
- Article
- Open access
- Published:
- Jennifer A. Simonovich1 na1,
- Sabrina L. Macias1 na1,
- Mingqi Zhou1 na1,
- Marija Portelli1,
- Isha Verma1,
- Arun Wanchoo1,
- Alexander J. Kwiatkowski1,
- Eric Y. Helm2,
- Tran B. Ngo1,3,
- Shamima Islam4,
- Carlos Moran Segura5,
- Jonathan V. Nguyen5,
- Robert Maile3,
- Shannon M. Wallet6,
- Gregory A. Hudalla1,
- Dorina Avram2,4,7 &
- …
- Benjamin G. Keselowsky1,8
npj Biomedical Innovations volume 2, Article number: 36 (2025) Cite this article
Subjects
Abstract
Localized anti-inflammatory treatment options for psoriasis are limited. Addressing this, a fusion protein was created consisting of the regulatory enzyme indoleamine 2,3-dioxygenase (IDO) fused to carbohydrate-binding protein galectin-3 (Gal3), termed IDO-Gal3, and therapeutic potential was investigated in a murine model of imiquimod-induced psoriasis. Prior work established that Gal3 fusion provides prolonged localized tissue retention of injected fusion protein, which colocalizes therapeutic to the site of inflammation, limits systemic distribution, and reduces potential for off-target side-effects. A single subcutaneous administration at disease onset remarkably decreased the disease metrics of skin redness, scaling and thickening, as measured by the psoriasis area and severity index (PASI) composite score. Infiltrating immune cells, particularly neutrophils and gamma delta T cells, and inflammatory responses were significantly reduced. Transcriptomic analyses indicated inflammation and fibrosis-associated programs were also substantially reduced following IDO-Gal3 treatment. These data demonstrate IDO-Gal3 treatment ameliorated psoriasis with concomitant remodeling of inflammatory and fibrotic programs.
Introduction
Plaque psoriasis is a chronic inflammatory skin disease characterized by thickened, scaley skin lesions1. Approximately 8 million individuals in the United States are diagnosed with psoriasis2, resulting in an estimated economic burden of $35.2 billion3. Though the exact cause of psoriasis is unknown, excessive inflammation of key immune cell populations have been implicated in initiating and maintaining inflammation. Plaque psoriasis is characterized through the activation of the TNFα-IL-23-IL-17 inflammatory pathway4,5,6,7. Keratinocyte overactivation amplifies IFN-γ, IL-17, IL-22, and other inflammatory cytokine signals from T helper (Th) 17 and Th1 CD4 + T cells, which then induce keratinocyte hyperplasia8,9,10,11,12. While psoriasis is considered a T cell mediated disease, innate immune cells present in the skin also play a role in driving pathology. Neutrophils, which can also produce IL-17A6, contribute to the T-cell mediated inflammatory loop seen between T cells and keratinocytes through the release of tumor necrosis factor alpha (TNFα)13. Neutrophil extracellular trap (NET) formation are enhanced in psoriatic patients14 and correlate to severity of disease15. The process of releasing NETs, NETosis, amplifies inflammation in psoriasis, reportedly through the release of RNA16. Macrophages also contribute to psoriasis pathology, as shown in murine models of psoriasis17, and human patients have a skewed M1-like (classically activated, or pro-inflammatory) to M2-like (alternatively activated, or anti-inflammatory) macrophage phenotype, where the ratio of M1 to M2 is correlated to disease severity18. The current standard of care for psoriasis is systemically administered steroids or immunosuppressants, and multiple biologic antibody immunotherapies, targeting TNF, IL-12/23, IL-17, or IL-23, have been approved for use by the US FDA1. Current therapies require a lifetime of continued administration, which can lead to off-target effects19 and render patients vulnerable to opportunistic infections20,21.
The immunomodulatory enzyme indoleamine 2,3-dioxygenase 1 (IDO) is the rate limiting enzyme of the kynurenine pathway and its potent immunomodulatory effects were first described in a key role in preventing rejection of allogeneic fetus during pregnancy22. Analysis of circulating leukocytes from patients with psoriasis correlate IDO deficiency with disease pathology23, suggesting administration of exogenous IDO may be advantageous in treatment. Along this line, both the delivery of kynurenine pathway metabolites as well as cells constitutively expressing IDO have been reportedly been effective in reducing IMQ-induced psoriasis24,25. Improving upon these approaches could include new strategies to bypass limitations related to 1) retention of viable cells and therapeutic molecules at site of administration, and 2) control over the effective dose of IDO and/or kynurenine pathway metabolites at the site of inflammation. Previous work in our group demonstrated immunomodulatory effects of IDO, where dendritic cells cultured with exogenous IDO maintained immature phenotype suppressive to antigen-specific T cell proliferation26. Additionally, IDO did not act as a damage-associated molecular pattern when supplemented in culture. In vivo delivery unmodified exogenous IDO, however, is quickly cleared from injected tissues. Addressing this, fusion enzymes to the carbohydrate-binding protein galectin-3 (Gal3) demonstrate enhanced tissue retention27. Fusion of IDO to Gal3 (IDO-Gal3) provided tissue retention of IDO enzyme at various sites of administration, locally produced and kynurenine product, and effectively resolved inflammation in multiple models, including psoriasis28.
Here, we follow our prior work by further investigating the imiquimod (IMQ)-induced psoriasis mouse model29, characterizing cellular and molecular mechanisms of action of IDO-Gal3 treatment. A single subcutaneous injection of IDO-Gal3 administered at the first sign of disease was sufficient to reverse disease severity, reduce epidermal thickening and resolve inflammation. Immune cell infiltration and activation was significantly reduced. RNA-sequencing (RNA-seq) of skin revealed a distinct transcriptomic profile associated with IDO-Gal3 treatment. Transcriptomic analysis on a panel of immunity-associated genes at midpoint and termination demonstrated sustained remodeling effects from a single IDO-Gal3 treatment, with significant reduction in inflammatory and fibrotic programs. Additional multiplex cytokine analyses are consistent with decreased inflammation. Overall, results demonstrate IDO-Gal3 treatment serves as a potent immunomodulatory protein therapy in this pre-clinical psoriasis model.
Results
A single therapeutic subcutaneous administration of IDO-Gal3 reverses psoriasis scores
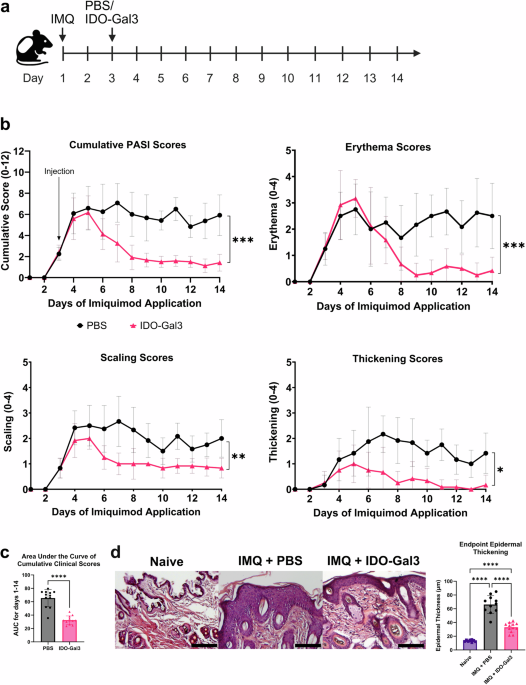
Imiquimod was applied daily on the back skin of mice for 14 consecutive days in this psoriasis model. Therapeutic and control groups received a single subcutaneous injection of IDO-Gal3 or sterile phosphate buffered saline (PBS) administered on day 3 (Fig. 1a). Scores quantifying the psoriasis area and severity index (PASI) metrics of skin erythema (redness), scaling and thickening were recorded. The first appearance of disease metrics occurred on day 3, while the peak of disease was established on approximately day 5 (Fig. 1b). Cumulative PASI and individual scores for mice which received IDO-Gal3 were significantly lower compared to control psoriatic mice (PBS group). Following the peak of disease, cumulative clinical scores of IDO-Gal3 treated decreased, while control animals maintained peak score values. Scores of IDO-Gal3 treated animals decreased to an average PASI score of ~2 by approximately day 8 and remained there until termination on day 14. This trend was observed for each of the individual scoring criteria. The 14-d area under the curve (AUC) of the cumulative PASI score for the IDO-Gal3 treatment group was ~50% of the control psoriatic (PBS) group (Fig. 1c), consistent with psoriasis mitigation. To confirm the necessity for localized immunomodulation through the IDO-Gal3 fusion protein, mice were treated with a non-immunomodulatory fusion of the NanoLuc luciferase to galectin-3 (NL-Gal3) or with wild-type IDO (Supplementary Fig. 2). There were no significant differences between cumulative scores of psoriatic animals that received NL-Gal3, wild-type IDO, or PBS. AUC from PBS, NL-Gal3, and wild-type IDO treated animals were significantly higher than IDO-Gal3 treated animals. These results indicate that the IDO arm is critical to the immunological mechanism by which the fusion protein decreases scores in IMQ-induced psoriasis and that galectin-3 is necessary for retaining the immunomodulatory capabilities of IDO.

a Timeline of the IMQ-induced mouse psoriasis model. b Clinical PASI scores of three criteria (erythema, scaling, and thickening of the skin) recorded daily from each animal (day 1–day 14). Psoriatic skin was subcutaneously injected with either sterile PBS (black circles) or IDO-Gal3 (red triangles) in equal volumes on day 3 of imiquimod application (black arrow labeled “Injection”). Cumulative score was calculated from the sum of individual criteria. c Area under the curve of cumulative clinical score between days 1 and 14 for each animal. d Representative images of skin from a naïve animal injected with sterile PBS (Naïve), a psoriatic animal injected with sterile PBS (IMQ + PBS), and a psoriatic animal injected with IDO-Gal3 (IMQ + IDO-Gal3). Average epidermal thickness (µm) measured from all naïve, psoriatic, and IDO-Gal3 animals. Scale bars 100 µm; data are mean ± SD; n = 12/group; clinical scores (cumulative and individual criteria) on day 14 compared using Kruskal-Wallis test with multiple comparisons; area under the curve and epidermal thickness compared using one-way ANOVA with Tukey’s multiple comparisons test; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001; ns not significant.
Epidermal thickening at day 14 endpoint was further quantified via histology for treated and control groups, as well as naïve tissue for reference (Fig. 1d, e). In corroboration with longitudinal PASI scores, IDO-Gal3 treatment reduced endpoint epidermal thickness to roughly 50% compared to psoriatic PBS control animals but remained higher than naïve epidermis. Additionally, gross histological inspection suggested concomitant cellular infiltration levels commensurate with epidermal thickness across groups, which motivated more detailed investigation.
IDO-Gal3 treatment reduces psoriatic infiltrating immune cells in the dermis and epidermis
Further histological observations at day 14 via immunofluorescence and H&E staining were consistent with reduced immune cell infiltration and inflammatory markers after IDO-Gal3 treatment. Specifically, in IDO-Gal3 treated skin, CD11b+ macrophages, Ly-6G+ neutrophils, and the neutrophil NETosis marker, citrullinated histone H3 (H3Cit), appeared to be trending lower (Fig. 2), as did total infiltrating cell populations via H&E stain. Quantitative assessment by image analysis, however, did not yield p-values meeting our criteria for significance (Supplement Fig. 3), limiting interpretation of these images to qualitative descriptions.

Representative immunofluorescence staining and light microscopy images of day 14 psoriatic skin after treatment with PBS (left) or IDO-Gal3 (right). IDO-Gal3 treatment resulted in the appearance of reductions in intensity of myeloid CD11b+ staining (teal), NETosis marker H3Cit (citrullinated histone H3 regions; red), and infiltrating Ly-6G+ neutrophil number (green), with cell nuclei stained with DAPI (blue). Infiltrating cell number observed via H&E stain appeared reduced with IDO-Gal3 treatment. 20X magnification, scale bars 100 µm.
Immune cell populations were next isolated from both dermis and epidermis and characterized by flow cytometry at day 7. The percent of granulocyte receptor-1 positive (GR-1 + ) neutrophils was decreased in IDO-Gal3 treated epidermis (Fig. 3a). In dermis, CD44, an activation marker increased in T cell populations in psoriatic skin30,31, was reduced by IDO-Gal3 treatment in both total TCRαβ + T cells (Fig. 3b) as well as in CD4+ helper T cells (Fig. 3c). The dermis was also depleted of the CD44+ activation marker in TCRγδ + T cells (Fig. 3d), a key pathogenic T cell subtype in psoriasis32,33. Furthermore, dermal Th17-like γδ T cell levels were reduced, with fewer CCR6+ (Fig. 3e) and RORγt + (Fig. 3f) TCRγδ cells in IDO-Gal3 treated mice compared to PBS control. CCR6 and RORγt are key markers for Th17 cells, and γδ T cells positive for these markers share similar characteristics, such as IL-17 expression, but may precede Th17 development33,34. These findings demonstrate IDO-Gal3 treatment provides modulation in immune cell populations of the skin, decreasing both myeloid populations and psoriasis-specific T cell populations and activation.

a Percent CD11b+ GR-1+ cells in the dermis and epidermis, with representative histograms. Fewer neutrophils are in the epidermis following IDO-Gal3 treatment as compared to PBS control. b Percent CD11b- TCRαβ + CD44+ cells in the dermis and epidermis, with representative histograms. c Percent CD11b- TCRαβ + CD4 + CD44+ cells in the dermis and epidermis, with representative histograms. Fewer activated αβ and CD4+ helper T cells are in the dermis following IDO-Gal3 treatment as compared to PBS. d Percent CD11b- TCRγδ + CD44+ cells in the dermis and epidermis, with representative histograms. e Percent CD11b- TCRγδ + CCR6+ cells in the dermis and epidermis, with representative histograms. f Percent CD11b- TCRγδ + RORγt+ cells in the dermis and epidermis, with representative histograms. Fewer activated TCRγδ+ cells are in the dermis following IDO-Gal3 treatment as compared to PBS. All populations were compared using an unpaired t-test; *P < 0.05; **P < 0.01; ***P < 0.001. No comparison indicates no significance.
IDO-Gal3 treatment reduces immune and fibrosis transcription programs in psoriatic skin
Skin samples were harvested at the day 14 endpoint and transcriptomic analysis was performed to investigate mechanisms related to IDO-Gal3 treatment. Differentially expressed gene (DEG) analysis showed more genes (4037) down-regulated than up-regulated (3,827) in the IDO-Gal3 treatment group than the psoriatic (PBS) control (Fig. 4a, b; Supplementary Data 1). Additionally, a larger fold-change in gene expression levels was observed in down-regulated DEGs compared to up-regulated DEGs (Fig. 4a). Down-regulated genes of interest were the Schlafen family member 3, Slfn3, where down-regulation of Slfn3 corresponds with induction of peripheral Tregs35, the CXC motif chemokine ligand 3, Cxcl3, associated with psoriatic keratinocytes36, and interleukin 1 beta, Il1b, which promotes and maintains differentiation of Th17 cells in psoraisis37. The interleukin 23 receptor, Il23r, which is necessary for IL-23-mediated activation of γδ T cells in the psoriatic dermis32,33, also was down-regulated with IDO-Gal3 treatment.

a Volcano plot representing differentially expressed genes (DEGs) from skin of IDO-Gal3 treated psoriatic animals when compared to PBS-injected control psoriatic animals at day 14 of the model. b Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways enriched using g:GOst from 4037 down-regulated DEGs and 3827 up-regulated DEGs, respectively. Significant enrichment is identified with P-value < 0.01 after g:SCS adjustment. c Volcano plot representing DEGs associated with macrophage polarization. Genes associated with pro-inflammatory “M1”-like macrophage phenotype and tolerogenic/wound healing “M2”-like macrophage phenotype were shown. d Volcano plot representing skin fibrosis related DEGs. Key functions were indicated for the up- and down-regulated DEGs related to skin fibrosis, respectively.
Pathway analysis using the Kyoto Encyclopedia of Genes and Genomes (KEGG) indicated enrichment of specific pathways in both up-regulated and down-regulated DEGs in response to treatment (Fig. 4b, Supplementary Data 2). Among down-regulated pathways were Toll-like receptor signaling, a pathogen response signaling pathway of the innate immune system38, TNF signaling, a pathogenic pathway involved in many autoimmune diseases39, T cell receptor signaling, the main activation pathway for T cells40, as well as FoxO, a key regulator of pathological immune responses41, and mTOR, which serves as central regulator of metabolism, integrating multiple pathways including amino acid sensing, and which is involved in many autoimmune diseases42,43. Pathways up-regulated in response to treatment included oxidative phosphorylation (OXPHOS) cellular metabolism44,45 and mitophagy, the selective removal of dysfunctional mitochondria46. In-depth analysis of macrophage polarization-related genes revealed that expression of pro-inflammatory “M1”-like macrophage genes was significantly decreased in IDO-Gal3 samples (Fig. 4c), consistent with the reduction observed in inflammatory myeloid cell populations (Figs. 2a, 3a). Among genes commonly associated with “M2”-like macrophages, only transforming growth factor beta 1 (Tgfb1) was significantly increased by IDO-Gal3 treatment, where TGFβ1 is a cytokine regulating cell differentiation and growth, with predominantly suppressive action on myeloid immune cells47,48.
Given the prominent role of fibrosis in psoriasis49,50, relative expression of skin fibrosis-associated genes51 was investigated, revealing 54 genes significantly altered following treatment with IDO-Gal3 (Supplementary Data 3). Among these, a number of genes related to inflammation were down-regulated (Fig. 4d), including some associated with immune cell migration, like WNK lysine deficient kinase 1 Wnk1, a regulator of lymphocyte migration52, and C-C chemokine receptor type 2 CCR2, a chemokine receptor involved in monocyte recruitment to psoriatic lesions53,54. Conversely, IDO-Gal3 treatment up-regulated suppressive genes such as Tgfb1, nuclear receptor subfamily 1 group H member 2 Nr1h2, a key gene for regulatory T cell function55, and interferon regulatory factor 5 Irf5, a regulator of innate and adaptive immune cells56, consistent with an immuno-protective, psoriasis-relevant modulation. Regulation of such genes and pathways with roles in inflammation and fibrosis help shed light on mechanisms related to the beneficial effects of IDO-Gal3 in psoriatic skin.
IDO-Gal3 fine-tunes immunity-associated genes to protect skin against inflammation
Targeted gene analysis via NanoString nCounter technology was employed as a complementary, amplification-free approach to quantify immune cell transcripts using the Mouse Immunology Panel57 of 770 metabolic and immune specific gene probes to further investigate the impact of IDO-Gal3 treatment of psoriasis at the day 7 midpoint. From this panel, 27 down-regulated and 33 up-regulated immunity associated DEGs were identified (Fig. 5a, Supplementary Data 4). Several up-regulated genes are involved in the negative regulation of inflammatory cytokines, such as CD200 receptor 1, Cd200r1, a negative regulator of Il-6 production and leukocyte migration58, complement component 3, C3, a mediator of T cell responses linked with promoting regulatory phenotype59,60, and MAF bZIP transcription factor, Maf, which regulates CD4 + T cells61. Also within the up-regulated gene population were several genes involved in adhesion molecule and extracellular matrix (ECM) formation. Three genes for ECM protein production collagen I alpha 1, Col1a1, fibronectin, Fn1, and collagen IV alpha 1, Col4a1 were all up-regulated at day 7, as were the cell adhesion molecules neural cell adhesion molecule 1, Ncam1 and vascular cell adhesion molecule 1, Vcam1. Genotypic level of chymase, Cma1, an enzyme involved in ECM degradation that is down-regulated in psoriatic plaques62, was also increased at day 7 after IDO-Gal3 treatment, suggesting a shift to a more homeostatic skin remodeling. Many of the genes down-regulated in the IDO-Gal3 treated animals are associated with psoriatic pathogenesis, such as interleukin 7 receptor, Il7r, which transmits inflammatory T cell activation signals by IL-763, interleukin 1 alpha, Il1a, an inflammatory cytokine64, interleukin 13 receptor alpha 2, Il13ra2, an IL-13 binding receptor that contributes to early psoriatic inflammation65, interleukin 22 receptor alpha 2, Il22ra2, a receptor for the highly inflammatory IL-22 cytokine66,67, and chemokine (C-C motif) ligand 20, Ccl20, a potential biomarker for psoriatic disease severity68. Additionally, two genes in the pro-inflammatory NFκB pathway69,70, nuclear factor kappa B subunit 1, Nfkb1, and interleukin 18, Il18, a positive regulator of NFκB71,72, were both down-regulated at day 7.

a Volcano plot representing differentially expressed genes (DEGs) from the Immunology Panel in skin samples comparing psoriatic animals treated either with IDO-Gal3 or PBS (control) at day 7. b KEGG pathways (8 down-regulated, 1 up-regulated) enriched using g:GOst from 27 down-regulated DEGs and 33 up-regulated DEGs from the Immunology Panel, respectively. Significant enrichment is identified with P-value adjusted by g:SCS (Padj) < 0.01. c Heatmap representing differential expression of the genes significantly changed in both the transcriptome survey at day 14 and the targeted NanoString analysis at day 7 of the model.
The down-regulated DEGs were enriched in multiple pathways of innate immune response and inflammatory signaling (Fig. 5b, Supplementary Data 5). Down-regulated pathways of note were IL-17 signaling pathway73 and Th17 T cell differentiation37, as both IL-17 and Th17 T cells are pathogenic in psoriasis. The NFκB and TNF signaling pathways, key inflammatory pathways in psoriasis70,74, were also down-regulated (Fig. 5b). This observed TNF signaling pathway down-regulation was consistent with persisting down-regulation to day 14 (Fig. 4b). Interestingly, DEGs in the AGE-RAGE signaling pathway also were up-regulated. This could be interpretated as incongruous, as AGE-RAGE signaling is involved in oxidative stress responses which can contribute to the pathogenesis of psoriasis75. However, the inflammatory signaling of AGE-RAGE converge onto NFκB76,77, whose down-regulation by IDO-Gal3 treatment (Fig. 5b) may serve to constrain such AGE-RAGE signaling responses.
Comparing the NanoString targeted analysis data at the day 7 midpoint and the bulk RNAseq transcriptomic survey at the day 14 endpoint, there were 36 DEGs detected in both data sets (Fig. 5c). Genes of encoding pro-inflammatory proteins Il22ra2, IL1rn, Il7r, and nuclear factors of activated T cell 3, Nfatc3, a promoter of inflammation and fibrosis78, were down-regulated across both data sets. Others, such as Cd38, a glycoprotein marker for M1 macrophages that is found in various skin diseases79,80, Cd86, a costimulatory molecule whose expression correlates to psoriatic disease severity81, interleukin 6 cytokine family signal transducer, Il6st, which is highly expressed in psoriatic skin82, and signal transducer and activator of transcription 2, Stat2, a transcription factor in the JAK/STAT pathway pathogenic in psoriasis83, were up-regulated at day 7 but down-regulated by day 14, suggesting a resolution of these inflammatory signals. Structural ECM and adhesion molecule genes encoding Fn1, Ncam1, and Vcam1 were also up-regulated at the midpoint but decreased expression by endpoint, suggesting a dynamic and evolving tissue remodeling effect throughout disease and treatment progression. Interesting, Lgals3, which encodes endogenous Gal-3, was down-regulated at day 7 and up-regulated at day 14, further implicating resolution of disease, as Gal-3 is significantly reduced in psoriatic lesions84. The fine-tuning of immunity-associated genes observed at day 7, along with the sustained modulation of immune signaling at day 14, demonstrated the long-term effective adjustment of innate immunity and inflammatory response by IDO-Gal3 in the treatment of psoriasis.
Out of the down-regulated genes and signaling pathways identified as relating to immune and fibrotic activity, many were involved in recruitment and activation of neutrophils and γδ T cells, with a high degree of overlap between the two cell populations (Fig. 6)34,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136. This down-regulation of pro-inflammatory genes and signaling pathways may contribute to disease amelioration. Additionally, there were multiple genes and pathways identified that were involved with both neutrophil and γδ T cell recruitment and activation and were down-regulated at both day 7 and day 14, such as Il22ra288,89, Il7r85, and the TNF signaling pathway98,99. Very few immunosuppressive genes and pathways were down-regulated, with only one instance of overlap between neutrophils and γδ T cells137,138,139,140,141, suggesting that IDO-Gal3 treatment has limited affect exacerbating disease on the genotypic level. Overall, as there were fewer genes up-regulated, there were also fewer up-regulated genes involved in recruitment and activation for either cell type of interest107,108,109,110,111,112,142,143,144,145,146,147,148,149,150,151,152,153,154, further demonstrating the minimal effect of IDO-Gal3 on worsening disease pathology. Roughly half of the neutrophil recruitment and activation genes identified were up-regulated at day 7 and later down-regulated at day 14, suggesting a reversal in neutrophil recruitment with IDO-Gal3 treatment. Fewer up-regulated genes were identified as immunosuppressive towards neutrophils and γδ T cells55,138,155,156,157,158,159,160,161,162,163, though there was more overlap seen between the two populations than within the down-regulated immunosuppressive genes. These findings are consistent with the decrease in neutrophil and γδ T cell populations seen at day 7 (Fig. 3), and suggest that the immunomodulatory effect of IDO-Gal3 may be due to a reduction in the recruitment of pathogenic immune cells.

Genes and signaling pathways with potential disease ameliorating effect are on the left, while those with potential disease exacerbating effect are on the right, with down-regulated genes on the top (blue circles) and up-regulated genes on the bottom (red circles). Text color reports day and detection method of gene expression, with Nanostring d7 only shown in pink, bulk RNA-seq d14 only shown in purple, and both metrics and time points shown in blue.
Expression of myeloid cytokines in psoriatic tissues pivots mid-model following injection with IDO-Gal3
Multiplex cytokine and chemokine protein expression levels of immune system markers were measured from tissue lysates harvested on day 7 (Fig. 7). Comparison of PBS and IDO-Gal3 groups revealed significant decrease expression of key cytokines associated with IMQ-induced psoriasis pathology. Consistent with gene expression data at day 7 and day 14, protein concentration of TNFα and macrophage inflammatory protein-2 (MIP-2) were significantly decreased in IDO-Gal3 treated skin lysate. In contrast, IL-10 protein concentration was unaltered, consistent with our previous finding that IDO-Gal3 maintains IL-10 at homeostatic basal levels28. These data represent that the anti-inflammatory effect of IDO-Gal3 treatment persists at both the genotypic and phenotypic level within the tissue. Assessment of number of other cytokines in the panel, however, were unable to detect differences between groups (Supplementary Fig. 4). Of note, differences were not detectable of IL-17A, a key product of the IL-23/Th17 axis driving psoriatic pathophysiology164, nor IL-23, an early driver of psoriasis33, despite the down-regulation of IL-17 signaling pathway transcriptomics seen at day 14 and the reduction of clinical scores.

Bar graphs representing multiplex cytokine and chemokine measurement from psoriatic skin tissue lysates collected mid-model (day 7). Concentrations observed in PBS treated (black circles) and IDO-Gal3 treated psoriatic (IDO-Gal3, red triangles) tissues for tumor necrosis factor-α (TNFα), macrophage inflammatory protein-2 (MIP-2 aka CXCL2), and interleukin-10 (IL-10). Data are mean ± SD; n = 7/group; concentrations compared using student’s t-test; *P < 0.05, ns not significant.
Discussion
This work presents compelling evidence of IDO-Gal3 halting and reversing presentation of disease severity in the pre-clinical IMQ-induced mouse model of psoriasis. IDO-Gal3 apparently reduced the inflammatory immune cell population and activation within both the epidermis and the dermis of the skin, and through changes in both the myeloid and lymphoid pathogenic immune cell population, throughout the course of disease. As neutrophils and macrophages both drive psoriasis pathology, decreased general myeloid and neutrophil specific markers in the epidermis suggest that this may be key to the improvement in clinical scores from IDO-Gal3 treatment. Reduction in myeloid cells at both the midpoint and endpoint of disease indicates a lasting effect of IDO-Gal3 on immune cell populations. Additionally, as NETosis is associated with increased psoriatic inflammation15, the data suggests not only the decreased number of neutrophils but also their decreased inflammatory behavior contributes to the therapeutic effect, indicating a potential mechanism of action. However, there was no significant changes in GM-CSF protein expression seen at day 7, which is contrary to what is expected, as GM-CSF stimulates neutrophil extravasation and localization to ligand-bearing sites in psoriatic skin165,166, potentially due to the timing of cytokine measurement within the scope of disease. Furthermore, we observed decreased levels of the chemokine MIP-2 at the midpoint of disease. MIP-2, also known as CXCL2, is one of the antimicrobial chemokines expressed by keratinocytes specifically for neutrophil recruitment167, pointing to a modulation of the cellular microenvironment not limited only to immune cell populations, which could prove advantageous given the keratinocyte ability to recruit and engage immune cells. Although not significantly different, the remaining cytokine and chemokine levels measured via ELISA show potential trends that may hint at the influence of IDO-Gal3 on protein expression. Lack of significant difference in target molecules in the panel may be due to the timing at which samples were harvested, or the discrepancies often observed between measurements of transcript expression and protein measurements168. Of note, both IL-23 and IL-17 genotypic expression levels peak by 72 h after initial imiquimod application29, suggesting that the midpoint of disease may be too late to detect notable changes in these cytokine expression levels. Future studies may benefit from earlier cytokine measurement, such as at the peak of disease on day 5, and by identifying up-stream and down-stream proteins throughout signaling pathways from gene expression data and subsequent protein level measurement based on transcriptional differences.
Within the lymphoid population of the dermis, IDO-Gal3 treatment resulted in similar reduction of pathogenic T cells, most notably within the γδ T cell population. As the expansion and activation of TCRγδ cells is a common feature of psoriasis169 and IL-23 predominantly stimulates γδ T cells to produce IL-17 and increase psoriasis disease progression33,170,171, the reduced activation of γδ T cells observed after IDO-Gal3 treatment implicates involvement of this mechanism of action. Furthermore, the observed reduction in CCR6+ γδ T cell populations is of interest, as CCR6+ γδ T cells have been shown to produce IL-17172,173, CCR6 is required for epidermal trafficking of pathogenic γδ T cells in the skin32,174. In contrast, in the epidermis, detection of differences in γδ T cell populations was not anticipated as CCR6+ and RORγt + γδ T in the epidermis is described as being low in both healthy and diseased tissue33,170. The decreased number of both activated myeloid and lymphoid cells of the dermis suggests a two-pronged anti-inflammatory mechanism of IDO-Gal3.
A single administration of IDO-Gal3 at the day 3 onset of disease decreases expression of key inflammatory pathways implicated in IMQ-induced psoriasis, regardless of daily administration of IMQ. Hyper-proliferation and hyper-keratinization are phenotypic hallmarks of IMQ-induced psoriasis, and the day 14 transcriptional down-regulation of pathways promoting cell turnover (FoxO signaling, mTOR signaling) is consistent with the observed decrease in skin scaling and thickening in the IDO-Gal3 treated animals175. Down-regulation of TNF signaling demonstrated the robust and long-lasting impact of IDO-Gal3 on immune and inflammatory responses, as IMQ results in significantly increased TNF production in the skin29,176. Pathways up-regulated at day 14 indicated metabolic reprogramming and suggest a shift toward homeostasis. Oxidative phosphorylation (OXPHOS), occurs in the mitochondria and is associated with suppressive and tolerogenic immune cell subtypes, such as M2 macrophages and regulatory T cells44,45,177. Interestingly, up-regulated mitophagy-associated pathways indicate IDO-Gal3 treatment may counteract psoriasis-associated malfunctions in mitophagy46,178,179, requiring further investigation.
Within the macrophage polarization-related gene set, pro-inflammatory “M1”-like genes were down-regulated, with Tgfb1 as the only gene increased by IDO-Gal3. Tgfb1 is a pleotropic factor implicated in M2 macrophage skewing and suppression of inflammation, but is also overexpressed in psoriasis47,48, associated with its ability to stimulate ECM protein production and promote fibrosis. Along this line, up-regulation of AGE-RAGE signaling is involved in oxidative stress responses contributing to psoriasis75. However, in the context of the observed down-regulation of NFκB, the AGE-RAGE-associated signaling pathways associated with cell cycle and vascular remodeling may serve a larger role. Indeed, there is cross-talk between the two pathways which can result in ECM regulation and turnover180. The convergence in these pathways may relate to the up-regulation of ECM and adhesion molecule gene expression observed at disease midpoint. Additionally, ECM components may have immunomodulatory roles in psoriasis, not fully elucidated181. Among fibrosis-related genes associated with inflammation, IDO-Gal3 treatment decreased expression of genes relating to the recruitment53,54 and migration52 of pathogenic cells and increased expression of genes that, which, when deficient, result in increased psoriatic inflammation56.
At the disease midpoint, IDO-Gal3 treatment evidenced overall trends of decreasing expression of psoriatic genes and pathways, similar to that seen at the endpoint of disease. However, while some genes of interest were down-regulated at both day 7 and day 14, a number of genes varied in their expression levels between the two time points, suggesting that IDO-Gal3’s therapeutic effect is dynamic over time, resulting in genotypic changes that vary with the stage of disease treatment. Reduction of one key pro-inflammatory cytokine, TNFα, stayed consistent across time points, with the TNF signaling pathway down-regulated at day 7 and day 14. Our genotypic findings were confirmed with phenotypic expression, as TNFα levels were decreased within psoriatic skin tissue at day 7 after IDO-Gal3 treatment, indicating the widespread protection against inflammatory molecules by IDO-Gal3 during immune response. TNFα, a pleotropic pro-inflammatory cytokine, was the first clinical target for psoriasis monoclonal antibody therapy20,21, and decreased TNFα in response to IDO-Gal3 implicates IDO-Gal3 as a robust intervention for psoriasis.
Within the genes and signaling pathways identified, many affect the recruitment and activation or immunosuppression of two pathogenic cell populations, neutrophils and γδ T cells. Overall, there were more genes and pathways of interest down-regulated at both day 7 and day 14 than up-regulated, and within those populations, most of the down-regulated genes were associated with recruitment and activation, rather than immunosuppression, as expected from additional work showing the reduction in neutrophil and γδ T cell numbers at disease midpoint. The relatively modest number of up-regulated genes and signaling pathways involved in immunosuppression may further correlate to our immune cell population findings, as we saw a decrease in activated pathogenic cell populations, rather than an increase in immunosuppressive regulatory T cell populations. Additionally, several of the genes identified in the up-regulated population at day 7 were down-regulated by day 14, demonstrating the dynamic effect of IDO-Gal3 on psoriatic skin.
In conclusion, this study has revealed new insight into the immunological role of IDO-Gal3 mediating anti-inflammatory processes within psoriatic tissues. The sustained inflammation suppression by IDO-Gal3 with only a one-time application is intriguing given the expectation of approximately one week of residence in situ following injection28, consistent with the notion of IDO as a general regulator of inflammation, as well as impacting psoriasis fibrotic programs. These data highlight the potential of IDO-Gal3 as a powerful therapeutic protein for psoriasis, acknowledging limitations such as the use of an acute model to extrapolate to a chronic inflammatory disease, as well as differences in cell populations and transcriptome between the IMQ mouse model and human plaque psoriasis182.
Methods
Mice and treatments
All experiments were carried out in 8-12-week-old female C57BL/6j (The Jackson Laboratory) mice in accordance with the Institutional Animal Care and Use Committee at the University of Florida. Animals were housed in standard ventilated SPF conditions and followed a standard light-dark cycle.
Imiquimod-induced psoriasis and scoring
The model used was a modified version of that reported by van der Fits et al.29. The model has been reproduced in the current study, with two main differences. The current study was carried out in C57BL6/j (B6) mice, as opposed to BALB/c, due to the fact that human plaque psoriasis is more closely recapitulated in the B6 mouse strain183. The second difference is in the length of the model. The current study follows a two-week model schedule, as opposed to a one-week schedule, to capture both the acute and chronic phases of the model184. Briefly, mice were anesthetized and rendered unconscious with isoflurane and their backs were shaved, followed by application of depilatory cream to remove any remaining fur. Each day for 14 days total, 5% IMQ cream (62.5 mg, Patterson Veterinary Supply, cat. num. 07-893-7787) was applied to the backs of the mice. IMQ was applied to anesthetized mice so that movement, grooming, and social behavior did not spread IMQ to distal areas. On the 3rd day of IMQ application, anesthetized mice were subcutaneously injected with five doses of IDO-Gal3 (10 μg) in sterile saline spread evenly throughout the back, or a sterile saline control (n = 12 per group). Disease severity was measured each day using a modified version of the Psoriasis Area and Severity Index (PASI) where area of effect is not considered. Erythema (redness), scaling, and thickening were scored independently and assigned a score on a scale of 0 to 4: 0, none, 1: slight, 2: moderate, 3: marked, 4: very marked. The cumulative score was reported as a measure of the severity of inflammation (scale 0–12). On the 14th day of the experiment, mice were euthanized by inhalation of 100% carbon dioxide, followed by cervical dislocation, and skin was collected for analysis.
IDO-Gal3 expression and purification
Protein was expressed and purified as described previously27,28. Purity was determined by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Coomassie staining (Supplementary Fig. 1a). Tryptophan catabolism was confirmed by incubating IDO with tryptophan and electron donors (methylene blue and ascorbic acid) and assessing the slope of the absorbance at 321 nm (Supplementary Fig. 1b). Affinity of IDO-Gal3 for lactose was determined using affinity chromatography in an AKTA Pure chromatography system (GE Life Sciences) equipped with consumer-packable glass column (GE Life Sciences) packed with α-lactose agarose affinity resin (Sigma-Aldrich). Proteins were eluted with a linear gradient of β-lactose (Sigma-Aldrich) in phosphate buffer (Supplementary Fig. 1c). Endotoxin content at concentrations used for in vivo experiments was reduced below 1.0 EU·mL-1, the maximum acceptable allowable dose for pre-clinical drugs185 using endotoxin removal solution (Millipore-Sigma). Endotoxin level was determined with Chromo-LAL and compared against Control Standard Endotoxin (Associates of Cape Cod, Inc.) at known concentrations, according to the manufacturer’s instructions.
Histology
After dissection skin samples were fixed overnight at 4 °C in 10% neutral buffered formalin. Following fixation, samples were dehydrated, cleared, and embedded in paraffin according to standard lab technique. Tissues were sectioned into slices (5 µm thick) and stained with hematoxylin and eosin (Vector Laboratories). Slides were scanned for high resolution images with an Olympus VS200 Whole Slide Scanner (EvidentScientific) and analyzed using QuPath and ImageJ.
Multiplex immune panel
Formalin-fixed and paraffin-embedded (FFPE) tissue samples from day 14 were immunostained using the OPAL TM 7-Color Automation IHC kit (AKOAYA Biosciences) on the BOND RX autostainer (Leica Biosystems). The OPAL 7-color kit uses tyramide signal amplification (TSA)-conjugated to individual fluorophores to detect various targets within the multiplex assay. Sections were baked at 65 °C for one hour, then transferred to the BOND RX (Leica Biosystems). All subsequent steps (ex., deparaffinization, antigen retrieval) were performed using an automated OPAL IHC procedure (AKOYA Biosciences). OPAL staining of each antigen occurred as follows: heat induced epitope retrieval (HIER) was achieved with EDTA pH 9.0 buffer for 20 min at 95 °C before the slides were blocked with AKOYA blocking buffer for 10 min. Slides were then incubated with primary antibody at RT for 30 min followed by OPAL HRP polymer and one of the OPAL fluorophores during the final TSA step. Staining used the following antibodies: Ly-6G (Cell Signaling Technologies, cat. no. 87048 s, E6Z1T, 1:100), CD86 (Cell Signaling Technologies, cat. no. 19589 s, 1:150), Citrullinated histone 3 (Cell Signaling Technologies, cat. no. 97272 s, (Arg17) (E4O3F), 1:500), CD8 (Cell Signaling Technologies, cat. no. 98941 s, D4W2Z, 1:50), CD32 (Abcam, ab28364, 1:200), CD11c (Abcam, ab219799, EPR21826, 1:50), and CD11b (Bio SB, cat. no. BSB6440, EP45, 1:200). Individual antibody complexes are stripped after each round of antigen detection. This was repeated five more times using the remaining antibodies on the panel. After the final stripping step, DAPI counterstain is applied to the multiplexed slide and is removed from BOND RX for coverslipping with ProLong Diamond Antifade Mountant (ThermoFisher Scientific). Autofluorescence slides (negative control) were included, which use primary and secondary antibodies omitting the OPAL fluors and DAPI. All slides were imaged with the Vectra®3 Automated Quantitative Pathology Imaging System. Cell population attributions were considered as follows. While integrin subunit CD11b is a marker shared among multiple immune cells, in the skin CD11b+ macrophages are typically the most abundant immune cell population186. Ly-6G is exclusively expressed by granulocytes187 and served as a neutrophil marker. The presence of citrullinated histone H3 (H3Cit), is evidence of NET formation188,189.
Quantitative image analysis
Multi-layer TIFF images are exported from InForm (AKOYA Biosciences) and loaded into HALO Image Analysis Platform version 3.6.4134 (Indica Labs) for quantitative image analysis. For the quantitative fluorescent phenotype analysis, the tissue is segmented into individual cells using the DAPI marker which stains cell nuclei. For each marker, a positivity threshold within the nucleus or cytoplasm is determined per marker based on visual intensity. After setting a positive fluorescent threshold for each staining marker, the entire image set is analyzed with the created algorithm. The generated data includes positive cell counts for each fluorescent marker in cytoplasm or nucleus, and percent of cells positive for the marker. Along with the summary output, a per-cell analysis can be exported to provide the marker status, classification, and fluorescent intensities for downstream bioinformatics analysis.
Flow cytometry
Skin was harvested at the midpoint of disease, day 7. Following the protocol adapted from Lou et al. 190, 1 cm2 pieces of skin were quartered and digested in Dispase II (5 mg mL-1, Sigma, cat. D4693-1G) for 1 hour at 37 °C. Dermis and epidermis were separated, and dermis layers were digested in Collagenase P (1 mg mL−1, Sigma, cat. 11213857001) and DNAse I (1 mg mL−1, Sigma, cat. 10104159001) for 80 minutes at 37 °C. Epidermis layers were further dissected and homogenized using a gentleMACS Dissociator (Miltenyi Biotec) before digestion in 0.05% Trypsin-EDTA for 10 minutes. Following digestion, cells were counted and pretreated with TruStain FcX (anti-mouse CD16/32, Biolegend). Cells were stained against CD45.2 [104, AF700, Biolegend], CD11b [M1/70, BV510, Biolegend], CD11c [N4180, BV711, Biolegend], MHCII [M5/114.15.2, BUV496, eBiosciences], CD86 [GL-1, BUV737, eBiosciences], F4/80 [BM8, FITC, Biolegend], Gr-1 [RB6-8C5, BUV395, eBiosciences], iNOS2 [CXNFT, PE-Cy7, eBiosciences], Arginase [A1ex5, APC, eBiosciences], TCRb [H57-597, eF450, eBiosciences], TCRγδ [GL3, PE, Biolegend], CD4 [RM4-5, SuperBright 780, eBiosciences], CD8α [53-6.7, BV605, eBiosciences], CD44 [IM7, BV650, Biolegend], CXCR3 [CXCR3-173, NovaFluor Blue 660-120S, eBiosciences], CCR6 [29-2L17, PE/Fire 810, Biolegend], Tbet [4B10, PE-Cy5, eBiosciences], Rorγt [Q31-378, PerCP-Cy5.5, BD Biosciences], and FoxP3 [MF23, PE-cf594, BD Biosciences]. All measurements were done on a five laser Aurora (Cytek Biosciences) and analyzed using FlowJo software (BD Biosciences).
RNA sequencing
Following dissection, day 14 skin samples were stored in RNAlater (Invitrogen) at 4 °C until the time of RNA extraction. Total RNA was extracted from the affected skin of the back using the RNeasy Lipid Tissue Mini Kit (QIAGEN). RNA concentration via Qubit and purity was measured via Nanodrop Spectrophotometer (Thermo Fisher Scientific). Library preparation and sequencing were carried out by GENEWIZ on the NovaSeq 6000 platform. FASTQ files from each time-point were trimmed using Seq TK191 and aligned to Mus musculus GRCm38 (ENSEMBL) reference genome with the Hisat2192 alignment method. Gene counts were obtained using featureCounts193 and differential expression was determined using DESeq2194. Differentially expressed genes (DEGs) were identified with the threshold of P value adjusted by false discovery rate (Padj) < 0.05. KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway enrichment analysis on DEGs was performed using g:GOst (https://biit.cs.ut.ee/gprofiler/gost). The items with P-value adjusted by g:SCS (Padj) < 0.01 were identified as significantly enriched KEGG pathways.
Nanostring
NanoString technology and the nCounter Mouse Pan Cancer Immunology Panel was used to simultaneous evaluate 770 immune and metabolic-related mRNAs in each day 7 skin sample57. Three biological replicates were used for each sample. Briefly, a total of 100 ng mRNA was hybridized to report-capture probe pairs (CodeSets) at 65 °C for 18 h. After this solution-phase hybridization, the nCounter Prep Station was used to remove excess probe, align the probe/target complexes, and immobilize these complexes in the nCounter cartridge. The nCounter™ cartridge was then placed in a digital analyzer for image acquisition and data processing. Color codes designating mRNA targets of interest were directly imaged on the surface of the cartridge. The expression level of each gene was measured by counting the number of times the color-coded barcode for that gene was detected, and the barcode counts tabulated. nSolver v4.0, an integrated analysis platform was used to generate appropriate data normalization as well as fold-changes, resulting ratios and differential expression57. Differentially expressed immune genes (immune DEGs) were identified with the threshold of P < 0.05.
Multiplex cytokine detection
Following dissection, day 14 skin samples were frozen at −80 °C until the time of homogenization. Samples were dissected into pieces approximately 2 mm × 2 mm in area a placed in tubes containing 800 μm glass beads and NP-40 lysis buffer supplemented with Halt Protease Inhibitor (Thermo Fisher Scientific). Tissues were homogenized using a Mini bead beater. Following bead beating samples were spun at 10,000 G for 10 min 4 °C, and supernatant was collected. Protein concentration was determined using BCA assay (Thermo Fisher Scientific) and samples were normalized to 4 mg·mL−1. Subsequent multiplex protein detection was completed by Eve Technologies, via a Luminex 200 (Thermo Fisher Scientific) and the MILLIPLEX Mouse High Sensitivity T-Cell 18-Plex Discovery Assay (Thermo Fisher Scientific). Additional cytokine detection for IL-23 was performed using the Mouse IL-23 DuoSet ELISA (Bio-techne).
Statistics of measurements
Clinical scores were compared using a Kruskal-Wallis test with multiple comparisons. Area under the curve values and epidermal thickness measurements were compared using One-way ANOVA with Tukey’s multiple comparisons test. Multiplex cytokine and chemokine data were compared using student’s t-test. Outliers were removed using Grubbs test for outliers (α = 0.05) when appropriate. P < 0.05 are considered as significant. In all figures; *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001, ns: not significant.
Data availability
The RNA-seq data have been deposited in the National Center for Biotechnology Information (NCBI) database with the accession number of PRJNA1227466.
References
-
Armstrong, A. W. & Read, C. Pathophysiology, clinical presentation, and treatment of psoriasis: A review. JAMA 323, 1945–1960 (2020).
-
Rachakonda, T. D., Schupp, C. W. & Armstrong, A. W. Psoriasis prevalence among adults in the United States. J. Am. Acad. Dermatol. 70, 512–516 (2014).
-
Vanderpuye-Orgle, J. et al. Evaluating the economic burden of psoriasis in the United States. J. Am. Acad. Dermatol. 72, 961–7.e5 (2015).
-
Rendon, A. & Schäkel, K. Psoriasis pathogenesis and treatment. Int. J. Mol. Sci. 20, 1475 (2019).
-
Blauvelt, A. New concepts in the pathogenesis and treatment of psoriasis: key roles for IL-23, IL-17A and TGF-β1. Expert Rev. Dermatol. 2, 69–78 (2007).
-
Matsuzaki, G. & Umemura, M. Interleukin-17 family cytokines in protective immunity against infections: role of hematopoietic cell-derived and non-hematopoietic cell-derived interleukin-17s. Microbiol. Immunol. 62, 1–13 (2018).
-
Mylonas, A. & Conrad, C. Psoriasis: Classical vs. Paradoxical. The Yin-Yang of TNF and Type I Interferon. Front. Immunol. 9, 2746 (2018).
-
Austin, L. M., Ozawa, M., Kikuchi, T., Walters, I. B. & Krueger, J. G. The majority of epidermal T cells in Psoriasis vulgaris lesions can produce type 1 cytokines, interferon-gamma, interleukin-2, and tumor necrosis factor-alpha, defining TC1 (cytotoxic T lymphocyte) and TH1 effector populations: a type 1 differentiation bias is also measured in circulating blood T cells in psoriatic patients. J. Invest. Dermatol. 113, 752–759 (1999).
-
Aggarwal, S., Ghilardi, N., Xie, M.-H., de Sauvage, F. J. & Gurney, A. L. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 278, 1910–1914 (2003).
-
Teunissen, M. B., Koomen, C. W., de Waal Malefyt, R., Wierenga, E. A. & Bos, J. D. Interleukin-17 and interferon-gamma synergize in the enhancement of proinflammatory cytokine production by human keratinocytes. J. Invest. Dermatol. 111, 645–649 (1998).
-
Lee, E. et al. Increased expression of interleukin 23 p19 and p40 in lesional skin of patients with psoriasis vulgaris. J. Exp. Med. 199, 125–130 (2004).
-
Eyerich, S. et al. Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling. J. Clin. Invest. 119, 3573–3585 (2009).
-
Terui, T., Ozawa, M. & Tagami, H. Role of neutrophils in induction of acute inflammation in T-cell-mediated immune dermatosis, psoriasis: a neutrophil-associated inflammation-boosting loop. Exp. Dermatol. 9, 1–10 (2000).
-
Meng, X. et al. RIPK1 downregulation enhances neutrophil extracellular traps in psoriasis. Postepy Dermatol. Alergol. 39, 72–80 (2022).
-
Zhang, J., Feng, Y. & Shi, D. NETosis of psoriasis: a critical step in amplifying the inflammatory response. Front. Immunol. 15, 1374934 (2024).
-
Herster, F. et al. Neutrophil extracellular trap-associated RNA and LL37 enable self-amplifying inflammation in psoriasis. Nat. Commun. 11, 105 (2020).
-
Clark, R. A. & Kupper, T. S. Misbehaving macrophages in the pathogenesis of psoriasis. J. Clin. Invest. 116, 2084–2087 (2006).
-
Lin, S.-H., Chuang, H.-Y., Ho, J.-C., Lee, C.-H. & Hsiao, C.-C. Treatment with TNF-α inhibitor rectifies M1 macrophage polarization from blood CD14+ monocytes in patients with psoriasis independent of STAT1 and IRF-1 activation. J. Dermatol. Sci. 91, 276–284 (2018).
-
Feyen, O. et al. Off-target activity of TNF-alpha inhibitors characterized by protein biochips. Anal. Bioanal. Chem. 391, 1713–1720 (2008).
-
Downey, C. Serious infection during etanercept, infliximab and adalimumab therapy for rheumatoid arthritis: A literature review. Int. J. Rheum. Dis. 19, 536–550 (2016).
-
Minozzi, S. et al. Risk of infections using anti-TNF agents in rheumatoid arthritis, psoriatic arthritis, and ankylosing spondylitis: a systematic review and meta-analysis. Expert Opin. Drug Saf 15, 11–34 (2016).
-
Munn, D. H. et al. Prevention of allogeneic fetal rejection by tryptophan catabolism. Science 281, 1191–1193 (1998).
-
Llamas-Velasco, M. et al. Immune cells from patients with psoriasis are defective in inducing indoleamine 2,3-dioxygenase expression in response to inflammatory stimuli. Br. J. Dermatol. 176, 695–704 (2017).
-
Elizei, S. S. et al. IDO-expressing Fibroblasts Suppress the Development of Imiquimod-induced Psoriasis-like Dermatitis. Cell Transplant 27, 557–570 (2018).
-
Clement, C. C. et al. 3-hydroxy-L-kynurenamine is an immunomodulatory biogenic amine. Nat. Commun. 12, 4447 (2021).
-
Bracho-Sanchez, E., Hassanzadeh, A., Brusko, M. A., Wallet, M. A. & Keselowsky, B. G. Dendritic Cells Treated with Exogenous Indoleamine 2,3-Dioxygenase Maintain an Immature Phenotype and Suppress Antigen-specific T cell Proliferation. J. Immun. Regen. Med. 5, 100015 (2019).
-
Farhadi, S. A. et al. Locally anchoring enzymes to tissues via extracellular glycan recognition. Nat. Commun. 9, 4943 (2018).
-
Bracho-Sanchez, E. et al. Suppression of local inflammation via galectin-anchored indoleamine 2,3-dioxygenase. Nat. Biomed. Eng. 7, 1156–1169 (2023).
-
van der Fits, L. et al. Imiquimod-induced psoriasis-like skin inflammation in mice is mediated via the IL-23/IL-17 axis. J. Immunol. 182, 5836–5845 (2009).
-
Reichrath, J. et al. Expression of integrin subunits and CD44 isoforms in psoriatic skin and effects of topical calcitriol application. J. Cutan. Pathol. 24, 499–506 (1997).
-
Lugović-Mihić, L., Novak-Bilić, G., Vučić, M., Japundžić, I. & Bukvić, I. CD44 expression in human skin: High expression in irritant and allergic contact dermatitis and moderate expression in psoriasis lesions in comparison with healthy controls. Contact Derm 82, 297–306 (2020).
-
Mabuchi, T. et al. CCR6 is required for epidermal trafficking of γδ-T cells in an IL-23-induced model of psoriasiform dermatitis. J. Invest. Dermatol. 133, 164–171 (2013).
-
Cai, Y. et al. Pivotal role of dermal IL-17-producing γδ T cells in skin inflammation. Immunity 35, 596–610 (2011).
-
Martin, B., Hirota, K., Cua, D. J., Stockinger, B. & Veldhoen, M. Interleukin-17-producing gammadelta T cells selectively expand in response to pathogen products and environmental signals. Immunity 31, 321–330 (2009).
-
Condamine, T. et al. Characterization of Schlafen-3 expression in effector and regulatory T cells. J. Leukoc. Biol. 87, 451–456 (2010).
-
Girolomoni, G., Mrowietz, U. & Paul, C. Psoriasis: rationale for targeting interleukin-17. Br. J. Dermatol. 167, 717–724 (2012).
-
Li, B. et al. The role of Th17 cells in psoriasis. Immunol. Res. 68, 296–309 (2020).
-
Kawasaki, T. & Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 5, 461 (2014).
-
Jang, D.-I. et al. The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. Int. J. Mol. Sci. 22, 2719 (2021).
-
Shah, K., Al-Haidari, A., Sun, J. & Kazi, J. U. T cell receptor (TCR) signaling in health and disease. Signal Transduct. Target. Ther. 6, 412 (2021).
-
Kim, M. E., Kim, D. H., & Lee, J. S. FoxO Transcription Factors: Applicability as a Novel Immune Cell Regulators and Therapeutic Targets in Oxidative Stress-Related Diseases. Int. J. Mol. Sci. 23, 11877 (2022).
-
Perl, A. mTOR activation is a biomarker and a central pathway to autoimmune disorders, cancer, obesity, and aging. Ann. N.Y. Acad. Sci. 1346, 33–44 (2015).
-
Suto, T. & Karonitsch, T. The immunobiology of mTOR in autoimmunity. J. Autoimmun. 110, 102373 (2020).
-
Cibrian, D., de la Fuente, H. & Sánchez-Madrid, F. Metabolic pathways that control skin homeostasis and inflammation. Trends Mol. Med. 26, 975–986 (2020).
-
O’Neill, L. A. J., Kishton, R. J. & Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 16, 553–565 (2016).
-
Xu, Y., Shen, J. & Ran, Z. Emerging views of mitophagy in immunity and autoimmune diseases. Autophagy 16, 3–17 (2020).
-
Baran, W., Szepietowski, J. C., Mazur, G. & Baran, E. TGF-beta(1) gene polymorphism in psoriasis vulgaris. Cytokine 38, 8–11 (2007).
-
Han, G. et al. A role for TGFbeta signaling in the pathogenesis of psoriasis. J. Invest. Dermatol. 130, 371–377 (2010).
-
Maybury, C. M. et al. Prevalence of advanced liver fibrosis in patients with severe psoriasis. JAMA Dermatol 155, 1028–1032 (2019).
-
Famularo, G. et al. Retroperitoneal fibrosis associated with psoriasis: a case series. Scand. J. Rheumatol. 38, 68–69 (2009).
-
Gu, C. et al. Identification of common genes and pathways in eight fibrosis diseases. Front. Genet. 11, 627396 (2020).
-
Köchl, R. et al. WNK1 kinase balances T cell adhesion versus migration in vivo. Nat. Immunol. 17, 1075–1083 (2016).
-
Behfar, S., Hassanshahi, G., Nazari, A. & Khorramdelazad, H. A brief look at the role of monocyte chemoattractant protein-1 (CCL2) in the pathophysiology of psoriasis. Cytokine 110, 226–231 (2018).
-
Vestergaard, C., Just, H., Baumgartner Nielsen, J., Thestrup-Pedersen, K. & Deleuran, M. Expression of CCR2 on monocytes and macrophages in chronically inflamed skin in atopic dermatitis and psoriasis. Acta Derm. Venereol. 84, 353–358 (2004).
-
Michaels, A. J., Campbell, C., Bou-Puerto, R. & Rudensky, A. Y. Nuclear receptor LXRβ controls fitness and functionality of activated T cells. J. Exp. Med. 218, e20201311 (2021).
-
Nakao, M., Miyagaki, T., Sugaya, M. & Sato, S. Exacerbated Imiquimod-Induced Psoriasis-Like Skin Inflammation in IRF5-Deficient Mice. Int. J. Mol. Sci. 21, 3681 (2020).
-
Kulkarni, M. M. Digital multiplexed gene expression analysis using the NanoString nCounter system. Curr. Protoc. Mol. Biol. Chapter 25, Unit25B.10.1-25B.10.17 (2011).
-
Li, D. et al. CD200-CD200R1 signalling attenuates imiquimod-induced psoriatic inflammation by inhibiting the activation of skin inflammatory macrophages. Int. Immunopharmacol. 78, 106046 (2020).
-
Zheng, Q.-Y. et al. Complement component 3 deficiency prolongs MHC-II disparate skin allograft survival by increasing the CD4( + ) CD25(+) regulatory T cells population. Sci. Rep. 6, 33489 (2016).
-
Zheng, Q.-Y. et al. Complement component 3 prevents imiquimod-induced psoriatic skin inflammation by inhibiting apoptosis in mice. Int. Immunopharmacol. 85, 106692 (2020).
-
Gabryšová, L. et al. c-Maf controls immune responses by regulating disease-specific gene networks and repressing IL-2 in CD4 + T cells. Nat. Immunol. 19, 497–507 (2018).
-
Zhang, Y. et al. Immune cell infiltration analysis demonstrates excessive mast cell activation in psoriasis. Front. Immunol. 12, 773280 (2021).
-
Meyer, A., Parmar, P. J. & Shahrara, S. Significance of IL-7 and IL-7R in RA and autoimmunity. Autoimmun. Rev. 21, 103120 (2022).
-
Kaneko, N., Kurata, M., Yamamoto, T., Morikawa, S. & Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 39, 12 (2019).
-
Cancino-Díaz, J. C. et al. Interleukin-13 receptor in psoriatic keratinocytes: overexpression of the mRNA and underexpression of the protein. J. Invest. Dermatol. 119, 1114–1120 (2002).
-
Voglis, S. et al. Regulation of IL-22BP in psoriasis. Sci. Rep. 8, 5085 (2018).
-
Wawrzycki, B. et al. Interleukin-22 and Its Correlation with Disease Activity in Plaque Psoriasis. Arch Immunol Ther Exp (Warsz) 67, 103–108 (2019).
-
Elnabawi, Y. A. et al. CCL20 in psoriasis: A potential biomarker of disease severity, inflammation, and impaired vascular health. J. Am. Acad. Dermatol. 84, 913–920 (2021).
-
Liu, T., Zhang, L., Joo, D. & Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2, 17023 (2017).
-
Goldminz, A. M., Au, S. C., Kim, N., Gottlieb, A. B. & Lizzul, P. F. NF-κB: an essential transcription factor in psoriasis. J. Dermatol. Sci 69, 89–94 (2013).
-
Kojima, H. et al. An essential role for NF-kappa B in IL-18-induced IFN-gamma expression in KG-1 cells. J. Immunol. 162, 5063–5069 (1999).
-
Wang, X. et al. Interleukin-18 and IL-18BP in inflammatory dermatological diseases. Front. Immunol. 14, 955369 (2023).
-
Mosca, M. et al. The Role of IL-17 Cytokines in Psoriasis. Immunotargets Ther 10, 409–418 (2021).
-
Holbrook, J., Lara-Reyna, S., Jarosz-Griffiths, H. & McDermott, M. Tumour necrosis factor signalling in health and disease. F1000Res. 8, 1–12 (2019).
-
Pleńkowska, J., Gabig-Cimińska, M. & Mozolewski, P. Oxidative stress as an important contributor to the pathogenesis of psoriasis. Int. J. Mol. Sci. 21, 6206 (2020).
-
Peng, Y. et al. AGE-RAGE signal generates a specific NF-κB RelA “barcode” that directs collagen I expression. Sci. Rep. 6, 18822 (2016).
-
Yano, T. et al. RAGE-dependent NF-kB inflammation processes in the capsule of frozen shoulders. J. Shoulder Elbow Surg 29, 1884–1891 (2020).
-
Nie, Y. et al. Nfatc3 promotes pulmonary inflammation and fibrosis by regulating production of CCL2 and CXCL2 in macrophages. Aging Dis 14, 1441–1457 (2023).
-
Mi, Q.-S. et al. Dysregulated CD38 expression in blood and skin immune cells of patients with hidradenitis suppurativa. Res. Sq. https://doi.org/10.21203/rs.3.rs-2609421/v1 (2023).
-
Li, W. et al. CD38: A significant regulator of macrophage function. Front. Oncol. 12, 775649 (2022).
-
Nguyen, C. T. H. et al. Up-regulated expression of CD86 on circulating intermediate monocytes correlated with disease severity in psoriasis. J. Dermatol. Sci. 90, 135–143 (2018).
-
Grossman, R. M. et al. Interleukin 6 is expressed in high levels in psoriatic skin and stimulates proliferation of cultured human keratinocytes. Proc Natl Acad Sci USA 86, 6367–6371 (1989).
-
Johansen, C. et al. STAT2 is involved in the pathogenesis of psoriasis by promoting CXCL11 and CCL5 production by keratinocytes. PLoS ONE 12, e0176994 (2017).
-
Shi, Z.-R. et al. Decrease of galectin-3 in keratinocytes: A potential diagnostic marker and a critical contributor to the pathogenesis of psoriasis. J. Autoimmun. 89, 30–40 (2018).
-
Kasten, K. R. et al. Interleukin-7 (IL-7) treatment accelerates neutrophil recruitment through gamma delta T-cell IL-17 production in a murine model of sepsis. Infect. Immun. 78, 4714–4722 (2010).
-
Lee, P. Y. et al. IL-1α modulates neutrophil recruitment in chronic inflammation induced by hydrocarbon oil. J. Immunol. 186, 1747–1754 (2011).
-
Yau, T. O. et al. Hyperactive neutrophil chemotaxis contributes to anti-tumor necrosis factor-α treatment resistance in inflammatory bowel disease. J. Gastroenterol. Hepatol. 37, 531–541 (2022).
-
Pavlidis, P. et al. Interleukin-22 regulates neutrophil recruitment in ulcerative colitis and is associated with resistance to ustekinumab therapy. Nat. Commun. 13, 5820 (2022).
-
Ness-Schwickerath, K. J. & Morita, C. T. Regulation and function of IL-17A- and IL-22-producing γδ T cells. Cell. Mol. Life Sci. 68, 2371–2390 (2011).
-
Lörchner, H. et al. Neutrophils for revascularization require activation of CCR6 and CCL20 by tnfα. Circ. Res. 133, 592–610 (2023).
-
Li, Z., Burns, A. R., Miller, S. B. & Smith, C. W. CCL20, γδ T cells, and IL-22 in corneal epithelial healing. FASEB J 25, 2659–2668 (2011).
-
Guo, Q. et al. NF-κB in biology and targeted therapy: new insights and translational implications. Signal Transduct. Target. Ther. 9, 53 (2024).
-
Mair, F. et al. The NFκB-inducing kinase is essential for the developmental programming of skin-resident and IL-17-producing γδ T cells. eLife 4, e10087 (2015).
-
Leung, B. P. et al. A role for IL-18 in neutrophil activation. J. Immunol. 167, 2879–2886 (2001).
-
Li, W. et al. Effect of IL-18 on expansion of gammadelta T cells stimulated by zoledronate and IL-2. J. Immunother. 33, 287–296 (2010).
-
McColl, S. R., Paquin, R., Ménard, C. & Beaulieu, A. D. Human neutrophils produce high levels of the interleukin 1 receptor antagonist in response to granulocyte/macrophage colony-stimulating factor and tumor necrosis factor alpha. J. Exp. Med. 176, 593–598 (1992).
-
Awla, D. et al. NFATc3 regulates trypsinogen activation, neutrophil recruitment, and tissue damage in acute pancreatitis in mice. Gastroenterology 143, 1352–60.e1 (2012).
-
Vieira, S. M. et al. A crucial role for TNF-alpha in mediating neutrophil influx induced by endogenously generated or exogenous chemokines, KC/CXCL1 and LIX/CXCL5. Br. J. Pharmacol. 158, 779–789 (2009).
-
Li, H., Luo, K. & Pauza, C. D. TNF-alpha is a positive regulatory factor for human Vgamma2 Vdelta2 T cells. J. Immunol. 181, 7131–7137 (2008).
-
Liu, R. et al. IL-17 Promotes Neutrophil-Mediated Immunity by Activating Microvascular Pericytes and Not Endothelium. J. Immunol. 197, 2400–2408 (2016).
-
McKenzie, D. R. et al. IL-17-producing γδ T cells switch migratory patterns between resting and activated states. Nat. Commun. 8, 15632 (2017).
-
Fan, X., Shu, P., Wang, Y., Ji, N. & Zhang, D. Interactions between neutrophils and T-helper 17 cells. Front. Immunol. 14, 1279837 (2023).
-
Jones, M. R., Simms, B. T., Lupa, M. M., Kogan, M. S. & Mizgerd, J. P. Lung NF-kappaB activation and neutrophil recruitment require IL-1 and TNF receptor signaling during pneumococcal pneumonia. J. Immunol. 175, 7530–7535 (2005).
-
Sokulsky, L. A. et al. A Critical Role for the CXCL3/CXCL5/CXCR2 Neutrophilic Chemotactic Axis in the Regulation of Type 2 Responses in a Model of Rhinoviral-Induced Asthma Exacerbation. J. Immunol. 205, 2468–2478 (2020).
-
Mayes-Hopfinger, L. et al. Chloride sensing by WNK1 regulates NLRP3 inflammasome activation and pyroptosis. Nat. Commun. 12, 4546 (2021).
-
Talbot, J. et al. CCR2 expression in neutrophils plays a critical role in their migration into the joints in rheumatoid arthritis. Arthritis Rheumatol 67, 1751–1759 (2015).
-
Meng, X. et al. FN1 Immunoregulation in Glioblastoma: Insights From Neutrophil-Centric Studies. Cancer Sci 116, 1758–1772 (2025).
-
Akbar, N. et al. Rapid neutrophil mobilization by VCAM-1+ endothelial cell-derived extracellular vesicles. Cardiovasc. Res. 119, 236–251 (2023).
-
Mohagheghpour, N., Bermudez, L. E., Khajavi, S. & Rivas, A. The VLA-4/VCAM-1 molecules participate in gamma delta cell interaction with endothelial cells. Cell. Immunol. 143, 170–182 (1992).
-
Lischke, T. et al. CD38 controls the innate immune response against Listeria monocytogenes. Infect. Immun. 81, 4091–4099 (2013).
-
Vono, M. et al. Neutrophils acquire the capacity for antigen presentation to memory CD4 + T cells in vitro and ex vivo. Blood 129, 1991–2001 (2017).
-
Snarr, B. D. et al. Galectin-3 enhances neutrophil motility and extravasation into the airways during Aspergillus fumigatus infection. PLoS Pathog 16, e1008741 (2020).
-
Prince, L. R., Whyte, M. K., Sabroe, I. & Parker, L. C. The role of TLRs in neutrophil activation. Curr. Opin. Pharmacol. 11, 397–403 (2011).
-
Wesch, D., Peters, C., Oberg, H.-H., Pietschmann, K. & Kabelitz, D. Modulation of γδ T cell responses by TLR ligands. Cell. Mol. Life Sci. 68, 2357–2370 (2011).
-
Hu, Y. et al. γδ T cells: origin and fate, subsets, diseases and immunotherapy. Signal Transduct. Target. Ther 8, 434 (2023).
-
Dong, G. et al. FOXO1 Regulates Bacteria-Induced Neutrophil Activity. Front. Immunol. 8, 1088 (2017).
-
Hedrick, S. M., Hess Michelini, R., Doedens, A. L., Goldrath, A. W. & Stone, E. L. FOXO transcription factors throughout T cell biology. Nat. Rev. Immunol. 12, 649–661 (2012).
-
Cai, Y. et al. Differential Roles of the mTOR-STAT3 Signaling in Dermal γδ T Cell Effector Function in Skin Inflammation. Cell Rep 27, 3034–3048.e5 (2019).
-
Yu, Z., Cheng, L., Liu, X., Zhang, L. & Cao, H. Increased expression of INHBA is correlated with poor prognosis and high immune infiltrating level in breast cancer. Front. Bioinform. 2, 729902 (2022).
-
Duerschmied, D. et al. Platelet serotonin promotes the recruitment of neutrophils to sites of acute inflammation in mice. Blood 121, 1008–1015 (2013).
-
Manoharan, R. R. et al. NADPH oxidase-dependent free radical generation and protein adduct formation in neutrophils. RSC Adv 14, 24765–24780 (2024).
-
Jiang, Q., Duan, J., Van Kaer, L. & Yang, G. The Role of Myeloid-Derived Suppressor Cells in Multiple Sclerosis and Its Animal Model. Aging Dis 15, 1329–1343 (2024).
-
Hussain, T. et al. IFNAR1 Deficiency Impairs Immunostimulatory Properties of Neutrophils in Tumor-Draining Lymph Nodes. Front. Immunol. 13, 878959 (2022).
-
Agerholm, R., Kadekar, D., Rizk, J. & Bekiaris, V. Type I interferon supports γδ T-cell homeostasis and immunity through direct and indirect receptor signaling in mice. Eur. J. Immunol. 51, 3186–3193 (2021).
-
Agerholm, R., Rizk, J., Viñals, M. T. & Bekiaris, V. STAT3 but not STAT4 is critical for γδT17 cell responses and skin inflammation. EMBO Rep 20, e48647 (2019).
-
Mehrpouya-Bahrami, P. et al. STAT4 is expressed in neutrophils and promotes antimicrobial immunity. JCI Insight 6, e141326 (2021).
-
Yagi, M. et al. PDK1 regulates chemotaxis in human neutrophils. J. Dent. Res. 88, 1119–1124 (2009).
-
Huang, Y. et al. The effect of PDK1 in maintaining immune cell development and function. Biochem. Biophys. Res. Commun. 721, 150106 (2024).
-
Serrano, R., Wesch, D. & Kabelitz, D. Activation of Human γδ T Cells: Modulation by Toll-Like Receptor 8 Ligands and Role of Monocytes. Cells 9, 713 (2020).
-
Tamassia, N. et al. Human neutrophils activated via TLR8 promote Th17 polarization through IL-23. J. Leukoc. Biol. 105, 1155–1165 (2019).
-
Kambe, N. et al. Neutrophils predominate as IL1B-expressing cells in Schnitzler syndrome: Insights from the SCan study to evaluate the efficacy and safety of canakinumab in Japanese patients. Allergol. Int. https://doi.org/10.1016/j.alit.2025.04.003 (2025).
-
Sutton, C. E. et al. Interleukin-1 and IL-23 induce innate IL-17 production from gammadelta T cells, amplifying Th17 responses and autoimmunity. Immunity 31, 331–341 (2009).
-
Buus, T. B., Ødum, N., Geisler, C. & Lauritsen, J. P. H. Three distinct developmental pathways for adaptive and two IFN-γ-producing γδ T subsets in adult thymus. Nat. Commun. 8, 1911 (2017).
-
Kim, Y. S. et al. CD200Rhigh neutrophils with dysfunctional autophagy establish systemic immunosuppression by increasing regulatory T cells. Cell. Mol. Immunol. 21, 349–361 (2024).
-
Groselj-Grenc, M., Ihan, A. & Derganc, M. Neutrophil and monocyte CD64 and CD163 expression in critically ill neonates and children with sepsis: comparison of fluorescence intensities and calculated indexes. Mediators Inflamm 2008, 202646 (2008).
-
Gillespie, A. et al. Gamma Delta TCR and the WC1 Co-Receptor Interactions in Response to Leptospira Using Imaging Flow Cytometry and STORM. Front. Immunol. 12, 712123 (2021).
-
Akitsu, A. et al. IL-1 receptor antagonist-deficient mice develop autoimmune arthritis due to intrinsic activation of IL-17-producing CCR2( + )Vγ6(+)γδ T cells. Nat. Commun. 6, 7464 (2015).
-
Schadeck, J. et al. Vdelta1 T cells are more resistant than Vdelta2 T cells to the immunosuppressive properties of galectin-3. Front. Immunol. 14, 1286097 (2023).
-
Itakura, A. & McCarty, O. J. T. Pivotal role for the mTOR pathway in the formation of neutrophil extracellular traps via regulation of autophagy. Am J Physiol, Cell Physiol 305, C348–C354 (2013).
-
Zhang, H. et al. CD109 Restrains Activation of Cutaneous IL-17-Producing γδ T Cells by Commensal Microbiota. Cell Rep 29, 391–405.e5 (2019).
-
Vorstenbosch, J. et al. Transgenic mice overexpressing CD109 in the epidermis display decreased inflammation and granulation tissue and improved collagen architecture during wound healing. Wound Repair Regen 21, 235–246 (2013).
-
Linley, H. et al. CD200R1 is required for the development of γδ17 T cells. BioRxiv https://doi.org/10.1101/2025.05.19.654867 (2025).
-
Nakagawa, H. & Komorita, N. Complement component C3-derived neutrophil chemotactic factors purified from exudate of rat carrageenin-induced inflammation. Biochem. Biophys. Res. Commun. 194, 1181–1187 (1993).
-
Toledano-Fonseca, M. et al. The Combination of Neutrophil-Lymphocyte Ratio and Platelet-Lymphocyte Ratio with Liquid Biopsy Biomarkers Improves Prognosis Prediction in Metastatic Pancreatic Cancer. Cancers (Basel) 13, 1210 (2021).
-
Zuberbuehler, M. K. et al. The transcription factor c-Maf is essential for the commitment of IL-17-producing γδ T cells. Nat. Immunol. 20, 73–85 (2019).
-
Ren, J., Da, J. & Hu, N. Identification of COL1A1 associated with immune infiltration in brain lower grade glioma. PLoS ONE 17, e0269533 (2022).
-
Okuno, D. et al. Two Distinct Mechanisms Underlying γδ T Cell-Mediated Regulation of Collagen Type I in Lung Fibroblasts. Cells 11, 2816 (2022).
-
Guven, A. et al. Development and validation of an immunoassay for quantification of NCAM-1 in human plasma. J. Pharm. Biomed. Anal. 197, 113981 (2021).
-
Chavakis, T. et al. The pattern recognition receptor (RAGE) is a counterreceptor for leukocyte integrins: a novel pathway for inflammatory cell recruitment. J. Exp. Med. 198, 1507–1515 (2003).
-
Weiss, M. et al. IRF5 controls both acute and chronic inflammation. Proc Natl Acad Sci USA 112, 11001–11006 (2015).
-
Yan, J., Pandey, S. P., Barnes, B. J., Turner, J. R. & Abraham, C. T Cell-Intrinsic IRF5 Regulates T Cell Signaling, Migration, and Differentiation and Promotes Intestinal Inflammation. Cell Rep 31, 107820 (2020).
-
Li, S. et al. IRF5 as a potential immunological biomarker in lung adenocarcinoma. Transl. Cancer Res. 13, 6936–6946 (2024).
-
Wang, Y. et al. Natural γδT17 cell development and functional acquisition is governed by the mTORC2-c-Maf-controlled mitochondrial fission pathway. iScience 26, 106630 (2023).
-
Guerau-de-Arellano, M., Piedra-Quintero, Z. L. & Tsichlis, P. N. Akt isoforms in the immune system. Front. Immunol. 13, 990874 (2022).
-
Linley, H., Jaigirdar, S., Mohamed, K., Griffiths, C. E. & Saunders, A. Reduced cutaneous CD200:CD200R1 signalling in psoriasis enhances neutrophil recruitment to skin. BioRxiv https://doi.org/10.1101/2022.04.01.486720 (2022).
-
Bruno, M. E. C. et al. Accumulation of γδ T cells in visceral fat with aging promotes chronic inflammation. Geroscience 44, 1761–1778 (2022).
-
Hein, L. E., SenGupta, S., Gunasekaran, G., Johnson, C. N. & Parent, C. A. TGF-β1 activates neutrophil signaling and gene expression but not migration. PLoS ONE 18, e0290886 (2023).
-
Beatson, R. E. et al. TGF-β1 potentiates Vγ9Vδ2 T cell adoptive immunotherapy of cancer. Cell Rep. Med. 2, 100473 (2021).
-
Thomas, D. G. et al. LXR Suppresses Inflammatory Gene Expression and Neutrophil Migration through cis-Repression and Cholesterol Efflux. Cell Rep 25, 3774–3785.e4 (2018).
-
Jeon, J.-H., Hong, C.-W., Kim, E. Y. & Lee, J. M. Current understanding on the metabolism of neutrophils. Immune Netw 20, e46 (2020).
-
Caielli, S. et al. Oxidized mitochondrial nucleoids released by neutrophils drive type I interferon production in human lupus. J. Exp. Med. 213, 697–713 (2016).
-
Wang, X.-Y. et al. Tribbles Pseudokinase 3 Converts Sorafenib Therapy to Neutrophil-Mediated Lung Metastasis in Hepatocellular Carcinoma. Adv Sci (Weinh) 12, e2413682 (2025).
-
Liu, G. et al. Kinase AKT1 negatively controls neutrophil recruitment and function in mice. J. Immunol. 191, 2680–2690 (2013).
-
Vičić, M., Kaštelan, M., Brajac, I., Sotošek, V. & Massari, L. P. Current concepts of psoriasis immunopathogenesis. Int. J. Mol. Sci. 22, 11574 (2021).
-
Takematsu, H. & Tagami, H. Granulocyte-macrophage colony-stimulating factor in psoriasis. Dermatologica 181, 16–20 (1990).
-
Schön, M., Denzer, D., Kubitza, R. C., Ruzicka, T. & Schön, M. P. Critical role of neutrophils for the generation of psoriasiform skin lesions in flaky skin mice. J. Invest. Dermatol. 114, 976–983 (2000).
-
Furue, M., Furue, K., Tsuji, G. & Nakahara, T. Interleukin-17A and Keratinocytes in Psoriasis. Int. J. Mol. Sci. 21, 1257 (2020).
-
Wang, D. et al. A deep proteome and transcriptome abundance atlas of 29 healthy human tissues. Mol. Syst. Biol. 15, e8503 (2019).
-
Castillo-González, R., Cibrian, D. & Sánchez-Madrid, F. Dissecting the complexity of γδ T-cell subsets in skin homeostasis, inflammation, and malignancy. J. Allergy Clin. Immunol. 147, 2030–2042 (2021).
-
Gray, E. E., Suzuki, K. & Cyster, J. G. Cutting edge: Identification of a motile IL-17-producing gammadelta T cell population in the dermis. J. Immunol. 186, 6091–6095 (2011).
-
Sumaria, N. et al. Cutaneous immunosurveillance by self-renewing dermal gammadelta T cells. J. Exp. Med. 208, 505–518 (2011).
-
Arunachalam, P. et al. CCR6 (CC Chemokine Receptor 6) Is Essential for the Migration of Detrimental Natural Interleukin-17-Producing γδ T Cells in Stroke. Stroke 48, 1957–1965 (2017).
-
Hammerich, L. et al. Chemokine receptor CCR6-dependent accumulation of γδ T cells in injured liver restricts hepatic inflammation and fibrosis. Hepatology 59, 630–642 (2014).
-
Campbell, J. J. et al. IL-17-Secreting γδ T Cells Are Completely Dependent upon CCR6 for Homing to Inflamed Skin. J. Immunol. 199, 3129–3136 (2017).
-
Zhang, M. & Zhang, X. The role of PI3K/AKT/FOXO signaling in psoriasis. Arch. Dermatol. Res. 311, 83–91 (2019).
-
Horváth, S. et al. Methodological refinement of Aldara-induced psoriasiform dermatitis model in mice. Sci. Rep. 9, 3685 (2019).
-
Macias, S. L. & Keselowsky, B. G. Perspectives on immunometabolism at the biomaterials interface. Mol. Aspects Med. 83, 100992 (2022).
-
Willemsen, J. et al. TNF leads to mtDNA release and cGAS/STING-dependent interferon responses that support inflammatory arthritis. Cell Rep 37, 109977 (2021).
-
Klapan, K. et al. Evidence for Lysosomal Dysfunction within the Epidermis in Psoriasis and Atopic Dermatitis. J. Invest. Dermatol. 141, 2838–2848.e4 (2021).
-
Serban, A. I., Stanca, L., Geicu, O. I., Munteanu, M. C. & Dinischiotu, A. RAGE and TGF-β1 Cross-Talk Regulate Extracellular Matrix Turnover and Cytokine Synthesis in AGEs Exposed Fibroblast Cells. PLoS ONE 11, e0152376 (2016).
-
Bhattacharjee, O., Ayyangar, U., Kurbet, A. S., Ashok, D. & Raghavan, S. Unraveling the ECM-Immune Cell Crosstalk in Skin Diseases. Front. Cell Dev. Biol. 7, 68 (2019).
-
Gangwar, R. S., Gudjonsson, J. E. & Ward, N. L. Mouse models of psoriasis: A comprehensive review. J. Invest. Dermatol. 142, 884–897 (2022).
-
Swindell, W. R. et al. Imiquimod has strain-dependent effects in mice and does not uniquely model human psoriasis. Genome Med 9, 24 (2017).
-
Terhorst, D. et al. Dynamics and Transcriptomics of Skin Dendritic Cells and Macrophages in an Imiquimod-Induced, Biphasic Mouse Model of Psoriasis. J. Immunol. 195, 4953–4961 (2015).
-
Malyala, P. & Singh, M. Endotoxin limits in formulations for preclinical research. J. Pharm. Sci. 97, 2041–2044 (2008).
-
Kolter, J. et al. A subset of skin macrophages contributes to the surveillance and regeneration of local nerves. Immunity 50, 1482–1497.e7 (2019).
-
Fleming, T. J., Fleming, M. L. & Malek, T. R. Selective expression of Ly-6G on myeloid lineage cells in mouse bone marrow. RB6-8C5 mAb to granulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family. J. Immunol. 151, 2399–2408 (1993).
-
Masuda, S. et al. NETosis markers: Quest for specific, objective, and quantitative markers. Clin. Chim. Acta 459, 89–93 (2016).
-
Kuczia, P. et al. Citrullinated histone H3, a marker of extracellular trap formation, is increased in blood of stable asthma patients. Clin. Transl. Allergy 10, 31 (2020).
-
Lou, F., Sun, Y. & Wang, H. Protocol for flow cytometric detection of immune cell infiltration in the epidermis and dermis of a psoriasis mouse model. STAR Protocols 1, 100115 (2020).
-
GitHub – lh3/seqtk: Toolkit for processing sequences in FASTA/Q formats. https://github.com/lh3/seqtk.
-
Kim, D., Langmead, B. & Salzberg, S. L. HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12, 357–360 (2015).
-
Liao, Y., Smyth, G. K. & Shi, W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014).
-
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol 15, 550 (2014).
Acknowledgements
This work was supported by the National Institutes of Health (DE027301 to BGK and GAH; DK129690 and AI171045 to BGK, DA, and GAH). Thanks to the University of Florida Interdisciplinary Center for Biotechnology Research for preparation and sequencing of RNA libraries. We would also like to thank the Karst lab at the University of Florida for use of their bead beater. Protein-level analysis of tissue lysate was made possible by Eve Technologies for multiplex ELISA. Thanks to the University of Florida Molecular Pathology Core, RRID:SCR_016601, for histopathological staining and image scanning. Lastly, we would like to thank the Center for Immunology and Transplantation at the University of Florida for the use of their Cytek Aurora spectral cytometer.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Simonovich, J.A., Macias, S.L., Zhou, M. et al. Indoleamine 2,3-dioxygenase-galectin-3 fusion protein remodels inflammatory and fibrotic programs, ameliorating imiquimod-induced psoriasis. npj Biomed. Innov. 2, 36 (2025). https://doi.org/10.1038/s44385-025-00040-y
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s44385-025-00040-y