
Introduction
Biotherapeutics constitute a large and growing segment of pharmaceutical products. Unlike non-proteinaceous chemical molecules, biotherapeutic proteins are subject to adaptive immune responses. Understanding the immunogenicity potential of biotherapeutic proteins is of major importance as these medicines can induce the body to produce anti-drug antibodies (ADAs). ADAs can have broad and diverse effects such as negating the clinical benefit of the biotherapeutic protein by reducing the efficacy and/or pharmacokinetic properties of the biotherapeutic protein, causing allergic reactions, and cross-reaction with endogenous proteins. These effects can be serious and, in some cases, lethal1,2,3,4. Monitoring the potential immune response of biotherapeutic proteins is an important regulatory requirement during drug development (Immunogenicity Assessment for Therapeutic Protein Products (fda.gov)). The identification and removal of immunogenic components from biotherapeutics is a critical objective for developing safe and effective medicines.
A central tenant of the humoral adaptive immune response is the presentation of peptides by professional antigen presenting cells (APCs) for immunosurveillance by CD4 T cells which orchestrate the response to a variety of conditions including infections, cancer, autoimmunity, allergy, and immunogenicity of protein therapeutics. The expression of human leukocyte antigen class II (HLA class II) molecules, which display fragments of self and foreign proteins for interaction with T cell receptors (TCRs), is restricted to APCs. HLA class II molecules are highly polymorphic and are encoded by multiple loci. HLA-DR is the predominant HLA class II isotype and is a heterodimer consisting of invariant alpha (DRA) and highly polymorphic beta (DRB) chains. Additionally, multiple HLA-DRB paralogs (DRB1/3/4/5) exist. An individual will have one (homozygous) or two (heterozygous) HLA-DRB1 alleles but the presence of additional DRB3, DRB4, or DRB5 paralogs is variable and dependent on the haplotype of the individual. Historically, HLA-DRB1 alleles have been the primary focus for the evaluation of immunity, disease, and immunogenicity given its ubiquitous presence in all APCs and high level of expression relative to the other HLA class II loci5,6.
Immunopeptidomics first described by Hunt et al.7,8 demonstrated that mass spectrometry is a powerful technology for the direct identification of HLA-bound peptides. MHC-associated peptide proteomics (MAPPs) is a specific extension that determines HLA class II peptides after the intentional pulsing of monocyte-derived dendritic cells (MDDCs) with exogenous antigen or protein of interest9. This technique has become an increasingly important tool to assess the immunogenicity potential of viral infection10, gene therapy delivery vectors11,12, and protein therapeutics13,14,15. These mass spectrometry-based approaches enable the precise elucidation of the protein sequences presented to TCRs by HLA class II molecules under endogenous peptide-loading conditions and greatly enhance our ability to both understand and modulate humoral responses. However, as most individuals are multi-allelic, uncertainty in the HLA-DRB1 peptide restriction persists. Each HLA-DRB1 allele can have distinct peptide binding specificities but most individuals are heterozygous which confounds the ability to discern the distinct, and sometime overlapping, repertoires of displayed peptides. This ambiguity also precludes accurate assessment of the proportion of the population at risk for developing immunogenicity to a given protein therapeutic or vaccine candidate. Finally, the reliance upon fresh peripheral blood mononuclear cells (PBMCs), which we and others have shown to be optimal for MAPPs sensitivity16, brings a degree of randomness and difficulty to sample alleles from different geographies that reflect the span of HLA-DRB1 alleles present in diverse patient populations.
B cells are another type of professional APC that express HLA class II molecules and interact with T cells during the cascade of events that occur during an adaptive immune response. Implementation of B cells into an immunopeptidomic workstream could take advantage of the ability to utilize immortalized cell lines17 which is currently not tractable with dendritic cells. While macrophages and dendritic cells have the capacity for universal antigen internalization, B cells have clonally distributed antigen-specific surface immunoglobulin receptors (BCRs) which restrict the diversity of antigen uptake. Lanzavecchia18 demonstrated that BCRs are required for antigen internalization under limiting concentrations to enable processing and presentation to T cells in an HLA-restricted manner indistinguishable from that of other APCs. Unfortunately, the antigen-specific nature of the BCR and difficulties in obtaining appropriate monoallelic HLA-DRB1 configurations has prevented the utilization of B cells as a general technology platform for routine immunogenicity assessments.
We report the development of a platform that addresses the shortcomings of using B cells for immunopeptidomics. We constructed a panel of HLA-DRB1 monoallelic immortalized B cell lines having a universal BCR capable of binding a wide range of clinically relevant biotherapeutic proteins. These cell lines correspond to >70% allele frequency usage from two distinct geographic regions. We show robustness and reproducibility of our system and compare results to those obtained using MDDCs. We extend our knowledge of HLA-DRB1 allele peptide binding motif preferences and show divergence between members of HLA-DRB1 supertypes. We characterize the immunogenic potential of clinical antibodies and show how understanding the precise allelic restriction of presented peptides provides higher resolution of patient populations at risk for developing ADA responses which can lead to decreased efficacy and adverse patient experiences. Finally, we present evidence that invariant cysteine residues from multiple antibodies can be modified and comprise distinct components of HLA-DRB1 binding.
Results
Establishing a universal BCR HLA-DRB1 monoallelic APC platform
To address shortcomings in the ability to interrogate HLA-DRB1 presentation of biotherapeutics and to better understand the specific allelic restriction from a broad range of alleles, we aimed to construct a monoallelic HLA-DRB1 APC platform capable of internalizing a wide range of biotherapeutic protein modalities (Fig. 1A). Raji cells are a human B cell lymphoblastoid cell line resulting from Epstein-Barr viral integration that has been in continuous culture since the early 1960’s19. The non-adherent growth characteristics, fast doubling times, and facile transfection efficiency made this an ideal B cell line to build the universal BCR monoallelic HLA-DRB1 platform. We used this cell line as starting point for our genetic manipulations.
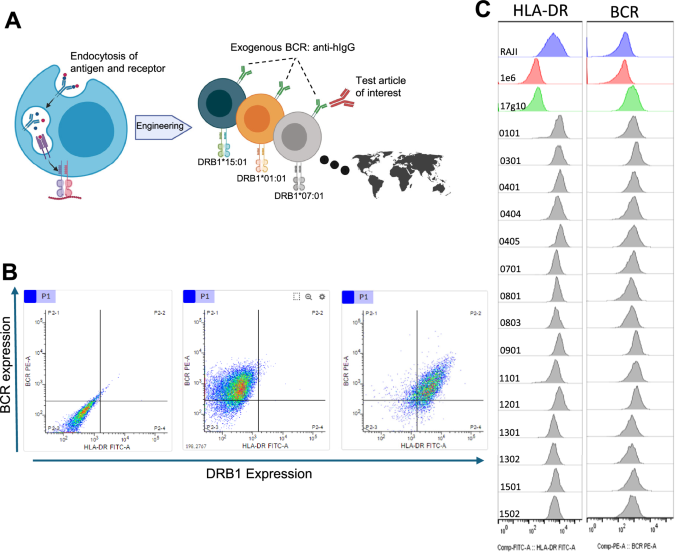
The endogenous HLA-DRB alleles in the Raji cell line are HLA-DRB1*03:01 and 10:01 and HLA-DRB3*02:02. We designed a series of guide RNAs for CRISPR-mediated disruption of the functional reading frames of the alleles. The absence of cell surface expression was confirmed by flow cytometry (Fig. 1B left panel and Fig. 1C 2nd row) and western analysis (Supplementary Fig. 1). Sequencing of genomic DNA reveals 7 nucleotide deletion in DRB1*03:01 allele, 10 nucleotide deletion in the exon 1 in DRB1*10:01, and 27 nucleotide deletion in DRB3:02:02 allele (Supplementary Fig. 1). Clone 1E6 was selected as the HLA-DRB knockout cell line for subsequent work.

A Generalized schematic depicting the platform. Raji B cells were engineered to (1) add a universal BCR capable of binding various human IgG subclasses and (2) reconstitute HLA-DRB1 alleles of interest following removing of endogenous alleles. B Flow cytometric profiling of surface expression of exogenous universal BCR (Y-axis) and exogenous HLA-DRB1 allele (X-axis) from representative cell lines during platform development. The base endogenous HLA-DRB1 knockout cell line (1E6) is shown in the left plot. The exogenous universal BCR cell line (17g10) is shown in the middle plot. An exogenous monoallelic HLA-DRB1 cell line (HLA-DRB1*01:01) is shown in the right plot. C Histograms of all cell lines. HLA-DRB1 surface expression is denoted in the left column and exogenous universal BCR expression is denoted in the right column. Parental Raji cells (top row, blue), endogenous DRB1 knockout line 1E6 (2nd row, red), exogenous BCR cell line 17g10 (3rd row, green) and all 15 monoallelic HLA-DRB1 allele cell lines (remaining rows with the 4-digit HLA-DRB1 allele denoted in the inset, gray). Universal BCR was detected using biotinylated hIgG and streptavidin-PE. HLA-DR expression was detected using anti-HLA-DR antibody (L243) and anti-mouse FITC labeled secondary reagent. Created in BioRender. Siegel, R. (2025) https://BioRender.com/9ioqt73.
Once the expression of functional endogenous HLA-DRB alleles had been eliminated, we next embarked to create a stable cell line expressing an exogenous BCR capable of binding all human IgG subclasses. A mouse hybridoma clone was identified that bound human IgG, but not human IgM, human IgE or bovine IgG. The murine variable regions were cloned into an expression vector encoding human IgD and IgK constant region sequences to express a chimeric exogenous BCR after stable cell integration using blasticidin selection upon transfection. The IgD isotype was chosen as the scaffold for our universal chimeric BCR to maximize the probability of surface expression and appropriate signal transduction upon antigen binding without depending on the ancillary Igα and Igβ molecules20 required for IgM-mediated BCR signal transduction. Clone 1E6-17g10 was selected for further manipulation based on robust surface expression of the exogenous BCR capable of binding human IgG (Fig. 1B middle panel).
We next set forth to re-introduce functional HLA-DR molecules by stable integration of various HLA-DRB1 alleles into the 17g10 universal BCR cell line. The coding sequence of each HLA-DRB1 allele of interest was cloned into an expression vector for stable integration using puromycin selection upon transfection. The alleles were chosen to reflect the highest frequencies across different ethnic groups from the United States and Japan populations (Table 1) and corresponding breadth of peptide binding specificities. We used allele frequencies derived from the Allele Frequency Net Database21 for our calculations. A total of 15 different HLA-DRB1 cell lines were constructed. This constellation of HLA-DRB1 alleles represents 9 of the top 10 alleles for U.S. population of European descent, 7 of the top 10 alleles for U.S. population of African American descent, 8 of the top 10 alleles for U.S. population of Hispanic descent, 9 of the top 10 alleles for U.S. population of Chinese descent, and 9 of the top 10 alleles for the Japan population which was not divided into any ethnic groupings. The 15 HLA-DRB1 alleles incorporate 71% and 75% allele frequency usage for the U.S. and Japan populations, respectively. These allele frequencies translate to covering ~92% and ~94% of the individuals in the U.S. and Japan populations, respectively, owing to heterozygosity. We sequence confirmed the HLA-DRB1 allele for each cell line generated (and developed a panel of allele-specific PCR primers that enable facile confirmation of cell line identity for quality control during continual cell growth (Supplementary Fig. 2 and Supplementary Table 1).
One uncertainty that remained was as to whether the exogenous HLA-DRB1 chain randomly inserted into the 17g10 genome would pair with the remaining endogenous HLA-DRA chain to form a functional molecule that was translocated to the cell surface. Flow cytometry was used to determine if the intact heterodimer is expressed after HLA-DRB1 transfection. Dual staining with human IgG to denote the exogenous universal BCR and HLA-DR antibody L243 show no signal with either reagent from the 1E6 cells (Fig. 1B, left panel), exogenous universal BCR with 17g10 cells (Fig. 1B, center panel) and dual staining of both the exogenous universal BCR and HLA-DR with a representative monoallelic cell line (Fig. 1B, right panel). The surface expression profile for all 15 HLA-DRB1 cell lines shows comparable levels to that observed from the parental Raji cell line with the endogenous HLA-DRB1*03:01 and 10:01 molecules (Fig. 1C). We further investigated and determined that the quantitative level of HLA-DRB1 expression for each cell line (range of 50,000 to 250,000 molecules/cell) was equivalent to, or higher than, that of the parental Raji cell line that contained both endogenous HLA-DRB1 alleles (Supplementary Fig. 3). We also confirmed that a reconstituted HLA-DRB1 molecule had to be present to identify immunopeptidomic peptides (Supplementary Fig. 4).
Characterization of the universal BCR monoallelic HLA-DRB1 technology platform
Upon generating the panel of monoallelic HLA-DRB1 cell lines with the universal BCR, we wanted to confirm the ability of the platform to bind, internalize, process, and load peptides for display on HLA-DRB1 from a wide variety of exogenous biotherapeutics. We dosed each of the 15 monoallelic cell lines with a panel of anti-PCSK9 antibodies that represented different Ig HC and LC subclasses. Alirocumab is IgG1/kappa, bococizumab is IgG2/kappa, evolocumab is IgG2/lambda, and frovocimab is IgG4-S228P, F234A, L235A/kappa subclasses, respectively. Peptides identified by MAPPs derived from the constant regions of the antibodies are aligned and displayed with heat maps (Fig. 2). Most, but not all, alleles display peptides from a least one area of the constant regions and many alleles display peptides from multiple regions. Constant region peptides were observed from 12 of 15 alleles for IgG1, IgG2, and IgG4. Constant region peptides were observed for 13 of 15 alleles for IgK and 10 of 15 alleles for IgL. Interestingly, a few alleles did not present peptides from most of the subclasses examined. For instance, HLA-DRB1*15:01 did not present peptides from IgG1, IgG2, IgG4, and IgK subclasses. We confirmed that the HLA-DRB1*15:01 cell line was functional as 1000’s of peptides from endogenous proteins, media components and variable region peptides from multiple therapeutic antibodies were identified during cell line characterization. Taken together, these results demonstrate that the universal BCR is capable of binding and internalizing several different antibody subclasses commonly used for therapeutic applications.
We also analyzed the unique HLA-DRB1 peptides derived from endogenous and serum components obtained from our studies analyzing the multiple antibody subclasses. In aggregate we identified 33,584 unique peptides from this dataset. The total number of unique peptides for each cell line are shown (Supplementary Fig. 5). Peptide numbers ranged from a maximum of 5765 for HLA-DRB1*09:01 to a minimum of 1412 peptides for DRB1*13:01. Interesting the peptide numbers were not correlated with HLA-DRB1 surface expression levels. Leveraging the monoallelic nature of our system, we were able to unequivocally assign peptide restriction for binding core predictions using MHCMotifDecon 1.0 (https://services.healthtech.dtu.dk/services/MHCMotifDecon-1.0/). We observed that the predicted HLA-DRB1 binding cores from our monoallelic B cell platform matched those obtained from larger muti-allelic MDDC immunopeptidome data sets (Supplementary Fig. 6). Our monoallelic system also enabled subtle binding differences between alleles within HLA supertypes to be unambiguously defined (Supplementary Fig. 7). For instance, HLA-DRB1*04:01 will accept F and Y residues at P1 and negatively charged resides at P4 and P6. However, HLA-DRB1*04:04 does not accept F and Y residues at P1 or negatively charged resides at P4. Finally, HLA-DRB1*04:05 accepts F and Y residues at P1 but prefers negatively charged residues at P9.
We next wanted to examine the effect of antigen titration on the ability to identify exogenous antigen-derived HLA-DRB1 peptides. Bococizumab is an antibody that resulted in high incidence of ADA that impacted efficacy22. We dosed bococizumab at concentrations ranging from 1 to 100 µg/ml on 4 different HLA-DRB1 cell lines that we had previously determined could present variable region peptides from this molecule. Three distinct clusters with varying numbers of peptides contained in each cluster were observed from the variable heavy (VH) region (Fig. 3A). Remarkably, the HLA-DRB1 display of all the clusters remained largely unchanged over concentrations that differed by 2 orders of magnitude. A closer examination of the H3 cluster (residues 87–105) from HLA-DRB1*07:01 cell line revealed that almost all the 15 peptide sequences identified at the 100 µg/ml dose level were also observed at the 1 µg/ml dose level (Fig. 3B).
We were also interested in examining the reproducibility of our results over the course of time. Five distinct clusters from golimumab VH were observed from four different HLA-DRB1 cell lines and the presence and density of display were unchanged over the course of >20 passages during continuous cell culture (Fig. 3C). The total number of all golimumab peptides observed from both the VH and VL regions observed from 7 different HLA-DRB1 cell lines are plotted as a function of passage number (Fig. 3D) and inter-experiment variability examined (Supplementary Table 2). In totality, this data demonstrates the robust reproducibility and sensitivity of our system. We used 10 or 100 µg/ml dosing concentrations for all following experiments.
Finally, we compared MAPPs results obtained from the universal BCR monoallelic HLA-DRB1 platform with results from traditional monocyte-derived dendritic cells (MDDCs). ATR-107 is a fully human anti-IL21 receptor antibody that generated a high degree of ADA with adverse events associated with hypersensitivity reactions during clinical investigation23. ATR-107 has been used as a positive control in preclinical immunogenicity assessment assays24,25.
We dosed MDDCs from 10 normal healthy donors (Supplementary Table 3) and the 15 HLA-DRB1 cell lines with ATR-107. Either a pan HLA class II antibody (Tu39) capable of binding a wide subset of HLA-DR1/3/5, -DQ, and -DP molecules or HLA-DR antibody (L243) was used for immunoprecipitation prior to MS analysis. Heatmaps of the results from both VH and VL regions of ATR-107 are shown (Fig. 4 left and right panels, respectively). Of foremost interest, the HLA-DRB1 cell lines recapitulated all the HLA class II clusters observed from MDDCs. The prominent VH CDR3 cluster (residues 90–105) observed from a majority of the MDDC donors is restricted to HLA-DRB1*01:01, 07:01, 09:01, 11:01 and 15:02 alleles. The VL CDR2 cluster (residues 37–59) is restricted to HLA-DRB1*13:01 and 13:02 alleles. Finally, the VL CDR3 cluster (residues 79–99) is restricted to HLA-DRB1*01:01, 04:01, 07:01, 08:01, 08:03, 09:01, 11:01, and 15:02 alleles. These results also indicate, at least for ATR-107, that all the clusters observed from MDDCs are likely restricted to HLA-DRB1 molecules as we did not observe any meaningful differentiation between results obtained with pan- or DR-specific antibodies. Most importantly, we conclude that the universal BCR monoallelic HLA-DRB1 platform is comparable to results obtained with MDDCs.

A MAPPs display heatmaps representing IgG1 (top), IgG2 (middle), and IgG4 (bottom) human heavy chain subclasses. The various constant region domains are depicted above the heatmaps for orientation. B MAPPs display heatmaps representing kappa (top) and lambda (bottom) human light chain isotypes. Each HLA-DRB1 allele is represented as a row in the heatmap. The x-axis denotes the position within the constant region sequence (EU numbering). Heatmap densities are normalized to the region of highest peptide display for each isotype, respectively.

A Four cell lines were dosed with 1, 10, or 100 µg/ml anti-PCSK9 bococizumab. Peptides identified from the VH are summarized and displayed as a heatmaps. Each row represents an individual condition (HLA-DRB1 and dose level). The x-axis denotes the position within the linear sequence. Darker shaded regions within the heatmap correspond to greater peptide coverage. The density scale is capped at 20 peptides. B Bococizumab H3 cluster peptides (residues 87–105) identified from DRB1*07:01 are listed on the right. Three columns represent 1, 10, and 100 µg/ml dose levels. Boxes shaded in gray depict peptides that were identified from the dosing concentration denoted below the chart. For example, the first peptide (RSEDTAVYYCARERPLY) was identified from the 100 µg/ml, but not 10 or 1 µg/ml dose levels. C The heatmap shows golimumab VH peptides identified from four different cell lines at passage 7, 13 and 30. Each row represents an individual condition (HLA-DRB1 and passage number). The x-axis denotes the position within the linear sequence. The density scale is capped at 20 peptides. D Golimumab was dosed on cell lines at various times across 30 passages. The graph depicts the sum of golimumab variable region peptides (VH + VL) identified from each sample. The peptide count is denoted on the Y-axis, the passage number is denoted on the x-axis, and colored lines indicate different HLA-DRB1 alleles.

MDDCs and B cell lines were dosed with anti-IL21R antibody ATR-107. Peptides identified from the variable regions were summarized for each sample and displayed as heatmaps. Each row of a heatmap represents an independent donor or monoallelic HLA-DRB1 cell line. The heatmaps on the left show peptides aligned to the VH using linear sequence numbering. The heatmaps on the right show peptides aligned to the VL using linear sequence numbering. The top plots correspond to MDDCs immunoprecipitated with a pan-HLA class II antibody (clone Tu39). The middle plots correspond to MDDCs and the bottom plots correspond to monoallelic B cell lines immunoprecipitated with an HLA-DR specific antibody (clone L243). The scale is capped at 20 peptides for each heatmap.
Population immunogenicity risk assessment of adalimumab with the monoallelic HLA-DRB1 universal BCR platform
Adalimumab is one of the most prescribed biologics in the world and has been associated with various degrees of immunogenicity26. We sought to determine if our HLA-DRB1 monoallelic system could indicate which proportion of both U.S. and Japanese populations may be at risk for developing ADA. Adalimumab was dosed at a concentration of 100 µg/ml with each of the 15 HLA-DRB1 monoallelic B cell lines. Adalimumab-derived MAPPs peptides were identified, aligned to the parent sequence, and displayed as a heatmap for relative peptide abundance visualization (Fig. 5 and Supplementary Data S1). A total of 221 unique adalimumab VH and VL derived peptides ranging in length from 10 to 23 residues with a median length of 15 were identified from 14 of the 15 monoallelic HLA-DRB1 B cell lines. No adalimumab variable region peptides were observed from HLA-DRB1*03:01 cell line. One can readily observe that certain portions of adalimumab variable regions are associated with particular HLA-DRB1 allele(s) with different density of display and that not all HLA-DRB1 alleles present peptides from the same regions of adalimumab. A low of 6 variable region peptides were observed from HLA-DRB1*12:01 and a high of 100 variable region peptides were observed from HLA-DRB1*01:01 (Supplementary Data S1). Interestingly, peptides from the VH domain were either the only presented peptides or were the variable domain with the higher number of presented peptides from every allele except DRB1*15:01 which only presented peptides from the VL domain.
HLA-DRB1 presentation of peptides that contain one or more non-germline residues are considered at risk for eliciting a T cell response that aid the generation of anti-drug antibody responses. Two overlapping clusters from CDR2 and two overlapping clusters from CDR3 of the VH region were identified that contain one or more non-germline residues (Fig. 5, top panel). One cluster from CDR1, one cluster from CDR2 and two overlapping clusters from CDR3 of the VL region (Fig. 5 bottom panel) were identified that contained one or more non-germline residues. Peptide clusters from other regions of adalimumab variable regions were also observed but did not contain any non-germline residues to indicate risk and are not annotated. In aggregate, 8 regions of concern were identified from 14 of the 15 monoallelic cell lines. The ability of a particular sequence to activate CD4 T cells depends on many factors including the peptide binding register and whether non-germline residue(s) are exposed for TCR interaction. While not every region presented for T cell surveillance would be expected to result in activation, it was interesting to note that most of the adalimumab regions that we identified from our panel of monoallelic HLA-DRB1 cell lines have been associated as CD4 T cell epitopes. Table 2 lists those clusters that have been shown to elicit a T cell response from either healthy drug naïve donors or patients who developed ADA to adalimumab27. Moreover, the percent of the population at risk for potential ADA can be calculated based on allele frequencies for both U.S. and Japan populations. The H3-1 cluster (VH resides 87-106) has the widest range of prevalence with nearly 70% of U.S. and 95% of Japanese populations at risk for developing anti-adalimumab antibodies mediated by this single region of the molecule. It is also interesting to note the magnitude of difference in exposure for the H2-2 cluster (residues 53–71) between U.S. and Japan populations at 38% and 66%, respectively. This data highlights how the same segment of a biotherapeutic molecule can have differential immunogenicity impact between populations with diverse HLA-DRB1 allele utilization.

Heatmaps of HLA-DRB1 peptides derived from adalimumab variable heavy (top panel) and variable light (bottom panel) domains from each of the 15 monoallelic B cell lines. Each row represents results from one HLA-DRB1 allele cell line. The heatmap density legend is displayed to the right. The top end of the heat map was fixed at 20 peptides. Linear residue numbering is indicated below each graph. VH variable heavy, VL variable light, CDR complementarity determining region; H1 = VH CDR1; H2 = VH CDR2; H3 = VH CDR3; L1 = VL CDR1; L2 = VL CDR2; L3 = VL CDR3. General locations of clusters are annotated at top of each heatmap.
Modified cysteine residues are present in HLA-DRB1 binding registers
Our current MAPPs protocol does not implement a reduction/alkylation step prior to LC-MS so that we may observe natural modifications in any cysteine-containing HLA-DRB1 peptide. The list of the cysteine modifications used in our database search strategy is indicated in the methods. Analysis of cysteine-containing peptides from our anti-PCSK9 antibody immunopeptidomic dataset indicate 30% of identified peptides contain a modified cysteine residue. Universal intra-domain disulfide bonds are contained in both VH and VL domains. A common HLA-DRB1 cluster that spans VH framework 3 (FR3) into the CDR3 containing an invariant cysteine residue was observed from many antibodies across several alleles. Interestingly, we observed several H3 CDR cysteine-containing peptides that contained various modifications. Even more surprising was an apparent synchronicity of the modification pattern between certain alleles for identical peptide sequences.
Representative cysteine-containing peptides from both adalimumab and ATR-107 are shown in Fig. 6. The adalimumab FR3-H3 cluster (residues 88-109) observed from HLA-DRB1*01:01 allele contains 25 unique cysteine-containing peptide sequences. None of the peptides associated with this allele contain modified cysteines (Fig. 6A). In stark contrast, 13 out of 14 unique adalimumab cysteine-containing peptides obtained from HLA-DRB1*13:01 contained a cysteine tri-oxidized to cysteic acid or di-oxidized to sulfinic acid. A similar allele-specific modification pattern is also observed with the ATR-107 FR3-H3 cluster (Fig. 6B). Modified cysteine residues were absent from the 15 unique cluster peptides (residues 88–106) identified from the HLA-DRB1*07:01 allele. Again, in stark contrast, all 8 of the unique ATR-107 FR3-H3 peptides (residues 88–105) obtained from HLA-DRB1*11:01 allele only contained tri-oxidized modified cysteine residues. We also observed other cysteine modifications (mono-oxidation, di-oxidation, and cysteinylation) from other alleles from this same region of both adalimumab and ATR-107 molecules. Mass spectrometry data supporting the modified cysteine identifications are shown in Supplementary Fig. 8.

Modification state of cysteine-containing VH CDR3 peptides identified from A adalimumab and B ATR-107. Each unique peptide is represented by a dot. Cysteine modifications (Y-axis) and HLA-DRB1 allele of the cell lines (X-axis) are denoted on each graph. C Predicted HLA-binding registers for representative cysteine-containing VH CDR3 peptides that are modified in one allele but not another. The antibody and the HLA-DRB1 alleles are denoted on the left side of table. Cluster depth represents the number of unique peptide sequences containing the predicted binding core from each allele. Peptide sequences of a representative peptide from each antibody are listed and the predicted HLA-binding core (underlined) for each HLA-DRB1 allele was determined using NetMHCIIpan 4.3 (NetMHCIIpan 4.3 – DTU Health Tech – Bioinformatic Services). The reliability of the binding core (core_rel), expressed as the fraction of networks in the ensemble selecting the optimal core, is reported. The modification state (SH or tri-oxidation), predicted core position (P3 or P4), and T cell receptor or HLA-DRB1 exposure status (TCR or HLA-DR) as a result of the predicted binding core of the cysteine residues are denoted.
We used NetMHCIIpan-4.328 to predict the binding core from adalimumab (AEDTAVYYCAKVSYL) and ATR-107 (AADTAVYYCARGGGIS) FW3-H3 peptides (Fig. 6C). These peptides were chosen as representative sequences that were identified from alleles that contained either unmodified or modified cysteines. The predicted 9-mer binding register from both peptides for each allele is underlined and is of high confidence. Binding core residues P1, P4, P6, and P9 are buried and critical binding contacts with HLA-DRB1 and residues P2, P3, P5, P7, and P8 are solvent exposed and could interact with TCR29. The unmodified cysteine from the adalimumab peptide is predicted to reside at P3 and be solvent exposed for TCR interaction when bound by HLA-DRB1*0101; whereas the modified cysteine is predicted to reside at P4 and be buried as part of interaction with HLA-DRB1*13:01. The unmodified cysteine residue for the ATR-107 peptide is also predicted to reside at P3 and be TCR-facing when bound by HLA-DRB1*07:01. The ATR-107 modified cysteine residue is predicted to reside at P4 and interact with the HLA-DRB1*11:01 allele. We find it curious that a buried cysteine residue would carry a modification, as we presumed its location within the binding pocket would be somewhat protective. However, it remains to be determined whether the binding register predictions are incorrect or if cysteine-modifications are an inherent property for binding to certain HLA-DRB1 alleles.
Discussion
We have developed a diverse, robust, and reproducible monoallelic HLA-DRB1 system in professional APCs currently capable of examining the immunogenic potential of any human IgG. Our panel of cell lines is currently composed of 15 HLA-DRB1 alleles that are present in >90% of individuals from two distinct geographic regions. The inclusion of an exogenous universal BCR allows the platform to interrogate a wide range of biotherapeutic protein modalities in a professional APC. Utilization of these cells preserves the natural expression of the ancillary HLA-DM and HLA-DO proteins required for accurate peptide loading which we believe best reflect in vivo conditions. The platform is highly scalable, and the throughput is amendable to analysis of multiple molecules on a weekly basis. We posit that this system can become a cornerstone of a preclinical immunogenicity risk assessment strategy and an invaluable resource to expand our understanding of HLA class II binding specificities for vaccine development.
The robustness and reproductivity of our immunopeptidomic results are due to the utilization of stable cell lines with our B cell platform which enables consistent expression levels of both the universal BCR and HLA-DRB1 allele of interest. We identified a highly conserved set of HLA-DRB1 peptides from multiple alleles across two orders of magnitude of antigen titration (Fig. 3A, B). We also show an ability to consistently isolate the same peptides after more than 20 passages from multiple HLA-DRB1 alleles (Fig. 3C, D). This consistency continues with all cell lines that have been in continuous culture for more than a year. Of critical importance, we show that our monoallelic B cell system recapitulates results obtained with traditional monocyte derived dendritic cells (Fig. 4). We see essential overlap of MDDC results using either HLA-DR specific or pan-HLA class II immunoprecipitations. We do observe the emergence of a few novel clusters from our monoallelic HLA-DRB1 B cell system. It is intriguing to speculate if these differences stem from the presence of HLA-DO which is absent in non-B cell APCs.
Monoallelic HLA class II immunopeptidomic approaches have been previously described. Most notably Abelin and colleagues have developed an elegant allele specific immunoprecipitation method that leverages transient transfection of tagged HLA class II molecules into non-professional APCs30,31. While capable of examining a range of non-HLA-DRB1 alleles, several limitations exist. First and foremost, the extracellular molecules that can be analyzed with this approach is limited to only those that have natural receptors or achieve cell entry via infection. While this approach is an excellent means to develop allele-specific training sets for improved prediction algorithms based on largely intracellular and limited extracellular proteins, it is incompatible for investigating the HLA-DRB1 repertoire of a multitude of biotherapeutic proteins. Second, utilization of non-APCs require co-transfections with HLA-DM to facilitate appropriate peptide exchange for high-affinity ligands as immunopeptidome differences have been noted across several HLA-DR alleles when HLA-DM is either omitted entirely or the ratio of expression relative to HLA-DR is changed30. Finally, the need for transient transfection of both HLA-DM and each HLA class II allele of interest for each experimental run reduces efficiency in a production environment and limits the scale that can be obtained. We also speculate that the lack of stable cell lines could impact reproducibility of results.
The HLA-DRB1 peptides isolated from our system can provide unprecedented resolution on the various populations at risk for developing anti-drug antibody responses. Evaluation of the peptide presentation pattern from adalimumab identified 8 clusters (nested set of peptides with a common core) containing one or more non-germline residues spread across 14 of the 15 alleles contained in our panel (Fig. 5). Additional clusters containing non-germline residues are ignored as they are unlikely to represent an immunogenicity risk. A recent publication determined adalimumab T cell epitopes from both drug naïve healthy donors and ADA+ patients27. Multiple CD4 T cell lines encompassing 5 different regions were generated after multiple rounds of stimulations by autologous DCs dosed with adalimumab. The T cell epitopes were determined using overlapping peptides. We were encouraged by the fact that our HLA-DRB1 B cell system was able to identify all 5 of these known T cell epitopes.
We leveraged the unambiguous monoallelic restriction of the observed peptides gleaned from our system to calculate a percentage of population at risk for each adalimumab cluster (Table 2). For example, the dominant T cell epitope represented by cluster H3-1 (residues 87–106) has the potential to facilitate immunogenicity for nearly 70% and 95% of the U.S. and Japan populations, respectively. We can also determine how many clusters are observed with any given allele. HLA-DRB1*07:01 is the most prominent allele (nearly 22%) in the U.S. population after adjustment for variable expression frequencies among the major ethnic group constituents. This allele presented peptides from 3 of the 5 verified T cell epitopes. HLA-DRB1*09:01 is the most prominent (nearly 27%) allele in the Japanese population. This allele presented peptides from 4 of the 5 verified T cell epitopes. The ability to use designed allele sets allows examination of risk for potential patients that are underrepresented in localities used for fresh PBMC collections when using a MDDC-derived MAPPs approach. One could also imagine scenarios where cells from different HLA-DRB1 allele cell lines are simply pooled to create a single batch to ease experimental throughput. Pooling eliminates the ability for experimental determination of the allelic restriction of HLA-DRB1 peptides but provides the ability to manipulate the composition of the pool to best reflect the allelic diversity of any population of interest for a global assessment of immunogenicity risk.
Our antibody immunopeptidomic datasets, composed of molecules containing multiple cysteine residues coupled with our unique approach of not reducing and alkylating cysteines prior to proteomic analysis, provided an opportunity to investigate the redox state of various HLA-DRB1 cysteine containing peptides. We observed peptides containing cysteine residues from most therapeutic antibodies analyzed. Fascinatingly, a subset of cysteine containing peptides in the immunopeptidomic dataset from our monoallelic B cell system were found to be cysteinylated or oxidized (primarily tri-oxidation). The fact that most of the cysteine-containing peptides from dosed antibody molecules, endogenous proteins or serum components identified in our study are not modified strongly implies that these findings are not an artifact of the MAPPs protocol. Even more dramatic, we observed several instances in which the same cysteine containing peptide sequence would be quantitatively modified when isolated from one HLA-DRB1 allele but completely unmodified when isolated from another allele (Fig. 6). This dichotomous pattern was not observed for any of the other post-translation modifications (e.g. deamidation and oxidation) included in our search strategy defined in “Methods”.
Understanding the biological mechanism that leads to these cysteine modifications in peptides derived from therapeutic antibodies is of interest. Does oxidation occur within the endo-lysosomal compartment during peptide loading onto HLA-DRB1 or does it occur when exposed to the extracellular environment? If cysteine modifications occur during peptide loading onto the HLA-DRB1 complex and the binding register predictions denoted in Fig. 6C are correct for the adalimumab and ATR-107 peptides, then these modifications may be a previously unrecognized integral component of binding recognition. If the modifications occur after loading the peptide onto the HLA-DRB1 complex and exposure to environmental conditions, then the modification status may be reflective of the solvent accessibility of the cysteine residue. This latter scenario then implies incorrect in silico binding register predictions for the adalimumab and ATR-107 cysteine peptides. This error could be explained by the fact that the eluted ligand data used to date to train prediction algorithms do not contain such modifications. More importantly, it is also unknown how omission of modified cysteines may impact functionality when testing the T cell proliferative potential of MAPPs-derived peptides. If cysteine modification is a component required for either HLA-DRB1 binding or TCR recognition, then omission may alter results and potentially fail to highlight immunogenicity risk
Presentation of peptides from biotherapeutic proteins is a necessary, but not sufficient, step for developing an ADA response. The monoallelic HLA-DRB1 MAPPs platform described greatly facilitates our ability to reproducibly identify the peptides and ascertain the density of display from a diverse range of alleles with limited material requirements. However, the context of the presented sequence is a key parameter that determines the viability of the peptide to become a CD4 T cell epitope. The presence of a non-germline residue(s) in the HLA-DRB1 binding core, primarily in a solvent exposed position for interaction with TCR, is a major determinant of potential antigenicity. Even if the HLA-DRB1 peptide can result in CD4 T cell activation, the manifestation of the antigenicity is governed by the propensity of the biotherapeutic to gain entry into professional APCs. While the nature of the antigen targeted by the biotherapeutic may dictate such entry, many times yet undetermined biophysical properties and/or aggregation leads to enhanced uptake. Consideration of all three of these parameters (degree of HLA-DRB1 display, T cell proliferative potential of the displayed peptide(s), and APC uptake) will lead to the most accurate prediction of the immunogenicity potential for biotherapeutics.
We envision that the optimum application of this system is early in biotherapeutic discovery with molecules having diverse sequences. Ideally one or more molecules has no or limited HLA-DRB1 presentation of sequence(s) of concern. If a molecule with concerning HLA-DRB1 presentation is selected, then the immunogenic potential of those peptides from the biotherapeutic can be characterized for T cell proliferation potential in an allele-specific manner if desired. Confirmation of immunogenicity potential can then inform subsequent engineering efforts to remove or limit HLA-DRB1 presentation prior to clinical development. We also envision that this technology platform could be useful for the development of vaccines designed to elicit CD4 T cell responses.
The platform can be expanded to assess the HLA-DRB1 ligands for any proteinaceous component for which a human chimeric bridge antibody can be generated. For instance, a murine monoclonal antibody that recognizes a biotherapeutic peptide can be converted to human chimeric IgG. Co-dosing of this chimeric antibody will bind to the peptide of interest and initiate internalization via the exogenous BCR binding the chimeric antibody. We look forward to expansion of the immunopeptidome repertoire of a variety of endogenous proteins associated with autoimmunity, biotherapeutic peptides, and pathogens. This platform also provides a system to evaluate the impact of HLA-DO on immunopeptide repertoires from the various HLA-DRB1 alleles included in this data set. Although we do not anticipate any differences, it is difficult to compare immunopeptidomic results from our B cell platform to primary B cells owing to the requirement of BCR internalization to faithfully mimic encounters with potential antigens in vivo. Finally, while a current limitation, further modifications of this platform will allow investigation of HLA-DRB1 paralogs HLA-DRB3/4/5, as well as prominent HLA-DQ and HLA-DP alleles to gain a fuller HLA class II understanding for any exogenous protein of interest.
Methods
Endogenous HLA-DRB1 allele knockout
A human DRB1 double knockout Raji cell line was created by using guide RNAs targeting DRB1*03:01 and HLA-DRB1*10:01 alleles. DRB1 crRNAs were purchased from Horizon targeting DRB1.03 (target sequence CGTGACAAGCCCTCTCACAG) and the DRB1.10 allele (target sequence ATGGTGTGTCTGAGGCTCCC).
The DRB1.03 or DRB1.10 crRNA was combined with tracrRNA (Horizon) and Cas9 EGFP Nuclease mRNA (Horizon) and transfected into Raji cells (RRID:CVCL_0511) by Nucleofection with the Amaxa IIb instrument (Lonza) using Nucleofector V solution and program M-013. Nucleofected cells were recovered with 500 µl of complete medium containing 10% FBS and dispensed into a 12 well plate containing 1 ml of pre-warmed media and incubated at 37 C, 5% CO2 until the cells were 80-90% confluent. Cells were sorted by FACs for single cells in a 96 well plate and incubated at 37 °C; 5% CO2. Cells were screened by the T7EI assay for gene editing and several cell clones were identified for evaluation by western blot with DRB1 antibody (Thermo).
Universal exogenous BCR cell line construction
The Raji-derived 1E6 HLA-DRB1 deficient cell line was transfected with the exogenous universal chimeric IGHD BCR expression plasmid. Blasticidin resistance was used for stable cell line selection. The Amaxa nucleofector system (Lonza) was used following manufacturer instructions. Transfected cells were seeded into 96-well plate with media (RPMI 1640 with 2 mM L-glutamine (Gibco), 5% FBS (Gibco), 1x Antibiotic/Antimycotic (Gibco), and 5 µg/ml Blasticidin (Gibco)) was added to cells after 24 h to select for stable plasmid integration. Cells were cultured for 3–4 weeks and growth was visually monitored. Wells with growth were expanded into 24-well plate with 1 ml of selective media.
Expression and function of the exogenous chimeric anti-human IgG BCR was confirmed using flow cytometry. A biotinylated human IgG4 (4 µg/ml) with streptavidin-phycoerythrin (Invitrogen) (1 µg/ml) was used to stain 1 x10E6 cells. Clone 17g10 was further expanded for cell bank generation and used for monoallelic HLA-DRB1 cell line development.
Transfection of exogenous HLA-DRB1 alleles
On day 1, 2 x10E6 17g10 IgD engineered Raji B cells were transfected with 2 µg of HLA-DRB1 allele of choice using the Super PiggyBac Vector (SBI) containing puromycin resistance gene for stable cell line selection. Expression constructs for the following HLA-DR alleles were generated: HLA-DRB1*01:01, 03:01, 04:01, 04:04, 04:05, 07:01, 08:01, 08:03, 09:01, 11:01, 12:01, 13:01, 13:02, 15:01, 15:02. The transfections were done using the Amaxa instrument, and the Amaxa Cell Line Nucleofector Kit V (Lonza) protocol was followed. After transfection, the cells were incubated in warm media (RPMI 1640 with 2mM L-glutamine (Gibco), 5% FBS (Gibco), 1x Antibiotic/Antimycotic (Gibco), 5 µg/ml Blasticidin (Gibco)) for 3 days. On day 4, cells were resuspended in fresh selection media (RPMI 1640 with 2mM L-glutamine (Gibco), 5% FBS (Gibco), 1x Antibiotic/Antimycotic (Gibco), 5 µg/ml Blasticidin (Gibco), and 0.5 µg/ml Puromycin (Gibco)). Cells were then expanded to become confluent in a T75 cell culture flask and analyzed for functionality and sequence confirmed for accuracy.
HLA-DRB1 allele frequency calculations
The population risk for each allele in the United States and Japan is calculated using the HLA-DRB1 allele frequency for various ethnic groups from The Allele Frequency Net Database—Allele, haplotype and genotype frequencies in Worldwide Populations (allelefrequencies.net). The United States population considered for this example contain European (EUR), African American (AFA), Hispanic (HIS) and Chinese (CHN) ethnic groups. The Japanese population was not broken into various ethnic groups. An adjusted United States allele frequency was calculated using data from the 2020 US census (U.S. Census Bureau QuickFacts: United States) for each HLA-DRB1 allele in the monoallelic B cell panel using:
Equation 1:
$${US} ; {Adjusted} ; {Allele};{frequency}= , left(U{S}_{{EU}{R}_{{DRB}1}frequency}* 0.6right) \ +left(U{S}_{{{HIS}}_{{DRB}1}frequency}* 0.185right) \ +left(U{S}_{{{AFA}}_{{DRB}1}frequency}* 0.134right) \ +left(U{S}_{{{CHN}}_{{DRB}1}frequency}* 0.061right)$$
Most, but not all, individuals contain two different HLA-DRB1 alleles. The allele frequency was then converted to population frequency which represents the fraction of population containing the particular HLA-DRB1 allele using:
Equation 2:
$${Population}; {frequency}=1-{left(1-{allele};{frequency}right)}^{2}$$
Monoallelic B cell surface expression flow cytometry
2.5 x10E6 cells were spun down at 300 rcf for 10 min and resuspended in culture media and placed in an incubator at 37 °C and 5% CO2 for 24 h. After incubation, cells were spun down and resuspended in FACs buffer (1x PBS/0.5% BSA) and stained with primary cell markers: 1:10,000 Mouse a-HLA-DR antibody L243 (Sigma-Aldrich Cat# SAB4700731, RRID:AB_3676496), and 1:1000 biotinylated hIgG (internal source) for 20 min. The samples were washed in FACs buffer and resuspended in FACs buffer with secondary antibodies: 1:200 Rat a-mIgG2a-FITC (BD Biosciences Cat# 553390, RRID:AB_394828) cytometry using the BD FACs Melody (Becton Dickinson). The median fluorescent intensity was determined using the FlowJo Software. Example gating strategy is depicted in Supplementary Fig. 9.
Monoallelic B cell culture and dosing
Monoallelic HLA-DRB1 B cells were maintained in complete RPMI cell culture media (RPMI 1640 with 2 mM L-glutamine (Gibco); 5% FBS (Gibco), 1x Antibiotic/Antimycotic (Gibco), 5 µg/ml Blasticidin (Gibco), and 0.5 µg/ml Puromycin (Gibco)). For test article dosing, viability and cell count was determined with Trypan blue staining (Thermo Fisher Scientific, Countess cell counter). Cells from each selected HLA-DRB1 cell line were resuspended at a density of 1x10E6 cells/ml in 30 ml and plated 5 ml per well in a 6 well cell culture plate. Cells were dosed with test article at either 1, 10, or 100 µg/ml and placed in an incubator at 37 °C and 5% CO2 for 24 h. The cells were lysed the next day with RIPA lysis and extraction buffer (Thermo Fisher Scientific, 25 mM Tris•HCl pH 7.6, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) containing 10 unit/ml DNase (Roche) and 1 tablet of EDTA free protease inhibitors (Roche) per 10 ml of lysis buffer. The lysates were frozen at −80 °C.
Monocyte-derived dendritic cell culture and dosing
PBMCs were obtained through the American Red Cross and differentiated into immature DCs. On day 0, CD14+ cells were separated from the PBMCs by use of an AutoMACS instrument with anti-CD14 magnetic beads (Miltenyi Biotech). After viability determination with Trypan Blue (Thermo Fisher Scientific, Countess cell counter) purified CD14+ mononuclear cells were resuspended in complete RPMI Media (RPMI 1640 with 2 mM L-glutamine (Gibco), 5% Serum Replacement (Thermo Fisher Scientific), 5 mM HEPES (Gibco), 1% of 100X MEM nonessential amino acids (Gibco), 1% of 10,000 U/ml Penicillin/Streptomycin (HyClone), 1 mM sodium pyruvate (Gibco), 50 µM β-mercaptoethanol (Fisher Chemical), and 3.5% of DMEM high glucose (Gibco)) containing 40 ng/ml granulocyte monocyte-colony stimulating factor (human GM-CSF; Sargramostim, Sanofi-Aventis) and 20 ng/ml IL-4 (R&D Systems) to a density of 1x10E6 cells/ml and were differentiated into mature DCs in 6 well cell culture dishes (5 ml final culture volume per well) at 37 °C and 5% CO2. On day 4, test antibody was added to each well at the final concentration of 250 µg/ml. Cells were then incubated for 5 h prior to the addition of LPS (Sigma-Aldrich) at 1 µg/ml to transform the cells into mature DCs. Approximately 10x10E6 cells per test antibody condition were lysed in 1 ml of RIPA lysis buffer (Thermo Fisher Scientific, 25 mM Tris•HCl pH 7.6, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) containing 10 unit/ml DNase (Roche) and 1 tablet of EDTA free protease inhibitors (Roche) per 10 ml of lysis buffer. The lysates were frozen at −80 ˚C.
Monocyte-derived dendritic cell immunoprecipitation
An Agilent AssayMAP robot was used to isolate the HLA class II molecules in the lysate. One hundred micrograms of biotinylated anti-pan HLA class II antibody Tu39 (BD Biosciences Cat# 555556, RRID:AB_395938) or anti-HLA-DR antibody L243 (Sigma-Aldrich Cat# SAB4700731, RRID:AB_3676496) was immobilized on streptavidin cartridges (Agilent) by passing over the cartridge at 5 µl/min. The cartridge was washed with 50 µl of PBS three times. Lysates were thawed, passed over a 0.22 µm filter, and 1 ml of each sample was loaded onto a 96 well polypropylene plate. The lysate was aspirated into the syringes and passed over the antibody loaded affinity cartridge at 5 µl/min at room temperature. The cartridge was washed twice with 50 µl 100 mM ammonium acetate and once with 50 µl of water. The cartridge was eluted with 50 µl of 5% acetic acid with 0.1% TFA at 2 µl/min into a 96 well polypropylene PCR plate. The eluted peptides were passed over a 10k MWCO spin filter treated with 1 mg/ml BSA and 100 µg/ml angiotensin I peptide and washed with 5% acetic acid. The filtrate was loaded in a 96 well polypropylene PCR plate for LC/MS/MS analysis.
Monoallelic B cell immunoprecipitation
An Agilent AssayMAP robot was used to isolate the HLA-DR molecules in the lysate. One hundred micrograms of biotinylated anti-HLA-DR antibody (L243) was immobilized on streptavidin cartridges (Agilent) by passing over the cartridge at 5 µl/min. The cartridge was washed with 50 µl of PBS three times. Lysates were thawed, passed over a 0.22 µm filter, and 900 µl of each sample was loaded onto a 96 well polypropylene plate. The lysate was aspirated into the syringes and passed over the antibody loaded affinity cartridge at 5 µl/min at room temperature. The cartridge was washed twice with 50 µl 100 mM ammonium acetate and once with 50 µl of water. The cartridge was eluted with 50 µl of 5% acetic acid with 0.1% TFA at 2 µl/min into a 96 well polypropylene PCR plate. The eluted peptides were passed over a 10k MWCO spin filter treated with 1 mg/ml BSA and 100 µg/ml angiotensin I peptide and washed with 5% acetic acid. The filtrate was loaded in a 96 well polypropylene PCR plate for LC/MS/MS analysis.
LC/MS analysis of HLA class II eluted peptides
The samples were analyzed with a Thermo LUMOS mass spectrometer using a Thermo easy 1200 nLC-HPLC system. The separation was carried out with a 75 μm x 15 cm Thermo Easy-Spray Column coupled to an EASY-Spray source with an electrospray potential of 1.9 kV. The solvents were A—0.1% formic acid in water and B—80% acetonitrile with 0.1% formic acid. The gradient was 75 min using a flow rate of 300 nl/min, starting with a 60 min 2%–55% B ramp followed by a 1 min 55%–98% B ramp and a 4 min hold at 98% B. For the final 10 min, one additional 2%–98%–2% ramp was performed to mimic a blank run. The Lumos was run with scans at 240,000 resolution in the orbitrap followed by a 3 s data dependent MS/MS cycle comprised of ion trap rapid scans where +2 ions were fragmented by HCD (CE of 15,22,28) and +3 and +4 ions were fragmented by HCD (CE of 15,22,28) and EThcD (Calibrated Charge-Dependent ETD parameters and supplemental HCD (CE of 50)).
Database searching
Peaks Online X v1.5 (Bioinformatics Solutions Inc.) was used to analyze raw data. Spectra were searched against a combined human (uniprot_2023_04, no isoforms) and bovine (uniprot_2023_05) database with antibody variable region sequences removed and test article sequences appended. No enzyme searches for peptides 9–30 residues in length were performed using 10 ppm precursor mass error and 0.30 Da fragment mass error tolerances. At maximum, 3 PTMs were considered per peptide. Variable PTMs included in the search were: deamidation (NQ), pyro-Glu (QE), oxidation (MHWC), additional Trp oxidations (kynurenine, hydroxykynurenine), additional Cys oxidations (cysteic acid, sulfinic acid), and other Cys specific modifications (Cysteinylation, glutathione disulfide, half of disulfide bridge). Results were filtered for 1% PSM false discovery rate. Data was exported from PEAKS as a project level protein-peptides.csv file.
Data summarization
The PEAKS protein-peptide.csv file was uploaded to KNIME 5.1.3 (Open for Innovation | KNIME)32 and peptides were filtered for length (9–25 residues) and searched against the dosed test article heavy chain and light chain sequences. Matching peptides were aligned against the parent sequence and summarized by count at each residue. Graphpad Prism 10.1.2 (GraphPad Software, Boston, Massachusetts USA, www.graphpad.com) was used to visualize peptide data as heatmaps. Adalimumab non-germline residues were defined after variable region alignment using ANARCI v1.1. Amino acid frequencies calculated from the repertoire at each ANARCI aligned position were used to identify potentially immunogenic peptide sequences. The repertoire was constructed using publicly available sequences. The antibody sequence repertoire used for this analysis consisted of 202,395,793 heavy chain and 20,300,442 light chain sequences (11,969,289 Kappa, 8,331,153 Lambda).
In silico tools
NetMHCIIpan 4.3 (NetMHCIIpan 4.3 – DTU Health Tech – Bioinformatic Services)28 was used to determine the binding register of peptides derived from adalimumab and ATR-107 H3 sequences. Peptide sequence and corresponding HLA-DRB1 allele were used as the input parameters.
For the generation of logo plots, data was summarized for 60 samples (4 samples per HLA-DR allele). For each allele, peptide lists from all runs were combined and deduplicated (disregarding modification and charge state). Peptide lists were paired with the HLA-DR allele in which they were identified and used as input to MHCMotifDecon-1.028,33 (MHCMotifDecon 1.0 – DTU Health Tech – Bioinformatic Services) using 9-25 peptide length and 20% Rank trash threshold.
Statistics and reproducibility
Graphpad Prism 10.1.2 was used to generate descriptive statistics (mean, Std. Dev, SEM, 95% CI and CV) for the golimumab variable region immunopeptidome reproducibility data. Individual replicates (N = 13–15) per HLA-DRB1 cell line were taken from distinct samples over the course of several cell line passages. A total of seven cell lines were examined.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Information. Adalimumab immunopeptidome data used for Fig. 5 and Table 2 is included in Supplementary Data S1. Data used for Fig. 3 and Supplementary Table 2 is included in Supplementary Data S2. Data used for Fig. 6 is included in Supplementary Data S3. In addition, .raw mass spectrometry files, identified peptide lists, and associated .fasta databases for the entire immunopeptidome data have been deposited in the database MassIVE (dataset MSV000094594). https://doi.org/10.25345/C5NP1WW03. No custom code was used for the manuscript.
References
-
Neumann, T. A. & Foote, M. Megakaryocyte growth and development factor (MGDF): an Mpl ligand and cytokine that regulates thrombopoiesis. Cytokines Cell Mol. Ther. 6, 47–56 (2000).
-
Casadevall, N. et al. Pure red-cell aplasia and antierythropoietin antibodies in patients treated with recombinant erythropoietin. N. Engl. J. Med. 346, 469–475 (2002).
-
Hunley, T. E. et al. Nephrotic syndrome complicating alpha-glucosidase replacement therapy for Pompe disease. Pediatrics 114, e532–e535 (2004).
-
Kessler, C. M. & Ortel, T. L. Recent developments in topical thrombins. Thromb. Haemost. 102, 15–24 (2009).
-
Yamamoto, F. et al. Capturing differential allele-level expression and genotypes of all classical HLA loci and haplotypes by a new capture RNA-Seq method. Front. Immunol. 11 https://doi.org/10.3389/fimmu.2020.00941 (2020).
-
Billiet, T. et al. Immunogenicity to infliximab is associated with HLA-DRB1. Gut 64, 1344–1345 (2015).
-
Hunt, D. F. et al. Characterization of peptides bound to the class I MHC molecule HLA-A2.1 by mass spectrometry. Science 255, 1261–1263 (1992).
-
Hunt, D. F. et al. Peptides presented to the immune system by the murine class II major histocompatibility complex molecule I-Ad. Science 256, 1817–1820 (1992).
-
Röhn, T. A. et al. A novel strategy for the discovery of MHC class II-restricted tumor antigens: identification of a melanotransferrin helper T-cell epitope. Cancer Res. 65, 10068–10078 (2005).
-
Knierman, M. D. et al. The human leukocyte antigen class II immunopeptidome of the SARS-CoV-2 spike glycoprotein. Cell Rep. 33, 108454 (2020).
-
Brito-Sierra, C. A., Lannan, M. B., Siegel, R. W. & Malherbe, L. P. The HLA class-II immunopeptidomes of AAV capsids proteins. Front. Immunol. 13, 1067399 (2022).
-
Brito-Sierra, C. A., Lannan, M. B., Malherbe, L. P. & Siegel, R. W. The HLA class I immunopeptidomes of AAV capsid proteins. Front. Immunol. 14, 1212136 (2023).
-
Hamze, M. et al. Characterization of CD4 T cell epitopes of infliximab and rituximab identified from healthy donors. Front. Immunol. 8 https://doi.org/10.3389/fimmu.2017.00500 (2017).
-
Quarmby, V., Phung, Q. T. & Lill, J. R. MAPPs for the identification of immunogenic hotspots of biotherapeutics; an overview of the technology and its application to the biopharmaceutical arena. Expert Rev. Proteom. 15, 733–748 (2018).
-
Walsh, R. E. et al. Post-hoc assessment of the immunogenicity of three antibodies reveals distinct immune stimulatory mechanisms. MAbs 12, 1764829 (2020).
-
Siegel, M. et al. Validation of a dendritic cell and CD4+ T cell restimulation assay contributing to the immunogenicity risk evaluation of biotherapeutics. Pharmaceutics 14 https://doi.org/10.3390/pharmaceutics14122672 (2022).
-
Lanzavecchia, A., Corti, D. & Sallusto, F. Human monoclonal antibodies by immortalization of memory B cells. Curr. Opin. Biotechnol. 18, 523–528 (2007).
-
Lanzavecchia, A. Antigen-specific interaction between T and B cells. Nature 314, 537–539 (1985).
-
Pulvertaft, J. V. Cytology of Burkitt’s tumour (African lymphoma). Lancet 1, 238–240 (1964).
-
Radaev, S. et al. Structural and functional studies of Igαβ and its assembly with the B cell antigen receptor. Structure 18, 934–943 (2010).
-
Gonzalez-Galarza, F. F. et al. Allele frequency net database (AFND) 2020 update: gold-standard data classification, open access genotype data and new query tools. Nucleic Acids Res. 48, D783–D788 (2020).
-
Ridker, P. M. et al. Lipid-reduction variability and antidrug-antibody formation with bococizumab. N. Engl. J. Med. 376, 1517–1526 (2017).
-
Hua, F. et al. Anti-IL21 receptor monoclonal antibody (ATR-107): safety, pharmacokinetics, and pharmacodynamic evaluation in healthy volunteers: a phase I, first-in-human study. J. Clin. Pharm. 54, 14–22 (2014).
-
Cohen, S. et al. Immunogenicity risk assessment for biotherapeutics through in vitro detection of CD134 and CD137 on T helper cells. mAbs 13, 1898831 (2021).
-
Xue, L., Hickling, T., Song, R., Nowak, J. & Rup, B. Contribution of enhanced engagement of antigen presentation machinery to the clinical immunogenicity of a human interleukin (IL)-21 receptor-blocking therapeutic antibody. Clin. Exp. Immunol. 183, 102–113 (2015).
-
Fleischmann, R. M. et al. Long-term efficacy, safety, and immunogenicity of the adalimumab biosimilar, PF-06410293, in patients with rheumatoid arthritis after switching from reference adalimumab (Humira®) or continuing biosimilar therapy: week 52-92 data from a randomized, double-blind, phase 3 trial. Arthritis Res. Ther. 23, 248 (2021).
-
Meunier, S. et al. Impact of human sequences in variable domains of therapeutic antibodies on the location of CD4 T-cell epitopes. Cell Mol. Immunol. https://doi.org/10.1038/s41423-019-0304-3 (2019).
-
Nilsson, J. B. et al. Accurate prediction of HLA class II antigen presentation across all loci using tailored data acquisition and refined machine learning. Sci. Adv. 9, eadj6367 (2023).
-
Stern, L. J. et al. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature 368, 215–221 (1994).
-
Abelin, J. G. et al. Defining HLA-II ligand processing and binding rules with mass spectrometry enhances cancer epitope prediction. Immunity 51, 766–779.e717 (2019).
-
Strazar, M. et al. HLA-II immunopeptidome profiling and deep learning reveal features of antigenicity to inform antigen discovery. Immunity 56, 1681–1698.e1613 (2023).
-
Fillbrunn, A. et al. KNIME for reproducible cross-domain analysis of life science data. J. Biotechnol. 261, 149–156 (2017).
-
Kaabinejadian, S. et al. Accurate MHC motif deconvolution of immunopeptidomics data reveals a significant contribution of DRB3, 4 and 5 to the total DR immunopeptidome. Front. Immunol. 13 https://doi.org/10.3389/fimmu.2022.835454 (2022).
Acknowledgements
We thank Drs. Laurent Malherbe and Andrea Ferrante for fruitful discussions and critical review of the manuscript. We thank Courtney Alexander for antibody production. We thank Vidula Garud for technical assistance with immunoprecipitations. This work was supported by Eli Lilly and Company.
Ethics declarations
Competing interests
All the authors have equity interest in Eli Lilly and Company. R.W.S. is inventor on patent application submitted by Eli Lilly and Company that covers monoallelic B cell immunopeptidomic platform.
Peer review
Peer review information
Communications Biology thanks Adán Pinto-Fernández and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Dr Isabela Pedroza-Pacheco and Dr Ophelia Bu. [A peer review file is available].
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lannan, M.B., Ming, W., Spindler, L.J. et al. A universal monoallelic human leukocyte antigen class II immunopeptidomic platform for defining the immunogenicity potential of therapeutic proteins. Commun Biol 8, 1243 (2025). https://doi.org/10.1038/s42003-025-08692-7
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s42003-025-08692-7