
Introduction
The most common cancer in women is breast cancer (BC). While most BC cases are benign and can be treated with surgery, 25% have a latent, insidious nature that causes them to grow slowly but spread rapidly. Current treatments significantly slow the growth of the tumors, but they recur, resulting in high mortality rates1,2. According to recent data, 40% of BC patients die from metastasis, even when they receive standard treatment. The essential characteristic of triple-negative BC is its propensity for metastasis, which has a very complicated mechanism3,4,5. In healthy human cells, the release and consumption of oxygen are strictly regulated. Hypoxia is caused by increased O2 consumption resulting from enhanced cancer cell proliferation. Cancer cells are characterized by dysregulated cell proliferation and physiologically abnormal blood vessels due to extreme hypoxia6,7. Hypoxia makes traditional cancer treatments, such as prevalent chemotherapy drugs, ineffective and also hamper medication delivery. Because the hypoxic cells are located deep in the center of the tumor, beyond blood vessels, they cannot be reached by NPs or by therapies that are routinely administered8,9. To overcome these problems, drugs that can destroy cancer cells and NPs that penetrate the deep layer of cancer cells can be deployed10.
Tirapazamine (TPZ; SR-4233), which acts under hypoxic conditions, is one of the clinical small molecule prodrugs against tumor hypoxia. Under hypoxia, TPZ is activated to form an extremely toxic free radical that damages DNA and eventually kills cells11. The harmful free radicals reoxidize to their inactive form when oxygen is present. TPZ exhibits a higher cytotoxic effect in the hypoxic regions of tumors due to its hypoxia-selective nature12. It is shown that TPZ has a hypoxia-selective effect in both in vitro and in vivo surveys. Research has demonstrated that TPZ can prevent DNA replication by inhibiting topoisomerase II activity under hypoxic conditions and by directly damaging DNAss12,13.
Computational approaches, particularly molecular docking and protein–protein interaction network analysis, have become invaluable tools for understanding drug mechanisms and optimizing delivery systems14. For hypoxia-activated drugs like TPZ, in silico analysis reveals critical protein interactions and binding mechanisms that guide therapeutic strategies. Understanding TPZ’s interactions with hypoxia-inducible factor 1-alpha (HIF1A) and related pathway proteins through computational modeling provides crucial insights into its selective activation under hypoxic conditions15. These molecular-level insights can guide the design of more effective drug delivery systems, particularly in selecting appropriate nanocarrier combination enhance drug targeting and efficacy16. By combining computational predictions with experimental validation, more rational and effective therapeutic approaches can be developed for hypoxic tumors17.
Nowadays, the field of nanotechnology is expanding, and most pharmaceutical researchers have adopted drug delivery methods in the form of nanoparticles (NPs), especially in the field of cancer research for targeted purposes. Lipid-based NPs (liposomes18, niosomes19), micells20, non-organic NPs21,22, hydrogels23,24,25,26,27, chitosan1,28,29,30, DNA nanostructure31,32 and dendrimers33,34 are some examples of nanosystems used for therapeutic purposes. The purpose of these NPs system is to enhance drug targets, enable drugs to escape the reticuloendothelial system, and reduce the side effects of drugs10,35,36,37. In recent decades, the number of dual-loaded NPs systems has increased to allow for the simultaneous release of co-encapsulated anticancer agents and chemosensitizers upon degradation or upon external cues and to improve uptake and penetration to eradicate tumors and overcome multidrug-resistant cancers38,39. To improve bioavailability and overcome drug synergy limitations, dual NP systems with sequential delivery of drug combinations have been utilized to treat cancer40,41,42. Dendrimers are highly branched, three-dimensional molecules, ranging in size from 1 to 10 nm, and their ability to enter cells with high efficiency due to numerous of amine groups is one of their unique features. Among the initial families of dendrimers to be thoroughly studied, manufactured, and marketed was PAMAM. PAMAM dendrimers are widely employed in research on targeted therapies and genetic material delivery because of their extremely symmetrical structure, which allows for multiple modifications to be made to their core and surface groups43.
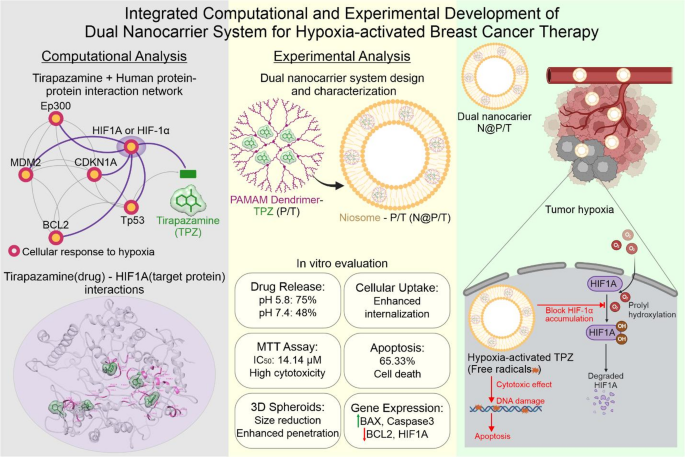
In the current study, we first employed computational studies to investigate the interaction of TPZ with HIF1A. Then, in the experimental section, TPZ was loaded in PAMAM (P/T) to improve the effects of P/T, such as preventing the excretion by the kidney, increasing the amount of P/T to inter the cancer cells, reducing the side effect of PAMAM, and overcoming the hydrophobic problem of P/T, and P/T itself was loaded in niosome (NIO) to realize a double-package nano drug delivery system (NDDS) (Fig. 1). NIOs are vesicular systems resembling liposomes obtained from the hydration of synthetic nonionic surfactants with the addition of cholesterol. They are non-immunogenic, biodegradable, and biocompatible. And are extensively studied for controlled release and targeted delivery in the treatment of cancer. NIO can entrap both hydrophilic and lipophilic drugs and prolong the circulation of the entrapped drug in the body19,44.

Concept design of double package niosome-dendrimer platform for Tirapazamine delivery to hypoxic breast cancer. From left to right, computational studies, major results and working principle of N@P/T. Figure is drawn by Corel DRAW version 2024v.25.2.1.313.
Thus, we designed and synthesized a dual NP delivery system for hypoxia-activated prodrug–Tirapazamine- delivery to MDA-MB-231 mammospheres. In our design, niosome can encapsulate high concentrations of small-sized cationic PAMAM G5 loaded with TPZ. Upon localization in acidic tumor microenvironment, niosomes decompose and release P/T which can swiftly penetrate into the cell membrane and deliver TPZ to the hypoxic region of tumor spheroids. Under hypoxic conditions, TPZ can be reduced into BTZ and generate toxic free radicals to kill cancer cells (Fig. 1).
Materials and methods
In silico analysis of TPZ interactions
(A) Protein–protein interaction network analysis
The STITCH database (Search Tool for Interactions of Chemicals, version 545) was employed to investigate the interactions between TPZ and human proteins. This database integrates multiple lines of evidence from experimental data, curated databases, and text mining to predict chemical-protein interactions. The UniProt database (the world’s leading comprehensive and freely accessible resource for protein research46), was used for protein annotation and gene ontology analysis of the identified proteins. Protein–protein interaction networks were visualized using Cytoscape v3.10.3 software47, with subsequent pathway analysis performed using the KEGG database (a comprehensive database for pathway research48) to identify BC-related pathways. Network centrality including degree was calculated using the CytoHubba49 plugin of Cytoscape software.
(B) Molecular docking analysis
The molecular interactions between TPZ and Hypoxia-inducible factor 1-alpha (HIF1A or HIF1A_HUMAN: UniProt ID Q16665) were investigated using COACH-D web-server (a template-based algorithm for protein–ligand binding site prediction50). The 3D structure of TPZ was obtained from the DrugBank database (Drug ID DB04858). Due to limited availability of complete HIF1A crystal structures (highest coverage PDB: 4H6J-Chain A, 13% coverage), the full 3D structure was modeled using I-TASSER51 and refined using GalaxyRefine52. Model quality was validated using PROCHECK53, ERRAT54, and ProSA-web55 web servers. Molecular interactions were analyzed using Chimera version 1.17.156, the best-docked poses with the highest C score were selected for visualization.
Materials
Cholesterol, Span 60, tween 80, and MTT were purchased from Sigma-Aldrich (St. Louis, MO, USA). Ethanol, methanol, and chloroform were obtained from Merck Co. MDA-MB-231 cell line was purchased from the National Cell Bank of Iran, Pasteur Institute (Tehran, Iran). Fetal bovine serum (FBS) and RPMI 1640 medium were provided by Gibco. Annexin V-FITC apoptosis detection kit was obtained by eBiosciences (MA, USA). PAMAM G5 dendrimer, FITC and TPZ were purchased from Sigma.
Synthesis of NPs
NIO was synthesized by the thin-film hydration method according to our previous works57,58,59. Briefly, Tween 80 (16 mg), cholesterol (8 mg), Span 60 (10 mg), and P/T (1 mg)33 were dissolved in organic solvents (chloroform (6 mL) and methanol (6 mL)). Next, the mixture was placed on a rotary evaporator (120 rpm, 60 °C, 1 h), and the solvents were evaporated, forming a thin film layer (pro-niosomes). Then, 10 mL of PBS was added to form N@P/T. A probe sonicator was used (for 8 min) to ensure uniformity of the N@P/T.
Characterization of the NPs
The size, surface charge, and shape of NPs were characterized using transmission electron microscopy (TEM), dynamic light scattering (DLS), and scanning electron microscopy (SEM). For TEM, a drop of the NP suspension was placed on a carbon-coated grid, dried, and analyzed at 200 kV. DLS analysis involved diluting the suspension in deionized water and measuring hydrodynamic size, polydispersity index (PDI), and zeta potential using a 633 nm laser. SEM imaging, conducted at 15 kV after gold-coating dried samples, provided complementary morphological data. Fourier transform infrared (FT-IR) spectroscopy confirmed TPZ encapsulation and identified functional group interactions by analyzing KBr pellets over the range of 400–4000 cm⁻1. These techniques collectively evaluated the physicochemical properties of NPs, ensuring their suitability for drug delivery applications while assessing size, stability, morphology, and structural integrity.
Assessment of encapsulation efficiency (EE) and drug loading (DL) in the N@P/T
To remove the unloaded P/T and assess the amount of P/T successfully loaded into N@P/T, a dialysis membrane (molecular weight cut-off of 12,000 Da) was used. Briefly, 10 mL of the N@P/T suspension was placed inside the dialysis membrane, which was then immersed in a container containing 70 mL of PBS. The container was kept on ice and stirred magnetically at 90 rpm for 30 min to ensure efficient separation of unloaded drug.
The concentration of the unloaded drug was subsequently determined using UV–Vis spectroscopy at a wavelength of 415 nm. A calibration curve was constructed to quantify the drug concentration based on absorbance values. The following formula was used to calculate the drug loading efficiency:
$$Encapsulation;Efficiencyleft( {EE} right),left( % right), = ,Mass,of,TPZ,in,N@P/Total,TPZ , mass, times ,100.$$
$$Drug;Loadingleft( {DL} right),left( % right), = ,Mass,of , TPZ,in,N@P/Total , N@P , mass, times ,100.$$
The drug release
In the beginning, the N@P/T was moved to two fixed dialysis bags. PBS (pH = 7.4) and PBS (pH = 5.8) were applied to simulate normal and tumor conditions, respectively. Containers were filled with 70 mL of buffer solutions with pH 7.4 and 5.8, and the dialysis bags were immersed in the buffers. Containers were then shaken (120 rpm) at 37 °C. At various intervals, 2 mL of buffers were replaced with 2 mL of fresh buffers. UV–Vis spectrophotometer was used at 415 nm to determine the release pattern. To assess the release curves, several kinetic models were applied60.
Uptake assessment
Niosome loaded with FITC (N@FITC) was used for uptake studies. The procedure was similar to niosome synthesis, except that P/T was replaced with FITC (1 mg) as a fluorescent probe. MDA-MB-231 cell lines were seeded at a density of 5.0 × 107 cells per well in six-well plates. Cells were treated with N@FITC, and after 4 h, cells were trypsinized, washed, and analyzed by flow cytometry (Becton Dickinson, San Jose, CA, USA).
MTT assay
The cytotoxicity of P/T, NIO, and N@P/T on the MDA-MB-231 cells was assessed using the MTT assay. Briefly, cells were seeded at a density of 5.0 × 103 in a 96-well plate. The next day the cells were treated with various doses of TPZ, NIO, and N@P/T and incubated for 48 h. After that, MTT solution was added to each well and then incubated for 4 h. The MTT solution was then removed and DMSO was added, and then the plate was mixed by thorough pipetting. Absorbance was measured at 570 nm using a microplate reader (BioTek Instruments, Winooski, USA) to determine cell viability.
Apoptosis/necrosis assessment
For this, MDA-MB231, cells were seeded at a density of 5.0 × 107 cells per well in a 6-well plate, and after 24 h treated with P/T and N@P/T (25μM). Next 48 h were trypsinized, washed, and then resuspended in 2μL binding buffer. Then, Annexin V-FITC (2μL), and PI (2μL) were sequentially added and kept in the dark for 15 min. Finally, cells were washed, and resuspended in binding buffer and analyzed by flow cytometry (FACS flow cytometry, BD FACSCalibur, BD bioscience, CA, USA).
Spheroid size test
In line with our previous works11,30,33,61, to examine how NPs affect the size of spheroids, a 3D culture was created using the liquid overlay technique (LOT). In brief, 50 μL of a 1.5% agarose solution dissolved in PBS was added to each well of a 96-well plate and then autoclaved. The plate was left at room temperature for 30 min to allow the agarose gel to solidify completely. Next, 5.0 × 103 MDA-MB-231 cells were added to each well and centrifuged at 1200 rpm for 5 min to concentrate the cells in the center of the well. The cells were then placed in an incubator for two days to allow them to interact and form spheroids. Subsequently, the spheroids were exposed to different concentrations of N@P/T. The size of the spheroids was then assessed and examined by Cell A at specific time points, namely 24, 48, and 72 h after the treatment.
qRT real time PCR
To evaluate the expression levels of BCL2, Bax, and caspase-3 mRNA, reverse transcription-quantitative polymerase chain reaction (RT-qPCR) was conducted2,62. For this experiment, 3D cultures of the MDA-MB-231 cell line were established. Afterward, the cells were treated with TPZ and N@P/T (25 μM) for a duration of 48 h. The resulting spheroids from each cell line were collected in a falcon tube and rinsed twice with PBS. The spheroids were then subjected to trypsinization and transferred to an incubator for 100 s, followed by neutralization with a complete culture medium. Subsequently, the spheroids were pipetted to obtain individual cells, which were subsequently centrifuged at 160 g for 10 min.
Based on our previous works30,33, we also evaluated the expression levels of HIF1A in MDA-MB-231 spheroids under normoxia (21% O2 level) and hypoxia condition (1% O2 concentration) after 24h treatment with N@P/T (25 μM) using a hypoxia incubator (Memmert, Germany).
Total RNA was extracted using a Trizol reagent. The concentration of the RNA was determined using a Nanodrop spectrophotometer. Following that, cDNA synthesis was performed using the Two-Step 2 × RT-PCR Pre-mix Taq and random hexamer primers, as directed by the instructions provided by the manufacturer. Gene expression analysis was carried out using SYBR® 2 × Real-Time PCR Master Mix High ROX on an AB StepOne Plus™ instrument. The temperature profiles for PCR included an initial denaturation step at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s, 60 °C for 30 s, and 72 °C for 30 s. The threshold cycle of the human GAPDH housekeeping gene was used as a reference for normalization. Fold changes in gene expression were determined using the Pfaffl (2−ΔΔCt) method. The primer sequences used for PCR amplification is reported elsewhere33,58,61.
Statistical analysis
ANOVA and the Tukey post-HOC test were applied to evaluate the mean difference among groups. GraphPad Prism software version 9.3. p values ˂0.05 mean difference tests were considered significant. All experiments were repeated at least twice in triplicates.
Results and discussion
Computational analysis of TPZ’s molecular interactions and network topology
(A) Protein–protein interaction network analysis reveals TPZ’s multi-target effects
The protein–protein interaction network analysis revealed significant interactions between TPZ and several key proteins involved in hypoxia response and cancer pathways. As shown in Fig. 2B, HIF1A emerged as a central node in the interaction network, showing direct connections with multiple proteins including EP300, CDKN1A, and TP53. The network analysis identified three major functional clusters:
- 1.
Cancer pathways (KEGG hsa05200): Including BAX, BCL2, BCL2L11, BRCA1, CASP3, CDKN1A, CDKN2A, EGLN1, EP300, HIF1A, MDM2, TP53, VHL
- 2.
HIF-1 signaling pathway (KEGG hsa04066): Comprising HIF1A and its direct interactors
- 3.
Cellular response to hypoxia (GO:0071456): Including key proteins like BCL2, EGLN1, HIF1A, MDM2, SIRT1, VHL

Molecular docking analysis of TPZ-HIF1A interactions and protein–protein interaction network. (A) Predicted binding sites between TPZ and Hypoxia-inducible factor 1-alpha (HIF1A). The refined 3D structure of HIF1A is shown in gray representation, with predicted binding residues highlighted in pink. Five distinct binding sites were identified: two primary sites (Cscore 1.0, cluster size 3), one intermediate site (Cscore 1.0, cluster size 2), and two additional sites (Cscore 1.0-0.96, cluster size 1). TPZ molecules are shown in green surface representation in their respective binding poses. (B) Protein–protein interaction network analysis of TPZ with human proteins. Nodes are colored according to pathway involvement: yellow nodes represent proteins involved in cancer pathways (KEGG hsa05200), grey nodes indicate proteins without specific cancer pathway assignment, and red-outlined nodes show proteins involved in HIF-1 signaling pathway (KEGG hsa04066) and cellular response to hypoxia (GO:0071456). Numbers in parentheses indicate each protein node’s degree parameter, representing their network connectivity. Purple lines highlight direct interactions between HIF1A and network proteins. www.kegg.jp/kegg/kegg1.html.
The protein–protein interaction network analysis revealed a complex interaction pattern between TPZ and various proteins involved in hypoxic response and cancer pathways. The network topology analysis highlighted varying degrees of connectivity among nodes, quantified by degree parameters (shown in parentheses next to each protein in Fig. 2B):
- 1.
High-degree nodes (≥ 10):
-
TP53 (14): The highest degree of connectivity, indicating its central role in the network
-
HIF1A (12): Second-highest connectivity, supporting its role as a primary target
-
CDKN1A (12): Equal connectivity to HIF1A
-
EP300 (11): High connectivity suggests an important regulatory role
-
BCL2 (11): Key mediator in apoptotic pathways
-
MDM2 (10): Critical regulator of p53 pathway
-
- 2.
Medium-degree nodes (7–9):
-
ATM (9): DNA damage response regulator
-
SIRT1 (8): Metabolic regulator
-
CDKN2A (8): Cell cycle regulator
-
BRCA1 (8): DNA repair protein
-
BAX (8): Apoptosis regulator
-
KAT2B (7): Histone acetyltransferase
-
- 3.
Low-degree nodes (≤ 6):
-
VHL (6): Hypoxia pathway regulator
-
CASP3 (6): Apoptosis executor
-
BCL2L11 (4): Pro-apoptotic protein
-
EGLN1 (2): Lowest connectivity, specific to oxygen sensing
-
The degree distribution analysis reveals that the network follows a scale-free topology, typical of biological networks, where a few hub proteins (particularly TP53, HIF1A, and CDKN1A) maintain the majority of connections. This architecture suggests that TPZ’s effects are primarily mediated through these high-degree nodes, with subsequent cascade effects through their respective pathways.
The integration of degree parameters with pathway assignments demonstrates that proteins with high degree centrality tend to be involved in multiple pathways, particularly those related to hypoxia response and cancer signaling. For example, HIF1A (degree 12) participates in both hypoxia response and cancer pathways, while TP53 (degree 14) serves as a master regulator connecting multiple pathways.
The presence of both high and low-degree nodes in hypoxia-related pathways suggests a hierarchical organization where TPZ’s effects are channeled through major regulators (high-degree nodes) to more specific effectors (low-degree nodes). This network structure supports the drug’s selective activation under hypoxic conditions while maintaining multiple pathway influences through its interaction with hub proteins.
This detailed network analysis provides a systems-level understanding of TPZ’s mechanism of action, highlighting both direct interactions and broader pathway effects that contribute to its therapeutic efficacy when delivered via the N@P/T system.
(B) Molecular docking identifies key binding sites between TPZ and HIF1A
The 3D structure of HIF1A, its refined model, and validation analyses including the Ramachandran plot and ProSA Z-score assessment is shown in Fig. 2A. The I-TASSER modeling generated five models, with Model 1 showing the highest confidence metrics. The initial model demonstrated promising quality parameters with a C-score of − 0.24, an estimated TM-score of 0.68 ± 0.12, and an RMSD of 9.0 ± 4.6Å. The C-score, ranging from − 5 to + 2, indicates good model confidence as it approaches the positive range. The TM-score exceeding 0.5 suggests the correct topology of our modeled structure.
The refined structure, processed through GalaxyRefine, showed significant improvements in stereochemical quality. The best-refined model (Model 1) exhibited a GDT-HA score of 0.8965 with an RMSD of 0.556Å relative to the initial structure, indicating successful refinement while maintaining the core structure. The refinement process substantially improved structural parameters:
-
Reduction in poor rotamers from 15.5 to 0.8%
-
Enhancement in Ramachandran favored residues from 64.1 to 85.8%
-
Improvement in MolProbity score from 3.057 to 2.261
The refined model validation metrics showed notable improvements:
-
Increase to 77.7% residues in most favored regions
-
Reduction to 17.0% in additionally allowed regions
-
Decrease to 2.3% in generously allowed regions
-
Slight improvement in disallowed regions to 3.0%
-
ProSA Z-score of − 3.78, within the range of experimentally determined structures
-
ERRAT quality factor of 70.822, above the threshold of 50 for high-quality models
The molecular docking analysis using COACH-D revealed detailed binding interactions between TPZ and the refined HIF1A structure. Five distinct binding poses were identified, each with specific interaction characteristics and with varying degrees of confidence:
-
Two equally significant primary binding sites (Highest Confidence sites 1 and 2) with Cscore 1.0 and cluster size 3
Features: hypoxia-response elements, pH-sensing histidine, Serine-rich region for phosphorylation regulation
-
One intermediate site (Moderate Confidence site 3) with Cscore of 1.0 and cluster size of 2
Features: Redox-sensitive cysteine, compact interaction surface
-
Two additional sites (Lower Confidence sites 4 and 5) with Cscore 1.0 and 0.96 respectively, each with cluster size 1
Features: Large interaction surface, multiple regulatory sites, hydrophobic pocket, compact binding region
This hierarchical organization of binding sites reveals a complex interaction mechanism between TPZ and HIF1A, supported by consistently high confidence scores (CScore ≥ 0.96) across all predicted binding sites. The primary sites (Sites 1 and 2) demonstrate both maximum confidence scores and the largest cluster sizes (3), strongly indicating their predominant role in TPZ-HIF1A interactions through hypoxia sensing and phosphorylation-dependent regulation. The interaction specificity is further supported by additional regulatory mechanisms through the intermediate and additional binding sites. The refined model’s improved stereochemical quality adds substantial validity to these predictions, while the specific binding residues (highlighted in pink in Fig. 2A) map out crucial interaction zones that collectively contribute to TPZ’s hypoxia-selective activity. This comprehensive binding profile suggests a sophisticated molecular mechanism whereby TPZ achieves its therapeutic effects through multiple, coordinated interaction points with HIF1A under hypoxic conditions. Beside in silico studies, measurement of HIF-1A mRNA/protein level expression in TPZ treated cancer cells can further strengthen and explain the possible anti-cancer mechanism of TPZ under hypoxia conditions.
Characterization of N@P/T
One of the most important properties of NPs is their size and more of the characterization of NPs, such as physical stability, drug release, biological distribution, and cellular absorption, are related to size. Several variables, including the method of production, surface charge, and various chemical compounds, have a significant effect on the size of NPs35. TEM and SEM images show that the shape of N@P/T is homogeneously spherical and has a size of 200 nm (Fig. 3A) and 191.30 nm (Fig. 3B), respectively. The DLS analysis showed a size and zeta potential of N@P/T as 234.9 nm (Fig. 3C) and − 4.4 mV (Fig. 3D), respectively. The discrepancy between DLS and TEM/SEM measurements is attributed to the hydrated diameter of NPs, which is typically larger due to the hydration shell57. From a biological standpoint, the size of NPs plays a crucial role in determining their biodistribution and clearance pathways. In general, both the liver and spleen can store NPs smaller than 70 nm and larger than 250 nm, respectively10. Our NPs, ranging between these sizes, are suitable for drug delivery, allowing efficient cellular uptake without rapid clearance by the reticuloendothelial system10,57.

Characterization of N@P/T NPs. (A) TEM images revealed nanosized NPs with uniform spherical morphology, (B) SEM highlights size and surface texture, (C) size distribution analysis showed an average diameter of 236.1 nm, and (D) zeta potential measurement represented a negative surface charge for developed niosomes.
The hydroxyl group in cholesterol molecules that does not have an even distribution of polarity might be linked to NIO’s negative zeta potential58. The zeta potential of NPs may indicate their physical stability in biological environments, and an inappropriate zeta potential can causes vesicle accumulation, insufficient target specificity, and toxicity. It is commonly believed that a zeta potential value less than − 30 mV and greater than + 30 mV is necessary to ensure the physical stability of NPs63. However, we find that our formulated niosomes with a low zeta potential (− 4 mV) were stable at 4 °C for more than 3 months, which could be explained by steric stabilization, favorable formulation conditions, or other stabilizing interactions. Furthermore, our niosome maintained structural integrity over three months with reduced and non-significant drug release, confirming their viability as robust carriers for drug delivery.
For optimal stability, niosomes typically benefit from zeta potentials greater than − 10 mV, thus it is suggested to formulate niosomes with more negative charge64. In comparison to other nanocarriers, the value of − 4mV is relatively low, suggesting that niosomes may be more prone to aggregation and less stable over time than liposomes (− 10 mV to + 30 mV)65, polymeric nanoparticles (− 20 mV and + 40 mV)66, or dendrimers (exceeding + 40 mV) with higher zeta potentials67. The size of the N@P/T was reduced by probe sonication. The thermodynamic stability of Span 60 is high and facilitates the downsizing of N@P/T compared to the use of other surfactants68. Previous studies on smaller nanoparticles (< 100 nm) emphasized the trade-off between size and stability; while smaller NPs penetrate tissues more effectively, they face challenges with rapid clearance and reduced payload capacity57,69. In this study, N@P/T NPs, with a size of 234.9 nm, a surface charge of − 4.4 mV, and a PDI of 0.357, strikes a balance between stability and cellular uptake, showing promise for nano drug delivery systems (NDDSs).
The primary disadvantage of NPs is their potential to expand or crack during storage. Studies indicate that NIO size, zeta potential, PDI, and drug quantity remain stable during refrigeration (4 ± 0.5 °C). Other research highlights that NP aggregation or fusion increases size over time, particularly under uncontrolled storage conditions. Maintaining refrigeration significantly reduces these changes, aligning with findings that lower temperatures minimize drug release70,71. In our experiments, drug release from refrigerated N@P/T was negligible, confirming NP stability.
The FT-IR technique was used to investigate the chemical structure of NIO and N@P/T (Fig. 4). The NIO spectrum, 3250–3550 cm−1 is related to the OH group, 1738 cm−1 indicates the peak of C=O stretching vibrations, 2923 is related to the C–H bond, and 1109 cm−1 is the bond of C–O stretching. 1249 cm−1 is related to ester bond stretching72,73. 1580–1650 cm−1 is related to the amine groups of PAMAM74.

FTIR spectra of NIO and N@P/T nanoparticles. There was no difference between NIO and N@P/T regarding the characteristic peaks which denotes physical loading of P/T inside niosome.
Dual nanoparticle system
The integration of TPZ into PAMAM dendrimers followed by encapsulation with NIOs marks an innovative approach to optimizing the delivery of chemotherapeutics while mitigating the adverse effects commonly associated with PAMAM75,76. Dendrimer-based platforms have attracted attention for use in pharmaceutical applications. Due to their size of 1–10 nm, dendrimers can efficiently diffuse into cells77. Dual NPs approach enhances tumor treatment efficacy by overcoming the limitations of single NPs and leveraging the synergistic effects of combined delivery systems78. NIOs can encapsulate a substantial amount of P/T in their hydrophobic regions, releasing it into cancer cells via endocytosis. The acidic tumor microenvironment promotes the release of P/T from NIOs, while the ultrasmall PAMAM dendrimers, rich in amino groups, facilitate the penetration and uptake of the nanocarrier into tumor cells.
Drug entrapment efficiency and drug release behavior
Improving encapsulation efficiency (EE) is a critical goal in the formulation of NP systems for drug delivery79. Our research highlights the effectiveness of using Span 60 in the synthesis of NIOs to achieve higher EE. The long alkyl chain of Span 60 enhances the stability of membrane bilayers, which translates into improved drug encapsulation40. Additionally, incorporating cholesterol stabilizes the surfactant bilayer, increasing microviscosity and reducing drug leakage80. The thin-film hydration method also contributes to increased drug loading in NIOs68. The EE in N@P/T was more than 84%, which indicates the high loading potential of NIO as a carrier for P/T.
The release of TPZ from N@P/T exhibited a controlled release pattern, being rapid in the first 5 h and then slowing until 48 h. As shown in Fig. 5, TPZ release was 28% at pH 5.8 and 20% at pH 7.4 after 5 h. After 48 h, cumulative drug release reached 75% at pH 5.8 and 48% at pH 7.4. The enhanced release at acidic pH demonstrates the pH-sensitive nature of the niosomal formulation, advantageous for targeting the acidic tumor microenvironment. NIOs respond to acidic pH due to the behavior of amphiphilic surfactants, which can enhance drug release81. The high acidity of tumor cells is attributed to hypoxia, or lack of oxygen owing to the inadequate blood supply27. In vitro drug release studies showed that N@P/T performs better at pH 5.8 than at pH 7.4, confirming its suitability for novel drug delivery systems (NDDS). This finding aligns with our recent study by Hashemi et al.59 that demonstrated NIOs containing Fingolimod coated by Hyaluronic acid are sensitive to acidic pH conditions. The kinetic models used to fit drug release curves at different pH are shown in Table 1. At pH = 7.4 and 5.8, the fitted models for the release curves was Zero order for both normal and acidic pH conditions. This indicates that niosome nanoparticles provide a consistent and predictable rate of drug release, independent of the concentration of the drug in the system which is advantageous for achieving sustained therapeutic effects (Table 1). There are numerous advantages to this mode of action: (i) Sustained Release: Niosomes can be engineered to provide a controlled release of drugs, maintaining a steady concentration in the bloodstream. This is particularly beneficial for chronic conditions such as cancer where consistent drug levels are necessary. (ii) Reduced Frequency of Dosing: With a zero-order release profile, the need for frequent dosing is reduced. Patients can benefit from improved adherence to treatment regimens, as they do not need to take medication as often. (iii) Minimized Side Effects: By preventing peaks and troughs in drug concentration, zero-order release can help minimize side effects associated with high drug levels while ensuring therapeutic efficacy. (iv) Enhanced Bioavailability: Niosomes can improve the solubility and stability of poorly water-soluble drugs, leading to enhanced bioavailability and prolonged therapeutic effects. (v) Targeted Delivery: Niosomes can be modified to target specific tissues or cells, allowing for prolonged drug action at the site of interest while reducing systemic exposure19,68.

The cumulative in vitro release profile of TPZ from N@P/T NPs was evaluated at pH 7.4 and 5.8, simulating physiological and acidic environments, respectively, at 37 °C. The results demonstrate a sustained release pattern, with a higher release rate observed at pH 5.8, indicating enhanced drug availability in acidic conditions.
Uptake
Analyzing the cellular uptake of NPs is essential for assessing DDs’ potential. It is demonstrated that slight physicochemical variations have a major biological impact on the way NPs are absorbed by cells and the way they function biologically. The size of NPs and zeta potential are the most important properties of NPs that impact the cells’ uptake, efficiency, and pathway10. Bannunah et al.82 studied the mechanisms of nanoparticle internalization across intestinal epithelial cells, highlighting that both size and surface charge play crucial roles. Previous studies have shown in vivo biodistribution NPs with slightly negative charges and a particle size around 150 nm tend to accumulate more efficiently in the tumor83. For instance, rod-shaped NPs tend to show the highest cellular uptake, followed by spherical ones84. Ma et al.85 discussed how NPs size greatly influences their cellular uptake, with medium-sized NPs often showing the most efficient uptake. In our previous study, the uptake of NIO NPs was evaluated in MCF-7 and MDA-MB-231 BC cells, demonstrating that the internalization of NIO by cancer cells is highly efficient44,57. In the current study, flow cytometry analysis demonstrated significant cellular uptake of FITC-labeled N@P/T in MDA-MB-231 cells after 4h compared to untreated controls (Fig. 6).

Flow cytometric analysis of cellular uptake in MDA-MB-231 cells. Representative flow cytometry histograms showing FITC fluorescence intensity of (a) untreated control cells and (b) cells treated with FITC-labeled NIO. The significant shift in fluorescence intensity demonstrates efficient cellular internalization of the nanoformulation. Analysis was performed after 4 h of incubation at 37 °C.
MTT assay for cytotoxicity evaluation
After a 48-h treatment of MDA-MB-231 cells with various concentrations (0–150 μM) of NIO, P/T, and N@P/T, an MTT assay was performed to evaluate cytotoxicity. NIO showed negligible toxicity, confirming its suitability as a drug carrier. In contrast, N@P/T exhibited significantly higher toxicity than P/T at all concentrations (p < 0.001), with cell viability dropping below 12% at 50 μM and above, while NIO maintained ~ 100% viability at 100 μM (Fig. 7).

Cytotoxicity evaluation of different formulations on MDA-MB-231 breast cancer cells after 48h treatment. The niosome carrier shows minimal cytotoxicity, while N@P/T demonstrates significantly enhanced cytotoxic effects compared to free TPZ and P/T at all tested concentrations. Data are presented as mean ± SD (n = 3). Two-way ANOVA, Tukey post-hoc test: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, ns = not significant.
IC50 values were determined to assess therapeutic efficacy. N@P/T had the lowest IC50 value (14.14 μM), indicating superior cytotoxicity compared to P/T (IC50 = 71.37 μM) and free TPZ (IC50 = 143.3 μM). This nanoformulation enhanced TPZ toxicity under 2D cell culture conditions (21% O2). Previous studies indicate that IC50 values for TPZ decrease in 3D versus 2D cultures, demonstrating a more favorable environment for hypoxia-activated drugs (Fig. 8)86. Using CCK-8 assay to examine cytotoxicity of different derivatives of TPZ on CT26 murine colorectal tumor cells, TPZ IC50 was determined 51.42 μM vs. 16.35 μM under normoxic (∼20% O2) or hypoxic (∼1% O2) conditions, respectively87. Our previous work with hypoxia-activated PAPAM NPs showed that PAP@TPZ at 2 μg/mL completely destroyed 3D cultured MDA-MB-231 cells33. Additionally, hypoxia-activated polymeric NPs (APAP@TPZ) at 50 μg/mL induced complete apoptosis of MDA-MB-231 cells. Research by Saunders et al.88 found TPZ IC50 values of 1758 μM under normoxia and 63 μM under hypoxia for the resistant A549c50 cell line. Combining NDDSs with other anti-cancer strategies can further lower IC50 values. For instance, hyaluronic-acid-based nanoparticles (PA/HA-Ce6@TPZ NPs) reduced IC50 to 0.2 μg/mL against 4T1 breast cancer cells89. The enhanced cytotoxicity of N@P/T is attributed to several mechanisms. TPZ activates under hypoxic conditions to generate toxic free radicals that damage DNA13. The small size of PAMAM’s enhances drug penetration into cells90, while its positive charge creates membrane pores that facilitate cellular uptake76. However, the high concentration of amine groups in G5 PAMAM may be toxic to cell membranes67. PAMAM dendrimers localize in mitochondria and generate reactive oxygen species (ROS), which leads to DNA damage and cell death. The cationic PAMAM enters through endosomes and reaches mitochondria, where acid–base equilibrium reactions between secondary amines and their conjugate bases produce ROS and alter mitochondrial pH. Different cell types show different toxic responses to PAMAM, possibly due to differences in membrane composition or antioxidant levels67,91. In our previous study, loading TPZ into a dendrimer significantly enhanced its therapeutic effects33. NIOs have been shown to enhance anticancer drug efficacy, as demonstrated by Masoumi et al.92 in targeting BC cells. Utilizing NIO helps address the hydrophobicity and toxicity issues associated with PAMAM dendrimers, as inherent cationic nature can still lead to disruption of the membrane67.The encapsulation within a niosomal system provides a balanced carrier that protects sensitive tissues by enhancing the therapeutic index of the encapsulated agents. This is achieved by increasing the amount of P/T that can enter the cells and allowing for high loading of P/T in NIO.

Dose–response curves and IC50 values for different formulations in MDA-MB-231 cells after 48h treatment. Data are presented as mean ± SD (n = 3). Statistical significance: *p < 0.05, **p < 0.01.
To optimize this system and increase P/T efficacy, we employed NIO as a carrier. NIO’s ability to load substantial amounts of drug and enhance toxicity specifically in cancer cells improved P/T delivery and therapeutic efficacy, as evidenced by the steeper dose–response curve and lower IC50 value of N@P/T.
Apoptosis assay
The MDA-MB-231 cells were incubated for 48 h with P/T and N@P/T. Annexin V was then used as a sensitive probe for the phospholipid phosphatidylserine. In addition, PI also stain DNA in flow cytometry due to its intercalating ability. Flow cytometry analysis revealed distinct patterns of cell death between treatment groups. Untreated control cells showed minimal apoptosis (Fig. 9A), while treatment with P/T induced significant apoptosis with 38.74% late apoptotic and 19.51% early apoptotic cells (Fig. 9B). Notably, N@P/T treatment demonstrated enhanced apoptotic effects with 44.85% late apoptotic and 6.31% early apoptotic cells (Fig. 9C). In total, P/T (44.28%) and N@P/T (65.33%) significantly increased the apoptosis of MDA-MB-231 cells compared to untreated controls (Fig. 9D).

Analysis of apoptotic by Flow cytometry on MDA-MB-231 cells. (A) Untreated group and treated group by (B) P/T, (C) N@P/T, (D) statistical analysis among studied groups, One-way ANOVA, **p < 0.01***p < 0.001.
Research by Bo Hong et al.13 indicated that TPZ selectively targets hypoxic environments to treat nasopharyngeal cancer (NPC), suppressing NPC cell proliferation and disrupting the cell cycle in S-phase, evidenced by PARP cleavage as a sign of apoptosis. They found that TPZ activated Chk1 and Chk2 checkpoint regulators in NPC cells under hypoxia. Similarly, Yingchao Hao et al.93 demonstrated that PAMAM G5 enhanced TPZ’s effects under hypoxic conditions in 4T1 breast cancer cells, inducing apoptosis. Our research experience aligns with others that the formulation of different type of drugs with NIO leads to increased apoptosis94,95. In a previous study, the formulation of tamoxifen-loaded NIO increased apoptosis in MDA-MB-231 and SKBR3 of BC through significant upregulation of caspase-9 and caspase-394.
Real-time assessment
Treatments of MDA-MB-231 cells by TPZ and N@P/T have shown a significant decrease in the proto-oncogene BCL2 and an increase in BAX and caspase3. A previous study showed that TPZ significantly increases caspase-3 and stimulates cells to undergo apoptosis96. Hongfang Chen et al.97 developed oxygen-absorbing perfluorotributylamine (PFA) and polydopamine (PDA)-coated UiO-66 with TPZ (TPZ/PFA@UiO-66@PDA). TPZ/PFA@UiO-66@PDA increased hypoxia, induced cell apoptosis by increasing BAX and caspase3 expression genes via the oxygen-dependent HIF1A pathway, and decreased oxygen levels in vivo after intratumoral injection. Meanwhile, treatment of cancer cells with TPZ has shown a significantly decreased expression of BCL298. In this study, while TPZ had no significant effect on BCL2 gene expression compared to the untreated control, N@P/T treatment resulted in ~ 7.3 folds drop in BCL2 expression. Also, both TPZ and N@P/T resulted in BAX upregualtion by ~ 16.99 and ~ 16.3 folds, respectively compared to the control. In case of caspase3, there were 2.19 and 1.12 folds upregualtion for TPZ and N@P/T respectively, compared to the control, demonstrating the key role of NIO in efficient delivering of P/T to kill cancer cells (Fig. 10). It is worth to mention compared to free TPZ, N@PT induced a much higher downregulation of BCL2 and a higher upregulation of caspase 3, however for BAX, there was no statistically difference between free TPZ and its nanoformualtion N@PT.

Effect of free drug and nanoparticle formulation on gene expression of apoptotic genes as well as HIF1A. One way ANOVA, Tukey post-hoc test, ns: non-significant, *p < 0.05, **p < 0.01.
As bioinformatic studies reveled HIF1A a possible target for TPZ, we further examined the role of TPZ on HIF1A mRNA expression. Our results demonstrated that the levels of HIF1A was markedly decreased to 1.86 and 2.70 folds in groups treated N@P/T under hypoxia and normoxia conditions compared to the untreated control, respectively. Our results are in line with previous reports that TPZ can reduce HIF1A protein level, in a manner independent of its topoisomerase IIα inhibition action99 (Fig. 10).
Spheroid size test
TPZ is preferentially cytotoxic in hypoxic areas of solid tumors. The 3D cell culture is beneficial for the preclinical development of new drugs by mimicking the physiological conditions and properties of cancer microenvironment including hypoxia1. It addresses the issues of conventional 2D in vitro culture, provides more comprehensive data on 3D interactions between cells and matrices, and exhibits a more clinically realistic response to treatment agents11. MDA-MB-231 spheroids were treated with P/T and N@P/T at concentrations ranging from 25 to 150 μM for 24, 48, and 72 h. As shown in Fig. 11, the spherical conditions and the effects of different doses of P/T and especially N@P/T on the morphology and size of MDA-MB-231 spheroids were investigated over time. The microscopy images and quantitative analysis demonstrate that cellular integrity is progressively lost with treatment. The size of spheroids treated with N@P/T was significantly decreased at 50 μM, with notable morphological changes visible by 48 h100. This data proved that N@P/T is suitable NDDS for P/T delivery to MDA-MB-231 spheroids.

Effect of P/T and N@P/T treatments on MDA-MB-231 spheroid size and morphology. (A) Representative microscopic images showing morphological changes in MDA-MB-231 spheroids treated with different concentrations of P/T and N@P/T for 24h, 48h, and 72h. (B) Quantitative analysis of relative spheroid size over time. Data are presented as mean ± SD (n = 3).
Conclusion
In this study, in silico analysis revealed complex protein–protein interaction networks centered on HIF1A and identified several high-confidence binding sites between TPZ and HIF1A, providing insights at the molecular level into TPZ’s selective activation under hypoxic conditions. Based on these computational predictions and further experimental validation, N@P/T was designed and successfully synthesized, incorporating TPZ into dendrimers and then loaded into NIOs. The potential of dual NPs against many diseases, especially cancer cells, has been studied and confirmed. In this study, TPZ as a target of cancer cells in the hypoxia condition loaded in the PAMAM, which has a great potential to penetrate into cancer cells because of plenty of amino groups. Then P/T was loaded into NIO to enhance the drug payload while eliminating dendrimer off-target toxicity and elimination. N@P/T was examined on the MDA-MB-231 and showed significant cytotoxicity compared to P/T. The results provided the proof-of-principle for efficacy of dual package NDDS to cross numerous biological barriers and provide selective delivery of TPZ to eliminate hypoxic tumors. For future studies, optimization of niosome with more negative charge, fluorescent tracking of P/T by live/dead staining, pharmacokinetic and pharmacodynamics of niosome with and without targeting agents (e.g. HA), and testing combination treatments with other modalities e.g. hypoxia-aggravating agents to improve TPZ’s efficacy is needed to provide valuable principle regarding clinical applicability of developed niosome systems for hypoxic tumors.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
-
Mahmudi, H. et al. Tumor microenvironment penetrating chitosan nanoparticles for elimination of cancer relapse and minimal residual disease. Front. Oncol. 12, 1054029 (2022).
-
Nejati-Koshki, K. et al. Inhibition of leptin gene expression and secretion by silibinin: Possible role of estrogen receptors. Cytotechnology 64(6), 719–726 (2012).
-
Huang, Z., Yu, P. & Tang, J. Characterization of triple-negative breast cancer MDA-MB-231 cell spheroid model. OncoTargets Ther. 13, 5395–5405 (2020).
-
Mazloomi, M. et al. Advanced drug delivery platforms target cancer stem cells. Asian J. Pharm. Sci. 20(3), 101036 (2025).
-
Karimi, S. et al. For and against tumor microenvironment: Nanoparticle-based strategies for active cancer therapy. Mater. Today Bio 31, 101626 (2025).
-
Semenza, G. L. Oxygen sensing, homeostasis, and disease. N. Engl. J. Med. 365(6), 537–547 (2011).
-
Semenza, G. L. Hypoxia-inducible factors: Mediators of cancer progression and targets for cancer therapy. Trends Pharmacol. Sci. 33(4), 207–214 (2012).
-
Riffle, S., Pandey, R. N., Albert, M. & Hegde, R. S. Linking hypoxia, DNA damage and proliferation in multicellular tumor spheroids. BMC Cancer 17, 1–12 (2017).
-
Jahanban-Esfahlan, R., Seidi, K. & Zarghami, N. Tumor vascular infarction: Prospects and challenges. Int. J. Hematol. 105(3), 244–256 (2017).
-
Azizi, M. et al. Multifunctional nanostructures: Intelligent design to overcome biological barriers. Mater. Today Bio 20, 100672 (2023).
-
Shahpouri, M. et al. Prospects for hypoxia-based drug delivery platforms for the elimination of advanced metastatic tumors: From 3D modeling to clinical concepts. J. Control. Release 353, 1002–1022 (2023).
-
Siim, B. G., Menke, D. R., Dorie, M. J. & Brown, J. M. Tirapazamine-induced cytotoxicity and DNA damage in transplanted tumors: Relationship to tumor hypoxia. Can. Res. 57(14), 2922–2928 (1997).
-
Hong, B. et al. Chan AT-C: Hypoxia-targeting by tirapazamine (TPZ) induces preferential growth inhibition of nasopharyngeal carcinoma cells with Chk1/2 activation. Invest. New Drugs 29, 401–410 (2011).
-
Kitchen, D. B., Decornez, H., Furr, J. R. & Bajorath, J. Docking and scoring in virtual screening for drug discovery: methods and applications. Nat. Rev. Drug Discov. 3(11), 935–949 (2004).
-
Lu, H. et al. Recent advances in the development of protein-protein interactions modulators: Mechanisms and clinical trials. Signal Transduct. Target Ther. 5(1), 213 (2020).
-
Salemi, A., Pourseif, M. M. & Omidi, Y. Next-generation vaccines and the impacts of state-of-the-art in-silico technologies. Biologicals 69, 83–85 (2021).
-
Tang, J. & Aittokallio, T. Network pharmacology strategies toward multi-target anticancer therapies: From computational models to experimental design principles. Curr. Pharm. Des. 20(1), 23–36 (2014).
-
Dianat-Moghadam, H. et al. Cancer stem cells-emanated therapy resistance: Implications for liposomal drug delivery systems. J. Control. Release 288, 62–83 (2018).
-
Hashemi, Z. et al. Engineered niosomes for cancer therapy: Classification, synthesis, and clinical applications. BioNanoScience 15(1), 34 (2024).
-
Pourbakhsh, M., Jabraili, M., Akbari, M., Jaymand, M. & Jahanban Esfahlan, R. Poloxamer-based drug delivery systems: Frontiers for treatment of solid tumors. Mater. Today Bio 32, 101727 (2025).
-
Massoumi, B. et al. A novel multi-stimuli-responsive theranostic nanomedicine based on Fe3O4@Au nanoparticles against cancer. J. Drug Dev. Ind. Pharm. 46(11), 1832–1843 (2020).
-
Samadian, H. et al. A de novo theranostic nanomedicine composed of PEGylated graphene oxide and gold nanoparticles for cancer therapy. J. Mater. Res. 35(4), 430–441 (2020).
-
Ayoubi-Joshaghani, M. H. et al. Potential applications of advanced nano/hydrogels in biomedicine: Static, dynamic, multi-stage, and bioinspired. Adv. Fun. Mater. 30(45), 2004098 (2020).
-
Eskandani, M., Jahanban-Esfahlan, R., Sadughi, M. M. & Jaymand, M. Thermal-responsive β-cyclodextrin-based magnetic hydrogel as a de novo nanomedicine for chemo/hyperthermia treatment of cancerous cells. Heliyon 10(11), e32183 (2024).
-
Derakhshankhah, H. et al. Folate-conjugated thermal-and pH-responsive magnetic hydrogel as a drug delivery nano-system for “smart” chemo/hyperthermia therapy of solid tumors. Mater. Today Commun. 30, 103148 (2022).
-
Jahanban-Esfahlan, R. et al. Multi-stimuli-responsive magnetic hydrogel based on tragacanth gum as a de novo nanosystem for targeted chemo/hyperthermia treatment of cancer. J. Mater. Res. 36, 858–869 (2021).
-
Sayadnia, S., Arkan, E., Jahanban-Esfahlan, R., Sayadnia, S. & Jaymand, M. Tragacanth gum-based pH-responsive magnetic hydrogels for “smart” chemo/hyperthermia therapy of solid tumors. Polym. Adv. Technol. 32(1), 262–271 (2021).
-
Massoumi, B. et al. Polymers: Electrically conductive nanofibers composed of chitosan-grafted polythiophene and poly (ε-caprolactone) as tissue engineering scaffold. Fibers Polym. 22(1), 49–58 (2021).
-
Dadashi, H. et al. A rapid protocol for synthesis of chitosan nanoparticles with ideal physicochemical features. Heliyon 10(11), e32228 (2024).
-
Mahmudi, H. et al. Self-activating chitosan-based nanoparticles for sphingosin-1 phosphate modulator delivery and selective tumor therapy. Int. J. Biol. Macromol. 272, 132940 (2024).
-
Jahanban-Esfahlan, A. et al. Dynamic DNA nanostructures in biomedicine: Beauty, utility and limits. J. Control. Release 315, 166–185 (2019).
-
Jahanban-Esfahlan, R. et al. Static DNA nanostructures for cancer theranostics: Recent progress in design and applications. Nannotechnol. Sci. Appl. 2019(12), 25–46 (2019).
-
Shahpouri, M. et al. Dual-stage acting dendrimeric nanoparticle for deepened chemotherapeutic drug delivery to tumor cells. Adv. Pharm. Bull. https://doi.org/10.34172/apb.2024.054 (2024).
-
Mollazade, M. et al. PAMAM dendrimers augment inhibitory effects of curcumin on cancer cell proliferation: possible inhibition of telomerase. Asian Pac. J. Cancer Prev. APJCP 14(11), 6925–6928 (2013).
-
Doustmihan, A. et al. Molecular targets, therapeutic agents and multitasking nanoparticles to deal with cancer stem cells: A narrative review. J. Control. Release 363, 57–83 (2023).
-
Baghban, R. et al. Tumor microenvironment complexity and therapeutic implications at a glance. Cell Commun. Signal. 18, 1–19 (2020).
-
Seidi, K. et al. Bioinspired hydrogels build a bridge from bench to bedside. Nano Today 39, 101157 (2021).
-
Hu, C.-M.J. & Zhang, L. Nanoparticle-based combination therapy toward overcoming drug resistance in cancer. Biochem. Pharm. 83(8), 1104–1111 (2012).
-
Jahanban-Esfahlan, R. et al. Dual stimuli-responsive polymeric hollow nanocapsules as “smart” drug delivery system against cancer. Polym. Plast. Technol. Mater. 59 (13), 1492-1504 (2020).
-
Ahmadi, S. M., Seyedabadi, M., Ebrahimnejad, P., Abasi, M. & Nokhodchi, A. Efficient delivery of gold nanoparticles and miRNA-33a via cationic PEGylated niosomal formulation to MCF-7 breast cancer cells. AAPS PharmSciTech 25(7), 213 (2024).
-
Seidi, K., Jahanban-Esfahlan, R. & Zarghami, N. Tumor rim cells: From resistance to vascular targeting agents to complete tumor ablation. Tumour Biol. 39(3), 1010428317691001 (2017).
-
Seidi, K., Neubauer, H. A., Moriggl, R., Jahanban-Esfahlan, R. & Javaheri, T. Tumor target amplification: Implications for nano drug delivery systems. J. Control. Release 275, 142–161 (2018).
-
Tarach, P. & Janaszewska, A. Recent advances in preclinical research using PAMAM dendrimers for cancer gene therapy. Int. J. Mol. Sci. 22(6), 2912 (2021).
-
Fakeri, M., Haghi, M., Jahanban Esfahlan, R., Fathi, M. & Hosseinpour Feizi, M. A. Targeted apoptosis in breast cancer cells via niosome-mediated delivery of cyclophosphamide and sodium oxamate. Mol. Biol. Rep. 52(1), 139 (2025).
-
Kuhn, M., von Mering, C., Campillos, M., Jensen, L. J. & Bork, P. STITCH: Interaction networks of chemicals and proteins. Nucl. Acids Res. 36, D684-688 (2008).
-
UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucl. Acids Res. 49(D1), D480–D489 (2021).
-
Otasek, D., Morris, J. H., Boucas, J., Pico, A. R. & Demchak, B. Cytoscape automation: Empowering workflow-based network analysis. Genome Biol. 20(1), 185 (2019).
-
Kanehisa, M., Furumichi, M., Sato, Y., Kawashima, M. & Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucl. Acids Res. 51(D1), D587–D592 (2023).
-
Chin, C. H. et al. cytoHubba: identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 8(Suppl4), S11 (2014).
-
Wu, Q., Peng, Z., Zhang, Y. & Yang, J. Coach-D: Improved protein-ligand binding sites prediction with refined ligand-binding poses through molecular docking. Nucl. Acids Res. 46(W1), W438–W442 (2018).
-
Zheng, W., Zhang, C., Bell, E. W. & Zhang, Y. I-TASSER gateway: A protein structure and function prediction server powered by XSEDE. Future Gener. Comput. Syst. 99, 73–85 (2019).
-
Heo, L., Park, H. & Seok, C. GalaxyRefine: Protein structure refinement driven by side-chain repacking. Nucl. Acids Res. 41, W384-388 (2013).
-
Laskowski, R. A., Rullmannn, J. A., MacArthur, M. W., Kaptein, R. & Thornton, J. M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 8(4), 477–486 (1996).
-
Colovos, C. & Yeates, T. O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 2(9), 1511–1519 (1993).
-
Wiederstein, M. & Sippl, M. J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucl. Acids Res. 35, W407-410 (2007).
-
Pettersen, E. F. et al. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 25(13), 1605–1612 (2004).
-
Kaveh Zenjanab, M. et al. Hyaluronic acid-targeted niosomes for effective breast cancer chemostarvation therapy. ACS Omega 9(9), 10875–10885 (2024).
-
Sharifi-Azad, M. et al. Codelivery of methotrexate and silibinin by niosome nanoparticles for enhanced chemotherapy of CT26 colon cancer cells. Biomed. Mater. 19(5), 055015 (2024).
-
Hashemi, Z. et al. Hyaluronic acid-modified theranostic niosomes for targeted fingolimod delivery and inhibition of triple-negative breast cancer metastasis. Med. Oncol. 42(7), 256 (2025).
-
Fathi, M. et al. Thermo-sensitive chitosan copolymer-gold hybrid nanoparticles as a nanocarrier for delivery of erlotinib. Int. J. Biol. Macromol. 106, 266–276 (2018).
-
Dadashi, H. et al. Chitosan nanoparticles loaded with metformin and digoxin synergistically inhibit MCF-7 breast cancer cells through suppression of NOTCH-1 and HIF-1α gene expression. Int. J. Biol. Macromol. 287, 138418 (2024).
-
Esfahlan, R. J. et al. The possible impact of obesity on androgen, progesterone and estrogen receptors (ERalpha and ERbeta) gene expression in breast cancer patients. Breast Cancer 5, 227–237 (2011).
-
Amiryaghoubi, N. et al. Smart chitosan–folate hybrid magnetic nanoparticles for targeted delivery of doxorubicin to osteosarcoma cells. Colloids Surf. B 220, 112911 (2022).
-
Khoee, S. & Yaghoobian, M. Chapter 6—Niosomes: A Novel Approach in Modern Drug Delivery Systems. In Nanostructures for Drug Delivery (ed. Andronescu, E.) 207–237 (Elsevier, 2017).
-
Allen, T. M. & Cullis, P. R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Deliv. Rev. 65(1), 36–48 (2013).
-
Crucho, C. I. C. & Barros, M. T. Polymeric nanoparticles: A study on the preparation variables and characterization methods. Mater. Sci. Eng. C 80, 771–784 (2017).
-
Fox, L. J., Richardson, R. M. & Briscoe, W. H. PAMAM dendrimer-cell membrane interactions. Adv. Coll. Interface. Sci. 257, 1–18 (2018).
-
Zenjanab, M. K., Pakchin, P. S., Fathi, M., Abdolahinia, E. D. & Adibkia, K. Niosomes containing paclitaxel and gold nanoparticles with different coating agents for efficient chemo/photothermal therapy of breast cancer. Biomed. Mater. 19(3), 035015 (2024).
-
Alimohammadvand, S. et al. Aripiprazole-loaded niosome/chitosan-gold nanoparticles for breast cancer chemo-photo therapy. BMC Biotechnol. 24(1), 108 (2024).
-
Chandrakala, V., Aruna, V. & Angajala, G. Review on metal nanoparticles as nanocarriers: Current challenges and perspectives in drug delivery systems. Emerg. Mater. 5(6), 1593–1615 (2022).
-
Abdelwahed, W., Degobert, G., Stainmesse, S. & Fessi, H. Freeze-drying of nanoparticles: Formulation, process and storage considerations. Adv. Drug Deliv. Rev. 58(15), 1688–1713 (2006).
-
Samed, N., Sharma, V. & Sundaramurthy, A. Hydrogen bonded niosomes for encapsulation and release of hydrophilic and hydrophobic anti-diabetic drugs: An efficient system for oral anti-diabetic formulation. Appl. Surf. Sci. 449, 567–573 (2018).
-
Mehta, S. & Jindal, N. Formulation of tyloxapol niosomes for encapsulation, stabilization and dissolution of anti-tubercular drugs. Colloids Surf. B 101, 434–441 (2013).
-
Pakchin, P. S., Fathi, M., Ghanbari, H., Saber, R. & Omidi, Y. A novel electrochemical immunosensor for ultrasensitive detection of CA125 in ovarian cancer. Biosens. Bioelectron. 153, 112029 (2020).
-
Baranei, M. et al. Anticancer effect of green tea extract (GTE)-Loaded pH-responsive niosome coated with PEG against different cell lines. Mater. Today Commun. 26, 101751 (2021).
-
McNerny, D. Q., Leroueil, P. R. & Baker, J. R. Understanding specific and nonspecific toxicities: A requirement for the development of dendrimer-based pharmaceuticals. Rev. Nanomed. Nanobiotechnol. 2(3), 249–259 (2010).
-
Kukowska-Latallo, J. F. et al. Nanoparticle targeting of anticancer drug improves therapeutic response in animal model of human epithelial cancer. Can. Res. 65(12), 5317–5324 (2005).
-
King, M. R. & Mohamed, Z. J. Dual nanoparticle drug delivery: The future of anticancer therapies?. Futur. Med. 2, 95–98 (2017).
-
Liu, J. F., Jang, B., Issadore, D. & Tsourkas, A. Use of magnetic fields and nanoparticles to trigger drug release and improve tumor targeting. Rev. Nanomed. Nanobiotechnol. 11(6), e1571 (2019).
-
Safari Sharafshadeh, M., Tafvizi, F., Khodarahmi, P. & Ehtesham, S. Folic acid-functionalized PEGylated niosomes co-encapsulated cisplatin and doxoribicin exhibit enhanced anticancer efficacy. Cancer Nanotechnol. 15(1), 14 (2024).
-
Rezaei, T. et al. Folic acid-decorated ph-responsive nanoniosomes with enhanced endocytosis for breast cancer therapy: In vitro studies. Front. Pharmacol. 13, 851242 (2022).
-
Bannunah, A. M., Vllasaliu, D., Lord, J. & Stolnik, S. Mechanisms of nanoparticle internalization and transport across an intestinal epithelial cell model: Effect of size and surface charge. Mol. Pharm. 11(12), 4363–4373 (2014).
-
He, C., Hu, Y., Yin, L., Tang, C. & Yin, C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials 31(13), 3657–3666 (2010).
-
Albanese, A., Tang, P. S. & Chan, W. C. The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annu. Rev. Biomed. Eng. 14, 1–16 (2012).
-
Ma, N. et al. Influence of nanoparticle shape, size, and surface functionalization on cellular uptake. J. Nanosci. Nanotechnol. 13(10), 6485–6498 (2013).
-
Nhan, N. et al. Anti-tumor activity of plant extracts against human breast cancer cells are different in monolayer and three-dimensional cell culture screening models: A comparison on 34 extracts. Biomed. Res. Ther. 7, 3667–3677 (2020).
-
Xu, Y. et al. Introducing urea into the tirapazamine derivatives to enhance anti-cancer therapy. Nat. Sci. Rev. 11, 038 (2024).
-
Saunders, M., Patterson, A., Chinje, E., Harris, A. & Stratford, I. NADPH: cytochrome c (P450) reductase activates tirapazamine (SR4233) to restore hypoxic and oxic cytotoxicity in an aerobic resistant derivative of the A549 lung cancer cell line. Br. J. Cancer 82(3), 651–656 (2000).
-
Zhu, R. et al. Cancer-selective bioreductive chemotherapy mediated by dual hypoxia-responsive nanomedicine upon photodynamic therapy-induced hypoxia aggravation. Biomacromol 20(7), 2649–2656 (2019).
-
Xie, Z. et al. Targeting tumor hypoxia with stimulus-responsive nanocarriers in overcoming drug resistance and monitoring anticancer efficacy. Acta Biomater. 71, 351–362 (2018).
-
Ainalem, M.-L. et al. On the ability of PAMAM dendrimers and dendrimer/DNA aggregates to penetrate POPC model biomembranes. J. Phys. Chem. B 114(21), 7229–7244 (2010).
-
Masoumi Godgaz, S., Asefnejad, A. & Bahrami, S. H. Fabrication of PEGylated SPIONs-loaded niosome for codelivery of paclitaxel and trastuzumab for breast cancer treatment: In vivo study. ACS Appl. Bio Mater. 7(5), 2951–2965 (2024).
-
Hao, Y. et al. A tumor microenvironment-responsive poly (amidoamine) dendrimer nanoplatform for hypoxia-responsive chemo/chemodynamic therapy. J. Nanobiotechnol. 20(1), 1–15 (2022).
-
Akbarzadeh, I. et al. The optimized formulation of tamoxifen-loaded niosomes efficiently induced apoptosis and cell cycle arrest in breast cancer cells. AAPS PharmSciTech 23(1), 57 (2022).
-
Fatemizadeh, M. et al. Apoptosis induction, cell cycle arrest and anti-cancer potential of tamoxifen-curcumin loaded niosomes against MCF-7 cancer cells. Iran. J. Pathol. 17(2), 183 (2022).
-
Bidkar, A. P., Sanpui, P. & Ghosh, S. S. Red blood cell-membrane-coated poly (lactic-co-glycolic acid) nanoparticles for enhanced chemo-and hypoxia-activated therapy. ACS Appl. Bio Mater. 2(9), 4077–4086 (2019).
-
Chen, H. et al. Polydopamine-coated UiO-66 nanoparticles loaded with perfluorotributylamine/tirapazamine for hypoxia-activated osteosarcoma therapy. J. Nanobiotechnol. 19, 1–18 (2021).
-
Zhao, H. et al. Biomimetic decoy inhibits tumor growth and lung metastasis by reversing the drawbacks of sonodynamic therapy. Adv. Healthcare Mater. 9(1), 1901335 (2020).
-
Zhang, J. et al. Suppression of hypoxia-inducible factor 1α (HIF-1α) by tirapazamine is dependent on eIF2α phosphorylation rather than the mTORC1/4E-BP1 pathway. PLoS ONE 5(11), e13910 (2010).
-
Balakrishnan, P. et al. Formulation and in vitro assessment of minoxidil niosomes for enhanced skin delivery. Int. J. Pharm. 377(1–2), 1–8 (2009).
Funding
RJE is supported by Tabriz University of Medical Sciences, Tabriz, Iran, grant number 70309.
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
This study as well as cell line authentication and mycoplasma testing is done according to the protocol approved by the Biomedical Research Ethical Committee of Tabriz University of Medical Sciences, Tabriz, Iran. (IR.TBZMED.VCR.REC.1402.191).
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kaveh Zenjanab, M., Salemi, A., Doustmihan, A. et al. Dual-loaded niosome-dendrimer nanoplatform enhances Tirapazamine delivery to hypoxic breast cancer cells. Sci Rep 15, 29308 (2025). https://doi.org/10.1038/s41598-025-14704-7
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-14704-7