycopeptides, a characteristic attributed to peptidoglycan precursor modifications in Pediococcus species51. The identified resistance to specific drug classes like ciprofloxacin and colistin from genomic analysis was validated phenotypically. However, a notable phenotypic-genotypic discrepancy was observed for tetracycline, where genomic determinants were present, but the strain remained susceptible. This phenomenon, where the mere presence of a resistance gene does not always correlate with phenotypic resistance, has been reported in other lactic acid bacteria52,53 and highlights the complexity of gene expression and functional protein activity. Such discrepancies emphasize the importance of complementing genomic predictions with in vitro validation for a complete understanding of a strain’s resistance profile.
P10’s robust antimicrobial activity is largely attributed to its predicted bacteriocin production. Our BaPres analysis identified 57 potential bacteriocins, with a high probability of activity, aligning with the well-documented bacteriocin-producing capabilities of Pediococcus species35. Specifically, the presence of genes encoding plaA (prebacteriocin) and a putative entrocin P bacteriocin further supports its broad-spectrum antimicrobial potential, as these bacteriocins are known in other Pediococcus strains like P. pentosaceus54. The presence of these and other putative antimicrobial genes (bhtA, Col, truE3) suggests a diverse repertoire of pathogen inhibition mechanisms.
A particularly significant finding is the discovery of a complete and highly conserved Type II-A CRISPR-Cas system in P10, a strain isolated from chicken. This system, characterized by a prominent CRISPR array with 20 unique spacers and the presence of core cas genes (cas9, cas1, cas2, csn2), indicates a robust, active, and adaptive immune mechanism55,56. The direct evidence of its role in phage defense is compelling, with 17 of 20 spacers showing 100% identity to various bacteriophage genomes, predominantly unclassified Metagenome-Assembled Genomes (MAGs)57. This high homology confirms past encounters with phages prevalent in its natural habitat, such as the chicken gut, and underscores P10’s acquired immunity58. Type II-A CRISPR-Cas systems are widespread among probiotic lactic acid bacteria and are crucial for phage resistance in both industrial and gut environments59,60. The high integrity (100% repeat identity) and completeness of P10’s CRISPR array, coupled with the functional cas1 and cas2 genes ensuring continuous spacer acquisition, differentiate P10 as a highly adaptable strain capable of responding to evolving phage threats61. This represents a superior intrinsic defensive capability compared to other P. acidilactici strains where CRISPR-Cas systems may be incomplete or have disrupted cas9 genes59.
Beyond phage defense, the identification of a functional cas9 gene positions P. acidilactici P10 as a powerful tool for precision genome editing62. Leveraging an endogenous CRISPR-Cas9 system in a probiotic bacterium like P10 offers a significant advantage; unlike models often relying on exogenous CRISPR components for gene editing63, P10’s native system potentially streamlines genetic modifications for food and feed applications without foreign material. This inherent system could be harnessed to enhance probiotic traits, target undesirable genes (e.g., antibiotic resistance), or develop live biotherapeutics64,65,66. The apparently intact and highly conserved cas9 in P10 suggests it could be a more readily applicable platform for direct genetic manipulation compared to strains requiring extensive repair or optimization67.
Finally, the pan-genome analysis revealed 59 strain-specific genes unique to P10. KEGG29 pathway analysis showed these genes are predominantly involved in diverse metabolic pathways, including specialized carbohydrate utilization (e.g., PTS, K02757)68, amino acid and cofactor/vitamin biosynthesis69,70, and even “Plant-pathogen interaction” (K00864) [Ref 83]. These findings collectively point to P10’s distinct metabolic capabilities and adaptive advantages, potentially conferring a competitive edge in nutrient acquisition and environmental response within complex ecological niches. While these inferences are based on genomic annotation, they provide strong hypotheses for P10’s unique ecological niche and probiotic advantages. Future studies focusing on the expression and functional characterization of these strain-specific genes are crucial to fully elucidate their contribution to P10’s probiotic efficacy.
Material and methods
Isolation and preparation of bacterial suspension
Gastrointestinal tracts (n = 15) were collected from native broiler chickens (average weight 2,650 g) sourced from various regions across Iran. All animal handling procedures adhered to protocols approved by the Institute of Animal Care and Use Committee at the National Institute of Genetic Engineering and Biotechnology, aligning with relevant national animal welfare guidelines and regulations, and are reported in accordance with the ARRIVE guidelines. Samples were transported to the laboratory under sterile conditions. The intestinal contents were homogenized in physiological peptone solution. Subsequently, 10 mL of the homogenate was inoculated into 190 mL of de Man, Rogosa, and Sharpe (MRS) and Brain Heart Infusion (BHI) broths (Ibresco, Iran) for bacterial enrichment. Cultures were incubated at 37 °C with shaking at 150 rpm for 24 h. To isolate lactic acid bacteria (LAB), a loopful of each broth culture was plated onto MRS agar and incubated at 37 °C for 24 h. Isolates exhibiting distinct morphologies were subjected to initial screening for in vitro probiotic properties, including Gram staining and catalase tests. Gram-positive and catalase-negative bacteria were selected for further analysis. Among these isolates, P. acidilactici strain P10 demonstrated particularly promising probiotic characteristics, as detailed in subsequent sections.
Hemolytic test
Hemolytic activity of the isolate was determined on blood agar plates containing 5% (w/v) defibrinated sheep blood. Plates were incubated at 37 °C for 48 h, with Bacillus licheniformis PTCC 1525 serving as a positive control71.
Bile salt and acid tolerance
We assessed the isolate’s tolerance to bile salts as previously described by Sahro et al. (2001). A 0.5 McFarland microbial suspension was inoculated at 10% (v/v) into MRS broth containing 0.3% (w/v) Oxgall bile salts (Sigma-Aldrich, USA). Cultures were incubated at 37 °C for 8 h, with samples collected in triplicate at 0 and 8 h. Growth was monitored by measuring the optical density (OD) at 600 nm. Acid tolerance was evaluated following the method of Yu et al.72. A 1 mL microbial suspension (0.5 McFarland turbidity) was prepared and cultured in 9 mL of liquid MRS medium adjusted to pH values of 3, 4, and 5.8. Cultures were incubated at 37 °C in triplicate. After 3 h of incubation, 1 mL aliquots were taken, serially diluted, and 100 µL of each dilution was plated onto MRS agar in duplicate. Plates were incubated at 37 °C for 48 h. A viable count of not less than 106 CFU/mL was considered indicative of acid tolerance.
Evaluation of antimicrobial activity
The antagonistic activity of the isolated strain against a panel of pathogenic bacteria (E. coli ATCC 25922, S. enterica serovar Typhimurium ATCC 14028, S. aureus ATCC 25923, L. monocytogenes ATCC 119115, and P. aeruginosa ATCC 27853) was evaluated using the agar well diffusion method, as described by Jorgensen et al.73. Pathogenic strains were initially cultured in nutrient broth at 37 °C with shaking at 150 rpm. Subsequently, 100 μL of a 0.5 McFarland standard equivalent from the logarithmic growth phase of each pathogen was inoculated onto the surface of appropriate agar media. After a 30-min drying period, the inoculated plates were refrigerated. Four wells were then aseptically created on each plate, and 100 μL of cell-free supernatant (filtered with a 0.22 μm filter) from the selected probiotic strain P10 was added to each well. Lactobacillus salivarius S4 served as a positive control, while MRS medium was used as the negative control. For each pathogen, three replicate plates were prepared. Finally, the plates were incubated at 37 °C for 48 h.
Antibiotic sensitivity test
Antibiotic susceptibility testing of the potential probiotic strain P10 was performed using the disk diffusion method. A panel of antibiotics was employed, including ampicillin (10 μg), gentamicin (10 μg), streptomycin (10 μg), tetracycline (10 μg), erythromycin (15 μg), kanamycin (30 μg), vancomycin (30 μg), rifampin (5 μg), chloramphenicol (30 μg), penicillin (10 μg), tetracycline (10 μg), ciprofloxacin (5 μg), clindamycin (2 μg), tylosin (30 μg), sulfamethoxazole/trimethoprim (sultrim) (10 μg), fosfomycin (200 μg), enrofloxacin (10 μg), colistin (10 μg), and florfenicol (30 μg) (all obtained from Padtan Teb Co., Iran). The interpretation of inhibition zones was based on Clinical and Laboratory Standards Institute (CLSI) guidelines to categorize the strain as resistant, intermediate, or susceptible to each antibiotic.
Caco-2 adhesion test
For the adhesion assay, we used the human colon carcinoma cell line, Caco-2, obtained from the National Institute of Genetic Engineering and Biotechnology (NIGEB). Cells were cultured in high-glucose Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin solution. We maintained cultures in a humidified incubator at 37 °C with 5% CO₂ (DNA Biotech Co., Iran), refreshing the medium every other day until cells reached approximately 80% confluency. We grew the probiotic strain P10 in MRS broth, adjusting its concentration to a 0.5 McFarland standard using fresh DMEM to ensure a uniform bacterial inoculum density. Following trypsinization and cell counting, we seeded Caco-2 cells (4 × 105 cells/mL) onto a 24-well plate. Each well received 100 μL of the standardized bacterial suspension, and we incubated the plate at 37 °C for 3 h to allow bacterial adhesion. After incubation, we washed the cells twice with phosphate-buffered saline (PBS) to remove non-adherent bacteria. We then fixed the cells with methanol for further processing and performed crystal violet staining for 5 min to visualize and quantify the adherent bacteria. We also included the non-pathogenic E. coli strain ATCC 25922 as a control to assess P10’s adherence potential74.
Genomic DNA extraction and sequencing
We extracted the total genomic DNA of strain P10 using the QIAamp DNA Mini Kit (Qiagen, CA). DNA quality was subsequently measured with a NanoDrop™ spectrophotometer (Thermo Scientific NanoDrop 2000c). For molecular identification, we performed polymerase chain reaction (PCR) to amplify the 16S rRNA fragment using general primers 27F and 1492R. The amplified fragments were separated via horizontal electrophoresis and sequenced using the Sanger method. The sequence from the selected probiotic strain was then registered in global nucleotide databases (NCBI). Furthermore, the whole genome was sequenced using the Illumina HiSeq 2500 platform, generating 150 bp paired-end reads. Libraries were prepared from 1 mg of genomic DNA using the Nextera DNA Library Preparation Kit following the manufacturer’s protocol. Raw sequence quality control was performed with FastQC v0.11.3, followed by adapter and quality trimming via Trimmomatic 0.32.
Genome assembly and annotation
To identify the bacterial species and its closest related organism, we conducted phylogenetic analysis using the TYGS webserver (https://tygs.dsmz.de/). De novo assembly was performed with SPAdes v3.9.0, testing various k-mer sizes and combinations. We improved the genome assembly using Pilon v1.16 and evaluated the quality of each assembly with QUAST v3.2. Finally, assembled genome scaffolds were ordered by RagTag using the identified closest reference genome, P. acidilactici (GCF_013127755.1), from the phylogenetic analysis. A circular genomic map was then created from the resulting genome using the CG View server. Gene detection and genome annotation of the ordered assembled genome were performed using Prokka (https://github.com/tseemann/prokka/). We filtered and reported general genome features, such as rRNA and tRNA, using annotations provided by an in-house script. Additionally, functional annotation analysis was performed using eggnog v5.0 (http://eggnog5.embl.de/) and BlastKOALA v2.175.
Safety assessment
To identify potential antimicrobial resistance (AMR) genes within the P10 genome, in silico analysis was conducted utilizing three databases: ResFinder v4.5.0, CARD, and KEGG (Kyoto Encyclopedia of Genes and Genomes)29. Furthermore, ABRicate was employed to predict antibiotic resistance genes, and bacteriocin prediction was performed using BaPres (https://github.com/suraiya14/BaPreS).
Gene pathway analysis
We analyzed the predicted and annotated gene sequences to determine the existence of specific pathways. Complete identification of corresponding pathways was then achieved by manually inspecting assigned gene functions and comparing them to KEGG29 pathways.
CRISPR-Cas system identification and analysis
We identified CRISPR-Cas loci and associated cas genes in the genome of P. acidilactici P10, a strain isolated from chicken, using a two-pronged bioinformatic approach. Initially, CRISPRDetect v2.276 was employed to identify potential CRISPR arrays based on characteristic repeat-spacer structures. The identified CRISPR array was then subjected to further analysis using CRISPRCasFinder v4.2.1977 to classify the CRISPR-Cas system type and identify associated cas genes (e.g., cas1, cas2, cas9, csn2). To assess potential phage resistance, the sequences of identified spacers within the CRISPR array were extracted. These spacer sequences were subsequently used as queries in BLASTn searches against the NCBI RefSeq Viral Genomes database to identify homologous protospacer sequences in known bacteriophage genomes. Only hits with high identity (≥ 95%) and coverage (≥ 50%) were considered as putative targets.
Pan-genome analysis
To comprehensively investigate the genetic diversity and functional repertoire of the P10 draft genome and its closely related strains, we conducted a pan-genome analysis using Roary. A comprehensive search of the NCBI database initially identified numerous P. acidilactici genome entries. However, for the pan-genome analysis, we restricted our dataset to publicly available strains with complete genome sequences, resulting in 37 genomes from NCBI. Including the P10 genome, the final dataset for pan-genome analysis comprised 38 complete P. acidilactici genomes. This analysis enabled the identification of core, soft-core, and shell genes, providing valuable insights into the genomic diversity of this bacterial population. Furthermore, a phylogenetic tree was constructed based on the presence and absence of core and unique genes within this pan-genome dataset.
Whole genome comparisons
To elucidate the genomic variation among closely related P. acidilactici strains, we selected seven genomes with the highest sequence similarity to the P10 genome from our preliminary pan-genome analysis. Subsequently, we retrieved and compared several genomic features of these seven strains from the NCBI database, including the number of genes, tRNAs, rRNAs, genome size, and GC content. To visualize the comparative genomic structure, the seven selected P. acidilactici genomes were aligned and compared with the P10 draft assembly using BRIG 0.95 (http://brig.sourceforge.net/). Additionally, a progressive Mauve alignment was performed using the software pyGenomeViz (https://github.com/moshi4/pyGenomeViz) to facilitate a detailed comparison of the P10 genomic sequence with the reference strain P. acidilactici PMC65 (GCF_013127755.1).
Conclusion
Our study provides a comprehensive genomic and functional analysis of Pediococcus acidilactici strain P10, a novel isolate with promising probiotic attributes. The genome sequence revealed a compact circular chromosome harboring a diverse repertoire of genes crucial for its survival and beneficial activities, including those involved in carbohydrate metabolism, stress response, adhesion, and antimicrobial production. Phylogenetic analysis placed strain P10 within a distinct P. acidilactici clade, suggesting a shared evolutionary history and potentially similar ecological niches with closely related strains. Strain P10’s potential as a probiotic and for industrial applications is underscored by the presence of genes related to robust stress tolerance (against acid, cold, heat, and oxidative conditions), strong adhesion capabilities, and potent antimicrobial production (bacteriocins). Most notably, the genome revealed a sophisticated defense system, including a complete, active, and highly conserved Type II-A CRISPR-Cas system. This intrinsic phage resistance offers robust, adaptive immunity against ecologically relevant bacteriophages, a critical advantage for its viability and persistence as a probiotic, especially when compared to strains with less robust or incomplete defense mechanisms. Furthermore, the intact cas9 gene unlocks significant potential for precision genome editing, positioning P10 as a valuable strain for developing tailored microbial solutions in agriculture, food science, and health, potentially requiring fewer engineering steps than strains with compromised endogenous systems. The identification of 59 unique, strain-specific genes in P10, particularly those involved in diverse metabolic pathways and membrane transport, highlights its distinct metabolic capabilities and adaptive advantages. While this research significantly contributes to our understanding of the genomic and functional diversity within the Pediococcus genus, further in-depth functional characterization of the predicted genes, especially those currently classified with unknown functions, is essential to fully elucidate the metabolic and physiological potential of P. acidilactici P10. Ultimately, these findings pave the way for future applications in food fermentation, probiotic development, and other biotechnological endeavors.
Data availability
To ensure data accessibility and reproducibility, the assembled genome and the 16S rRNA gene sequence (MG255949.1) are available through the NCBI Sequence Read Archive (SRA) under BioProject accession number PRJNA1243677. This comprehensive dataset allows researchers to replicate our analyses and conduct further genomic investigations.
References
-
Yang, S. J., Kim, K.-T., Kim, T. Y. & Paik, H.-D. Probiotic properties and antioxidant activities of Pediococcus pentosaceus SC28 and Levilactobacillus brevis KU15151 in fermented black gamju. Foods 9, 1154 (2020).
-
Hill, C. et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514 (2014).
-
Begum, J., Buyamayum, B., Lingaraju, M. C., Dev, K. & Biswas, A. Probiotics: Role in immunomodulation and consequent effects: Probiotics and immunity. Lett. Anim. Biol. 1, 1–6 (2021).
-
Samot, J. & Badet, C. Antibacterial activity of probiotic candidates for oral health. Anaerobe 19, 34–38 (2013).
-
Śliżewska, K., Markowiak-Kopeć, P. & Śliżewska, W. The role of probiotics in cancer prevention. Cancers 13, 20 (2020).
-
Homayouni Rad, A., Pourjafar, H. & Mirzakhani, E. A comprehensive review of the application of probiotics and postbiotics in oral health. Front. Cell. Infect. Microbiol. 13, 1120995 (2023).
-
Alayande, K. A., Aiyegoro, O. A. & Ateba, C. N. Probiotics in animal husbandry: Applicability and associated risk factors. Sustainability 12, 1087 (2020).
-
Anee, I. J., Alam, S., Begum, R. A., Shahjahan, R. M. & Khandaker, A. M. The role of probiotics on animal health and nutrition. J. Basic Appl. Zool. 82, 1–16 (2021).
-
Mahesh, M. S., Mohanta, R. K. & Patra, A. K. Probiotics in livestock and poultry nutrition and health. In Advances in Probiotics for Sustainable Food and Medicine 149–179 (2021).
-
Lambo, M. T., Chang, X. & Liu, D. The recent trend in the use of multistrain probiotics in livestock production: An overview. Animals 11, 2805 (2021).
-
Jiang, S., Cai, L., Lv, L. & Li, L. Pediococcus pentosaceus, a future additive or probiotic candidate. Microb. Cell Fact. 20, 1–14 (2021).
-
Qi, Y. et al. Pediococcus pentosaceus: scrEening and application as probiotics in food processing. Front. Microbiol. 12, 762467 (2021).
-
Pot, B. et al. The genus Lactobacillus. Lact. acid Bact. Biodivers. Taxon. 249–353 (2014).
-
Song, A.A.-L., In, L. L. A., Lim, S. H. E. & Rahim, R. A. A review on Lactococcus lactis: From food to factory. Microb. Cell Fact. 16, 1–15 (2017).
-
Hemme, D. & Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 14, 467–494 (2004).
-
Porto, M. C., Kuniyoshi, T. M., Azevedo, P. O., Vitolo, M. & Oliveira, R. D. Pediococcus spp.: An important genus of lactic acid bacteria and pediocin producers. Biotechnol. Adv. 35, 361–374 (2017).
-
Shokryazdan, P., Faseleh Jahromi, M., Liang, J. B. & Ho, Y. W. Probiotics: From isolation to application. J. Am. Coll. Nutr. 36, 666–676 (2017).
-
Song, Y.-R., Lee, C.-M., Lee, S.-H. & Baik, S.-H. Evaluation of probiotic properties of Pediococcus acidilactici M76 producing functional exopolysaccharides and its lactic acid fermentation of black raspberry extract. Microorganisms 9, 1364 (2021).
-
Pérez-Ramos, A., Mohedano, M. L., Pardo, M. Á. & López, P. β-Glucan-producing Pediococcus parvulus 2.6: Test of probiotic and immunomodulatory properties in zebrafish models. Front. Microbiol. 9, 1684 (2018).
-
Snauwaert, I., Stragier, P., De Vuyst, L. & Vandamme, P. Comparative genome analysis of Pediococcus damnosus LMG 28219, a strain well-adapted to the beer environment. BMC Genomics 16, 1–12 (2015).
-
Ayyash, M. et al. Physicochemical, bioactive and rheological properties of an exopolysaccharide produced by a probiotic Pediococcus pentosaceus M41. Carbohydr. Polym. 229, 115462 (2020).
-
Wanna, W., Surachat, K., Kaitimonchai, P. & Phongdara, A. Evaluation of probiotic characteristics and whole genome analysis of Pediococcus pentosaceus MR001 for use as probiotic bacteria in shrimp aquaculture. Sci. Rep. 11, 18334 (2021).
-
Cabello-Olmo, M. et al. Pediococcus acidilactici pA1c® improves the beneficial effects of metformin treatment in type 2 diabetes by controlling glycaemia and modulating intestinal microbiota. Pharmaceutics 15, 1203 (2023).
-
Yavorov-Dayliev, D. et al. Glucose-lowering effects of a synbiotic combination containing Pediococcus acidilactici in C. elegans and mice. Diabetologia 66, 2117–2138 (2023).
-
Aziz, T. et al. Genomic profiling of Pediococcus acidilactici BCB1H and identification of its key features for Biotechnological innovation, food technology and medicine. Sci. Rep. 15, 6050 (2025).
-
Lu, K., Wang, X., Zhou, Y. & Zhu, Q. Genomic characterization and probiotic potential assessment of an exopolysaccharide-producing strain Pediococcus pentosaceus LL-07 isolated from fermented meat. BMC Microbiol. 24, 142 (2024).
-
Zhao, M. et al. Probiotic characteristics and whole-genome sequence analysis of Pediococcus acidilactici isolated from the feces of adult beagles. Front. Microbiol. 14, 1179953 (2023).
-
Li, Z. et al. Comparative genomics analysis of Pediococcus acidilactici species. J. Microbiol. 59, 573–583 (2021).
-
Kanehisa, M., Furumichi, M., Sato, Y., Matsuura, Y. & Ishiguro-Watanabe, M. KEGG: Biological systems database as a model of the real world. Nucleic Acids Res. 53, D672–D677 (2025).
-
Li, O. et al. Complete genome sequence and probiotic properties of Lactococcus petauri LZys1 isolated from healthy human gut. J. Med. Microbiol. 70, 1397 (2021).
-
Song, H. et al. Metagenomic analysis of the gut microbiota of wild mice, a newly identified reservoir of Campylobacter. Front. Cell. Infect. Microbiol. 10, 596149 (2021).
-
Sun, Y. et al. Assessments of probiotic potentials of Lactiplantibacillus plantarum strains isolated from Chinese traditional fermented food: Phenotypic and genomic analysis. Front. Microbiol. 13, 895132 (2022).
-
Liu ChenJian, L. C. et al. Complete genome sequences and comparative genome analysis of Lactobacillus plantarum strain 5–2 isolated from fermented soybean (2015).
-
Gao, Y. et al. Physiological function analysis of Lactobacillus plantarum Y44 based on genotypic and phenotypic characteristics. J. Dairy Sci. 103, 5916–5930 (2020).
-
Altuntaş, E. G., Ayhan, K., Peker, S., Ayhan, B. & Demiralp, D. Ö. Purification and mass spectrometry based characterization of a pediocin produced by Pediococcus acidilactici 13. Mol. Biol. Rep. 41, 6879–6885 (2014).
-
Diep, D. B., Godager, L., Brede, D. & Nes, I. F. Data mining and characterization of a novel pediocin-like bacteriocin system from the genome of Pediococcus pentosaceus ATCC 25745. Microbiology 152, 1649–1659 (2006).
-
Pérez-Ramos, A. et al. Draft genome sequence of Pediococcus parvulus 2.6, a probiotic β-glucan producer strain. Genome Announc. 4, 10–1128 (2016).
-
Rocha, E. P. C. & Danchin, A. Base composition bias might result from competition for metabolic resources. TRENDS Genet. 18, 291–294 (2002).
-
Sharifi, S. et al. Integrated co-expression analysis of regulatory elements (miRNA, lncRNA, and TFs) in bovine monocytes induced by Str. uberis. Sci. Rep. 13, 15076 (2023).
-
Novick, R. P. Pathogenicity and other genomic islands (2013).
-
Jia, Y., Huang, C., Mao, Y., Zhou, S. & Deng, Y. Screening and constructing a library of promoter-5′-UTR complexes with gradient strength in Pediococcus acidilactici. ACS Synth. Biol. 12, 1794–1803 (2023).
-
Zhang, M. et al. Proteomic profiling and stress response in Pediococcus acidilactici under acetic acid. J. Agric. Food Chem. 70, 12708–12721 (2022).
-
Escobar-Sánchez, M. et al. Probiotic properties and proteomic analysis of Pediococcus pentosaceus 1101. Foods 12, 46 (2022).
-
Jaiswal, S., Pradhan, S. N., Jain, D., Dhassiah Peter, M. P. & Antony, U. Probiotic and functional characterization of Pediococcus acidilactici isolated from Bhaati jaanr, traditional fermented rice porridge. Appl. Biochem. Biotechnol. 194, 5734–5747 (2022).
-
Monteagudo-Mera, A., Rastall, R. A., Gibson, G. R., Charalampopoulos, D. & Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 103, 6463–6472 (2019).
-
Castex, M., Lemaire, P., Wabete, N. & Chim, L. Effect of probiotic Pediococcus acidilactici on antioxidant defences and oxidative stress of Litopenaeus stylirostris under Vibrio nigripulchritudo challenge. Fish Shellfish Immunol. 28, 622–631 (2010).
-
Zhang, H., Liu, J., Wen, R., Chen, Q. & Kong, B. Metabolomics profiling reveals defense strategies of Pediococcus pentosaceus R1 isolated from Harbin dry sausages under oxidative stress. LWT 135, 110041 (2021).
-
Zhang, H., Zhang, C., Liu, H., Chen, Q. & Kong, B. Proteomic response strategies of Pediococcus pentosaceus R1 isolated from Harbin dry sausages to oxidative stress. Food Biosci. 44, 101364 (2021).
-
Wang, Y. et al. Pediococcus pentosaceus PP04 ameliorates high-fat diet-induced hyperlipidemia by regulating lipid metabolism in C57BL/6N mice. J. Agric. Food Chem. 68, 15154–15163 (2020).
-
Bergsveinson, J., Ewen, E., Friesen, V. & Ziola, B. Transcriptional activity and role of plasmids of Lactobacillus brevis BSO 464 and Pediococcus claussenii ATCC BAA-344T during growth in the presence of hops. AIMS Microbiol. 2, 460–478 (2016).
-
Sarkar, P., Yarlagadda, V., Ghosh, C. & Haldar, J. A review on cell wall synthesis inhibitors with an emphasis on glycopeptide antibiotics. Medchemcomm 8, 516–533 (2017).
-
Chokesajjawatee, N. et al. Safety assessment of a nham starter culture Lactobacillus plantarum BCC9546 via whole-genome analysis. Sci. Rep. 10, 10241 (2020).
-
Kandasamy, S. et al. Probiogenomic in-silico analysis and safety assessment of Lactiplantibacillus plantarum DJF10 strain isolated from Korean raw milk. Int. J. Mol. Sci. 23, 14494 (2022).
-
Nes, I. F., Diep, D. B. & Holo, H. Bacteriocin diversity in Streptococcus and Enterococcus. J. Bacteriol. 189, 1189–1198 (2007).
-
Jiang, W. & Marraffini, L. A. CRISPR-Cas: New tools for genetic manipulations from bacterial immunity systems. Annu. Rev. Microbiol. 69, 209–228 (2015).
-
Barrangou, R. & Marraffini, L. A. CRISPR-Cas systems: Prokaryotes upgrade to adaptive immunity. Mol. Cell 54, 234–244 (2014).
-
Achudhan, A. B. & Saleena, L. M. Genomic reconstruction of unclassified microorganisms: Analysis of CRISPR arrays and genes involved in defense mechanisms. Gene 928, 148808 (2024).
-
Rocha, E. P. C. & Bikard, D. Microbial defenses against mobile genetic elements and viruses: Who defends whom from what?. PLoS Biol. 20, e3001514 (2022).
-
Xin, Y., Guo, T. & Qiao, M. Current application and future prospects of CRISPR-Cas in lactic acid Bacteria: A review. Food Res. Int. 209, 116315 (2025).
-
Rostampour, M., Panahi, B. & Masoumi Jahandizi, R. The CRISPR-Cas system in Lactiplantibacillus plantarum strains: Identification and characterization using a genome mining approach. Front. Microbiol. 15, 1394756 (2024).
-
Panahi, B., Dehganzad, B. & Nami, Y. CRISPR-Cas systems feature and targeting phages diversity in Lacticaseibacillus rhamnosus strains. Front. Microbiol. 14, 1281307 (2023).
-
Doudna, J. A. & Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 346, 1258096 (2014).
-
Makarova, K. S. et al. An updated evolutionary classification of CRISPR–Cas systems. Nat. Rev. Microbiol. 13, 722–736 (2015).
-
Selle, K. & Barrangou, R. Harnessing CRISPR–Cas systems for bacterial genome editing. Trends Microbiol. 23, 225–232 (2015).
-
Culligan, E. P., Hill, C. & Sleator, R. D. Probiotics and gastrointestinal disease: Successes, problems and future prospects. Gut Pathog. 1, 1–12 (2009).
-
Roberts, A. & Barrangou, R. Applications of CRISPR-Cas systems in lactic acid bacteria. FEMS Microbiol. Rev. 44, 523–537 (2020).
-
Liu, L. et al. High-efficiency genome editing based on endogenous CRISPR-Cas system enhances cell growth and lactic acid production in Pediococcus acidilactici. Appl. Environ. Microbiol. 87, e00948-e1021 (2021).
-
Dong, Z. et al. Core antibiotic resistance genes mediate gut microbiota to intervene in the treatment of major depressive disorder. J. Affect. Disord. 363, 507–519 (2024).
-
Lebeer, S., Vanderleyden, J. & De Keersmaecker, S. C. J. Host interactions of probiotic bacterial surface molecules: comparison with commensals and pathogens. Nat. Rev. Microbiol. 8, 171–184 (2010).
-
Rossi, M., Amaretti, A. & Raimondi, S. Folate production by probiotic bacteria. Nutrients 3, 118–134 (2011).
-
Huligere, S. S. et al. Isolation and characterization of lactic acid bacteria with potential probiotic activity and further investigation of their activity by α-amylase and α-glucosidase inhibitions of fermented batters. Front. Microbiol. 13, 1042263 (2023).
-
Yu, Z. et al. Evaluation of probiotic properties of Lactobacillus plantarum strains isolated from Chinese sauerkraut. World J. Microbiol. Biotechnol. 29, 489–498 (2013).
-
Jorgensen, J. H. & Turnidge, J. D. Susceptibility test methods: dilution and disk diffusion methods. In Manual of Clinical Microbiology 1253–1273 (2015).
-
Bazireh, H., Shariati, P., Azimzadeh Jamalkandi, S., Ahmadi, A. & Boroumand, M. A. Isolation of novel probiotic Lactobacillus and Enterococcus strains from human salivary and fecal sources. Front. Microbiol. 11, 597946 (2020).
-
Kanehisa, M., Sato, Y. & Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 428, 726–731 (2016).
-
Biswas, A., Staals, R. H. J., Morales, S. E., Fineran, P. C. & Brown, C. M. CRISPRDetect: A flexible algorithm to define CRISPR arrays. BMC Genomics 17, 1–14 (2016).
-
Couvin, D. et al. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 46 W246–W251 (2018).
Acknowledgment
This study has been approved by the National Institute of Genetic Engineering and Biotechnology under project number 764.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Tabashiri, R., Mahmoodian, S., Pakdel, M.H. et al. Comprehensive in vitro and whole-genome characterization of probiotic properties in Pediococcus acidilactici P10 isolated from Iranian broiler chicken. Sci Rep 15, 28953 (2025). https://doi.org/10.1038/s41598-025-13470-w
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-13470-w