
Introduction
Botanical insecticides are a safer, eco-friendly alternative to synthetic chemicals, offering effective pest control with minimal environmental impact1. Simarouba glauca (Simaroubaceae), the paradise tree or bitterwood, or aceitillo, is scientifically named, and it is indigenous to the Caribbean, Central America, and northern South America. This tree is highly valued not only for its timber but also for its medicinal properties, with traditional uses spanning a variety of ailments. The bark, leaves, and seeds of S. glauca are rich in bioactive compounds that demonstrate potential insecticidal capabilities. Research has shown that the antifeedant effect of bark extracts is considerably more pronounced compared to those from leaves and seeds, primarily due to the cytotoxic and insecticidal properties of quassinoids in the extracts2. Specifically, the bark extract achieved a notable 100% inhibition of feeding against Plutella xylostella Linnaeus (Plutellidae: Lepidoptera) and Helicoverpa armigera Hubner (Noctuidae: Lepidoptera) at concentrations of 20% and 25%, while the leaf extract showed a 95% inhibition at 25%3. Nonetheless, the specific metabolites responsible for these effects in S. glauca remain unreported. Furthermore, extracts from S. glauca have been shown to significantly reduce key lepidopteran pests such as H. armigera and P. xylostella, leading to high mortality rates and pronounced antifeedant effects3 The application of S. glauca seed oil resulted in 100% larval mortality in Eligma narcissus Cramer (Nolidae: Lepidoptera), Hyblaea puera,(Hyblaeidae: Lepidoptera), attributed to hydrolytic dissociation and excessive water loss4. These studies lay the groundwork for understanding the effects of S. glauca extracts.
Maize Zea mays (Poaceae), the “Queen of cereals,” is cultivated globally, with immense economic importance due to its diverse applications as food, feed, and raw material for various industries5. In 2020, global maize production reached 1,162.4 million metric tonnes (Mt) from 202 million hectares (Mha)6. It is projected that by 2030, maize will become the leading cereal crop in terms of production volume7. India accounts for 4% of the world’s maize area and 2% of total production, placing it fourth in area and seventh in production among maize-growing nations. In the 2018–19 season, 9.2 Mha were dedicated to maize cultivation in India. Production increased significantly from 1.73 Mt in 1950–51 to 27.8 Mt in 2018–19, nearly a 16-fold rise8. In India, maize is the third most important cereal crop in area and annual production, next to wheat and rice9. The successful cultivation of maize is notably endangered by biotic and abiotic factors, with biotic factors particularly impacting yields. Among these biotic factors, a dozen insect pests and pathogens pose the greatest threats to maize production10. Concerning pests, the fall armyworm Spodoptera frugiperda, a notorious polyphagous pest that attacks over 353 species from 76 different plant families11, enormously destroys maize, its primary host plant. This pest remained confined to the Americas until 2016, spreading to other continents, particularly West Africa12. Its spread continued to Asia and Africa, reaching Australia in 201813. The first instance of S. frugiperda in maize was reported in Karnataka state, India, in 201814, and within three years, it spread across all maize-growing regions of the country14,15. Following the S. frugiperda invasion, a loss of 8.3 to 20.6 Mt of maize yields each year and a decline of 20 to 50% of maize production without adequate control methods is projected16,17. S. frugiperda targets maize in its initial growth stages, where adults deposit eggs in groups on the underside of young leaves. Once hatched, early instar larvae feed by scraping the leaves, while later instar larvae conceal themselves in leaf whorls, consuming the developing shoot tissues. This feeding activity results in significant damage, such as distortion, stunting of the plant’s aerial parts, wilting, and leaf rolling18, ultimately resulting in yield losses exceeding 30% within a single season19,20,21. The larvae also feed on tassels and cobs, leading to damage to seeds and a decrease in their quality21. Pest management in maize gained significant attention in India following the S. frugiperda invasion, given the considerable round-the-year damage it inflicts throughout the crop’s lifecycle. The hidden feeding habits of S. frugiperda larvae pose a challenge in its management, reducing the effectiveness of insecticides, including newer classes of chemicals12,22,23. Inappropriate application of traditional pesticides causes pest resistance and unfavourable impacts on species that are not the intended target.
As a result, the persistent interactions between pests and plants require multiple applications of insecticides since current methods frequently prove inadequate. This scenario has sparked interest in alternatives to synthetic insecticides and prompted ongoing research into botanical pesticides that offer safe pest control solutions, aiming to reduce negative impacts on the environment and agriculture while safeguarding human and animal health. However, it is crucial to identify botanical insecticides with specific functionalities tailored to different pests and comprehend their mechanisms of action against these targeted insects. The present study intends to investigate the insecticidal attributes of S. glauca, employing its various plant parts and solvent extracts to effectively address the threat posed by S. frugiperda to maize.
Results
Isolation, extraction, and quantification of crude extract from different parts of the S. glauca
The extraction of bioactive compounds from various plant parts of S. glauca using different solvents resulted in significant variability in the yield of final crude extracts. The recovery percentages of the extractants provided preliminary insights into the concentration of bioactive compounds across the different plant parts, revealing differences based on the solvent used. Among the plant parts analyzed, seeds exhibited a remarkable recovery, with hexane yielding 78.66% and cold-pressed oil yielding 52.32 percent, while no recovery was observed for ethanol and ethyl acetate. A lower final crude recovery was recorded with ethanol (10.58%) and ethyl acetate (3.28% and 3.63%) extracts of leaves and bark, respectively (Table 1).
Bioassay
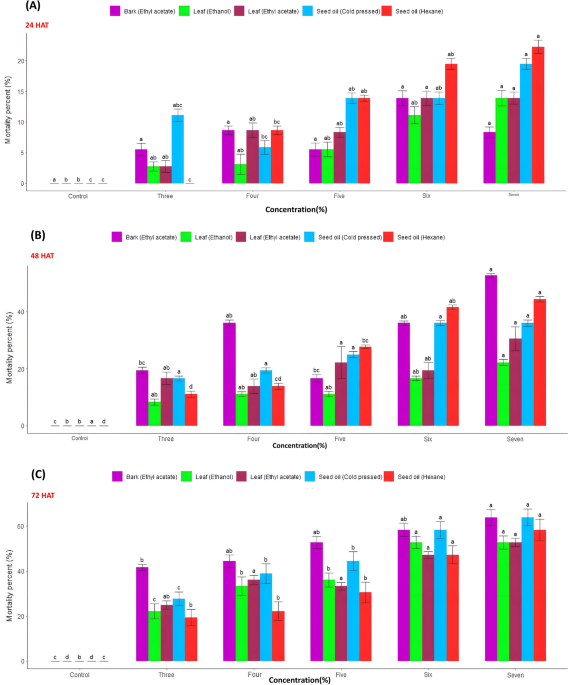
The probit analysis of S. glauca extracts against S. frugiperda third instar larvae demonstrated varying effectiveness. The ethyl acetate extract of bark showed the lowest Lethal Concentration (LC₅₀) (4.80%), followed by leaf extracted with ethanol LC50 (5.94%), while leaf ethyl acetate extract showed the highest LC₅₀ of 6.98% (Table 2 and Supplementary Fig. 1). All extracts fitted the probit model (p > 0.05) (Table 2 and Supplementary Fig. 1). The percent mortality of different extracts is illustrated in Fig. 1.

Percent mortality of S. frugiperda third instar larvae treated with extracts of different plant parts [bark (ethyl acetate), leaf (ethyl acetate), leaf (ethanol), seed oil (cold pressed) and seed oil (hexane)] of S. glauca tested at various concentrations. Means were separated by Tukey’s HSD (honestly significant difference) test (p = < 0.5%); different letters above the Mean ± SE bars indicate statistically significant differences among extracts at each tested concentration. (A) 24 Hours After Treatment (HAT); (B) 48 HAT; (C) 72 HAT.
The antifeedant activity demonstrated remarkable variability among the extracts (bark, seed, and leaf) assessed at different concentrations. Notably, the cold-pressed seed oil of S. glauca emerged as the most potent, exhibiting impressive antifeedant activity of 46.1, 51.0, and 52.3% after 24, 48, and 72 HAF (Hours After Feeding) at 7%, respectively. In contrast, the control group treated with acetone exhibited no antifeedant effects and recorded no mortalities. In contrast, the hexane-extracted seed oil demonstrated considerable efficacy, recording activity levels of 34.60%, 42.10%, and 44.21% after 24, 48, and 72 HAF, respectively, at the same concentration. Conversely, the leaf extract using ethanol exhibited the lowest feeding inhibition, with values of 36.2%, 39.0%, and 42.1% after 24, 48, and 72 HAF at 7% concentration (Fig. 2). The control group showed no antifeedant activity. Notably, all other extractants revealed significant feeding inhibition, ranging from 21 to 45% (Fig. 2), underscoring their potential as effective natural deterrents. Additionally, the bioassay results for bark extracted with ethyl acetate fractions indicated that fractions 2 (50.0% and 55.34%), 3 (41.7% and 44.89%), 4 (52.8% and 56.1%), and 5 (44.4% and 47.25%) exhibited the highest percentage of mortality and antifeedant activity, respectively. In contrast, the control group treated with acetone showed neither mortality nor antifeedant activity. This highlights a significant difference in their effectiveness (Fig. 3).

Percent antifeedant activity of S. frugiperda third instar larvae treated with extracts of different plant parts of S. glauca tested at various concentrations. Means were separated by Tukey’s HSD test (p = < 0.5%); Different letters above the Mean ± SE bars indicate statistically significant differences among extracts at each tested concentration. (A) 24 Hours After Treatment (HAT); (B) 48 HAT; (C) 72 HAT.

Percent mortality of S. frugiperda third instar larvae treated with different fractions of bark extracts of S. glauca tested at various concentrations. Means were separated by Tukey’s HSD test (p = < 0.5%); Different letters above the Mean ± SE bars indicate statistically significant differences among extracts at each tested concentration. (A) 24 Hours After Treatment (HAT); (B) 48 HAT; (C) 72 HAT.
Metabolite dynamics in plant parts using different solvents
The metabolite composition of extractants revealed significant differences based on both the plant parts and the solvents used for their extraction. Results from Gas Chromatography-Tandem Mass Spectrometry (GC–MS/MS) indicated a differential pattern of the existence of metabolites in the seeds, leaves, and bark of S. glauca and their chromatograms, which were presented in Supplementary Fig. 5. The metabolomic profiles of the ethyl acetate extract from the bark and the ethanol extract from the leaves contain overlapping compounds. The n-hexane extract of seeds demonstrated a diverse range of metabolites particularly rich in fatty acids. Notably, a few metabolites that tested and were reported to have insecticidal activity, including gamma-Sitosterol, Octadecanal, Phytol, Linoleic acid ethyl ester, Octadecanoic acid, and Campesterol, were also identified from plant parts, substantiating the insecticidal potential. The distinctive and shared metabolite storage characteristics across various plant parts and fractions are summarized in Supplementary File 1.
The analysis of differential regulation across tissue comparisons of bark vs. seed, seed vs. leaf, and leaf vs. bark revealed significant patterns. Highly upregulated features, such as m/z 383.3 and m/z 88.05 log₂ Fold change (log₂FC), were prominent in seed and leaf samples, indicating robust activation in these tissues. Conversely, features like 129.05 log2FC were strongly downregulated, particularly in seeds and leaves compared to the bark. Several features exceeded statistical thresholds (p < 0.05), demonstrating biological and statistical significance. Tissue-specific differences were evident, with bark suppressing certain features, while seed and leaf tissues exhibited strong activation of specific genes. These findings highlight distinct transcriptional profiles across tissues, offering potential insights into tissue-specific gene functions (Fig. 4).


Volcano plots showing the differential metabolite concentrations in the extracts of different plant parts of S. glauca analyzed using GC–MS/MS. (A) Leaves versus seeds; (B) Bark versus seeds; (C) Bark versus leaves.
Principle Component Analysis (PCA) and Heatmap
The PCA plot illustrated the variable important projections (VIP) scores for selecting chemical compounds, highlighting the significant metabolites that warranted retention for subsequent downstream analysis. Among the compounds evaluated, notable metabolites such as 9-octadecenoic acid, Methyl cis-11-eicosenoate, and gamma-Sitosterol exhibited the highest VIP scores within the various plant parts examined. In contrast, the bark fractions demonstrated elevated VIP scores for compounds including coniferyl aldehyde, elaidic acid, and 3,4-dihydroxymethylbenzoate, indicating their importance. Each compound was represented by a distinct dot on the plot, strategically positioned according to its corresponding VIP score. To enhance interpretation, a color scale was incorporated on the right side of the plot, showcasing a gradient that ranged from low (depicted in blue) to high VIP scores (illustrated in red). The PCA biplots illustrated the variability and clustering patterns across distinct datasets, highlighting similarities and differences among the analyzed groups. The first biplot focused on plant parts, with principal component 1 (PC1) and PC2 explaining 46.7% and 26.6% of the variance, respectively. Three clusters were formed, with bark in a distinct and separate group along PC1, indicating unique biochemical or structural characteristics. Leaf extracts (Ethyl acetate and Ethanol) and seed oils (Hexane and Cold pressed) formed a cluster with overlapping points, reflecting shared chemical compositions or functional similarities. The tight grouping of these samples contrasted with the clear separation of bark, emphasizing differences in traits between these plant parts (Fig. 5A and C). In the second biplot, fractions were distributed across three distinct clusters, with PC1 and PC2 explaining 30.8% and 24.7% of the variance, respectively (Fig. 5). Fractions 1 and 2 clustered closely together, suggesting they shared similar characteristics. In contrast, Fractions 3 and 4 overlapped within a separate cluster. Fractions 5 and 6 formed another distinct cluster, indicating unique features compared to the others. The observed separation suggested that similar properties or conditions influenced fractions within the same cluster, whereas fractions in different clusters had distinct properties (Fig. 5B and D). The quantitative expression of metabolites was represented in the heatmap (Fig. 6).

Principal Component Analysis (PCA) indicating the variance of distribution and clustering patterns among different extracts of S. glauca; Biplot for (A) Plant parts, (C) Bark fractions, (B) scree plot for plant parts and (D) scree plot for Bark fractions.

Heatmap showing the metabolite distribution and their relative abundance among different extracts of S. glauca; (A) Plant parts and (B) Bark fractions.
PLS, sPLS, and SOM analysis
The results showed that the bark was clustered solely. In contrast, extracts of cold-pressed and hexane seed oil were clustered together in both k-means and SOM analysis. This revealed the metabolic complexity and superiority of bark over other extractants. The correlation analysis showed the interdependence of compound expressions from different plant parts (Fig. 6). Concerning bark fractions, PLSDA, sPLSDA, and SOM analysis showed that a similar clustering pattern withstood the above PCA results. Correlation analysis of bark fractions also revealed no relationship among the fractions (Supplementary Figs. 2, 3, and 4).
Functional enrichment and network analysis
Since S. glauca is renowned for its medicinal properties, much of the existing annotation was linked to human diseases. The results of the functional enrichment analysis and the potential biological interactions for different plant part fractions are illustrated in Fig. 7. Network analysis of the metabolites obtained from the various parts and fractions indicated that many of these metabolites were associated with pathways related to fatty acid and terpenoid biosynthesis (Figs. 7 and 8). Furthermore, the metabolite-to-gene and gene-to-gene interaction plot suggested that the terpenoids identified in this study might interact with CYP genes, which indirectly highlighted the insecticidal potential of the bioactive compounds present in S. glauca (Figs. 7, 8, 9, and 10).

Enrichment plot showing the metabolite distribution and their relative abundance among different extracts of S. glauca; (A) Plant parts and (B) Bark fractions.

Metabolite to metabolite interaction plot among different extracts of S. glauca; (A) Plant parts and (B) Bark fractions.

Global network of metabolites derived from different extracts of S. glauca associated with biological pathways (www.kegg.jp/kegg/kegg1.html).

Metabolites associative network of metabolites derived from different extracts of S. glauca; (A) Metabolites to gene interaction plot and (B) Gene to gene interaction of terpenoid pathway.
Differential gene expression analysis
The transcriptome assembly was performed using the Spliced Transcripts Alignment to a Reference (STAR) aligner, employing a reference-based strategy. For quantifying transcripts, RNA-Seq by Expectation–Maximization (RSEM) was utilized to attain accurate measurements of gene expression. The RNA sequencing produced 30 million reads, resulting in a dataset size of 6 GB. Differential expression analysis using DESeq2 identified a total of 734 differentially expressed genes (DEGs). Among these, 383 genes showed upregulation, while 392 were downregulated; 44 genes did not demonstrate significant changes and PCA biplot illustrated that the bark-treated and control formed distinct clusters, explaining 94% of total variations (Fig. 11a–c). The heatmap and correlation heatmap demonstrated a noticeable difference in transcriptional responses, highlighting the potential of insecticidal activity (Fig. 11d and e). To ensure that the findings were biologically relevant, DEGs were filtered using a significance threshold of p < 0.05 and a log₂FC exceeding 2 or falling below − 2. The results of DEGs and the top three significantly upregulated and downregulated genes are given in Supplementary File 2 and Fig. 7. These findings offer valuable insights into the transcriptional changes occurring under experimental conditions, enhancing our understanding of the regulatory pathways involved in the investigation.

Differentially expressed genes after 48 HAT in S. frugiperda fourth instar larvae treated with bark extract of S. glauca at LC50 concentration; (A) Volcano plot; (B) Metabolites associative plot; (C) PCA biplot; (D) Heatmap; (E) Correlation heatmap.
Overrepresentation analysis
This functional annotation identifies biological processes, molecular functions, and KEGG pathways that may be related to insect mortality and antifeedant activity. There is an enrichment of primary metabolic pathways, macromolecular metabolic pathways, and protein digestion pathways, which implies that interference with these processes can compromise normal physiological functions (Fig. 12A and B). A disruption in protein digestion and absorption could result in malnutrition and hinder insect development, leading to increased insect mortality. In addition, thyroid hormone signaling pathways may influence insect growth and survival due to possible hormonal imbalances. As a result of disruptions in muscle function via the cytoskeleton in muscle cells and the cGMP-PKG signalling pathway (Fig. 12C), paralysis or diminished feeding behavior may occur. Feeding inhibition may result from dysfunctional ion binding and organic cyclic compound binding proteins during loss of coordination. Furthermore, lysosomal pathways and membrane-related gene ontology terms are significantly overrepresented (Fig. 12B).

Differentially expressed genes after 48 HAT in S. frugiperda fourth instar larvae treated with bark extract of S. glauca at LC50 concentration; (A) Gene ontology (GO) of up and down-regulated genes; (B) GO with category; (C) KEGG pathway enrichment.
Molecular docking analysis
The docking results with Cytochrome P450 family 6 (CYP6) and Glutathione S-transferase 2 (GST) revealed that gamma-Sitosterol (− 9.9 and − 10.5 kcal/mol) and Lanosta–8,24–dien–3–one (− 10.4 and 9.5 kcal/mol) exhibited the highest binding affinities, respectively. These results set a foundation for predicting the presumed growth inhibitory activities of the ligands. This prediction computationally proves that these ligands contribute to larval mortality and affect the metabolic efficiency of enzymes involved in detoxifying xenobiotics. Furthermore, botanicals are recognized for their significant antifeedant and repellent properties for insects. In this study, molecular docking interactions employing Odorant-binding protein (OBP) sequences derived from S. frugiperda demonstrated that the primary antennal sensor, OBP 1, exhibited the highest binding energy scores of − 10.5 and − 10.8 kcal/mol with gamma-Sitosterol and Lanosta-8,24-dien-3-one, respectively (Table 3 and Fig. 13). Moreover, across various subclasses of OBP, gamma-Sitosterol and Lanosta-8,24-dien-3-one consistently displayed considerable binding, often exceeding − 8 kcal/mol across all OBP proteins. (Table 3).

Molecular docking results against S. frugiperda protein targets; a) CYP with gamma-Sitosterol (a1) and Lanosta (a2), b) GST with gamma-Sitosterol (b1) and Lanosta (b2) and c) OBP1 with gamma-Sitosterol (c1) and Lanosta (c2) GST with Gamma-Sitosterol; (B) CYP with Lanosta-8,24-dien-3-one; (C) OBP1 with Gamma-Sitosterol and Lanosta-8,24-dien-3-one.
Discussion
S. glauca is native to central and northern south America and is well-known for its medicinal properties and the edible oil extracted from its seeds24. Numerous studies have highlighted its potential as an anticancer agent. While the metabolomic composition of S. glauca has been extensively investigated, its insecticidal properties have not been thoroughly explored. Insecticidal compounds are essential for effective pest management, with a strong emphasis on environmental safety. We utilized GC–MS/MS to analyze bioactive compounds extracted from various plant parts, ensuring a thorough evaluation of their potential. The extraction was done through a dry matter method, employing a Soxhlet apparatus, followed by solvent removal via rotary evaporation. The resulting compounds were then subjected to preliminary dose–response analysis to evaluate their insecticidal activity. Notably, extracts from the seed kernel and bark demonstrated significant insecticidal properties.
The bioactive compounds in these extracts were profiled to identify metabolic differences based on the solvents used. S. glauca has been extensively studied for its medicinal properties, particularly in cancer treatment. Various solvents, including water, 70% ethanol, ethyl acetate, hexane, and chloroform, were used to extract bioactive compounds from S. glauca leaves25. Triterpenoids, including betulinic acid, were identified in the terpene fraction of S. glauca leaves through gas chromatography-mass spectrometry (GC–MS) and high-performance thin-layer chromatography26. Lin et al. (2021) identified 102 insecticidal compounds, including nor-triterpenes, tetracyclic triterpenes, and pentacyclic triterpenes, which are recognized for their antifeedant, growth-inhibitory, and toxic properties. Numerous compounds exhibited significant insecticidal activity against various insect taxa, highlighting the prospective role of terpenes in pest management. The current study has additionally identified terpenoids, indicating the necessity for further investigation regarding their insecticidal characteristics. This reinforces the potential of terpenes within metabolomic fractions, and future investigations could further analyze their insecticidal effectiveness and pinpoint promising insecticidal candidates from the extensive data profiles. A phytochemical study of S. glauca bark extracts revealed alkaloids, phenols, tannins, and flavonoids, suggesting antioxidant activity27. These findings align with the current study, which identified similar chemical groups. Plant solvent extracts, particularly those rich in flavonoids, terpenoids, and phenolic compounds, are known for their potent anticancer activities28. Quassinoids, a group of bitter compounds found in the Simaroubaceae family, are recognized for their anticancer properties29. Research by Jose, et al.30 confirmed the presence of gallic acid, gentisic acid, and other phenolic components in chloroform and ethyl acetate extracts, with S. glauca exhibiting potent anticancer activity against various malignant cells. Furthermore, S. glauca leaf methanolic extract was shown to inhibit SCC9 (squamous cell carcinoma) and HCT 116 cell (colorectal carcinoma cells) proliferation at a concentration of 312.2 μg/ml31. These reports demonstrated the presence of growth-inhibitory compounds, providing preliminary evidence that SG extract may also possess growth-inhibitory activity against insects.
The current study further supported the growth-inhibitory activity against insects with a significant number of phenolic compounds potentially contributing to insect mortality, as confirmed by the dose–response analysis conducted. Phenolic compounds can induce cancer cell death by elevating oxidative stress, blocking proliferative pathways, and activating apoptotic signaling cascades32. Apoptosis is a key mechanism in anticancer therapies, and phytochemical fractions rich in anticancer compounds have been shown to trigger this process33,34. Apoptosis generally occurs in early and late stages, marked by plasma membrane modifications, caspase-3 activation, mitochondrial signaling, membrane damage, phagocyte recognition, and the release of intracellular contents35. The findings of this study demonstrate that the activity of S. glauca extract is mediated through comparable mechanisms. GC–MS analysis unequivocally identified a diverse array of metabolites in the methanol and acetone extracts, including significant compounds such as umckalin (1.19%), cis-sinapyl alcohol (2.33%), 2,3-trimethylene-4-pyrone (1.79%), and undecane (8.52%). This further reinforces the validity of the metabolomic profiling conducted in this research36. Remarkably, the present research has revealed that lanosterol and gamma-sitosterol are present in the highest concentrations, and they are classified within the sterol chemical group. Beta-Sitosterol, a compound derived from Abutilon indicum, exhibited notable larvicidal activity, leading to an increase in larval mortality observed within 24 h37. Furthermore, Gamma-Sitosterol is categorized as a WHO Class III toxicant, which induced significant DNA damage38. These findings substantiate the theoretical assertions regarding the insecticidal properties of these compounds. Bioactive chemicals affect insect mortality and antifeedant effects in a complex manner, including cellular damage, metabolic abnormalities, and neuromuscular dysfunction. These substances interfere with vital physiological functions, which results in developmental problems and malnutrition. The next sections concentrate on these crucial features. Malignancy in insects results from bioactive chemicals interfering with macromolecule metabolism and protein digestion. Moreover, antifeedants may disrupt neural functioning, lowering feeding activity by affecting muscle performance and signaling pathways39.
A phytochemical study identified bioactive compounds, such as steroids and terpenoids, in S. glauca seeds, suggesting insecticidal properties. Research by de Souza, et al.40 demonstrated that aqueous extracts of Simarouba species suppressed the Plutella xylostella. Additionally, ethanolic extracts of S. glauca have shown potent inhibitory actions against pathogenic fungi like Fusarium oxysporum and Aspergillus parasiticus41. These findings collectively suggested that S. glauca possesses insecticidal and fungicidal properties, making it a promising candidate for natural insect control strategies.
Overall, the PCA conducted for plant extracts and bark fractions exhibited distinct clustering patterns, with variability driven by the principle components, suggesting meaningful differences in their underlying features. The present study showed that the seed kernel and bark extracts contained significant bioactive compounds, including triterpenoids (such as betulinic acid), phenolic compounds, flavonoids, and quassinoids. Preliminary dose–response analysis revealed that these extracts exhibited substantial insecticidal properties. The likely mechanisms of action include phenolic compounds inducing oxidative stress, flavonoids and triterpenoids disrupting insect nervous systems, and the diverse biological activities of quassinoids. Future research shall focus on understanding their detailed molecular mechanisms of action on insects, conducting field trials, and developing practical formulations for pest management to use S. glauca, which has the potential to be developed into a natural insecticide, providing an environmentally friendly alternative to synthetic chemicals. Furthermore, although S. glauca extracts demonstrate considerable insecticidal potential, several limitations necessitate attention. It is imperative to undertake field validation to ascertain efficacy under natural conditions, as environmental factors such as temperature, humidity, and ultraviolet exposure may affect the stability and effectiveness of bioactive compounds. Moreover, the long-term persistence and degradation rates of these extracts remain inadequately understood. This limitation may be mitigated through the optimization of formulation techniques, such as microencapsulation, to enhance stability and improve field applicability for sustainable pest management42.
Methods
Experimental approach
Insect source
The young larvae of S. frugiperda were collected from infested maize plants at the research farm of the Department of Millets, Tamil Nadu Agricultural University, Coimbatore (11.0162913° N, 76.9286577° E), brought to the insect rearing laboratory to establish the culture in controlled conditions (28 ± 1 °C, 65 ± 2% relative humidity, and a 16-h light/8-h dark photoperiod). For the first 5–6 days, approximately 100 larvae were placed in rectangular boxes (4 cm height × 15 cm width × 21 cm length) containing a thin layer (3–4 mm) of a lab-lab protein-based artificial diet (Supplementary Table 1) (200 ml), modified slightly from the method suggested by Truzi, et al.43. The late instar larvae were transferred individually into small containers (3 cm diameter, 2.3 cm depth) and maintained until they pupated. The pupae were taken out and sterilized by dipping them in a 2% sodium hypochlorite solution. Then, they were placed in a Petri plate (20 cm diameter). and kept inside adult emergence cages (45 × 45 × 45 cm). Adult moths were fed with a 10% honey solution. Adults were collected and released in 10 pairs (male and female in equal numbers) inside egg-laying cages (60 × 60 × 60 cm) containing shoots of Nerium (30–40 cm) with their cut stem parts immersed in conical flasks filled with water. Egg masses deposited on Nerium leaves were collected daily and incubated inside egg boxes (30 × 25 × 5 cm) for hatching. Newly hatched larvae were reared on a semi-synthetic artificial diet for repeating the rearing cycle. For bioassays, larvae of specific instars were selected and used from the culture44.
Plant source
The leaves, seeds, and bark of S. glauca were collected from ten randomly chosen plants in agroforestry fields at the Forest College and Research Institute, Tamil Nadu Agricultural University in Mettupalayam (11.323828863887853° N, 76.93619953654411° E), as well as in Coimbatore, Tamil Nadu, India, during 2024. Initially, the collected plant materials were decontaminated by thoroughly washing them and subsequently shade-dried at room temperature (30 ± 2 °C for 15 days until achieving a brittle texture. They were powdered using a Wiley mill (Tanco PTL-210). The powder was sieved using a mesh (Size No.40) and stored in amber-colored bottles until further use.
Preparation and extraction of botanicals
The extraction of bioactive compounds from the powdered samples of S. glauca bark, seed, and leaves by following the method suggested by Mokaizh, et al.45. Briefly, 20 g of powdered samples were placed in the extraction chamber attached to the Soxhlet apparatus (TECHNICO®, Bengaluru, India) fitted with a 500 ml round-bottom flask containing solvents (ethanol, ethyl acetate, and hexane). This extraction process was repeated continuously with each solvent for the specified duration. Afterward, the solution was allowed to cool to room temperature. The extract was filtered through Whatman No. 1 filter paper (Sigma-Aldrich, Inc., USA) and concentrated using a rotary evaporator (Heidolph®, Heidolph Instruments GmbH & Co. KG, Germany) to remove the solvents45. The concentrated extracts were stored at 4 °C for future analysis and use. The final weight of the extract was recorded to calculate the recovery percentage of the botanical extracts46.
$${text{Reovery percentage}} = { }frac{{text{Weight of the final crude}}}{{text{weight of the sample used }}} times 100$$
Dose–response study to validate the insecticidal activity of extractant against S. frugiperda
The leaf dip bioassay method, outlined by Tabashnik and Cushing47, was used to assess the insecticidal properties of crude extracted from different parts of S. glauca and the bark fractions against the third instar larvae of S. frugiperda. Five concentrations (3%, 4%, 5%, 6%, and 7%) were prepared from the stock solution using either bracketing or ad hoc methods, as described by48. Freshly collected maize leaves from glass house-grown plants, aged 35–45 days, were thoroughly washed with tap water and air-dried for 3 min at room temperature (30 ± 2 °C), while the individual fractions were taken as a treatment and replicated twice. The leaves were then carefully cut into 3 × 3 cm2 leaf discs. Each disc was immersed for 15 s in the prepared concentrations and subsequently placed on filter paper to drain any excess solution. After drying, the treated leaf discs were individually placed in each well of 12-well tissue culture plates (Sigma-Aldrich) lined at the bottom with a 2–3 mm layer of 2.0% agar to maintain freshness. In each plate, 12 pre-starved (for 2 h) third-instar S. frugiperda larvae were released. The plates were incubated in a Biological Oxygen Demand (BOD) chamber (Sanyo MLR-350H, Sanyo, Kyoto, Japan) at controlled conditions of 25 ± 2 °C, 60 ± 5% relative humidity, and a 16:8 (L:D) photoperiod49. Four replicates of 30 larvae were maintained for each treatment. Acetone-treated leaves were used as the control. Larval mortality was assessed at 24-, 48-, and 72 h after treatment (HAT), and the percentage of larval mortality was corrected using Abbott’s formula when needed50.
$${text{Percent Mortality}} = frac{{% {text{ Test mortality }} – , % {text{ Control mortality}}}}{{{1}00 , – % {text{ Control mortality}}}} times { 1}00$$
Antifeedant activity
The Antifeedant activity was assessed using a method adapted from51, with slight modifications. As explained in the previous section, maize leaves were cut into 4.5 cm discs. These discs were dipped quickly in extract solutions prepared at various concentrations (3%, 4%, 5%, 6%, and 7%) using acetone and placed on filter paper to drain excess solution, then air-dried for 3 to 5 min at room temperature. Once dry, the treated leaf discs were individually placed in 9-cm glass Petri dishes lined at the bottom with a 2-3 cm layer of 2% agar. Pre-starved (2 h) third-instar S. frugiperda larvae were individually placed on each treated leaf disc and allowed to feed for 24, 48, and 72 h. The study was conducted with a completely randomized design (CRD) with four replicates for each treatment. Each concentration had four replicates, using about 40 larvae per replicate. Acetone-treated leaf discs served as the control. At the end of the experiment, the uneaten area of the leaf discs was measured using a Portable leaf area meter (Scitek, Model LAM-PB) at 24, 48, and 72 Hours after feeding (HAF). The percent antifeedant activity was calculated using the formula below52,53.
$${text{Antifeedant Index }} = frac{{left( {C – T} right)}}{{left( {C + T} right)}} times 100$$
where C represents the leaf area consumed on control leaves, and T indicates the leaf area consumed on treated leaves by S. frugiperda.
Metabolic profiling of solvent extracts in GC–MS/MS
The crude extracts were dissolved in respective solvents and analyzed in Gas Chromatography-Tandem Mass Spectrometry (Triple Quadrupole GC–MS/MS, Shimadzu Analytical India Pvt. Ltd., New Delhi, India) to identify the compounds in the different solvent extracts. One microliter of the eluted samples was injected in splitless mode into DB-5 MS capillary GC/MS column with a 60 min total run by following the specifications: initially, the temperature was run at 40 °C for 5 min, followed by 145 °C (Temperature rise at 7 °C/min) for 15 min, 285 °C (5 °C/min) for 30 min and finally post-run for 10 min at 300 °C. The injector and column temperatures were set at 250 °C. Using an ionization voltage of 70 eV, mass spectral data were recorded using scanning mode from 50 to 800 m/z. The mass spectrometer has an integral library of compounds that automatically searches and matches the spectrum produced by the sample mass spectrums with those in the National Institute of Standards & Technology (NIST) database.
Bio-fractionation of bark extracts obtained using ethyl acetate through analytical and preparatory HPLC
Analytical/preparatory high-performance liquid chromatography (HPLC) was used to fractionate the bark crude extracted with ethyl acetate. The analytical configuration included a Security Guard cartridge (C18, 4.0 × 3.0 mm, Phenomenex, Torrance, CA) and a reverse-phase (RP) column (HyPurity®, C18, 3 µm, 150 × 4.6 mm, Thermo Scientific). The mobile phase comprised acetonitrile (solvent A) and 0.1% aqueous formic acid in HPLC water (solvent B). Gradient elution was performed at a flow rate of 1 mL/min with a detection wavelength of 270 nm and an injection volume of 100 µL. The gradient program began with 40% solvent A for 5 min, increased to 80% over 25 min, and finally reached 100% held for 3 min, reverted to 40% in 0.01 min, and was maintained at 40% for 5 min, resulting in a total run time of 38 min. For preparative HPLC, we employed an RP column (Luna®, C18, 5 µm, 250 × 1.0 mm, Phenomenex) integrated into a Shimadzu Prominence system equipped with a photodiode detector. The solvents and the gradient program were identical to the analytical setup. Based on the analytical run, ten fractions were collected at each three-minute retention time interval starting from the 3 min after running time (Supplementary Figs. 5 and 6; analytical Chromatogram). The gradient program commenced with 40% solvent A for 3 min, ramped to 80% over 30 min, held at 100% for 3 min, returned to 40% in 0.01 min, and was maintained at 40% for 3 min, resulting in a total run time of 39 min to collect. Preparative HPLC was performed with an injection volume of 500 µL and a flow rate of 15 mL/min.
Total RNA isolation, library preparation, and sequencing
The bioassay was conducted as described in the prior section. The larvae were exposed to the LC50 levels, after which they were divided into control and bark-treated groups, with ten larvae placed in separate Eppendorf tubes for each replicate. These samples were then quickly flash-frozen in liquid nitrogen. Two biological replicates were acquired for each condition. Total RNA extraction from both control and bark-treated samples was performed utilizing TRIzol reagent (Sigma-Aldrich, USA), adhering to the manufacturer’s protocol. The quality and concentration of the RNA were assessed using a Thermo Scientific NanoDrop 8000 UV–VIS Spectrophotometer (Thermo Scientific, USA). Furthermore, RNA integrity was evaluated through an Agilent 2100 Bioanalyzer (Agilent Technologies, USA) and additionally 1% agarose gel electrophoresis. Only RNA samples exhibiting an OD260/280 ratio of ≥ 1.8 and an RNA integrity number (RIN) of ≥ 7.0 were chosen for subsequent analyses. High-quality RNA (1.0 μg per sample) was utilized for the efficient preparation of the cDNA library, following the protocol outlined in the Illumina TruSeq RNA Sample Preparation Kit (Illumina©, USA). The libraries underwent amplification, and the final cDNA yield was documented at approximately 400 ng, with an average fragment size measuring 300–350 bp. The libraries prepared from all six samples were later subjected to paired-end sequencing (2 × 150 bp) utilizing the Illumina NovaSeq 6000 system platform (ONEOMICS PVT LTD, India).
Bioinformatic analysis
The quality of raw sequencing reads was evaluated using FastQC, followed by preprocessing with Trim Galore. This step involved the removal of sequences shorter than 20 bp, the trimming of low-quality bases (Q < 20), the elimination of Illumina adapters, and the filtration of ambiguous nucleotides (Ns) at the read end. A genome index was constructed using the reference genome of S. frugiperda for the initial read alignment. Read mapping was performed using STAR54, allowing a maximum of 30 multiple alignments per read pair (-outFilterMultimapNmax) while setting a mismatch-to-mapped bases ratio threshold of ≤ 0.1 (-outFilterMismatchNoverLmax). To enhance the efficiency of read mapping, unmapped reads from the initial alignment underwent further processing, which included trimming of poly-A/T tails (> 4 bases), filtering for a minimum length of 25 nucleotides, and ensuring that the GC content remained above 20% utilizing PReprocessing and INformation of SEQuence data—lite version (PRINSEQ-lite)55. The filtered reads were subsequently remapped to the genome.
Differential gene expression analysis
Gene expression levels were determined using RSEM and normalized based on fragments per kilobase of transcripts per million mapped reads (FPKM). Differentially expressed genes (DEGs) were identified with the DESeq2 (version 1.46.0) tool, setting FDR < 0.05 and log2(fold-change) (> 1.5 to − 1.5 <)56. The KEGG database (Kanehisa et al. 2004) facilitated the KEGG pathway enrichment analysis57,58,59,60, while Gene Ontology (GO)61 analysis was performed using the goseq R program. All analyses were performed in the R version (4.4.2).
Homology modeling and molecular docking
Target proteins were identified based on their metabolic functions, and their amino acid sequences were retrieved from the National Center for Biotechnology Information (NCBI) and the Protein Data Bank (PDB). This selection included glutathione S-transferase (GST), cytochrome oxidase P450 family 6 (CYP6), and five odorant-binding proteins (OBPs). These proteins play integral roles in detoxification, signal transduction, and metabolic regulation, rendering them essential targets for structure-based drug design. Initially, homology modeling was conducted utilizing MODELLER v9.1762 to generate six models for each target, which were subsequently refined through energy minimization in UCSF Chimera63. Structural validation was achieved using PROCHECK, ensuring high-quality models with appropriate stereochemical properties. Functional regions were delineated using CASTp, which predicts active site pockets and solvent accessibility, thereby facilitating the selection of key ligand-binding regions. The results obtained from the GC–MS/MS metabolomic profiling of S. glauca indicate that gamma-Sitosterol and Lanosta-8,24-dien-3-one exhibited the highest intensity and were used to assess the probability of metabolic disruption. These phytochemicals were systematically evaluated for their molecular weight, hydrogen bond donors and acceptors, as well as various drug-likeness properties in accordance with Lipinski’s Rule of Five.The selected ligands were subsequently sourced from PubChem64 and subjected to optimization using Chimera, which involved adjustments to net charge, stereochemistry, and energy minimization. These procedures were instituted to ensure an enhanced binding affinity.
Molecular docking was performed utilizing AutoDock Vina version 4.265, a tool recognized for yielding rapid and precise predictions regarding ligand–protein interactions. Grid maps were generated for each ligand–protein complex to ensure the appropriate alignment of active site residues with the corresponding ligands. The Lamarckian Genetic Algorithm (LGA) was employed to ascertain the optimal binding conformations and the stability of interactions. Binding energies were computed, and an analysis of hydrogen bonding patterns, hydrophobic interactions, and π-π stacking interactions was conducted to evaluate the efficiency of ligand binding.
Statistical analysis
The data recorded from the dose–response study was analyzed to compute the lethal concentration using Finney’s Probit approach using R software66. The significance of the means of different treatments was statistically tested following one-way ANOVA using R software, and the means were separated using the Honest Significant Difference (HSD) test. The results obtained from the GC–MS/MS were subjected to statistical analysis, including Principal compound analysis using R software (v.4.3.2) and Partial least square (PLS), sparse partial least square (sPLS), to optimize the selection of the compound based on the variable important projection (VIP) that was weighted average of loading. The variance is accounted for by the percentage of each differentiated solvent extractant used. Clustering was carried out using k-means clustering algorithms and a self-organizing map (SOM), an unsupervised neural network-based clustering algorithm. The functional enrichment and network analysis were also done by employing MetaboAnalyst-6.0 (https://www.metaboanalyst.ca/MetaboAnalyst/ModuleView.xhtml) using the KEGG database to map biological function, and subsequently, metabolites to gene interaction network were constructed to terpenoid compounds to know the genetic influences.
Conclusion
The study investigated the insecticidal properties of the crude solvent extracts of Simarouba glauca against S. frugiperda. The metabolic profiling conducted through gas chromatography-mass spectrometry (GC–MS/MS) provided significant insights into identifying and quantifying compounds present in various solvent extracts. The findings indicated that the crude extracts of S. glauca possess potential as a natural insecticide for controlling S. frugiperda. Future research and development in this area could facilitate the formulation of effective and environmentally friendly pest control solutions for agricultural applications. Researchers are examining the insecticidal benefits of S. glauca through multiple solvent extracts derived from its bark, seeds, and leaves. By standardizing extraction processes and optimizing the application of bioactive compounds from S. glauca, it is plausible to develop a sustainable and eco-friendly alternative to synthetic pesticides for pest management. Continued research and development in this domain shall promote the integration of S. glauca-based products into pest management strategies, thereby offering farmers a more sustainable approach to pest control. This study also investigated the molecular mechanisms of insecticidal activity through differential gene expression and molecular docking analyses. It identifies 734 differentially expressed genes and significant enrichment in metabolic and signaling pathways, suggesting strong transcriptional responses linked to insect mortality and feeding inhibition. Disruptions in protein digestion, ion binding, and cytoskeletal function indicate serious physiological issues that lead to paralysis and reduced feeding. Molecular docking shows that bioactive compounds like gamma-Sitosterol and Lanosta-8,24-dien-3-one strongly interact with CYP6 and odorant-binding proteins, potentially hindering detoxification and olfactory functions essential for insect survival. These results pinpoint genetic and biochemical targets for pest management, emphasizing the promise of plant-derived compounds as eco-friendly alternatives to synthetic insecticides. Future studies should validate these targets through in vivo research and improve their application in integrated pest management for better agricultural sustainability.
Data availability
The RNA-Seq data generated in this study have been deposited in the NCBI Sequence Read Archive (SRA) and are publicly available under the BioProject accession number PRJNA1242543. The BioSample accession numbers are SAMN47595159 (https://www.ncbi.nlm.nih.gov/biosample/47595159), SAMN47595160 (https://www.ncbi.nlm.nih.gov/biosample/47595160), SAMN47595161 (https://www.ncbi.nlm.nih.gov/biosample/47595161), and SAMN47595162 (https://www.ncbi.nlm.nih.gov/biosample/47595162). All other relevant data supporting the findings of this study are provided within the manuscript and supplementary materials. Any additional data from the study are available from the corresponding author upon reasonable request.
References
-
Isman, M. B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 51, 45–66 (2006).
-
Satpathi, B. Simarouba glauca An Oil-Yielding Tree (Springer, 1984).
-
Bangar, S., Dudhare, M., Deshmukh, A. & Wagh, H. Bioefficacy of crude extracts from Simarouba glauca DC. against Plutella xylostella and Helicoverpa armigera. Agric. Update 12, 436–440 (2017).
-
Bharathi, N. S., Babu, D. S., Sumathi, R. & Senthilkumar, N. Simarouba glauca DC: An effective biopesticide against leaf defoliators of Ailanthus and Tectona plants. J. Biopestic. 13, 34–39 (2020).
-
Behera, P. & Mishra, B. K. Biological control of maize stem borer, Chilo partellus (Swinhoe) in Kharif maize through combined releases of Trichogramma chilonis (Ishii) and Cotesia flavipes (Cameron) in Odisha. J. Entomol. Zool. Stud. 8, 1647–1651 (2020).
-
Stat, F. http://www.fao.org/faostat/en/#data.QC (2020, accessed 1 November 2020).
-
Erenstein, O., Jaleta, M., Sonder, K., Mottaleb, K. & Prasanna, B. M. Global maize production, consumption and trade: trends and R&D implications. Food Secur. 14, 1295–1319 (2022).
-
ICAR-Indian Institute of Maize Research. https://iimr.icar.gov.in/(2024).
-
Bennett, B. A mass mortality of fish associated with low salinity conditions in the Bot River estuary. Trans. R. Soc. S. Afr. 45, 437–447 (1985).
-
Christensen, S. A. et al. Fungal and herbivore elicitation of the novel maize sesquiterpenoid, zealexin A4, is attenuated by elevated CO₂. Planta 247, 863–873 (2018).
-
Montezano, D. G. et al. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 26, 286–300 (2018).
-
Goergen, G., Kumar, P. L., Sankung, S. B., Togola, A. & Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 11, e0165632 (2016).
-
Early, R., González-Moreno, P., Murphy, S. T. & Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. BioRxiv 2018, 391847 (2018).
-
Sharanabasappa, S., Kalleshwaraswamy, C., Maruthi, M. & Pavithra, H. Biology of invasive fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) on maize. Indian J. Entomol. 80, 540–543 (2018).
-
Ganiger, P. et al. Occurrence of the new invasive pest, fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), in the maize fields of Karnataka, India. Curr. Sci. 115, 621–623 (2018).
-
PlantwisePlus Annual Report 2021—CABI.org. CABI (2023).
-
Mendesil, E. et al. The invasive fall armyworm, Spodoptera frugiperda, in Africa and Asia: responding to the food security challenge, with priorities for integrated pest management research. J. Plant Dis. Prot. 130, 1175–1206 (2023).
-
Chisonga, C., Chipabika, G., Sohati, P. H. & Harrison, R. D. Understanding the impact of fall armyworm (Spodoptera frugiperda J.E. Smith) leaf damage on maize yields. PLoS ONE 18, e0279138 (2023).
-
De Groote, H. et al. Spread and impact of fall armyworm (Spodoptera frugiperda J.E. Smith) in maize production areas of Kenya. Agric. Ecosyst. Environ. 292, 106804 (2020).
-
Abro, Z. et al. Socioeconomic and health impacts of fall armyworm in Ethiopia. PLoS ONE 16, e0257736 (2021).
-
Makgoba, M. C., Tshikhudo, P. P., Nnzeru, L. R. & Makhado, R. A. Impact of fall armyworm (Spodoptera frugiperda) (J.E. Smith) on small-scale maize farmers and its control strategies in the Limpopo province, South Africa. Jàmbá J. Disaster Risk Stud. 13, 5639 (2021).
-
Kansiime, M. K., Rwomushana, I. & Mugambi, I. Fall armyworm invasion in Sub-Saharan Africa and impacts on community sustainability in the wake of Coronavirus Disease 2019: reviewing the evidence. Curr. Opin. Environ. Sustain. 62, 101279 (2023).
-
Deshmukh, S. et al. Field efficacy of insecticides for management of invasive fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) on maize in India. Fla. Entomol. 103, 221–227 (2020).
-
Antony, J., Thomas, A. & Gnanasekaran, D. Review study on pharmacological importance of Simarouba glauca. Int. J. New Technol. Res. 2, 263412 (2016).
-
Jose, A., Kannan, E., Kumar, P. R. A. V. & Madhunapantula, S. V. Therapeutic potential of phytochemicals isolated from Simarouba glauca for inhibiting cancers: a review. Syst. Rev. Pharm. 10, 73–80 (2019).
-
Ashida, S., Beevi, N., Raj, R. S. & Prabhakumari, C. Qualitative detection of triterpenes and quantification of betulinic acid from hexane extract of Simarouba glauca leaves by gas chromatography-mass spectrometry and high-performance thin layer chromatography. Indian J. Pharm. Sci. 84, 6 (2022).
-
Osagie-Eweka, S. Phytochemical analyses and comparative in vitro antioxidant studies of aqueous, methanol and ethanol stem bark extracts of Simarouba glauca DC (Paradise tree). Afr. J. Plant Sci. 12, 7–16 (2018).
-
Jamil, M., Mirza, B., Yasmeen, A. & Khan, M. A. Pharmacological activities of selected plant species and their phytochemical analysis. J. Med. Plants Res. 6, 5013–5022 (2012).
-
Alves, I. A., Miranda, H. M., Soares, L. A. & Randau, K. P. Simaroubaceae family: botany, chemical composition and biological activities. Rev. Bras. Farmacogn. 24, 481–501 (2014).
-
Jose, A., Kannan, E. & Madhunapantula, S. V. Anti-proliferative potential of phytochemical fractions isolated from Simarouba glauca DC leaf. Heliyon 6, 4 (2020).
-
Umesh, T. In-vitro antioxidant potential, free radical scavenging and cytotoxic activity of Simarouba glauca leaves. Int. J. Pharm. Pharm. Sci. 7, 411–416 (2015).
-
Anantharaju, P. G., Gowda, P. C., Vimalambike, M. G. & Madhunapantula, S. V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 15, 1–16 (2016).
-
Katdare, M., Osborne, M. P. & Telang, N. T. Inhibition of aberrant proliferation and induction of apoptosis in pre-neoplastic human mammary epithelial cells by natural phytochemicals. Oncol. Rep. 5, 311–316 (1998).
-
Surh, Y.-J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 3, 768–780 (2003).
-
Bursch, W., Ellinger, A., Gerner, C. & Schulte-Hermann, R. Caspase-independent and autophagic programmed cell death. In When Cells Die. II. A Comprehensive Evaluation of Apoptosis and Programmed Cell Death, 2nd edn 275–310 (Wiley, 2004).
-
Priyadharshini, B. & Prakash, M. Pharmacoinformatics and molecular docking studies on Psoralea corylifolia (L.) derived seed compounds against tumor-induced cancer protein. Int. Res. J. Pharm. 15, 23–19 (2024).
-
Abdul-Rahuman, A., Gopalakrishnan, G., Venkatesan, P. & Geetha, K. Isolation and identification of mosquito larvicidal compound from Abutilon indicum (Linn.) Sweet. Parasitol. Res. 102, 981–988. https://doi.org/10.1007/s00436-007-0864-5 (2008).
-
Sirikhansaeng, P., Tanee, T., Sudmoon, R. & Chaveerach, A. Major phytochemical as γ-sitosterol disclosing and toxicity testing in Lagerstroemia species. Evid. Based Complement. Alternat. Med. 2017, 7209851. https://doi.org/10.1155/2017/7209851 (2017).
-
Ade-Souza, S. A. et al. An interesting relationship between the insecticidal potential of Simarouba sp. in the biology of diamondback moth. Sustainability 15, 7759 (2023).
-
Koul, O. The Handbook of Naturally Occurring Insecticidal Toxins (Springer, 2016).
-
Jayashanthini, S. & Murugan, K. Phytochemical screening of Simarouba amara seed oil: A potent mosquitocidal effect against malarial vector, Anopheles stephensi (2024).
-
Machado, S., Pereira, R. & Sousa, R. M. O. Nanobiopesticides: Are they the future of phytosanitary treatments in modern agriculture?. Sci. Total Environ. 896, 166401 (2023).
-
Truzi, C. C., Vieira, N. F., De Souza, J. M. & De Bortoli, S. A. Artificial diets with different protein levels for rearing Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Insect Sci. 21, 2 (2021).
-
Kumar, S., Chauhan, M., Singh, N. & Saxena, M. K. Efficacy of different bioassays for the assessment of toxicity of plant extracts. Indian J. Ecol. 47, 485–492 (2020).
-
Mokaizh, A. A. B., Nour, A. H. & Yunus, R. M. Extraction and characterization of phenolic compounds from Commiphora gileadensis bark using ultrasonic-assisted extraction. Pharmacol. Res. Nat. Prod. 4, 100066 (2024).
-
Larkem, I. et al. Chemical profile and antioxidant activity of Sesbania bispinosa (Jacq.) W. Wight aerial parts and seeds extracts. J. Food Process. Preserv. 45, e15468 (2021).
-
Tabashnik, B. & Cushing, N. Leaf residue vs. topical bioassays for assessing insecticide resistance in the diamond-back moth, Plutella xylostella L. (1987).
-
Reda, M., Mashtoly, T. A., El-Zemaity, M.E.-S., Abolmaaty, A. & Abdelatef, G. M. Phylogenic and antagonistic characteristics of novel Bacillus cereus isolates against desert locust, Schistocerca gregaria Forskal (Orthoptera: Acrididae). Egypt. J. Biol. Pest Control 28, 1–7 (2018).
-
Ahmed, A. M. et al. Development of protein-based artificial diet for mass rearing of Spodoptera frugiperda J.E. Smith (Lepidoptera: Noctuidae). Pak. J. Agric. Res. 36, 4 (2023).
-
Abbott, W. S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18, 265–267 (1925).
-
Rahman, A. et al. Larvicidal and antifeedant effects of copper nano-pesticides against Spodoptera frugiperda (J.E. Smith) and its immunological response. Insects 13, 1030 (2022).
-
Belles, X., Camps, F., Coll, J. & Piulachs, M. D. Insect antifeedant activity of clerodane diterpenoids against larvae of Spodoptera littoralis (Boisd.) (Lepidoptera). J. Chem. Ecol. 11, 1439–1445 (1985).
-
Sreelatha, T. et al. A new benzil derivative from Derris scandens: structure-insecticidal activity study. Bioorg. Med. Chem. Lett. 20, 549–553 (2010).
-
Dobin, A. & Gingeras, T. R. Mapping RNA-seq reads with STAR. CPBI 51(1), 11–14 (2015).
-
Cantu, V.A., Sadural, J. & Edwards, R. PRINSEQ++, a multi-threaded tool for fast and efficient quality control and preprocessing of sequencing datasets (2016).
-
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 1–21 (2014).
-
Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Prot. Sci. 28, 1947–1951 (2019).
-
Kanehisa, M. Post-Genome Informatics (OUP Oxford, 2000).
-
Kanehisa, M., Furumichi, M., Sato, Y., Kawashima, M. & Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 51, D587–D592 (2023).
-
Kanehisa, M. & Goto, S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30 (2000).
-
Ashburner, M. et al. Gene ontology: tool for the unification of biology. Nat. Genet. 25, 25–29 (2000).
-
Webb, B. & Sali, A. Comparative protein structure modeling using MODELLER. Curr. Protoc. Bioinform. 54, 5–37 (2016).
-
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
-
Kim, S. et al. PubChem substance and compound databases. Nucleic Acids Res. 44, D1202–D1213 (2016).
-
Trott, O. & Olson, A. J. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 31, 455–461 (2010).
-
Trott, O. Probit analysis. J. Pharm. Sci. 60, 1432. https://doi.org/10.1002/jps.2600600940 (1971).
Acknowledgements
The author sincerely thanks the Department of Agricultural Entomology and Centre for Plant Molecular Biology and Bioinformatics for providing instrumentation facilities.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
G, K., Mookiah, S., Marimuthu, M. et al. Untargeted metabolomics-based study of extracts from Simarouba glauca plant parts for insecticidal effects against Spodoptera frugiperda. Sci Rep 15, 30643 (2025). https://doi.org/10.1038/s41598-025-01605-y
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-01605-y