Introduction
Based on area and production worldwide, lentil (Lens culinaris Medikus Sub spp. culinaris) ranks as the sixth food legume. Due to the large amounts of protein, fibre, minerals, and micronutrients that its seed contains, it is a highly popular food legume in Bangladesh and throughout the world1,2. While more than 70 countries cultivate lentils, the Indian subcontinent, Africa, America, and Australia represent almost 70% of the world’s production3,4. According to FAOSTAT (2020)5 the top ten countries in the world that produce lentils are Canada, India, Australia, Turkey, Nepal, Bangladesh, Russia, China, the USA, and Ethiopia. 6.33 million tons of lentils are produced annually on 6.10 million hectares of land worldwide, with a productivity of 1037 kg/ha5. Abiotic and biotic stresses are the major constraints for reducing the potential yield of lentil. The most common and deadly biotic stress on lentils is the foliar fungal disease Stemphylium blight, which is brought on by the necrotrophic ascomycete Stemphylium botryosum Wallr and has been documented in practically all lentil-growing countries, including Bangladesh. Since the initial epidemic occurred in Bangladesh in 1986, the disease is widespread in countries that grow lentils, including India, Nepal, Canada, Syria, and the USA6. Moreover, significant yield loss was increased in lentil production in South Asia and North America7 and 22% yield reduction was recorded due to Stemphylium blight disease globally8. The overall, economic loss was recorded up to 80% by severe disease infection in susceptible varieties of lentil in Bangladesh9. It is the major threat for lentil production and is widespread the whole country with the maximum in Jashore, Pabna, Kushtia, Faridpur, Madaripur and Dhaka10. Very recent, Stemphylium blight is also a constraint for lentil cultivation in Barisal and Bhola districts, the southern parts of Bangladesh11. It also causes serious yield losses in the major lentil growing countries, like in India disease severity was observed up to 83% and yield loss was recorded 93%12, in Nepal caused 100% yield loss due to serious infestation in susceptible variety of lentil13. Recently, Stemphylium blight infection is raising in lentils fields of Saskatchewan14. However, severe disease infection in the susceptible lentil genotypes under favorable weather conditions, yield loss might be happened up to 100% in the others lentil producing countries of the world15.
Yield loss due to Stemphylium blight might be minimized in two ways; cultivation of resistant cultivars and application of fungicides6. Among these remedies’ cultivation of resistant cultivars or varieties is the most durable and ecofriendly approaches for minimization of yield loss in lentil. For instances, breeding attempts have to be focused on the development of varieties or identification of cultivars from existing germplasm with resistance to this biotic stress. Moreover, with narrow genetic variability in legume crop is a keen risk for crop vulnerability to biotic and abiotic constraints, a major concern shared among the plant breeders and growers of legume crops alike16. Hence, screening is the first and only one sustainable method to identify resistant gene from the existing sources of germplasm. In addition, exploring the sources of resistance from the available cultivated gene pool is often a first step before exploring secondary or tertiary gene pool. Although a limited report of resistance against Stemphylium blight in lentil are available which are focused on the screening of cultivated germplasm from several parts of the world17. Considering the above-mentioned facts, the screening of lentil germplasm was under taken as a time demanding task for development of Stemphyllium blight resistance lentil variety which will increase the lentil production in Bangladesh as well as globally.
Results
Disease reaction of Stemphylium blight of lentil genotypes
Sixty lentil genotypes were screened against Stemphylium blight disease under natural epiphytotic condition for two consecutive growing seasons, 2019-20 and 2020-21, respectively. Significant disease symptoms and condia of Stemphylium botryosum were detected from the experimental field during both growing seasons 2019-20 and 2020-21 (Fig. 1a,b). Disease severity was recorded in a 1 to 9 disease rating scale at three different growing stages such as flowering, pod filling and maturity stages. The disease rating scale of the infected genotypes is given in the Fig. 2.

(a) Symptoms of Stemphylium blight on lentil shown by arrow heads on leaflets at experimental field at PRC, BARI, Ishurdi, Pabna, Bangladesh within 89.050E longitudes and 24.030N attitudes during flowering stage; (b) Microphotograph of conidia of Stemphylium botryosum isolated from infected lentil leaves under compound (E-mail: akzaman118@yahoo.com).

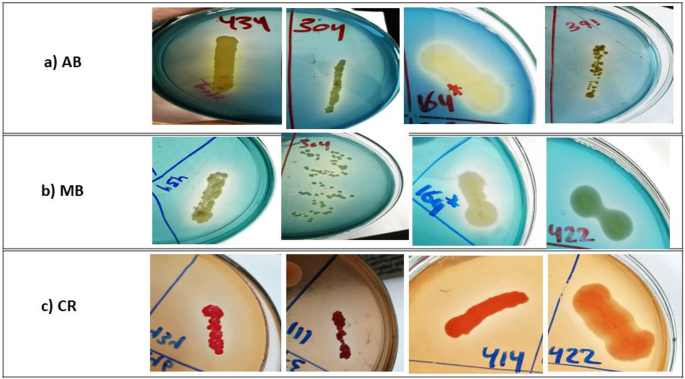
Infected genotypes by Stemphylium blight pathogen showing disease rating scale (“a” to “i” indicated the disease rating scale 1 to 9 respectively).
The disease symptom of the genotypes was detected around 24 January in each growing season and the disease severity progress was taken every 7 days interval. The average disease progress severity is shown in the Appendix I, Appendix II, and Appendix III. Significant difference to the reaction of Stemphylium botryosum was observed at different growth stages of 60 lentil genotypes.
At flowering stage
At flowering stage, from starting of infection around 24 January disease severity was scored three times at seven days interval. Based on the last disease severity scale (among three time scaling at flowering stage) the frequency distribution of Stemphylium blight disease score is shown in the Figs. 3 and 7 for each year respectively. From the first trial, only 3 genotypes viz.; G7 (BARI Masur-9), G16 (PRECOZ), and G40 (BLX-12004-5) were found to be resistant to the disease at flowering stage. 28 genotypes found to be Moderately Resistant-Moderately Susceptible (MR-MS) disease reaction, 10 genotypes Resistant-Moderately Resistant (R-MR), 14 genotypes Moderately Resistant (MR) (Fig. 3).

Frequency distribution of Stemphylium blight disease rating score in 60 lentil genotypes at flowering stage under natural epiphytotic condition during 2019-20.
In the second trial, 37 genotypes showed Moderately Resistant disease reaction, 17 genotypes showed Moderately Resistant-Moderately Susceptible disease reaction. Four genotypes G7 (BARI Masur-7), G8 (LRIL-21-67-1-1-1-1), G20 (BLX-01013-21-5), and G52 (BLX 10001-1) showed Resistant-Moderately Resistant disease reaction, and the genotypes G24 (BLX-0200-08-4), and G26 (BLX-04004-10) were found as Moderately Susceptible against Stemphylium blight disease (Fig. 4).

Frequency distribution of Stemphylium blight disease severity rating scale on 60 lentil genotypes at flowering stage under natural epiphytotic condition during 2020-21.
After inoculation at pre-flowering stages
Twenty-seven genotypes showed Moderately Resistant-Moderately Susceptible disease reaction and 22 genotypes as Moderately Susceptible (Fig. 5). No immune or resistant source was found among the tested 60 genotypes after inoculation of Stemphylium bottyosum inoculum in the field at pre-flowering stage. Eight genotypes viz.; G7 (BARI Masur-9), G15 (LR 9–25), G16 (PRECOZ), G40 (BLX-12004-5), G48 (RL-12-181), G58 (BLX-05002-3), G59 (BLX 09015), and G60 (BLX 04005-9 were identified as MR against Stemphylium blight disease of lentil. Three genotypes viz.; G24 (BLX-0200-08-4), G53 (BLX 10002-15), and BLX 10002-20 were found as Moderately Susceptible-Susceptible.

Frequency distribution of Stemphylium blight disease rating scale on 60 lentil genotypes at flowering stage under artificial inoculation during 2021-22 at PRC, BARI, Ishurdi, Pabna.
At pod filling stage
In case of first trial, 19 genotypes were found to be MS-S disease reaction, 15 genotypes as Moderately Susceptible. Eight genotypes were identified as MR, 7 were MR-MS, and rest of 7 genotypes were with S disease reaction at the pod filling stage during 2019-20 (Fig. 6). In the second-year trial, 19 genotypes were S, 18 were identified as MS-S, and 14 genotypes were identified as MS. Only one genotype G7 (BARI Masur-9) was MR and three genotypes were found as S-VS at Podding filling stage during 2020-21 under natural epiphytotic condition (Fig. 7).

Frequency distribution 60 lentil genotypes against Stemphylium blight disease rating scale at Pod filling stage under natural epiphytotic condition during 2019-20.

Frequency distribution of 60 lentil genotypes against Stemphylium blight disease ratingscale at Pod filling stage under natural epiphytotic condition during 2020-21.
Artificial inoculation at podding stage
In case of artificial inoculation, maximum 31 genotypes were identified as S, 18 genotypes as S-VS (Fig. 8), One genotype G16 (PRECOZ) was identified as MR-MS, 5 genotypes were MS and 5 showed as MS-S among the 60 lentil genotypes at podding stage in this study.

Frequency distribution of 60 lentil genotypes based on Stemphylium blight disease rating scale at Pod filling stage under artificial inoculation during 2021-22.
Area under disease progress curve (AUDPC)
Under natural epiphytotic condition
Significant variation was observed among the genotypes based on the estimated mean AUDPC values and the maximum AUDPC was found in case of the genotype G44 (Bagura local) followed by G30 (BLX-05004-8), G39 (BLX 01013), G10 (IRLL-22-68-1-1-1-1-0), G41(BLX-12009-6), G12 (ILL-5143), and G13 (91517) (Fig. 9). The minimum AUDPC was found in case of the genotype G16(PRECOZ) followed by the genotypes G7 (BARI Masur-9), G59 (BLX 09015), G48 (RL-12-181), G15 (LR-9-26), G40 (BLX-12004-5), and G9 (LRIL-21-112-1-1-1-1-6) during 2019-20 crop season. During 2020-21 maximum AUDPC (256.67) was recorded in case genotype 24 (BLX-0200-08-4), followed by G13(91517), G1(BARI Masur-1), G44 (Bagura local), G26 (BLX-04004-10), and G53 (BLX 10002-15) (Fig. 10). The minimum AUDPC (94.50) was found in case of genotypes G7 (BARI Masur-9) followed by G9 (LRIL-21-112-1-1-1-1-6), G16 (PRECOZ), G59 (BLX 09015), G53 (BLX 10002-15), G48 (RL-12-181) genotypes.

Estimated AUDPC of 60 lentil genotypes under natural epiphytotic condition during 2019-20 at PRC, BARI, Ishurdi, Pabna. Each column represents mean with standard error of each genotype; n = 3.

Estimated mean AUDPC of 60 lentil genotypes under natural epiphytotic condition during 2020-21 at PRC, BARI, Ishurdi, Pabna. Each column represents mean with standard error of each genotype; n = 3.
Under artificial inoculation
The maximum estimated AUDPC (241.33) was found in case of genotype G24 (BLX-0200-08-4) followed by the genotypes G44 (Bagura local) (220.40), G1 (BARI Masur-3) (219.33), G26 (BLX 04004-10) (215.83), and G39 (BLX 01013) (212.33) (Fig. 11) after applied artificial inoculation of the Stemphylium botryosum at pre-flowering and flowering stage of lentil. The minimum AUDPC 103.33 was recorded in genotype G28 (BLX-05001-10) followed by the genotypes G52 (BLX 10001-1) (110.83), G40 (BLX-12004-5) (110.83), and G7 (BARI Masur-9) (115.50).

Estimated mean AUDPC of 60 lentil genotypes under artificial inoculum condition during 2021-22 at PRC, BARI, Ishurdi, Pabna. Each column represents mean with standard error of each genotype; n = 3.
Overall frequency distribution of Stemphylium blight disease reaction of 60 lentil genotypes
Under natural epiphytotic condition
Based on the last two consecutive years disease severity and estimated average AUDPC of 60 lentil genotypes thirty genotypes {G2 (BARI Masur-4), G3 (BARI Masur-5), G5 (BARI Masur-7), G6 (BARI Masur-8), G10 (IRLL-22-68-1-1-1-1-0), G11 (ILL-8008), G12 (ILL-5143), G17 (Flip-2009–52 L), G19 (BLX-99033-19), G20 (BLX-01013-21-5), G21 (BLX-01014-9), G22 (BLX-02009-11-1), G23 (BLX-02009-17-1), G25 (BLX-0200-18-3), G26 (BLX-04004-10), G27 (BLX-04010-15), G31 (BLX-05005-3), G32 (BLX-05006-7), G33 (BLX-05007-5), G34 (BLX-05008-3),, G35 (BLX-05008-11), G36 (BLX-05010-3), G38 (BLX-05010-6), G41 (BLX-12009-6), G42 (BLX-12011-6), G47 (Maitree), G49 (BLX 11014-8), G50 (BLX 11014-10), G58 (BLX-05002-3), G60 (BLX 04005-9)} were identified as being susceptible to the disease with disease rating scale 7, 61–80% infection rate, and average estimated AUDPC 161–200 (Table 1). A total of 8 genotypes {G1 (BARI Masur-3), G4 (BARI Masur-6), G13 (91517), G24 (BLX-0200-08-4), G30 (BLX-05004-8), G39 (BLX 01013), G44 (Bagura local), G55 (LRIL-18-102)} were identified as susceptible-very susceptible with disease rating scale 8, 81–90% infection rate, and average estimated AUDPC 201–240. Only 6 genotypes; G8 (LRIL-21-112-1-1-1-1), G9 (LRIL-21-112-1-1-1-1-6), G28 (BLX-05001-10), G45 (RL-12-178), G5 1(BLX 11014-11), and G57 (LG-198) were identified as moderately susceptible genotypes with disease severity scale 3, 31 to 40% disease infection rate, and average estimated AUPC 121–140. Nine genotypes viz.; G14 (9945), G18 (BLX-98001-9), G29 (BLX 05002-6), G27 (BLX-04010-15), G43 (L 4717), G46 (RL-12-171), G53 (BLX 10002-15), G54 (BLX 10002-20), and G56 (LRIL-22-158) were determined as moderately susceptible with disease rating scale 5, 31 to 45% infection rate, and average estimated AUDPC 121–140 during the consecutive two years (2019-20, 2020-21). The genotypes G7 (BARI Masur-9), G15 (LR-9-26), G16 (PRECOZ), G40 (BLX-12004-5), G48 (RL-12-181), G52 (BLX 10001-1), and G59 (BLX 09015) were identified as moderately resistance-moderately susceptible lentil genotypes against Stemphylium blight disease with disease rating scale 4, 16 to 30% disease infection rate, and average estimated AUDPC 81–120. There was no genotype identified as resistant, resistant to moderately resistant, moderately resistant, and very susceptible genotypes among the 60 lentils under the under natural epiphytotic condition grown at experimental station of PRC, BARI, Ishurdi, Pabna.
Under artificial inoculation in field condition
Artificially developed inoculum was sprayed at two times viz.; first time at pre-flowering stage on 35 DAS and second time at 50 DAS. Then disease rating was recorded just after 13 days of the inoculum spray day (Table 2). Only three genotypes such as G7 (BARI Masur-9), G16 (PRECOZ), and G59 (BLX 09015) were identified as moderately susceptible genotypes among the studied genotypes. Whereas only one genotype G15(LR-9-26) was identified as moderately susceptible to susceptible genotype, 13 genotypes were found as susceptible, and 36 genotypes were identified as susceptible to very susceptible disease reaction with the average estimated AUDPC 201–240. The seven genotypes; G22 (BLX-02009-11-1), G23 (BLX-02009-17-1), G24 (BLX-0200-08-4), G30 (BLX-05004-8), G39 (BLX 01013), G54 (BLX 10002-20), and G55 (LRIL-18-102) were identified as very susceptible genotypes with the average estimated AUDPC more than 240.
Yield performance of 60 lentil genotypes under Stemphylium blight disease condition
Yield and yield contributing traits of 60 lentil genotypes under natural epiphytotic condition
Significant variations were observed among 60 lentil genotypes based on yield and all yield contributing traits except branches plant− 1 and pods plant− 1 under natural epiphytotic condition during the crop season 2019-20, and 2020-21(Table 3). The days to flowering and days to maturity ranged from 51 to 74 days, and 88–105 days, respectively. Genotypes G50 flowered early (51 days), and genotype G58 matured early (88 days). The filled grain plant− 1(FGP) ranged from 27 to 50, and the maximum FGP (50) was recorded from the genotype G9 (LRL-21-112-1-1-1-1-6), G7 (BARI Masur-7) followed by G12 (ILL-5143). The range of grain yield plant− 1 was recorded from 0.32 to 0.75 g plant− 1. The range of grain yield ha− 1 was recorded 529.15 kg to 1182.82 kg, with the maximum grain 1182.82 kg ha− 1 from genotype G36 (BLX-05010-3) followed by genotypes G32 (BLX-05006-7), and G54 (BLX 10002-20) with grain yield 1022.78 kg ha− 1, and 1018.37 kg ha− 1 respectively.
Yield and yield contributing traits of 60 lentil genotypes under artificial inoculation in field
Significant variation was observed among the studied genotypes in case of yield and all yield contributing traits except PH, BPP, FGP, and UFGP (Table 4). The days to flowering ranged from 51 to 74 days, and days to maturity range was 88–105 days. The genotype G54 (BLX 10002-20) flowered early but genotype G17 (Flip-2009–52 L) matured early. The minimum 100swt was 1.41 g recorded in the genotype G22 (BLX-02009-11-1) and the maximum 100swt 2.85 g was obtained from the genotype G16 (PRECOZ). The maximum GYP was 0.75 g from the genotype G41 (BLX-05010-3) with maximum yield performance 1182.82 kg ha− 1 followed by the genotypes G56 (LRIL-22-158) (0.73 g), and G16 (PRECOZ) (0.72 g). The minimum yield 529.15 kg ha− 1 was obtained from genotype G27 (BLX-04010-15) with 0.36 g grain yield plant− 1.
Molecular characterization of the causal organism of Stemphylium botryosum
PCR amplification of ITS region of Stemplylium botryosun isolates
PCR amplification of ITS region of Stemphylium botryosum isolates from lentil in Bangladesh is first initiative. The amplicon size ~ 550 bp of the ITS region were amplified by using ITS4 and ITS5 primers which include partial sequence of 18 S ribosomal RNA gene, ITS-1, 5. 8 S ribosomal RNA gene, ITS2 complete sequence and partial sequence of 28 S ribosomal RNA sequence (Fig. 12).

Agarose gel electrophoresis showing the amplification of glyceraldehyde-3-phosphate dehydrogenase (gdp) and internal transcribed spacer (ITS) region. Lane M = DNA Marker and lane 1–3 fungal genomic DNA of isolate SBI1; SBI2 & SBI3 of gdp2 and 4–6 lane are ITS region of Stemphylium botryosum.
Phylogenetic placement of Stemphylium botryosum isolates from Lentil in Bangladesh
A total of 03 isolates ITS region and gdp2 were sequenced and all sequences showed 96–100% similarity with sequences of Stemphylium botryosum from GenBank by nBLAST search analysis. Phylogenetic tree was constructed using MEGA V10 where Alternaria alternata is an out group. For construction of a phylogenetic tree, sequences of Stemphylium botryosum and related species of Stemphylium were collected retrieving from GenBank. From the multi-locus sequencing of Stemphylium botryosum from Bangladesh clustered with the Stemphylium botryosum isolates from other part of the world (Fig. 13). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches18. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree (Fig. 13). The evolutionary distances were computed using the Tajima-Nei method19 and are in the units of the number of base substitutions per site. The analysis involved 27 nucleotide sequences. All positions containing gaps and missing data were eliminated. Evolutionary analyses were ducted in MEGA1020. The description of the sequences used in this study are presented in the Table 5.

Phylogenetic tree constructed by Neighbor-Joining method from concatenated rDNA-ITS/gdp sequences using MEGA 11which developed by Tamura et al. (2021)54. * The partial sequencing data of the three isolate SBI1, SBI2, and SBI3 were available in the “National Center for Biotechnology Information (NCBI) Archive with the primary accession code “OQ442935.1; OQ442936.1; and OQ442937.1” respectively. The web link of these accession numbers are available in the following data base weblink system- https://www.ncbi.nlm.nih.gov/nuccore/OQ442935.1; https://www.ncbi.nlm.nih.gov/nuccore/OQ442936.1; https://www.ncbi.nlm.nih.gov/nuccore/OQ442937.1.
Discussion
Disease reaction of Stemphylium blight
This is the first attempt at the “Pulses Research Centre” to thoroughly screen lentil germplasm of resistance source against Stemphylium blight. In field screening program, the disease pressure was created in several ways: (i) seeds were sown one month late compared to the optimum seeding date, (ii) each tested genotype was sown in the middle two rows surrounding the both rows with Stemphylium blight susceptible variety BARI Masur-1, (iii) the previous year infected plant debris was sprayed as source of inoculation to the middle two rows of the each tested genotype for enhancing the disease infection, and (iv) mist irrigation was sprayed during the pre-flowering stage.
Three growing seasons 2019-20, 2020-21 and 2021-22 were more conducive for disease development in natural epiphytotic condition as well as artificial inoculum condition. However, it was field screening program under natural epiphytotic condition, that’s why, scrapping the infected leaves were observed under compound microscope for confirmation of the pathogen inoculumbefore rating the disease severity during the both growing seasons. Significant considerable variations were observed in resistance/susceptibility to Stemphylium blight among the studied 60 genotypes. Screening of lentil germplasm under field condition has been reported by many researchers globally11,20,21,22,23,24,25.
In this screening program disease severity rating scores were taken at three different growth stages like flowering, pod filling and maturity stages during the growing season 2019-20, 2020-21, and 2021-22 and the disease severity rating scale was recorded starting from the first severity stage at 48 days after sowing to till up to maturity stage within 7 days intervals. Area Under Disease Progress Cure also was estimated for identifying the resistant source among the tested genotypes against the Stemphylium blight disease of lentil.
At flowering stage
During 2019-20 crop season maximum 28 genotypes were identified as MR-MS followed by MR (14) and only three genotype Precoz, BARI Masur-9, and G59 (BMX 09005) was identified as R, and 10 genotypes were identified as R-MR. In case of second year trial, maximum 37 genotypes were identified as MR, followed by MR-MS (17 genotypes), and four genotypes were identified as R-MR source against Stemphylium blight disease. Considering two years disease severity at flowering stage, it was observed that disease infestation was more severe in the year 2020-21 than 2019-20. But disease severity under artificial inoculation in 2021-22 indicated that maximum genotypes showing as MR-MS disease reaction. So, based on disease severity under natural epiphytotic and artificial inoculation, many genotypes were identified as MR-MS which indicates that the genotypes might have more genetic potential at their early growth stages to overcome the Stemphylium blight disease infestation. However, some genotypes viz.; G7 (BARI Masur-9), G16 (PRECOZ), G15 (LR-9-26), G40 (BLX-12004-5), G48 (RL-12-181), G58 (BLX 05002-3), G59 (BLX 09015), and G60 (BLX 04005-9) showed as Moderately Resistant capacity against the Stemphylim blight disease up to their flowering stage in spite of artificial inoculation of those genotypes just before their pre-flowering stage. Similar study was conducted by Alam et al. (2019)26 and Razzak et al. (2018)20 under natural epiphytotic condition and they found several genotypes to be MR against the Stemphylium blight disease at flowering stage.
At pod filling stage
At pod filling stage, numerous genotypes were identified as MS-S against Stemphylium blight disease two years disease severity which indicated that the disease severity increased gradually and with the increase in temperature and relative humidity of the field condition. It also found that genetic resistance of the genotypes decreased after flowering stage and at the later stages of respective genotypes. But in case of artificial inoculation, quite a number of genotypes were identified as susceptible, followed by susceptible-very susceptible to Stemphylium blight disease at their pod filling stage which indicates that, application of artificial inoculum on the tested genotypes enhanced the disease severity.
Overall frequency distribution of Stemphylium blight disease reaction of 60 lentil genotypes
The disease severity results under the natural epiphytotic condition and artificial inoculation clearly indicated that no resistant or moderately resistant sources were available in these 60 lentil genotypes. Some genotypes viz.; G7 (BARI Masur-9), G16 (Precoz), G59 (BLX 09015), G15 (LR-9-26), G40 (BLX-12004-5), and G52 (BLX 10001-1), were identified as moderately resistance-moderately susceptible genotypes against the Stemphylium blight disease of lentil up to a certain growth stage like pod filling stage.
The present research findings also indicated that the studied genotypes have been exhibited a broad variation in reaction to Stemphylium blight disease during their different growth stages under field condition. The resistance capacity of the genotypes against Stemphylium blight was identified flowering stage > pod filing stage ˃maturity stage. We also found that the disease severity increased with age of plants and daily increase in temperature and humidity of the growing environments. Similar findings were reported by Alam et al. (2019)26and Razzak et al. (2018)20.
Six genotypes were identified as moderately resistant to moderately susceptible (MR-MS); two genotypes BARI Masur-9, LRIL-21-112-1-1-1-1-6 were originated from the ICARDA materials. ICARDA has significant impact on lentil breeding program in Bangladesh, and every year providing a large number of accessions with improved Stemphylium blight disease resistance. PRECOZ is the popular Argentine lentil variety, and at early stage it showed resistance to Stemphylium blight disease25. But the present study indicated that PRECOZ showed as MR to MS against Stemphylium blight which indicated that the resistance capacity of the variety has deteriorated at later growth stage. Other three genotypes BLX-12004-5, BLX 10001-1, BLX 09015 are the breeding lines of PRC, and the reason of their resistance capacity against Stemphylium blight is not known. Most of the breeding lines of PRC are susceptible against Stemphylium blight disease and the reasons of susceptibility of these breeding lines is unknown and further studies are required to confirm their pathogenic sensitivity. No resistant lines or varieties were identified from the present study against Stemphylium blight disease. Similar findings were also reported by many researchers globally17,27,28,29.
Exhibition of several reaction of the tested genotypes against Stemphylium blight under natural epidemic condition indicated different reasons behind the variation among the tested genotypes at their three different growing stages. The first and major reasons might be the genetic mechanisms or potentiality of the genotypes or varieties which could play the resistance mechanism of the plant against Stemphylium blight disease. Another reason might be the pathogenic strain/races alternation of S. botryosum, and that’s why some genotypes might have shown resistant mechanism against respective pathogen. Crops cellular structure also depends like a thicker cuticle, thickened epidermal cell layer, fewer stomata and a large number of epidermal hairs have been reported as the resistance mechanism compared to the susceptible genotypes30. Similar findings were reported by Razzak et al. (2018)20, Bakr et al. (1994)31 and Mia et al. (1986)32. From this study it was also clearly proved that environmental factors like temperature, relative humidity (RH), and leaf wetness are the major influencing parameters for the development of Stemphylium blight disease. Based on different surveys and conducted experimental at Bihar, India and Bangladesh by Suheri and Price (2000)33 Basallote-Ureba et al. (1999)34, and Sinha et al. (1993)12 confirmed that a wide range of environmental conditions like temperature and RH are the major factors for development of S. botryosum in lentil for causing the blight disease. The average temperature 18 ± 2 °C, 85–90% RH in the morning, RH > 50% at afternoon, and average 7.7 h of sunshine with number of cloudy and foggy days were most essential factors for causing, appearance, development, and spread of Stemphylium blight disease in lentil.
Yield and yield contributing traits under disease pressure in natural and artificial condition
Significant reduction in yield and all yield contributing traits was observed across the all studied genotypes under Stemphylium disease infestation either disease inoculation was happened in natural epiphytotic or artificially inoculated conditions. The observed disease rating scale values revealed that the disease severity was higher in artificial inoculation than that of natural epiphytotic condition in the field. That’s why comparatively lower yield reduction as well as adverse effect on all yield contributing traits were obtained in natural epiphytotic condition during two consecutive growing seasons in all studied genotypes than artificially fungus inoculated condition in the field. The reduction of yield, and yield contributing traits happened due to loss of green leaflets of the leaves which enhanced the reduction of photosynthetic capacity at flowering and pod filling stage, which ultimately hampered the grain quality and quantity in lentil35. About 80-92.5% yield reduction was recorded in Banglaesh36 and 82.55% disease incidence and up to 93.4% yield loss was recorded in India37. Therefore, screening of lentil germplasm under artificial inoculation is the more effective and accurate method for selecting the Stemphylium resistant genotypes of lentil.
Molecular characterization of the causal pathogen of Stemphylium blight
Among the fungal diseases Stemphylium blight is the severe pathogen that causes enormous losses to lentil production. This fungal pathogen could be found alone or in combination with other fungal pathogens causing severe yield loss due to severe foliage disease like Stemphylium blight in lentil. Therefore, it is urgent need for appropriate and cost-effective research to properly manage of this important disease.
In this present study, molecular characterization of 3 isolates of Stemphylium botryosum (SBI1; SBI2& SBI3) were collected from our infected experimental plot, and nearby plot of the experiment for using genome sequencing of the rDNA sequencing of the ITS region (contains internal transcribed spacer 1, 5.8 S ribosomal RNA, internal transcribed spacer 2, and large subunit of ribosomal RNA) and glyceraldehyde-3-phosphate dehydrogenase (gdp) from the Pulse Research Centre, Ishurdi, Bangladesh. The BLASTn search revealed that the isolated fungi derived from Stemphylium blight symptomatic leaves of lentil was Stemphylium botryosum. On the other hand, phylogenetic analysis based on the rDNA sequence data played a significant role in distinguishing species in Stemphylium 38,39,40. Previous studies used combined ITS regions to clarify the taxonomy and phylogenetic relationships of species in Stemphylium41,42,43. The current phylogenetic analyses with combined ITS sequence data gave good resolution of phylogenetic separations among Stemphylium species and provide insights in to taxonomic novelties. Yanez (2001)44; Chou and Wu (2002)45; Inderbitzin et al. (2009)42; and Crous et al. (2014)38 have amplified and sequenced the target regions of 5.8 S rRNA (ITS) and gpd genes for the detection of Stemphylium from the infected lentil. Wang et al., (2010)41 also reported that utilization of the phylogenetic relationships is essential to identify the new species of Stemphylium based on combined analyses of morphological and molecular data. Bakr et al. (1981)9 firstly identified the pathogen of Stemphylium botryosum as a causal organism of Stemphylium blight disease in lentil based on the conidia morphological features.
Based on the phylogenetic relationship of the present study it is clear that a variation exists among the Stemphylium botryosym isolates. From the phylogenetic analysis the isolates SBI1, and SBI2, yielded a same group in a same cluster, while other SBI3 isolate had clustered differently. Although these three isolates were collected from the same location but they are showing difference cluster. So, the microclimates of different area may influence the pathogen morphology and genetic diversity. Therefore, further studies need to be conducted with a huge number of isolates from different areas to know the genetic variability of Stemphylium botryosum by sequencing others isolates form different hosts and agro-ecological zones from Bangladesh.
Conclusion
PRC breeding program has been benefited from the development of a screening method that would identify susceptible lines which could be eliminated. Furhthermore, exotic lines with better resistance than commercial varieties were identified and that could be utilized as parents for enhancing the resistance in new varieties development programs. However, from this study, it was concluded that there was no resistance source or very susceptible genotypes of lentil against Stemphylium blight disease in Bangladesh. The maximum genotypes were identified as susceptible to Stemphylium blight disease. Only six genotypes viz.; G7; BARI Masur-7, G9; LRIL-21-112-1-1-1-1-6, G16; PRECOZ, G40; BLX-12004-5, G52; BLX 10001-1, and G59; BLX 09015 were identified as moderately resistance to moderately susceptible and four genotypes viz.; G8; LRIL-21-67-1-1-1-1, G15; LR-9-25, G28; BLX-05001-10, G29; BLX-05002-6 were as moderately susceptible genotypes with better yield performance. So, these genotypes might be used in the future breeding program of lentil for getting Stemphylium blight disease resistance source with stable and higher yield performance. Molecular study for the confirmation of the causal organism of Stemphylium blight was done This study confirmed that Stemphylium botryosum is the causal organism of Stemphylium blight disease of lentil, and this is the first reporting of molecular characterization of Stemphylium botryosum in Bangladesh.
Materials and methods
Plant materials
A total of 60 lentils genotypes including, local land race, commercial varieties, exotic lines, International Centre for Agricultural Research in the Dry Areas ICARDA developed advance lines, Pulses Research Centre (PRC) breeding lines were used in this study (Appendix IV).
Experimental site and design
The study was conducted at experimental field of Pulses Research Center (PRC), Bangladesh Agricultural Research Institute (BARI), Ishurdi, Pabna (24.030 N, 89.050 E, and 16 m elevation) followed by Alapha Lattice design with six blocks for minimizing the experimental errors. The unit plot size was 2.5 m long with 4 rows having row to row distance of 30 cm. The test entry was sown in the middle two rows with the surrounding the susceptible variety BARI Masur-1 in the border two rows in each plot. The experiment was conducted during 2019-20 and 2020-21 with consecutive growing seasons, maintaining the similar sowing date, disease pressure and same crop husbandry. Similar experimental design was followed in case of artificial inoculation during the year 2021-22. In case of artificial inoculated experiment, the unit plots size was 2.5 m long with single row of each tested genotypes. All intercultural operations were done for ensuring optimum plant growth.
Creating disease pressure
Creating the disease environment in the experimental plots; (i) the experiment was sown one month later (10 December, 2019 and 10 December 2020) compared with the optimum date of sowing. (ii) the tested lines were sown in the middle two rows, and the Stemphylium blight disease susceptible variety BARI Masur-1 was sown in the two border lines of the both sides of the tested genotypes in each plot, and (iii) The previous year infected plants debris was sprayed as source of inoculum46 to the middle two rows for enhancing the disease pressure into the tested entries. In addition, mist irrigation was applied for creating favourable weather condition at experimental plot and enhancing the natural epiphytotic condition of the pathogen Stemphylium botryosum.
Stemphylium botryosum isolation, identification, inoculum preparation and inoculation procedure in the experimental field
Three isolates were collected from the nearby infected lentil field of experimental plots at Research station, PRC, BARI, Ishurdi, Pabna. Collected infected leaf samples were taken in the sealed zipper polythene bags and it was carried in an ice-box from PRC, Ishurdi, Pabna to Plant Pathology Division, BARI, Gazipur for inoculum development. The leaf blight symptomatic samples were placed on blotting papers after well washed with chlorox and distilled water. After one week, mycelial mass with discs from the blotter paper were placed on V8 agar juice media plates (V8: 200 mL of Campbell’s V8 juice, 4 g calcium carbonate and 15 g Difco Technical agar). The V8 agar juice plates were stored in Incubator at 25 °C under cool white, fluorescent light (44 µmol/m2/s) for a photoperiod of 8 h light/day (9.00 am to 5.00 pm). After two weeks of incubation, the conidia were developed in the V8 agar juice media plates, and conidia of Stemphylium botryosum were transferred in to the Petri plates for sub-culture for preparation of the large amount of conidia. After three weeks incubation at 25 °C the subculture, conidia from the sub-cultured Petri pates (9 mm) of fungi Stemphylium botryosum were harvested by adding 10 mL of double distilled water into each Petri dish to develop a conidial suspension. The suspensions were filtered twice through cheese cloth and every sample was counted three times using a hemocytometer [Neubauer Improved, PRECICOLOR (HBG), Germany] for estimation of concentration of conidia in the suspension, where conidial concentration was obtained 1.3 × 105 spores/mL. The harvested conidia with 1.3 × 103 conidia/mL were sprayed with 0.02% v/v Tween®20 as adhesive materials for spore inoculation in the experimental plots. These artificial spore suspensions were sprayed by power spray machine with the finest setting until run off at 40 days old genotypes just first flowering stage or pre-flowering stage of the test entries. The whole procedure of inoculum development and spray in the field has been presented in the Fig. 14. After two hours of inoculations, mist irrigation was applied for maintaining the high humidity (80–90% RH) for 48 h in the experimental field. Artificial spore culture development process was done in the Plant Pathology Division, BARI, Gazipur and artificial inoculation was done in the experimental site at PRC, BARI, Ishurdi, Pabna.

Different steps for inoculum development and inoculation suspension preparation at Lab, and artificial inoculation in the field at PRC, BARI, Ishurdi, Pabna, Bangladesh within 89.050E longitudes and 24.030N attitudes during flowering stage; (A) Collected spores from infected plant, (B) Spores placement at media, (C) Spores culture at media, (D) Cultured pathogen, (E) Sub-cultured for pure cultured develop., (F) Confirmed spores by Microscrope, (G) Collection of spores (H) Suspension preparation, (I) Estimation of spores concentration by Haemocytometer, (K) Inoculation of spores in the field by power sprayer (E-mail: akzaman118@yahoo.com).
Pathogenicity test
After 14 days of artificial inoculation the disease symptoms were observed in leaves of the infected plants that was similar to previous symptoms of Stemphylium botryosum. For further confirmation of pathogen, the disease samples were taken and observed in the light microscope at PRC pathological lab, BARI, Ishurdi, Pabna. The morphology of these conidia under microscopic view was confirmed to the Stemphylium botryosum. Then these conidia were used for pure culture of Stemphylium botryosum for further molecular identification.
Data collection and analysis
Symptoms were observed after two weeks of inoculation, and scored on a 1–9 disease rating scale as described in Table 6 where, 1 indicated no infection or tiny non-spreading lesion, and 9 indicated the 100% plant death. The disease severity of the present investigation was recorded at three different growing stages like flowering, pod filling and maturity stages at seven days interval starting from 24 January of each growing season at flowering or pre-flowering stage based on the whole plot disease severity. Finally, Area Under Disease Progress Curve (AUDPC) was calculated using the following formula47:
$$AUDPC = sumlimits_{{i = 1}}^{n} {[(Y_{{i + n1}} } + Y_{i} )/2][X_{{i + 1}} – X_{i} ]$$
where, Yi = disease severity score at the ith observation, Xi = time (days) at the ith observation, and n = total number of observations.
Molecular study was done followed by the following procedures for confirmation of Stemphylium botryosum as a causal pathogen of Stemphylium blight disease of lentil.
Raising of Stemphylium botryosum culture for genomic DNA extraction
Preparation of culture
Collected three isolates from the artificially inoculated infected plants were grown on PDA. Hyphal tip of 4 days old pure cultures was transferred into 250 ml conical flask containing 100 ml potato dextrose (PD) broth and placed on electric shaker for four days at 120 rpm at 25 °C. Four days old mycelia of Stemphylium botryosum from potato dextrose broth were filtered with filter paper to remove water. The filtered mycelium mate was blotted, and dried to remove moisture as much as possible. The blotted, and flattened mycelium of Stemphylium botryosum was folded into an aluminium foil paper and frozen at -20 °C.
Fungal genomic DNA extraction
DNA was extracted separately from three isolates (SBI1; SBI2 & SBI3) of Stemphylium botryosum following the method of Reader and Broda (1985)48. Frozen mycelia were grounded in to fine powder using mortar and pestle by liquid Nitrogen. An amount of 0.5gm powdered mycelia were poured into 1.5 µl Centrifuge tube and homogenized with 600 µl of extraction buffer (250mM NaCl, 100mM Tris-HCl,100 m MEDTA, Sigma Aldrich) by stirring on vortex for lysis of the cells. 100 µl of 10% sodium dodecyl sulphate (SDS) was added in to centrifuge tube containing powdered mycelia and extraction buffer and then mixed by inverting the tube 5–6 times and incubated for 30 min at 65 °C in water bath. After incubation, 264 of 3 M sodium acetate (pH.5.2) was added into the centrifuge tube, mixed gently and kept in ice for 15 min. The mixture was then centrifuge (Hawkd15/05) 13,000 rpm at 4 °C for 10 min and supernatant was transferred in to a new tube. After that, (500 µl) of Chloroform was added in to the collected supernatant and mixed thoroughly by vortex. The mixer was centrifuge at 12,500 rpm at 4 °C for 30 min to precipitate the protein and long chain polysaccharides from the DNA. The upper aqueous phase was collected into new tubes very carefully without disturbing the interface.
Precipitation and re-suspension of DNA
Precipitation of DNA was done following the procedure of Wilson (2001)49. Two volumes of pure ethanol (99%) were added into the DNA suspension, kept on ice for 10 min to allow the DNA molecules to aggregate. The DNA was precipitated down to the bottom of the centrifuge tube as a pellet by centrifuging at 13,000 rpm at 4 °C for five minutes. Then the supernatant was discarded using a wide bore pipette and preserved the centrifuge tube with the DNA pellet. An amount of 800 µl of 70% ethanol was added into the centrifuge tube, stirred on vortex for washing the DNA pellet and centrifuge at 13,000 rpm at 4 °C for two minutes, ethanol was discarded using a micropipette. The centrifuged tube containing DNA pellet was dried into vacuum desiccators for two minutes. The DNA pellet was re-suspended by 50 µl TE buffer (10mM Tris, pH-8.0, 1 M EDTA, Sigma Aldrich). The DNA solution was preserved at -20 °C for further studies.
Quantification of genomic DNA
Prior to set of PCR reaction the extracted fungal genomic DNA was check by run on 0.7% agarose gel and then quantify by spectrophotometer (Nanodrop-1000).
Preparation of 0.7% agarose gel
For preparation of 0.7% agarose gel, 0.7 g agarose was weighed in a conical flask. An amount of 100 ml of 1×TAE buffer was poured into the conical flask having 0.7 g agarose and melted by heating in a microwave oven until dissolved fully. The melted agarose gel was cooled down under flowing tap water. The plate of the gel tank was placed flat on the table. The gel was poured smoothly and continuously starting from one corner until reached to the other corner. Then comb was inserted halfway into the gel immediately for making well, allowing an hour for polymerization before removing the comb gently. The casted plate was assembled in the tank. The gel was submerged into in to the 1×TAE running buffers in gel tank.
Gel loading and running
PCR product (10 µl) stained with 2 µl of 6× loading dye was loaded in to the well of the agarose gel and electrophoresed for 25 min at 100 volts to move the negatively charged DNA towards the anode.
Visualization of the PCR product in 2% agarose gel
The gel was taken out of the gel tank and stained with ethidium bromide solution (0.5 µl) for an hour. The stained gel was rinsed with water for distaining, illuminated on UV trans-illuminator and photographed by gel documentation (Bio-Rad) for measuring the bands of amplified DNA fragments. The selected RAPD primer was used for the rest of the studies with 03 selected isolates for amplification of DNA. The gel used to resolve the amplified DNA was stained, illuminated under UV light and the fingerprints photographed by gel documentation.
Amplification of multiple locus of the isolated fungi for sequencing (ITS region & glyceraldehyde-3-phosphate dehydrogenase)
PCR was performed using BIORAD Thermo-cycler (USA). The primer pairs ITS-1 (5’TCCGTAGGTGAACCTGCGG3’) and ITS4 (5ʹTCCTCCGCTTATTGATATGC3ʹ)50 were used for rDNA amplification. For the amplification glyceraldehyde-3-phosphate dehydrogenase (gdp) gene the primer pair gdp1 5ʹCAACGGCTTCGGTCGCATTG3ʹ, and gdp2 5ʹGCCAAGCAGTTGGTTGTG3ʹ were used respectively by Berbee et al. (1999)51. The PCR amplification was carried out in 25 µl reaction mixture containing 1ng of DNA sample, 5 µl of 5× PCR buffer, 2.5mM MgCl2, 2.0 µl of 2mM dNTPs (Promega, USA), 20pmol of each forward and reverse primer (1.0 µl) and 0.2 µl of Taq DNA Polymerase and made up to 25 µl with nuclease free water. The PCR conditions include initial denaturation at 94 °C for 3 min, 30 cycles of denaturation at 94 °C for 30 s, primer annealing at 55 °C for 30 s, followed by primer extension for 30 s at 72 °C and final extension at 72 °C for 10 min. The amplicon was gel purified by PCR clean up kits (Promega, USA) and sending for sequencing to National Institute for Biotechnology (NBR), Bangladesh. Based on the morphological characteristics of the fungi we have sequenced three isolates namely SBI1, SBI2 and SBI3 respectively.
Sequence analysis
The assembly of fungal genome was performed using the web service of CAP3 program (http://doua.prabi.fr/software/cap3) and Bio-Edit V5. ITS-rDNA sequences were aligned using the CLUSTALW2 program52. The assembled sequenced contain partial sequence of 18s ribosomal RNA gene, ITS-1, 5.8s ribosomal RNA gene, ITS2 complete sequence and partial sequence of 28 S ribosomal RNA partial sequence and the partial sequence of the glyceraldehyde-3-phosphate dehydrogenase (gdp) were submitted to NCBI (https://www.ncbi.nlm.nih.gov/) with the accession number OQ442935, OQ442936 & OQ442937 for ITS and OQ534370, OQ534371 & OQ534372 for gdp respectively.
The submitted sequence information of all 03 isolates that were responsible for Stemphylim blight disease of lentil in Bangladesh were presented in the Appendix V, VI, and VII. Phylogenetic tree was constructed based on the Neighbor-Joining [NJ] Method in MEGA version 11 (Fig. 13) using Kimura-two parameter model with 1000 bootstrap replications53.
Weather data
The weather data of the three growing seasons were collected from the Meteorological Department, Ishurdi, Pabna (Appendix VIII).
Data availability
The partial sequence data that support the findings of this study have been deposited in National Center for Biotechnology Information (NCBI) Archive with the primary accession code “OQ442935.1; OQ442936.1; OQ442937.1 ”. The web link of these accession numbers are available in the following data base weblink system- https://www.ncbi.nlm.nih.gov/nuccore/OQ442935.1; https://www.ncbi.nlm.nih.gov/nuccore/OQ442936.1; https://www.ncbi.nlm.nih.gov/nuccore/OQ442937.1.
References
-
Gupta, S. D. et al. Lentils (Lens culinaris L.) a rich source of folates. 61 (2013), 7794–7799. (2013).
-
Kissinger, G. Pulse crops and sustainability: A framework to evaluate multiple benefits. (Accessed 29 May 2019) http://www.fao.org/pulses (2016).
-
Bayaa, B. & Erskine, W. Lentil pathology. In: Pathology of Food and Pasture Legumes D. Allen and J. Lenne (Eds), 423–472. (Commonwealth Agricultural Bureaux International, U.K in association with International Crop Research Center for the Semi-Arid Tropics, Patancheru 502 324. Andhra Pradesh, India, 1998).
-
Mwakutuya, E. & Banniza, S. Influence of temperature and wetness periods on the development of stemphylium blight on lentil. Plant. Dis. 94, 1219–1224 (2010).
-
FAOSTAT Database. Food and agriculture organization of the United Nations. Rome, Italy. FAO. (Accessed 29 May 2020). http://www.fao.org/faostat/en/#home.
-
Kant, P., Materne, M., Rodda, M. S. & Slater, A. T. Screening lentil germplasm for stemphylium blight resistance. Australasian Plant. Pathol. 46, 1–8 (2017).
-
Morrall, R., Carriere, B., Pearse, C., Schmeling, D. & Thomson, L. Seedborne pathogens of lentil in Saskatchewan in 2005. Can. Plant. Disease survey-Inventaire Des. Maladies Des. Plant.s Au Can. 86, 1404–1406 (2006).
-
Kumar, S., Barpete, S., Kumar, J., Gupta, P. & Sarker, A. Global lentil production: constraints and strategies. SATSA Mukhapatra Annu. Tech. Issue 17, (2013).
-
Bakr, M. A. & Ahmed, F. Development of Stemphylium blight of lentil and its chemical control. Bangladesh J. Plant. Pathol. 8, 39–40 (1991).
-
Bakr, M. A. & Ahmed, F. Development of Stemphylium blight on lentil genotypes and its chemical control. Bangladesh J. Plant. Pathol. 8, (1&2), 39–40 (1992).
-
Islam, S. M. A. Search for resistance and chemical control against Stemphylium blight disease of lentil. MS thesis, Department of Plant Pathology, Sher-e-Bangla Agricultural University, Dhaka, Bangladesh (2014).
-
Sinha, J. N. & Singh, A. P. Effect of environment on the development and spread of Stemphylium blight of lentil. Indian Phytopathol. 46, 252–253 (1993).
-
Islam, M. A., Islam, S. M. A. & Sathi, M. A. Identification of lentil varieties/lines resistant to stemphylium blight considering disease reaction and yield. Malays J. Sustain. Agric. 4 (1), 22–25. https://doi.org/10.26480/mjsa.01.2020.22.25 (2020).
-
Dokken-Bouchard, F. Stemphylium blight of lentil. Crop Prod. News. 32 (7), 7–9 (2010).
-
Hosen, M. I., Ahmed, A. U., Zaman, J., Ghosh, S. & Hossain, K. M. K. Cultural and physiological variation between isolates of Stemphylium botryosum the causal agent of Stemphylium blight disease in lentil (Lens culinaris). World J. Agric. Sci. 5, 94–98 (2009).
-
Zamir, D. Improving plant breeding with exotic genetic libraries. Nat. Rev. Genet. 2 (12), 983–989 (2001).
-
Beare, M. Investigation into Stemphylium botryosum resistance in lentil. Undergraduate thesis, Department of Plant Sciences, University of Saskatchewan, Saskatoon, Saskatchewan (2002).
-
Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39, 783–791 (1985).
-
Tajima, F. & Nei, M. Estimation of evolutionary distance between nucleotide sequences. Mol. Biol. Evol. 1 (3), 269–285 (1984).
-
Razzak, M. A., Islam, M. A., Rahman, M. R., Sathi, M. A. & Atikuzzamman, M. Screening of lentil germplasm against Stemphylium blight by observing disease reaction in three different stages. Malalysian J. Halal Res. 1 (2), 15–18 (2018).
-
Yadav, N. K. et al. Source of resistant against Fusarium wilt and Stemphylium blight in lentil (Lens culinaris Medikus). Inter J. Appl. Sci. Biotech. 5, 102–110 (2017).
-
Subedi, S. et al. S. Integrated approach for the management of new threat Stemphylium botryosum Walr causing blight of lentil (Lens culinaris Medik.) Turkish J. Agric. Nat. Sci. (2014).
-
Khadka, R. B., Pathak, N., Gharti, D. B. & Joshi, S. Screening of lentil genotypes for resistance against Stemphylium blight at RARS, Nepalgunj. Proceedings of the 29th Winter Crops Workshop Held at 11 & 12 June 2014 at RARS (2014).
-
Gharti, D. B., Joshi, S., Darai, R., Ghimire, T. N. & Chadaro, M. B. Identifying sources of resistance to major diseases of lentil. Paper presented in the 28th National Winter Crops Research Workshop held at RARS, Lumle, Kaski. Falgun 25–26. (2011).
-
Kumar, P. Genetics of resistance to Stemphylium leaf blight of lentil (Lens culinaris) in the cross BARI Masur-4 × CDC Milestone. MS thesis, Department of Plant Science, Saks Saskatchewan, Saskatoon, Canada (2007)
-
Alam, K. H., Ali, M. K., Rashid, M. H., Haque, A. H. M. M., Sultana, N. A. Screening of lentil lines resistant to Stemphylium blight. Bangladesh Int. J. Adv. Res.Tech. 8 (1), 2278-7763 (2019).
-
Francovig, P. C., Mehta, Y. R., Fonseca, N. S., Reis, M. Sources of resistance to Stemphylium solani in cotton cultivars. Summa Phytopathol. 25 (3), 217-222 (1999).
-
Bakr, M. A. Plant protection of lentil in Bangladesh. In: Erskine W, Saxena MC (Eds), Lentil in South Asia, Proceedings of the Seminar on lentil South Asia 236, (ICARDA, 1993).
-
Ahmed, H. U., Bakr, M. A. & Alam, K. B. Pathogen survey of major winter and summer pulses in Bangladesh. Proceedings of National workshop on Pulses BARI, Joydebpur during 18–19 August 1981 (1981).
-
Chowdhury, A. M., Ahmed, A. & Zaman, M. Studies on the defense structural factors of some susceptible and resistant varieties of lentil plants. Mycological Res. 35, 35–39 (1997).
-
Bakr, M. A. Check list of pulse dis. In Bangladesh. J. Plant. Pathol. 10 (1–2), 13–16 (1994).
-
Mia, M. Z., Mian, M. A. K. & Rahman, M. M. Performance of exotic lentil germplasm in Bangladesh. LENSs Newsl. 13 (2), 12–13 (1986).
-
Suheri, H. & Price, T. V. Stemphylium leaf blight of Garlic (Allium sativum) in Australia. Australasian Plant. Pathol. 29 (3), 192–199 (2000).
-
Basallote-Ureba, M. J., Prados-Ligero, A. M., Melero-Vara, J. M. Aetiology of leaf spot of garlic and onion caused by Stemphylium vesicarium in Spain. Plant Pathol. 48 (1), 139-145 (1999).
-
Hay, F. S., Sharma, S., Hoepting, C., Strickland, D., Luong, K., Pethybridge, S. J. Emergence of Stemphylium leaf blight of onion in New Yorkassociated with fungicide resistance. Plant Dis.103 (12), 3083-3092. https://doi.org/10.1094/PDIS-03-19-0676-RE (2019).
-
Bakr, M. A. & Zahid, M. I. Stemphylium blight a new foliar disease of lentil in Bangladesh. J. Plant. Pathol. 2 (1), 69–70 (1986).
-
Mandal, D., Bhattacharyya, P. K., Das, R. & Bhattacharyya, S. Screening of lentil germplasms for Stemphylium blight resistance in West Bengal condition. J. Crop Weed. 15 (2), 155–162 (2019).
-
Crous, P. W. et al. The genera of fungi: fixing the application of type species of generic names. IMA Fungus. 5, 141–160 (2014).
-
Kurose, D. et al. Taxonomic re-examination of several Japanese Stemphylium strains based on morphological and molecular phylogenetic analyses. J. Gen. Plant. Pathol. 81, 358–367 (2015).
-
Leavitt, S. D., Moreau, C. S. & Lumbsch, H. T. The dynamic discipline of species delimitation: progress toward effectively recognizing species boundaries in natural populations, p11-44, In: Recent Advances in Lichenology, India: Springer (2001).
-
Wang, Y., Geng, Y., Pei, Y. & Zhang, X. Molecular and morphological description of two new species of Stemphylium from China and France. Mycologia 102, 708–717 (2010).
-
Inderbitzin, P., Mehta, Y. R. & Berbee, M. L. Pleospora species with stemphylium anamorphs: a four-locus phylogeny resolves new lineages yet does not distinguish among species in the pleospora herbarum clade. Mycologia 101, 329–339 (2009).
-
Camara, M. P. S., O’Neill, N. R. & Berkum, P. Phylogeny of Stemphylium spp. Based on ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 94, 660–672 (2002).
-
Yanez, M. J. M. Molecular and morphological analyses of Alternaria and Stemphylium from onion. Master Thesis. Cornell University, Ann Arbor, Michigan, United States of America (2001).
-
Chou, H. & Wu, W. Phylogenetic analysis of internal transcribed spacer regions of genus alternaria, and the significance of filament-beaked conidia. Mycological Res. 106, 164–169 (2002).
-
Mwakutuya, E. Epidemiology of Stemphylium blight on lentil (Lens culinaris) in Saskatchewan. MS thesis, Department of Plant Sciences, 51 Campus Drive, University of Saskatchewan, Saskatoon, Saskatchewan S7N 5A8 (2006).
-
Shaner, G. & Finney, R. E. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat. Phytopathology 67, 1051–1056 (1977).
-
Reader, U. & Broda, P. Rapid Preparation of DNA from filamentous fungi. Letter Appl. Microbiol. 1, 17–20 (1985).
-
Wilson, K. Preparation of genomic DNA from bacteria. Curr. Proto Mol. Biol. 56 (1), 2–4 (2001).
-
White, T. J., Bruns, T., Lee, S. & Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: (eds Innis, M. A., Gelfand, D. H., Sninsky, J. J. & White, T. J.) PCR Protocols a Guide To Methods and Applications. Academic; San Diego, CA, USA, 315–322 (1990).
-
Berbee, M. L., Pirseyedi, M. & Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 91, 964–977 (1999).
-
Larkin, M. A. et al. Clustal W and clustal X version 2.0. Bioinformatics 23, 2947–2948 (2007).
-
Kumar, S., Stecher, G., Li, M., Knyaz, C. & Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing plat-forms. Mol. Biol. Evol. 35 (6), 1547–1549 .
-
Tamura, K., Stecher, G. & Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38 (7), 3022–3027. https://doi.org/10.1093/molbev/msab120 (2021).
Acknowledgements
This research has been financed by OCPF-ICARDA-BARI Collaborative Project, Regional Coordinator, ICARDA, New Delhi. The authors would like to acknowledge the support of OCP Foundation, ICARDA, Morocco.
Funding
The publication of this article in Open Access mode and finding provided by the Regional Coordinator of South Asia and China, ICARDA, New Delhi. India.
Ethics declarations
Competing interests
The authors declare no competing interests.
Consent for publication
All authors have approved the manuscript and agree with its submission to Journal of Scientific Report’s.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Aktar-Uz-Zaman, M., Haque, M.A., Sarker, A. et al. Screening of lentil genotypes against stemphylium blight disease and molecular identification of causal organism. Sci Rep 15, 32318 (2025). https://doi.org/10.1038/s41598-025-15587-4
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-15587-4