Introduction
Antimicrobial Resistance (AMR) is quickly becoming one of the most concerning global health problems, with 1-1.28 M deaths worldwide associated with AMR infections each year1,2. Studies have estimated that in a few decades AMR infections will be the main cause of non-natural death, increasing the yearly toll to 10 M deaths associated with AMR1,3 and causing an annual loss of 1% of the world GDP4. AMR is particularly alarming in the clinical setting due to the generalised use of antibiotics in hospitals, which expedites the evolution of antimicrobial resistant bacteria5. In the past decades, a growing number of studies has focused on studying AMR evolution in clinical bacteria6,7,8. Still, the vast majority of scientific studies on AMR evolution to date have been performed with laboratory bacterial strains, which don’t fully represent the complexity of clinical ones. We are therefore in dire need of more scientific studies performed in clinical strains, ones that are able to capture AMR in its full, real-world complexity9.
AMR can be achieved through different mechanisms10. Out of them, conjugative plasmids play an essential role in the spread of AMR among clinical Enterobacterales11. These extrachromosomal genetic elements often carry AMR genes and have the ability to be transferred horizontally to cells that are nearby, promoting the dissemination of AMR genes across bacterial populations12. Plasmids can be double-edged swords for the bacteria that carry them. On the one hand, they carry genes that can be useful in the right conditions (e.g., a specific AMR gene when a given antibiotic is prescribed to a patient). On the other hand, they alter the fine-tuned metabolism of the cell, often causing changes to the fitness of their bacterial hosts13. Multiple studies have shown plasmids to produce variable fitness effects across bacterial hosts11,14,15,16. These host-specific responses to carrying the same plasmid help explain why plasmids are frequently associated with a small fraction of hosts, regardless of their often wide host range.
Plasmid-associated fitness costs arise from genetic conflicts between the novel plasmid genes and the genetic background of the host17,18. The molecular mechanisms behind these interactions are multifactorial and poorly understood19,20. Plasmid’s costs can originate during the different phases of their life-cycle in the cell: from plasmid reception21, to replication22, gene expression23 and conjugation24. However, recent studies suggest that the main source of plasmid fitness costs are the interactions between plasmid-encoded proteins and the host bacterium25. For example, plasmids often encode for homologues of bacterial regulators which they can use to re-tune the regulatory networks of the bacterium to their advantage20,26. While beneficial to plasmid survival, these changes in metabolic flux might affect essential cellular processes and create a fitness cost for the host. Similarly, by encoding alleles of genes also present in the bacterial chromosome, plasmids might disturb the optimal protein concentration, leading to a fitness cost23. Finally, by encoding homologues of highly connected bacterial proteins, multiple metabolic processes might be affected at once in the host27. Several studies have centred on trying to understand how plasmid fitness costs play a role in plasmid evolution. A trade-off between horizontal and vertical transmission of the plasmid has been proposed: a very efficient plasmid conjugation machinery tends to be costly to the plasmid host14,28. A second trade-off between the expression of resistance genes and vertical transmission has also been proposed: in the absence of antibiotics the costs associated with plasmids have been shown to often arise from the expression of their resistance genes29,30,31. pOXA-48 is a highly-conjugative plasmid of great clinical relevance, which promotes the dissemination of the carbapenemase-encoding gene blaOXA-48 across clinical Enterobacterales32,33. Carbapenemases are able to hydrolyse carbapenems, which are used in hospitals as last-resort treatments of multidrug resistant (MDR) infections. Because of this, pOXA-48 dissemination in the nosocomial context represents a global public-health threat. Despite its order-wide host range, clinical data shows the plasmid to be strongly associated with specific high-risk Klebsiella pneumoniae clones (such as those belonging to sequence type ST1134,35).
The aim of this study is to uncover the origin and molecular basis of pOXA-48-associated fitness effects at the gene level. Specifically, we investigated the fitness effects associated with silencing each gene in the plasmid in different clinical strains of Enterobacterales using CRISPR interference screens (CRISPRi). Our results show that although carrying pOXA-48 leads to very different fitness responses in its different hosts, the gene-level effects are conserved amongst them. We confirm the importance of the toxin-antitoxin (TA) system PemI-PemK in the vertical transmission of the plasmid in clinical strains, and highlight the previously unknown role of plasmid-encoded modulators of expression in pOXA-48 stability in E. coli. Our data also suggests that pOXA-48 might be better adapted to K. pneumoniae than to Escherichia coli, which could explain its strong association in the clinic with the first species. Most importantly, we show that the carbapenemase OXA-48 is the main responsible for the fitness costs associated with the plasmid.
Results
Designing a pOXA-48-wide CRISPRi screen for clinical Enterobacterales
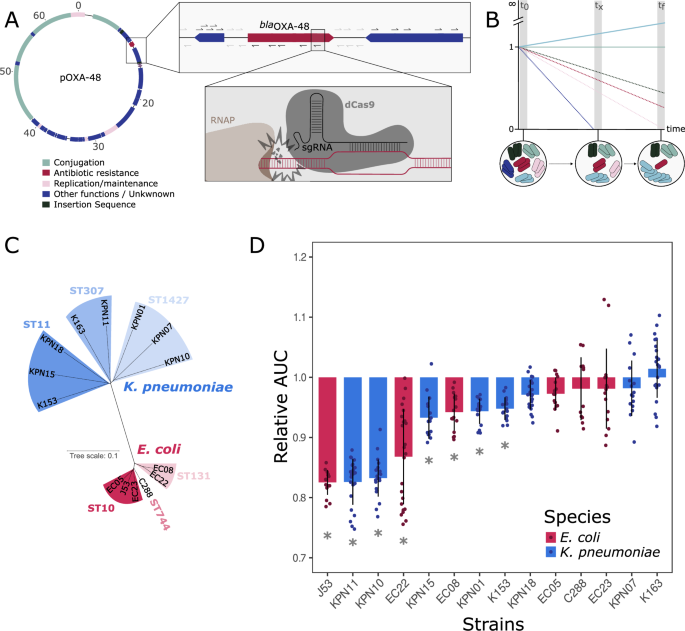
We started by designing and building a CRISPRi set-up to study the gene-level fitness effects of pOXA-48 in clinical strains of Enterobacterales. CRISPRi works by using dCas9, a DNA-binding protein that can be directed to virtually any region of DNA guided by a small RNA molecule named single-guide RNA (sgRNA). Once bound to DNA, dCas9 inhibits the expression of the targeted region (Fig. 1A). In CRISPRi screens, a pool of bacteria is built so that each individual bacterium carries dCas9 programmed to silence an individual gene through a specific sgRNA. This pool of bacteria can then be propagated, and the fitness effects of each gene in the given context can be determined through the change in frequency of gene-specific sgRNAs in the population (Fig. 1B). For the pOXA-48-wide CRISPRi screen, we designed 5 sgRNAs targeting each plasmid gene and up to 10 sgRNAs per intergenic region (depending on the size of the region, Supplementary Data 1). Each sgRNA was cloned in pFR56apm, a vector expressing the CRISPRi machinery under DAPG induction. The resulting 568-plasmid library was named OXAlib.

Depiction of the mechanism of CRISPRi screens and fitness costs associated with carrying pOXA-48 in the strains under study. A Schematic representation of CRISPRi gene-silencing and the set-up of CRISPRi screens. Values in pOXA-48 correspond to Kb. Each individual arrow represents an sgRNA targeting the region, and the orientation of the arrow indicates the orientation of dCas9 when binding to the DNA strand (RNAP stands for RNA polymerase). B Schematic representation of CRISPRi screens. Each colour corresponds to bacteria with a different pOXA-48 gene silenced (i.e., carries pFR56apm with an sgRNA programmed against said gene). Bacteria that carry a guide silencing a beneficial gene will decrease in frequency in the population during the screening, lowering the amount of the guide over time (e.g., red cells). On the other hand, bacteria carrying guides that silence costly genes will be enriched during the screening (e.g., light blue cells). C Unrooted phylogenetic tree of the Enterobacterales clinical strains used in this study. Multi Locus Sequence Types (STs) are indicated. D Distribution of pOXA-48 fitness effects (Relative Area Under the Growth Curve (see “Methods”), n = 16, error bars correspond to Standard Deviation) in the clinical strains selected for the CRISPRi screening. Asterisks indicate significant cost of pOXA-48 in each of the strains (two-sided paired t-tests against each relative plasmid-free strain after Bonferroni correction; n = 16; p < 0.05).
We selected a collection of 13 clinical strains of K. pneumoniae (n = 8) and E. coli (n = 5), representative of the diversity of MDR Enterobacterales isolated from hospitalised patients at a large hospital in Madrid35, to perform the CRISPRi screen. This collection included clones that are typically associated with pOXA-48 in our hospital and others, such as K. pneumoniae ST11, ST307 and E. coli ST1035 (Fig. 1C). As a control we also included E. coli J53, a lab strain derived from K1236. For more information on the strains used, consult the supplementary data (Supplementary Data 2).
We first determined the fitness effects of pOXA-48 (variant K8, GenBank MT441554, belonging to plasmid taxonomy unit L/M [IncL]) in each individual host, by comparing the growth of pOXA-48 carrying and pOXA-48-free isogenic versions of each strain (Fig. 1D). As previously described, the plasmid produced variable fitness effects in the 14 strains16,37.
One of the limitations of using CRISPRi in wild type, clinical strains is that the efficiency of CRISPRi silencing can vary between strains. These differences can be due to several factors, such as different levels of expression of the CRISPRi machinery in each strain, different guide strength for each guide in the library, or aspects intrinsic to the CRISPR machinery that vary from strain to strain. Therefore, prior to performing the CRISPRi screens, we confirmed the efficiency of dCas9-mediated silencing in all strains using two different approaches. First, we targeted the essential gene rpsL38 with a guide expressed from the pFR56apm vector and confirmed a strong reduction in the viability of all strains (SFig. 1). Second, we introduced a GFP-producing version of pOXA-48 (pOXA-48A_nuc::sfGFP) in each strain and measured the reduction in GFP expression associated with targeting the gfp gene by flow cytometry (SFig. 2). Our results confirmed that the CRISPRi machinery is highly active in all strains.
CRISPRi screens reveal conserved pOXA-48 gene-level effects across clinical strains
We introduced the OXAlib library in the 14 pOXA-48-carrying strains and performed the CRISPRi screens with continuous dCas9 expression for ~85 generations (72 h). The screens were performed in each strain individually, in triplicate, both without antibiotic pressure and in the presence of sub-inhibitory concentrations of ertapenem, to which pOXA-48 confers resistance. Due to the differences in MIC for the different strains, different concentrations of ertapenem were used for each strain (Supplementary Data 1). To determine the fitness effects of pOXA-48-encoded genes, we collected samples at 0, ~25, ~55 and ~85 generations and measured the fitness effect associated with silencing each gene individually. In CRISPRi screens, changes in the distribution of each sgRNA in the pool throughout time are used as proxy for the fitness effects associated with silencing the gene they target. These changes are represented as the log2 fold-change (log2FC) of each sgRNA in the population. For simplicity, we refer to log2FC as “genes scores” in the rest of this manuscript. The temporal gene score dynamics of each gene were analysed for each time-point (SFig. 3, 4). The data analysed in the rest of this paper corresponds to ~85 generations, as that’s when the effect of each sgRNA was most visible. In parallel, the stability in the population of both pOXA-48 and pFR56apm were also assessed (SFig. 5).
Despite the variable fitness effects produced by pOXA-48 across the clinical strains (Fig. 1D), we observed that silencing each pOXA-48 gene individually led to similar fitness effects across strains (Fig. 2A). To confirm this observation, we correlated the gene scores of all the genes and intergenic regions between all the strains per experimental condition. Pearson correlation indexes (r) resulted in highly positive values for most of the pairwise comparisons between strains both in absence and presence of ertapenem (0.25 < r < 0.90, median = 0.64 without ertapenem; −0.07 < r < 0.91, median = 0.66 with ertapenem. Figure 2B; Supplementary Data 3).

Representation of the fitness effects associated with silencing each gene in pOXA-48 individually in the presence and absence of antibiotic pressure, and how conserved those effects are amongst strains. A Heatmap of CRISPRi gene scores at the end of the screening (t = 72 h, ~85 generations). Fitness effects associated with silencing each individual gene / intergenic region in the absence of selection (top panel) and in the presence of ertapenem (bottom panel). The score for each gene / intergenic region corresponds to the median log2 fold-change of the sgRNAs targeting the element in the population (see “Methods”). Blue shades indicate those genes which silencing is detrimental (i.e., genes that are beneficial in that condition), whereas yellow shades correspond to genes which silencing is beneficial during the experiment (i.e., genes that are costly in that condition). Coding regions of pOXA-48 are shown with different colours indicating common functional clusters as shown on top. Genes and surrounding intergenic regions showing a significant deviation in the gene scores in both conditions are highlighted in the top. A fully annotated version of the figure is available in SFig. 7. B Pearson correlation matrices between strains’ gene scores at the end of the screening without (left) and with ertapenem (right). Blue shades indicate positive Pearson correlation coefficients, meaning similar tendencies in the overall screening results between the strains. Circle sizes indicate correlation absolute values from 0 to 1 (blue) or −1 to 0 (red).
Finally, we aimed to confirm that the overall gene-level effects revealed in CRISPRi screens were able to recapitulate the fitness effects produced by the entire pOXA-48 plasmid. Our results showed that the median gene scores of plasmid-targeting sgRNAs in the absence of ertapenem (compared to the control guides) anticorrelated with the relative AUC (Area Under the growth Curve) of pOXA-48-carrying strains (R = −0.73, P value = 0.0045). Therefore, silencing plasmid genes is more beneficial as the plasmid increases in cost. These results confirmed that our CRISPRi data is able to reflect overall plasmid fitness effects (SFig. 6).
Analysing pOXA-48 gene-level fitness effects
To identify which genes play a significant role in pOXA-48 fitness effects, we performed a permutation test to pinpoint the genes whose silencing led to statistically significant changes in frequency both in presence and absence of ertapenem (Fig. 3A). These genes were (i) blaOXA-48, the β-lactamase gene; (ii) repA, the replication initiation gene, and repC, transcriptional repressor involved in plasmid copy number control (in further experiments we study repA as representative of the repCBA locus); (iii) pri, which is involved in conjugation, and shows homology to DNA primases; (iv) korC, a transcriptional repressor with unknown function in pOXA-48; (v) a H-NS-like family regulator gene; (vi) an operon with unknown function formed by DNDJGHEP_0013 and _0014, predicted to be an ABC-transporter substrate binding protein and an Xre-like transcriptional regulator, respectively; and (vii) pemI, the antitoxin of the pemI-pemK system, responsible for post-segregational killing in case of plasmid mis-segregation39. In line with our previous results (Fig. 2, SFig. 7), the genes showing significant effects were highly conserved across strains (SFig. 8). Interestingly, the response was more generally conserved in K. pneumoniae than in E. coli (Welch two sample t-test, t = −2.1694 P value = 0.04234 without ertapenem; two-sided Kruskal-Wallis test, chi-squared = 5.6326, P value = 0.01763 with ertapenem; SFig. 8).

Identification of pOXA-48 genes producing significant fitness effects and their effect on plasmid stability. A Volcano plots obtained from two-sided permutation test assays in absence of selection (left) and in the presence of ertapenem (right). The Y axis shows the logarithmic transformation of the adjusted P value (FDR) from the permutation test, while the x axis indicates the median gene score (log2 fold-change values for sgRNAs targeting each gene from CRISPRi screens). Grey points indicate genes and intergenic regions which silencing was non-significant (i.e., −0.5 < gene score < 0.5; and P > 0.05). Genes and intergenic regions which silencing was detrimental in each condition are indicated in red, whereas genes which silencing was beneficial are coloured in blue. The labels of genes encoding hypothetical proteins are shown in grey. B pOXA-48 stability after 24 h of CRISPRi gene silencing of individual plasmid genes (X axis) in a subset of 5 strains (Y axis; 2 E. coli: EC22 and C288; and 3 K. pneumoniae: KPN18, KPN11 and KPN10) from our screenings (in presence of apramycin to select for pFR56apm). C pOXA-48 Copy Number measured silencing individual plasmid genes relative to control guide values for each strain tested (Log10 scale). Negative values indicate PCN decrease when silencing the indicated gene. Each point shows an individual replicate (n = 6). Bars correspond to median relative PCN values for all the replicates in each of the strains tested. Bars shaded in blue correspond to K. pneumoniae strains, and in pink to E. coli strains.
Our analysis detected genes that, when silenced, produced a significant advantage or disadvantage for the bacterial host. In the absence of ertapenem, silencing five of the genes led to a negative effect (pri, H-NS, DNDJGHEP_0013 and _0014, and pemI), while silencing of blaOXA-48 and repA led to a positive fitness effect (Fig. 3A, left panel). In the presence of ertapenem, silencing of all seven genes led to a detrimental effect (Fig. 3A, right panel).
Investigating the role of individual genes on plasmid stability
Our results showed that most of the genes producing a significant signal when silenced led to a fitness disadvantage. For blaOXA-48, the fitness disadvantage in the presence of ertapenem can be easily explained because the OXA-48 carbapenemase is responsible for the resistance phenotype. In the case of pemI, silencing the antitoxin gene probably leads to a PemK-mediated toxic effect (although this is likely mitigated by the polar silencing of pemK, since pemI-pemK form an operon40). The effect of PemK is analysed in depth in the following section. For the remaining genes, we hypothesised that the fitness defects could be due to a reduction in plasmid stability. Specifically, in the presence of ertapenem, plasmid loss would produce an obvious growth defect, since the cells would no longer produce the OXA-48 carbapenemase. Moreover, plasmid loss should also lead to a PemK-mediated post-segregational killing effect, and this should hold true both in presence and absence of ertapenem. Therefore, we decided to investigate the potential role of silencing these genes in pOXA-48 stability. We individually cloned the strongest sgRNA from the CRISPRi screen for each gene in pFR56apm. Then, we transformed the pFR56apm plasmids individually in a subset of the strains used in our screen, namely K. pneumoniae KPN10 (ST1427), KPN11 (ST307) and KPN18 (ST11), and E. coli C288 (ST744) and EC22 (ST131). Using these constructions we investigated how blocking the expression of each gene affected plasmid stability.
We blocked the expression of each gene individually over a 24 h growth cycle and measured the stability of pOXA-48 using replica plating (Fig. 3B). We confirmed the phenotypic results by measuring pOXA-48 copy number in the same populations by qPCR (Fig. 3C). Our results showed that silencing all the genes under study, except blaOXA-48, led to a reduction in pOXA-48 stability compared to the non-targeting control guide at least in one of the strains (one-tailed z-tests after Bonferroni adjustment, n = 5, p < 0.05). The most dramatic decrease in plasmid stability was mediated by silencing repA, which led to the complete curing of the plasmid in 4/5 of our strains. This was to be expected, given the role of RepA as the replication initiation protein of pOXA-48. Surprisingly, pri seems to play a similar role in plasmid stability as repA. This gene had been described as essential for conjugation in a similar plasmid to pOXA-48 belonging to the plasmid taxonomic unit L/M (IncM)41. However, its potential activity as a DNA primase could play a role in plasmid replication as well.
H-NS, korC and the DNDJGHEP_0013 and _0014 genes produced a stronger effect in pOXA-48 stability in E. coli than in K. pneumoniae. These results are further supported by a parallel study working with a transposon insertion library of pOXA-48 in E. coli MG1655, in which Baffert et al. showed that KorC and DNDJGHEP_0014 (orf20) are essential for plasmid stability42. The specific mechanisms by which these genes affect plasmid stability are difficult to assess. H-NS, korC and DNDJGHEP_0014 (which forms an operon with DNDJGHEP_0013 and is therefore also silenced by guides targeting this upstream gene) encode for transcriptional regulators. Further work will be needed to clarify the transcriptional effects associated with silencing these genes and how they impact plasmid stability. Interestingly, although silencing these genes produced a negative effect in most K. pneumoniae strains in our CRISPRi screens, they affected pOXA-48 stability to a lower extent in this second experiment. However, it is important to remember that the results of silencing individual genes were obtained after a single growth cycle (8-10 generations), while the CRISPRi screens were performed for 85 generations. Finally, and as expected, pemI produced a very modest effect on plasmid stability, and blaOXA-48 produced no effect at all.
PemK enhances vertical transmission of pOXA-48
Previous studies performed in laboratory E. coli strains had shown that PemK is responsible for the fitness defects associated with pOXA-48 loss39. We hypothesised that this effect should also hold true in MDR clinical strains of Enterobacterales. To test this hypothesis, we first cloned pemK in an expression vector under the control of the pTet promoter. We introduced this vector in K. pneumoniae KPN10 and E. coli EC22. Interestingly, even if the expression of pemK was not induced, we could not obtain viable transformants for KPN10 carrying the intact vector, suggesting that even a leaky expression of pemK is lethal in this strain. We tried to transform the vector in a different K. pneumoniae strain, KPN11, with the same result. For E. coli EC22, we could transform this vector, probably due to a tighter pTet repression in this species. Our results showed that pemK expression led to a dramatic growth defect in this strain (SFig. 9). These results confirm that PemK produces a toxic effect in our wild type clinical strains.
To further confirm the relevance of PemK in pOXA-48 stability, we constructed a version of pOXA-48 lacking the toxin gene pemK (pOXA-48∆pemK), and introduced it in K. pneumoniae KPN10, KPN11 and KPN18, and E. coli C288 and EC22. We performed plasmid stability assays for 5 days to determine the stability of both wt-pOXA-48 and pOXA-48∆pemK in the different strains (Fig. 4A). Wild-type pOXA-48 was fully stable in K. pneumoniae and in E. coli EC22, but it was slightly unstable in E. coli C288. pOXA-48∆pemK was surprisingly stable in K. pneumoniae, only starting to be lost in KPN11 and KPN18 after 4 and 3 days, respectively. Contrary, pOXA-48∆pemK was very unstable in E. coli, being lost up to 90% in EC22. Both pOXA-48 and pOXA-48∆pemK showed an erratic behaviour in E. coli C288. The fact that pOXA-48 does not produce a fitness cost in this strain, and therefore there is a small benefit associated with losing it, may help explain this behaviour (Fig. 1D). Together, these results suggest that pOXA-48 segregation might be more efficient in K. pneumoniae than in E. coli. This hypothesis will have to be tested in future studies.

The PemI-PemK TA system promotes pOXA-48 vertical transmission. A Stability of pOXA-48 (darker lines) and pOXA-48∆pemK (lighter dashed lines) in the different clinical strains over 5 days in LB culture (with a daily 1:1000 dilution). B pOXA-48 and pOXA-48∆pemK stability after 24 h of CRISPRi gene silencing of individual plasmid genes.
Finally, we decided to test the effect of silencing the three genes producing partial plasmid instability when silenced (korC, H-NS, and DNDJGHEP_0013-0014) in the pOXA-48∆pemK plasmid, including the repA guide as control. Our hypothesis was that if PemK is eliminating the cells where pOXA-48 is lost, once the pemK gene is removed, we should be able to observe a decrease in plasmid stability associated with silencing these genes. We performed these experiments in K. pneumoniae KPN10, KPN11 and E. coli EC22. In line with our hypothesis, we observed an increase in plasmid loss associated with silencing these three genes (Fig. 4B). These results confirmed the relevance of korC and the DNDJGHEP_0013-14 operon in pOXA-48 stability in both species. The H-NS gene, on the other hand, seems to produce a species-specific effect. This observation is supported by our previous results (Fig. 3B).
Taken together, our results underline the key role of the PemI-PemK TA system promoting pOXA-48 vertical transmission in clinical Enterobacterales.
bla OXA-48 is responsible for pOXA-48 fitness costs
Only two pOXA-48 genes produce a significant fitness benefit when silenced in the absence of ertapenem, repA and blaOXA-48. The results for repA are puzzling, since we have demonstrated that repA silencing produces a rapid plasmid loss, which leads to a toxic effect (Fig. 3B, C). In fact, when looking at the CRISPRi screen results, repA is the only gene with significant effect that changes in sign over time. Silencing repA is associated with a fitness defect in the first time point and it becomes beneficial later on (SFig. 10). We comment on these counterintuitive results in more depth in the discussion section. Results for blaOXA-48, on the other hand, are more straightforward. Previous reports had already indicated that blaOXA-48 expression was associated with a fitness cost in a clinical Enterobacterales strain31. Therefore, we decided to study blaOXA-48 effects in more detail.
First, we correlated the CRISPRi score of blaOXA-48 with the relative growth of pOXA-48-carrying strains, obtaining a strong negative relationship (Fig. 5 A, Pearson R-squared = −0.91, P value = 0.0016, for K. pneumoniae and Pearson R-squared = −0.88, P value = 0.02, for E. coli). This correlation was in fact stronger than the one obtained including the scores of all the genes in the CRISPRi screen (SFig. 6), as well as stronger than the correlation obtained with the subset of genes producing significant fitness effects (SFigs. 11, 12). Therefore, these results strongly suggest that the main contributor to pOXA-48 cost is the blaOXA-48 gene.

The blaOXA-48 gene is responsible for pOXA-48-associated fitness costs. A Two-sided Pearson correlation between the relative Area Under the growth Curve (AUC) of pOXA-48-carrying strains compared to their pOXA-48-free counterparts (shown in Fig. 1D) and the gene score (log2 fold change) associated with silencing blaOXA-48 in the CRISPRi screen. Lines indicate the best fit for the correlation of each species. Shadowed areas indicate the 95% CI. Each colour represents a species. B Plasmid cost associated with carrying pOXA-48 and pOXA-48∆blaOXA-48 in different clinical strains of Enterobacterales measured as relative AUC of pOXA-48-carrying compared to pOXA-48-free clones. Colours of strain names in the x-axis are again indicative of species: in pink E. coli, in blue K. pneumoniae, and in black C. freundii. Asterisks indicate significant cost/benefit of either pOXA-48 or pOXA-48∆blaOXA-48 in each of the strains (two-sided paired t-tests against each relative plasmid-free strain after Bonferroni correction; n = 8, except for KPN11, KPN10, KPN18, K163, EC22, C288 carrying pOXA-48, in which n = 16; p < 0.05; error bars correspond to Standard Deviation).
To experimentally validate these results, we used a version of plasmid pOXA-48 with a small deletion affecting the blaOXA-48 gene (pOXA-48∆blaOXA-48). This plasmid was recently recovered from a clinical strain and does not produce a functional OXA-48 carbapenemase, but is otherwise isogenic to pOXA-4831. pOXA-48∆blaOXA-48 has a small deletion of 199nt that comprises the full promoter region of blaOXA-48 and the first 36nt of the gene. Transcription of a truncated blaOXA-48 mRNA takes place, possibly from a secondary promoter, but levels of transcription are very low (between 1.8-8.4% of the wt blaOXA-48 expression) and does not lead to a resistant phenotype (SFig. 13). To further confirm that our previous results are generalisable, and not due to our choice of initial set of strains of work, we conjugated pOXA-48 and pOXA-48∆blaOXA-48 in a second set of clinical Enterobacterales including Klebsiella spp., E. coli, and Citrobacter freundii clones (Supplementary Data 2). As previously described16, pOXA-48 produced variable fitness effects across the collection (average relative AUC = 0.938, SD = 0.075), causing a significant reduction in relative AUC in 7 out of 13 strains (paired t-tests against each relative plasmid-free strain after Bonferroni correction; n = 8-16; p < 0.05, Fig. 5B). Interestingly, and in line with the hypothesis of blaOXA-48 being responsible for pOXA-48-associated fitness costs, pOXA-48∆blaOXA-48 produced milder fitness effects in this collection (average relative AUC = 1.002, SD = 0.059), leading to significant costs in 2 of the strains and to significant fitness advantages in 2 other strains (paired t-tests against each relative plasmid-free strain after Bonferroni correction; n = 8-16; p < 0.05, Fig. 5B). Taken together, these results strongly suggest that the main cause of the fitness cost produced by pOXA-48 is the expression of the blaOXA-48 gene.
Discussion
Unveiling the complex interactions between a plasmid and its bacterial hosts is essential for understanding plasmid ecology and evolution, and therefore for deciphering the dynamics of AMR in real-life scenarios, such as the hospital set-up or the patient’s gut. To better understand these interactions, in this work we study the fitness effect of each gene in pOXA-48, using 13 different clinically relevant clones of Enterobacterales as hosts. To our knowledge, this is the first study to date to perform such an extensive screen in clinical strains, which allowed us to discover clinically-relevant interactions that could be missed by performing the experiments in laboratory strains. Our results showed that although pOXA-48 produces a wide range of fitness effects across the clinical clones (Fig. 1), the effects of silencing each plasmid gene are overall conserved (Fig. 2). This difference is driven by the fact that most of the gene-level fitness effects detected were “post-segregational killing” deleterious effects, associated with plasmid loss (Figs. 3, 4). These effects emerge in the CRISPRi screen due to the silencing of genes associated with plasmid stability, but they do not necessarily contribute to the fitness effects of the wild-type plasmid with a correct partitioning. However, these results highlight the efficiency of plasmid-encoded mechanisms, such as the pemI-pemK TA system, promoting vertical plasmid transmission in clinical strains (Fig. 4). Concurrently, while these effects are qualitatively generalisable to all the strains, the degree to which each gene plays a role differs in each of the clinical strains. This could potentially be explained by the extensive pangenome of these strains (in particular their plasmidome, with each strain carrying very different sets of natural plasmids – whose gene-content could complement the silenced genes in a strain-to-strain manner). We claim that these observations highlight the need to perform genome-wide CRISPRi screens in clinical strains to better capture all potential genetic interactions in the cell. Finally, we could show that despite pOXA-48 carrying more than 90 genes and CDS, the fitness costs associated with this plasmid could be explained almost entirely by the expression of the carabapenemase gene blaOXA-48 (Fig. 5).
This study shows the potential of using CRISPRi screens to deepen our understanding of plasmid biology at a gene level. For example, we showed that apart from the previously known role in plasmid conjugation41, Pri plays a role in plasmid replication, likely as a primase (Fig. 3B, C). We also unveil the importance, particularly in E. coli, of three modulators of gene expression: KorC, DNDJGHEP_0014 and an H-NS-like protein. Individually silencing the genes coding for those proteins led to plasmid loss in E. coli (Fig. 3B, C). Plasmids are known to encode for transcription regulators that control the expression of genes on both the plasmid and the host chromosome, thus manipulating the host cell behaviour to their benefit. Examples of such plasmid-encoded regulators are VirBR, which promotes transmission of IncX3 plasmids by upregulating their Type IV secretion system43, RsmQ, which switches its host from a motile to a sessile lifestyle, thus promoting direct contact between individuals and promoting conjugation of the plasmid pQBR10320, or several plasmid-encoded H-NS-like proteins, which have been shown to, amongst other functions, control the extracellular matrix composition of its host to regulate biofilm formation44. The precise role of KorC, DNDJGHEP_0014 and the H-NS-like protein, more specifically the plasmid and/or host genes under their modulation, will be explored in more detail in the future. The importance of KorC and DNDJGHEP_0014 (orf20) for pOXA-48 stability was however also observed in E. coli MG1655 in an elegant parallel study by Baffert et al. working with a transposon insertion library of pOXA-4842. In this simultaneous study exploring the role of each plasmid gene in conjugation, the authors were not able to obtain pOXA-48 mutants disrupting korC. In addition, the authors showed that plasmid mutants disrupting DNDJGHEP_0014/orf20 were only stable when an intact copy of the gene was provided in trans. Baffert et al.’s results further support the hypothesis of these genes playing a role in pOXA-48 stability. Likewise, we observed that although deletion of the H-NS-like protein from pOXA-48 is possible, it has a strong effect on plasmid stability in E. coli (SFig. 14). We also show that, as previously described45, the PemI-PemK TA system promotes the vertical transmission of pOXA-48 to cell progeny. However, we observed that the role PemI-PemK play in vertical plasmid transmission varied among clones. pOXA-48∆pemK was stably transmitted to the progeny in the 3 K. pneumoniae strains tested but not in 2 E. coli strains (Fig. 4A). Given that our data indicated that PemK is toxic both in K. pneumoniae and E. coli, these results suggest that plasmid segregation may be more efficient in the first species, which could help to explain why pOXA-48 is more strongly associated with K. pneumoniae than E. coli in the clinic35. We hypothesise that this higher stability could be explained by the fact that pOXA-48’s plasmid copy number is higher in K. pneumoniae than in E. coli (K. pneumoniae, mean = 2.21 copies; E. coli mean = 1.41), as we recently showed11. Alternatively, this difference could be due to the partition machinery being more efficient in the first species than the latter, which would suggest that pOXA-48 is better adapted to the K. pneumoniae than E. coli. Finally, it could also be possible that additional TA systems in pOXA-48 may explain the differences in plasmid stability between species. In fact, Baffert et al. just reported the existence of a second TA system in pOXA-48, composed of a toxin, DqlB, and a non-coding RNA trans-acting antitoxin, agrB42. This system had not been previously annotated in pOXA-48 due to its small size, and is located in the intergenic region between repA and trbB. Because of the lack of annotation, we did not design guides specifically targeting this region in the OXAlib. However, three of our repA-trbB intergenic guides target the antitoxin agrB (SFig. 15A). In line with the results from Baffert et al., we observed a dramatic decrease in the gene score of the guides silencing the antitoxin throughout our CRISPRi screens (SFig. 15B-C), supporting the role of TrbB-agrB as a second TA system in pOXA-48. The effect associated with silencing agrB was however similar for K. pneumoniae and E. coli strains, so it did not help to explain the higher stability of pOXA-48 in K. pneumoniae.
In our CRISPRi screens only two genes produced a fitness benefit when silenced in the absence of antibiotic selection: repA and blaOXA-48. The results for repA are difficult to explain, and highlight some of the limitations of our set-up. On the one hand, it is possible that a small subpopulation of the cells that carried a repA guide survived an early pOXA-48 loss in the experiment, overcoming the PemK and Dlbq toxic effect. This subpopulation would be free of the cost associated with pOXA-48 carriage and could therefore proliferate in the pool of cells. This would explain the rapid initial decline of guides targeting repA in the population, and their subsequent enrichment in later time-points (SFig. 10). On the other hand, it has been shown that weak and unpredictable CRISPRi gene repression can arise from targeting CDSs under negative regulatory feedback loops46,47. Rep proteins are known to follow this type of self-regulation and this could also contribute to the seemingly erratic behaviour of guides targeting the repA locus48.
The main finding of this work is the fact that the gene encoding for the β-lactamase OXA-48 is the main contributor to pOXA-48’s fitness cost across clinical strains (Fig. 5). Β-lactamases had already been shown to have an effect on bacterial cell physiology and lead to a decrease in fitness30. These effects may have different origins. First, they could be produced by the β-lactamase’s signal peptide49, responsible for folding, stability and translocation of the protein to the periplasm50. By leading to a host-specific inefficient processing of the protein and subsequent envelope stress, the signal peptide effectively dictates the host-range of some β-lactamases51, which could explain why plasmids encoding for these genes are more costly in some hosts than others. Second, the fitness costs associated with β-lactamases have also been associated with their residual DD-endopeptidase activity, which leads to the hydrolysis of the peptide-cross bridge and alters the structure and stability of the cell wall52. This means that carrying plasmids such as pOXA-48 can lead to a severe destabilisation of the bacterial envelope which can have downstream repercussions, such as fitness costs and even collateral sensitivity to other antibiotics53,54. This also paints a somewhat hopeful picture about the dissemination of these plasmids in the population: since the main source of cost of the plasmid is the AMR gene itself, in the absence of antibiotic treatment the cells will be under pressure to either lose the plasmid or mutate blaOXA-48. The latter was already observed in a previous study analysing within-patient evolution of pOXA-48, where a small deletion affecting the blaOXA-48 gene and abolishing the AMR phenotype was associated with reduction of plasmid cost in the gut microbiota of a hospitalised patient31. Lastly, this also opens a gateway to explore new ways to exploit these changes in the cell envelope as potential new antimicrobial strategies, ones highly specific to cells carrying genes within the β-lactamase family53,55.
In summary, our study highlights the role of different plasmid-encoded genes in the biology of pOXA-48 and opens new doors to explore alternative ways to fight AMR in Enterobacterales. Our results also highlight the usefulness of using high-throughput techniques to explore previously unknown biological phenomena at a genetic level. And lastly, given that previous works indicate that both the fitness and transcriptomics response to carrying plasmids are highly host-specific26,56,57, our study paves the way to performing genome-wide CRISPRi screens in order to dissect plasmid-bacteria interactions in clinical bacteria. Set-ups such as this, although complex and labour-intensive, will help us unveil aspects of plasmid biology with applications in real-life scenarios.
Methods
Bacterial strains and culture conditions
All clinical strains used in this study were obtained from the previously characterised R-GNOSIS collection of over 28,000 rectal samples collected from patients admitted to the Hospital Universitario Ramón y Cajal in Madrid between the years of 2014 and 2016 (R-GNOSIS-FP7-HEALTH-F3-2011-282512)).
For the CRISPRi screens, a subset of these strains was selected based on previous works. First, a collection of strains that did not carry pOXA-48 naturally and where pOXA-48 transconjugants had been obtained37, and, secondly, a collection of strains that carried pOXA-48 naturally and where pOXA-48 had been successfully cured31. Out of these two collections, strains were selected based on sensibility to apramycin – antibiotic resistance gene used for selection of the OXAlib library -, conjugation rate efficiency, belonging to a sequence type of clinical relevance, their plasmid profile and, lastly, activity of dCas9 – tested by measuring cell death by silencing the essential gene rpsL, as in Cui et al38. and sfGFP (Supplementary Methods, SFig. 1, 2)). The resulting selection comprises 14 strains, 8 K. pneumoniae strains and 6 E. coli strains (including the lab strain J53) (Supplementary Data 2). The clinical strains belong to STs of clinical relevance, namely K. pneumoniae ST1427 (KPN01, KPN07 and KPN10), ST11 (KPN15, KPN18 and K153) and ST307 (KPN11 and K163), and E. coli ST10 (EC23), ST131 (EC08 and EC22), ST744 (C288) and ST4669 (EC05).
The variant of pOXA-48 used in this work is pOXA-48_K8 (accession number MT441554), and its sequence and complete genomes of the strains used in this study were obtained by the lab in previous works, available under the BioProjects PRJNA641166 and PRJNA803387 in the SRA repository of the National Centre for Biotechnology Information (NCBI).
All experiments were performed in Lennox lysogeny broth (LB), supplemented with 15 g/L of agar when indicated. The different antibiotics used throughout the study were amoxicillin / clavulanic acid at a proportion of 5 mg amoxicillin / 1 mg clavulanic acid, ertapenem, and apramycin. 50 µM of 2,4-diacetylphloroglucinol (DAPG) was used for induction of dCas9.
Calculation of pOXA-48 fitness effects in different clinical strains
The fitness effect associated with pOXA-48 in each strain was calculated by comparing differences in the Area Under the Curve (AUC) of isogenic plasmid-carrying and plasmid-free clones, as described in DelaFuente et al.58. Raw data can be found in the supplementary materials (Supplementary Data 4).
Designing the OXAlib library
dCas9 is reprogrammed to target a specific position using guide RNAs (sgRNAs), the design of which needs to follow a specific set of rules in bacteria59. (i) First, it has been shown that the specific sequence of the sgRNA has an effect on the activity of dCas9. (ii) Due to design limitations intrinsic to the CRISPRi technique, sgRNAs targeting coding regions have to be limited to the coding strand. The rules for efficiently targeting non-coding regions of DNA are not known, and therefore sgRNAs targeting those regions should be designed to target both strands of DNA. (iii) If dCas9 is directed to a gene within an operon, this binding will have a polar effect on the downstream genes. (iv) It is also important to avoid potential off-targets resulting from a permissiveness of dCas9 to bind to similar regions of DNA to bona fide targets.
The OXAlib library was designed using the CRISPR@Pasteur design tool (https://crispr-browser.pasteur.cloud/) which is trained on E. coli activity data, and which has been successfully used to design such libraries in different species of Enterobacterales59. The tool was adapted to our setup (looking for targets on a plasmid and for off-targets on a set of genomes – see Code Availability). Using this tool, the 5 best-performing sgRNAs were selected for each gene in pOXA-48_K8 (called pOXA-48 in this paper). sgRNAs targeting both DNA strands were also designed against intergenic regions of the plasmid (up to 5 targeting each strand, depending on the size of the intergenic region). The logic behind targeting both strands in intergenic regions is to attempt to capture effects of targeting any possible genetic elements present in the regions. 20 non-targeting control sgRNAs were also designed, amounting to a total of 568 sgRNAs (Supplementary Data 1).
Guides were selected with the previously described sorting strategy and ordered for synthesis as ssDNA oligos from Twist Bioscience. The sequence of each individual oligo is as follows: 5’-ACAGCTAGCTCAGTCCTAGGTATAATACTAGT-(20nt guide sequence)-GTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGT-3’.
Cloning the OXAlib library
The ssDNA oligo pool was amplified using primers AC74 and AC75 (Supplementary Data 2) for subsequent cloning into pFR56apm26. pFR56apm is based on pFR5660, with an apramycin resistance gene as a selection marker. This antibiotic permits selection of the vector in all the different clinical strains (which due to their plasmidsome are naturally resistant to several antibiotics). Amplification was performed using New England Biolabs’ Phusion High-Fidelity DNA polymerase, with the HF buffer, DMSO and the recommended protocol. To minimise PCR amplification biases of specific oligos in the pool, the reaction was limited to 9 cycles (1 cycle for generating dsDNA oligos and 8 cycles of amplification), and performed with 1 ng of template and 50 pmol of each primer. The resulting product was gel purified. The pFR56apm vector was digested with BsaI and gel purified. The OXAlib plasmid library was assembled using the Gibson Assembly method61, using New England Biolabs’ NEBuilder HiFi DNA Assembly, 200 ng of total DNA and a ratio of 1:5 vector:insert. Cloning was performed in E. coli MG1655. Quality control of the final OXAlib library can be found in the Supplementary Methods.
Conjugating the OXAlib library into clinical strains
pFR56apm is a mobilisable plasmid – meaning it can be conjugated to compatible hosts, but requires expression of the conjugation machinery in trans. The OXAlib library was minipreped from E. coli MG1655 and transformed (with a coverage > 1000x of OXAlib) into E. coli ß3914ø + pTA-MOB62,63. This strain is auxotrophic for diaminopimelic acid (DAP), allowing for its counter-selection as a donor strain.
Conjugation of the OXAlib from ß3914ø to the different strains was performed by mixing 108 donor cells with 108 of recipient cells (ensuring a coverage > 1000x of OXAlib) for 3 h. After conjugation, the cells were selected in apramycin (50 µg/mL) and let grow overnight at room temperature. Absence of DAP counter-selects against ß3914ø. Conjugation efficiency was assessed using serial dilutions and a good coverage of the OXAlib library achieved for each strain (Supplementary Data 1).
CRISPRi screens
Experiment
First of all, it is crucial when working with CRISPRi screens to ensure that each step guarantees a good coverage of the library (i.e., that no individual guides are lost from the population due to over-dilution). To ensure this, we always work with an excess of cells of at least 1000x the size of OXAlib.
Strains carrying the OXAlib library were diluted 1:1000 in fresh LB / LB supplemented with ertapenem (subinhibitory concentrations of the antibiotics were used, Supplementary Methods, Supplementary Data 1). After 1 h, DAPG (50 µM) was added to the cultures for dCas9 induction. Every 8 h, a 1:1000 dilution was performed in fresh LB + DAPG / LB + DAPG + ertapenem. Prior to each dilution, OD was measured (to calculate number of generations). At 24, 48 and 72 h, apart from the dilution and OD measurement, a miniprep was performed using 10 mL of culture. The resulting pool of plasmids correspond to the timepoints at ~25, ~55 and ~85 generations. All screens were performed in triplicate (3 biological replicates), in aerobic conditions, at 37 °C, with shaking (250 r.p.m.).
Simultaneously, to assess whether the CRISPRi machinery encoded in pFR56apm is still active after 85 generations of growth, we grew the different strains harbouring pOXA-48::sfGFP and pFR56apm loaded with an sgRNA targeting sfGFP for 85 generations with induction of the CRISPRi machinery (DAPG 50 µm), then diluted the cultures in LB / LB supplemented with DAPG, grew the resulting sub-cultures overnight and measured sfGFP expression using cytometry. Our results suggest that the CRISPRi machinery is fully active until the end of the CRISPRi experiments performed in this manuscript (SFig. 16).
Illumina sample preparation and sequencing strategy
Library sequencing was performed as previously described38, with some small adaptations. The PCR primers used for the nested PCR can be found in the supplementary materials (Supplementary Data 2). To maintain good coverage of the OXAlib library, 100 ng of plasmid miniprep were used as template for each reaction. To minimise PCR biases, 19 cycles were used in the 1st PCR, and 13 cycles on the 2nd. Each replicate was given a different i5-i7 index combination, achieved by amplification with specific pairs of primers. Lastly, Illumina sequencing quality depends strongly on the variability of the regions sequenced. To bypass variability issues arising from amplicons sharing a large 5’ identical sequence, 3 different staggered primers were used in the 1st PCR.
Each sample was quantified using a bioanalyzer. 150 ng of each sample were pooled together and sequenced using the standard protocol of a NextSeq 500 benchtop sequencer (Illumina).
Data analysis
Quality control of Illumina sequencing
First, a QC of the raw Illumina data sequences (55 bp amplicon sequencing) was performed using FastQC v.0.11.964 and MultiQC v.1.1165. After confirming high quality per base score in the guide region (mean Phred score > 30), the sequences were used in downstream analyses.
Guide count per sample
For getting each individual guide count in each sample, each fastq file was parsed using Bash (see Code Availability). Briefly, knowing the sequence of the promoter preceding the guides in each read, 15 bp of it were used to match the reads position in which the guide began, and the following 20 bp (i.e., the sgRNA) were extracted. For each sample, the number of times each individual guide appeared was calculated using Bash commands. The list of guides was sorted, each unique occurrence was counted, and the output was reformatted to save each strain guides count in an individual csv file (see Source Data). Within the individual csv counts files, the guides with less counts than 5 for timepoints 0 and with less counts than 2 for the rest of the timepoints were filtered. The results of all the samples were merged in the same table, and only those guides present in the plasmid library were kept. The sequencing coverage for each guide was calculated as the median of the counts of guides (1 sgRNA per read; avoiding outliers from guides counted in excess) multiplied by 5 (as there are 5 guides per gene). The counts for the three replicates per time point were merged after checking the reproducibility of the replicates in each condition (LB + DAPG or LB + DAPG+ertapenem).
Gene score calculation
For calculating the gene scores (median log2 fold change by gene) a script which took the table with the replicates merged by sample, and calculated the log2FC for each gene, then centreing the values by removing the median of control guides, was written based on the code of Rousset et al.60 (https://gitlab.pasteur.fr/dbikard/ecocg/-/blob/master/Notebook.ipynb).
Phylogenetic analysis
The phylogenetic distances between the different strains were calculated using Mashtree v.1.2.066 (NJ method) and visualised using the iTOL web server (https://itol.embl.de/). Phylogenetic information was used to represent the strains clustered by relatedness in the heatmap (Fig. 2A).
Conservation of CRISPRi screening results in different strains
To analyse whether the effect of the OXAlib library on the strains tested was conserved, we studied the similarity between strains gene scores at the end of the screening. We calculated the correlation values (Pearson correlation coefficients) between the strains based on all the gene scores at the end of the screening for each condition by using the corrplot package v.0.95 in R. Additionally, we compared the genes whose silencing had the most notable effect in each strain. To do so, we listed the significant genes and intergenic regions for each strain and condition calculated from individual permutation tests for either LB + DAPG or LB + DAPG+ertapenem (see following Methods section). With these, we measured the Sørensen-Dice Similarity Coefficient (SDC) for all pairwise comparisons between strains of the same species screened in the same condition. The SDC was calculated using the following formula:
$$S{DC}=frac{2{{{rm{cdot }}}}|{G}_{x}{,cap G}_{y}|}{|{G}_{x}|+|{G}_{y}|}$$
Where |Gx| and |Gy| are respectively the number of significant genes in strain X and Y (i.e. the number of elements compared from each strain), and |Gx ∩ Gy| is the number of common significant genes between X and Y strains in the given condition (i.e. full set of significant genes for both strains in either ertapenem or no antibiotics).
Permutation tests for gene importance
For checking statistically significant differences from the score of each gene from the distribution of gene scores in the CRISPRi screen, a permutation test for all the strains considered together per each condition (LB + DAPG or LB + DAPG + ERTA) was carried out. Briefly, the mean of each gene score was calculated considering all the strains from the screens (n = 14). The difference of this value and the mean of the scores from the rest of the group was assessed as the statistics of interest in the permutation test. The same statistic was calculated for 100000 iterations (i.e., minimum computable P value = 0.00001) taking random samples (n = 14) in each iteration. This number of permutations was selected to get more precise probabilities, as the P values of some genes resulted marginally significant when performing the permutation tests with fewer repeats (1000−10000). Lastly, the number of times that the permuted statistic was more extreme than the mean of each gene (i.e., the P value) was calculated. The same test was conducted for each species, as well as for each strain, for both experimental conditions. Finally, the p-values were adjusted by FDR. To report the statistical results from the permutation tests as well as the CRISPRi screening results, Volcano Plots were represented to simultaneously show each gene P value and its score using the R package EnhancedVolcano v.1.18.0. As in this type of screen, most genes showed scores close to 0. Thus, a log2FC low threshold (i.e., +−0.5) was set, close to the centre of the distribution. For the P value threshold, the default one (10e-06; i.e., P value < 0.05) was selected.
Ridge regression model for plasmid cost
To simultaneously consider the contribution of multiple genes to pOXA-48 cost, a multiple regression model with Ridge regularisation was developed using the glmnet v.4.1-7 R package separately for both E. coli and K. pneumoniae. First, feature selection of all the genes and intergenic regions tested was performed, as there were 8 observations of plasmid cost for K. pneumoniae and 6 observations of plasmid cost for E. coli (n=strains), and 106 explanatory variables (n=genes & intergenic regions included in CRISPRi screen). Then, only those genes with significant P values in the permutation test performed for each of the species (i.e., expected to be more informative) in absence of antibiotics during the CRISPRi screen were included in the model. From these, intergenic regions were kept out, as their effect introduced multicollinearity issues by showing similar behaviour as their surrounding genes in the CRISPRi screening (e.g., blaOXA-48 intergenic surrounding regions). Multicollinearity between explanatory variables was assessed after feature selection using the R package corrplot v.0.95. Due to the limited number of observations for each species, the models were trained using Leave-One-Out Cross Validation. Moreover, to include all the genes resulting from the feature selection process, Ridge regularisation was selected, as the regression coefficients are not shrunk to 0. Optimal parameters from the trained models were selected to then build the final models. The ridge coefficients for each gene, and the R-squared for each species were calculated from the final models tuned with the optimal parameters (lambda [λ]; SFig. 8).
Plasmid cost based on guide distribution
To infer the plasmid cost in each strain, a proxy based on the general effect of blocking the plasmid genes was employed. The effect of each plasmid gene on bacterial fitness is expected to be reflected on the CRISPRi screen in absence of antibiotics, as guides which block costly genes that cause deleterious effects are expected to show higher scores, and vice versa. Then, if the plasmid is highly costly in a strain, the general effect of blocking its genes should reflect this. Thus, the general effects of blocking the plasmid genes were correlated with the relative fitness of plasmid-carrying strains. The difference between the mean effect of blocking pOXA-48 genes (mean CRISPRi gene scores for each strain) and the mean effect of the control guides, which are supposed to have no effect, was calculated. Then, these values were correlated (Spearman method) with the fitness effect of pOXA-48 in each strain.
Validation of CRISPRi screen’s results
Strain selection
From the initial set of strains used in this study, two E. coli (C288, EC22) and three K. pneumoniae strains (KPN10, KPN11 and KPN18) were chosen for validation of results based on the overall strongest fold-changes observed.
Cloning of individual sgRNAs in pFR56apm and conjugation into clinical strains
Cloning of individual sgRNAs (Supplementary Data 1) in pFR56apm was performed using Golden Gate assembly as in Rousset et al.60. All cloning was performed in E. coli MG1655 and followed by conjugation into the clinical strains as described previously.
blaOXA-48 associated fitness effects
To validate the results associated with silencing the gene blaOXA-48, growth curves were performed to study plasmid fitness effects as explained before in this method section. This time, a second set of strains was selected from the RGNOSIS collection, to avoid overfitting to the original strains used in the screen. These strains were once again selected based on the fitness effects associated with the plasmid, as described in Fernández-Calvet et al.16. A variant of pOXA-48 lacking a complete blaOXA-48 was identified previously in the lab31. This variant was conjugated on the cured version of the strain, obtaining three isogenic set-ups for each strain: the wild-type carrying pOXA-48, a pOXA-48-free cured version, and a pOXA-48∆blaOXA-48 transconjugant.
Plasmid stability
Plasmid stability of pOXA-48 and pOXA-48 versions was measured by growing cells carrying the plasmid in LB over subsequent periods of 24 h. After each cycle, cells were plated in LB, guaranteeing the isolation of individual colonies. Cells were then streaked in presence and absence of ertapenem using replica plating. Percentages of plasmid loss / plasmid presence were calculated by comparing growth in presence and absence of selection. Each experiment was performed in triplicate. Number of 24 h cycles used in each experiment are indicated throughout the text. To statistically analyse whether the stability of pOXA-48 was significantly affected in at least one strain when silencing individual genes (Fig. 3B), we performed a one-tailed z-test for each guide. Briefly, we calculated the maximum deviation from the control group (guide “control”) per individual guide, from which we inferred their Z-score. Finally, we corrected the P values by Bonferroni adjustment (n = 8).
qPCR for PCN calculation
Changes in pOXA-48’s PCN were calculated for the different clinical strains carrying pFR56apm programmed to silence different plasmid genes. In order to do so, the different strains were grown in triplicates in LB supplemented with apramycin 50 µg/mL and DAPG 50 µM for 24 h. 50 µL samples were taken of each replicate, centrifuged to collect the pellet and boiled for 10 min. 2 µL of each boiling reaction were used per qPCR reaction, using the NZYSupreme qPCR Green Master Mix (2X), ROX plus kit from NZYTech. Targeted plasmid and chromosome genes were blaOXA-48 (amplicon size 100 bp; efficiency 107,47%) and dnaE (chromosomal gene with 1 copy, amplicon size 200 bp, efficiency 105.79%), respectively. Efficiency was calculated (taking into account amplicon size difference) as in San Millán et al.67. The amplification conditions were: 5 min of initial denaturation (95 °C), followed by 30 cycles of 15 s denaturation, 30 s annealing (55 °C) and 30 s extension (60 °C). The relative PCN was calculated as in San Millán et al.67. Raw data for qPCR can be found in the supplementary data (Supplementary Data 4).
Lambda-Red recombination for construction of deletion mutants
A version of pOXA-48 lacking pemK was built using lambda-red recombination68. The recombination cassette with extremes matching the beginning and end of pemK, and a kanamycin gene flanked by FRT sides was amplified from pKD4, as described in Datsenko et al.68. Primers used for construction of the cassette and validation of the construct can be found in the supplementary data (Supplementary Data 2).
Statistics & reproducibility
All statistical analyses were performed using RStudio v.2024.12.1 + 563 and R v.4.3. Parametric or non-parametric tests were used depending on the structure of the data (see each specific subsection analysis). Tests were controlled for multiple comparison testing by applying FDR or Bonferroni adjustment (see each specific subsection analysis). To analyze correlation data, we used Pearson correlation for normally distributed data. No statistical method was used to predetermine sample size. No data were excluded from the analyses. The experiments were not randomised. The Investigators were not blinded to allocation during experiments and outcome assessment.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The sequences generated during this project are available under the BioProject ID PRJNA1203587 in the SRA repository of the National Centre for Biotechnology Information (NCBI). Source data for all figures is provided with this paper (see Supplementary Data 3 and 4, Source Data, Supplementary Information). Source data are provided with this paper.
Code availability
All the code developed for the analyses included in this work can be found at the Github repository69.
References
-
Review on Antimicrobial Resistance. Antimicrobial Resistance – Tackling a Crisis for the Health and Wealth of Nations. (2014).
-
Naghavi, M. et al. Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. Lancet 404, 1199–1226 (2024).
-
Ho, C. S. et al. Antimicrobial resistance: a concise update. Lancet Microbe 0, 100947 (2024).
-
World Bank. Drug-Resistant Infections: A Threat to Our Economic Future. (2017).
-
Baquero, F. et al. Evolutionary Pathways and Trajectories in Antibiotic Resistance. Clin. Microbiol. Rev. https://doi.org/10.1128/CMR.00050-19 (2021).
-
Song, S. et al. Adaptive evolution of carbapenem-resistant hypervirulent Klebsiella pneumoniae in the urinary tract of a single patient. Proc. Natl. Acad. Sci. USA. 121, e2400446121 (2024).
-
Dorado-Morales, P., Garcillán-Barcia, M. P., Lasa, I. & Solano, C. Fitness cost evolution of natural plasmids of staphylococcus aureus. mBio 12, https://doi.org/10.1128/mbio.03094-20 (2021).
-
Hernando-Amado, S. et al. Ciprofloxacin resistance rapidly declines in nfxB defective clinical strains of Pseudomonas aeruginosa. Nat. Commun. 16, 4992 (2025).
-
Souque, C., Ojeda, I. G. & Baym, M. From petri dishes to patients to populations: scales and evolutionary mechanisms driving antibiotic resistance. Annu. Rev. Microbiol. 78, 361–382 (2024).
-
Smith, W. P. J., Wucher, B. R., Nadell, C. D. & Foster, K. R. Bacterial defences: mechanisms, evolution and antimicrobial resistance. Nat. Rev. Microbiol. 21, 519–534 (2023).
-
Alonso-del et al. Antimicrobial resistance level and conjugation permissiveness shape plasmid distribution in clinical enterobacteria. Proc. Natl. Acad. Sci. 120, e2314135120 (2023).
-
Smillie, C., Garcillán-Barcia, M. P., Francia, M. V., Rocha, E. P. C. & de la Cruz, F. Mobility of plasmids. Microbiol. Mol. Biol. Rev. 74, 434–452 (2010).
-
San Millan, A. & MacLean, R. C. Fitness costs of plasmids: a limit to plasmid transmission. Microbiol. Spectr. 5, 10 (2017).
-
Porse, A., Schønning, K., Munck, C. & Sommer, M. O. A. Survival and evolution of a large multidrug resistance plasmid in new clinical bacterial hosts. Mol. Biol. Evol. 33, 2860–2873 (2016).
-
Hall, J. P. J. et al. Plasmid fitness costs are caused by specific genetic conflicts enabling resolution by compensatory mutation. PLOS Biol. 19, e3001225 (2021).
-
Fernández-Calvet, A. et al. The distribution of fitness effects of plasmid pOXA-48 in clinical enterobacteria. Microbiology 169, 001369 (2023).
-
Baltrus, D. A. Exploring the costs of horizontal gene transfer. Trends Ecol. Evol. 28, 489–495 (2013).
-
Billane, K., Harrison, E., Cameron, D. & Brockhurst, M. A. Why do plasmids manipulate the expression of bacterial phenotypes?. Philos. Trans. R. Soc. B Biol. Sci. 377, 20200461 (2021).
-
Hall, R. J., Snaith, A. E., Thomas, M. J. N., Brockhurst, M. A. & McNally, A. Multidrug resistance plasmids commonly reprogram the expression of metabolic genes in Escherichia coli. mSystems 9, e01193–23 (2024).
-
Thompson, C. M. A. et al. Plasmids manipulate bacterial behaviour through translational regulatory crosstalk. PLOS Biol. 21, e3001988 (2023).
-
Prensky, H., Gomez-Simmons, A., Uhlemann, A.-C. & Lopatkin, A. Conjugation dynamics depend on both the plasmid acquisition cost and the fitness cost. Mol. Syst. Biol. 17, e9913 (2021).
-
Petrova, V., Chitteni-Pattu, S., Drees, J. C., Inman, R. B. & Cox, M. M. An SOS inhibitor that binds to free RecA protein: the PsiB protein. Mol. Cell 36, 121–130 (2009).
-
Park, C. & Zhang, J. High expression hampers horizontal gene transfer. Genome Biol. Evol. 4, 523–532 (2012).
-
Koraimann, G. & Wagner, M. A. Social behavior and decision making in bacterial conjugation. Front. Cell. Infect. Microbiol. 4, 54 (2014).
-
Toribio-Celestino, L. & Millan, A. S. Plasmid–bacteria associations in the clinical context. Trends Microbiol. 0, https://doi.org/10.1016/j.tim.2025.04.011 (2025).
-
Toribio-Celestino, L. et al. A plasmid-chromosome crosstalk in multidrug resistant enterobacteria. Nat. Commun. 15, 10859 (2024).
-
Cohen, O., Gophna, U. & Pupko, T. The complexity hypothesis revisited: connectivity rather than function constitutes a barrier to horizontal gene transfer. Mol. Biol. Evol. 28, 1481–1489 (2011).
-
Turner, P. E., Cooper, V. S. & Lenski, R. E. Tradeoff between horizontal and vertical modes of transmission in bacterial plasmids. Evol. Int. J. Org. Evol. 52, 315–329 (1998).
-
Vogwill, T. & MacLean, R. C. The genetic basis of the fitness costs of antimicrobial resistance: a meta-analysis approach. Evol. Appl. 8, 284–295 (2015).
-
Rajer, F. & Sandegren, L. The role of antibiotic resistance genes in the fitness cost of multiresistance plasmids. mBio https://doi.org/10.1128/mbio.03552-21 (2022).
-
DelaFuente, J. et al. Within-patient evolution of plasmid-mediated antimicrobial resistance. Nat. Ecol. Evol. 6, 1980–1991 (2022).
-
Poirel, L., Bonnin, R. A. & Nordmann, P. Genetic features of the widespread plasmid coding for the carbapenemase OXA-48. Antimicrob. Agents Chemother. 56, 559–562 (2012).
-
Rozwandowicz, M. et al. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 73, 1121–1137 (2018).
-
David, S. et al. Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 4, 1919–1929 (2019).
-
León-Sampedro, R. et al. Pervasive transmission of a carbapenem resistance plasmid in the gut microbiota of hospitalized patients. Nat. Microbiol. 6, 606–616 (2021).
-
Matsumura, Y., Peirano, G. & Pitout, J. D. D. Complete genome sequence of Escherichia coli J53, an azide-resistant laboratory strain used for conjugation experiments. Genome Announc. https://doi.org/10.1128/genomea.00433-18 (2018).
-
Alonso-del et al. Variability of plasmid fitness effects contributes to plasmid persistence in bacterial communities. Nat. Commun. 12, 2653 (2021).
-
Cui, L. et al. A CRISPRi screen in E. coli reveals sequence-specific toxicity of dCas9. Nat. Commun. 9, 1–10 (2018).
-
Reuter, A. et al. Targeted-antibacterial-plasmids (TAPs) combining conjugation and CRISPR/Cas systems achieve strain-specific antibacterial activity. Nucleic Acids Res 49, 3584–3598 (2021).
-
Tsuchimoto, S. & Ohtsubo, E. Autoregulatiou by cooperative binding of the PemI and PemK proteins to the promoter region of the pem operon. Mol. Gen. Genet. MGG 237, 81–88 (1993).
-
Dmowski, M., Gołębiewski, M. & Kern-Zdanowicz, I. Characteristics of the conjugative transfer system of the IncM plasmid pCTX-M3 and Identification of its putative regulators. J. Bacteriol. https://doi.org/10.1128/jb.00234-18 (2018).
-
Baffert, Y. et al. Genetic determinants of pOXA-48 plasmid maintenance and propagation in Escherichia coli. Preprint at bioRxiv https://doi.org/10.1101/2025.01.23.634512 (2025).
-
Ma, T. et al. VirBR, a transcription regulator, promotes IncX3 plasmid transmission, and persistence of blaNDM-5 in zoonotic bacteria. Nat. Commun. 15, 5498 (2024).
-
Benomar, S., Di Venanzio, G. & Feldman, M. F. Plasmid-encoded H-NS controls extracellular matrix composition in a modern Acinetobacter baumannii urinary isolate. J. Bacteriol. https://doi.org/10.1128/jb.00277-21 (2021).
-
Tsuchimoto, S., Ohtsubo, H. & Ohtsubo, E. Two genes, pemK and pemI, responsible for stable maintenance of resistance plasmid R100. J. Bacteriol. 170, 1461–1466 (1988).
-
Vigouroux, A., Oldewurtel, E., Cui, L., Bikard, D. & van Teeffelen, S. Tuning dCas9’s ability to block transcription enables robust, noiseless knockdown of bacterial genes. Mol. Syst. Biol. 14, e7899 (2018).
-
Rousset, F. et al. Genome-wide CRISPR-dCas9 screens in E. coli identify essential genes and phage host factors. PLOS Genet 14, e1007749 (2018).
-
del Solar, G., Giraldo, R., Ruiz-Echevarría, M. J., Espinosa, M. & Díaz-Orejas, R. Replication and control of circular bacterial plasmids. Microbiol. Mol. Biol. Rev. 62, 434–464 (1998).
-
Marciano, D. C., Karkouti, O. Y. & Palzkill, T. A fitness cost associated with the antibiotic resistance enzyme SME-1 β-lactamase. Genetics 176, 2381 (2007).
-
Socha, R. D., Chen, J. & Tokuriki, N. The molecular mechanisms underlying hidden phenotypic variation among metallo-β-lactamases. J. Mol. Biol. 431, 1172–1185 (2019).
-
López, C., Ayala, J. A., Bonomo, R. A., González, L. J. & Vila, A. J. Protein determinants of dissemination and host specificity of metallo-β-lactamases. Nat. Commun. 10, 3617 (2019).
-
Fernández, A. et al. Expression of OXA-type and SFO-1 β-lactamases induces changes in peptidoglycan composition and affects bacterial fitness. Antimicrob. Agents Chemother. 56, 1877–1884 (2012).
-
Tu, M. M. et al. Exploiting the fitness cost of metallo-β-lactamase expression can overcome antibiotic resistance in bacterial pathogens. Nat. Microbiol. 10, 53–65 (2025).
-
Herencias, C. et al. β-lactamase expression induces collateral sensitivity in Escherichia coli. Nat. Commun. 15, 4731 (2024).
-
Furniss, R. C. D. et al. Breaking antimicrobial resistance by disrupting extracytoplasmic protein folding. eLife 11, e57974 (2022).
-
Dunn, S., Carrilero, L., Brockhurst, M. & McNally, A. Limited and strain-specific transcriptional and growth responses to acquisition of a multidrug resistance plasmid in genetically diverse Escherichia coli lineages. mSystems, https://doi.org/10.1128/msystems.00083-21 (2021).
-
Buckner, M. M. C. et al. Clinically relevant plasmid-host interactions indicate that transcriptional and not genomic modifications ameliorate fitness costs of klebsiella pneumoniae carbapenemase-carrying plasmids. mBio. https://doi.org/10.1128/mbio.02303-17 (2018).
-
DelaFuente, J., Rodriguez-Beltran, J. & San Millan, A. Methods to Study Fitness and Compensatory Adaptation in Plasmid-Carrying Bacteria. Methods Mol. Biol. Clifton NJ 2075, 371–382 (2020).
-
Calvo-Villamañán, A. et al. On-target activity predictions enable improved CRISPR–dCas9 screens in bacteria. Nucleic Acids Res 48, e64–e64 (2020).
-
Rousset, F. et al. The impact of genetic diversity on gene essentiality within the Escherichia coli species. Nat. Microbiol. https://doi.org/10.1038/s41564-020-00839-y (2021).
-
Gibson, D. G. et al. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 6, 343–345 (2009).
-
Le Roux, F., Binesse, J., Saulnier, D. & Mazel, D. Construction of a vibrio splendidus mutant lacking the metalloprotease gene vsm by use of a novel counterselectable suicide vector. Appl. Environ. Microbiol. 73, 777–784 (2007).
-
Strand, T. A., Lale, R., Degnes, K. F., Lando, M. & Valla, S. A New and Improved Host-Independent Plasmid System for RK2-Based Conjugal Transfer. PLOS ONE 9, e90372 (2014).
-
Andrews S. FastQC: a quality control tool for high throughput sequence data. http://www.bioinformatics.babraham.ac.uk/projects/fastqc (2010).
-
Ewels, P., Magnusson, M., Lundin, S. & Käller, M. MultiQC: summarize analysis results for multiple tools and samples in a single report. Bioinformatics 32, 3047–3048 (2016).
-
Katz, L. S. et al. Mashtree: a rapid comparison of whole genome sequence files. J. Open Source Softw. 4, 1762 (2019).
-
San Millan, A., Heilbron, K. & MacLean, R. C. Positive epistasis between co-infecting plasmids promotes plasmid survival in bacterial populations. ISME J. 8, 601–612 (2014).
-
Datsenko, K. A. & Wanner, B. L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. 97, 6640–6645 (2000).
-
Sastre-Dominguez, J. Dissecting pOXA-48 fitness effects in clinical Enterobacterales using plasmid-wide CRISPRi screens. https://doi.org/10.5281/zenodo.16609943.
Acknowledgements
We thank the R-GNOSIS group, the volunteers and the medical staff from the Hospital Universitario Ramón y Cajal (Madrid, Spain) involved in the sample isolation process. We thank David Bikard and the Institut Pasteur for plasmid pFR56. We thank the Unidad de Genómica del Parque Científico de Madrid for helping with the Illumina sequencing. We thank Sarah Bigot for the kind gift of plasmid pOXA-48A_nuc::sfGFP. This work was supported by the European Research Council (ERC) under the European Union’s Horizon Europe research and innovation programme (ERC-2022-CoG Project 101086992 – PLAS-FIGHTER) and by MCIN/AEI/10.13039/501100011033 and the European Union NextGenerationEU/PRTR (Project PCI2021- 122062- 2 A). A.C.V. was funded by an EMBO postdoctoral fellowship (ALTF 322-2022). We acknowledge financial support from the Spanish State Research Agency, AEI/10.13039/501100011033, through the “Severo Ochoa” Programme for Centres of Excellence in R&D [SEV-2013-0347, SEV-2017-0712, CEX2023-001386-S].
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Enas Newire, and the other, anonymous, reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Calvo-Villamañán, A., Sastre-Dominguez, J., Barrera-Martín, Á. et al. Dissecting pOXA-48 fitness effects in clinical Enterobacterales using plasmid-wide CRISPRi screens. Nat Commun 16, 7700 (2025). https://doi.org/10.1038/s41467-025-63082-1
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41467-025-63082-1