
Introduction
Natural sources have been explored for numerous active compounds having unique chemical structure and potent bioactivities since time immemorial. The Cinchona or Quina belonging to the Rubiaceae family is industrially valued for its quinoline alkaloids. This plant is native to South America and contains 0.4 to 4% alkaloids in its bark. Except for South American countries major cinchona plantation is located in the Darjeeling districts of West Bengal, India. This area of plantation covers some of the important recognized Cinchona species viz. C. officinalis, C. succirubra, C. calisaya and C. ledgerianana that contain an array of quinoline alkaloids1. The dried bark of Cinchona exhibits alkaloids like quinine, quinidine, cinchonine and cinchonidine besides other flavonoids, steroids, and terpenoids2,3,4. Quinine and related compounds belong to the aryl amino alcohol class of quinolines. Quinine and quinidine have been used effectively against the asexual erythrocytic stages of malaria protozoa. Quinine interferes with the detoxification of heme in protozoa leading to oxidative damage and parasite death5. The antimicrobial role of quinine against S. cerevisiae points out tryptophan starvation as the primary cause6,7. The succession of research on quinine and its derivatives has led to the development of alternate commercial applications of Cinchona bark8,9. However, very little information is available about the mode of action of quinine and its derivatives against plant pathogens.
Quinine and its derivatives have been classified as potent inhibitors of plant pathogens and exhibit strong antifungal effects against Botrytis cinerea, Fusarium graminearum, Rhizoctonia solani and Sclerotinia sclerotiorum resulting upto 60% growth inhibition. Modulation in the chemical structure of quinine in its heterocyclic state (Thiosemicarbazide) can enhance its antimicrobial efficiency10,11,12,13. Quinine-derived compounds have increased the solubility and bioactivity in appropriate solvent retaining their antifungal properties14. Indoquinoline derivatives have a deleterious effect on the oxidative phosphorylation pathway of R. solani fungal cells that disrupt the mitochondrial membrane15.Silver nano prepared with turmeric extracts showed a similar disruption of the respiratory system of the fungus16. Although plant-based liquid formulations typically contain a complex mix of secondary metabolites, studies have indicated that a single active compound plays the primary role in antifungal activity17.
The surge in global population requires accelerated agricultural production. The detrimental effect of phytopathogens imposes a significant hindrance to crop production. Rhizoctonia solani belonging to Basidiomycota is a predominant necrotrophic fungus that causes rice sheath blight disease infecting leaf sheath and lamina. The persistent sclerotia in soil germinate into leaf penetrating mycelia in appropriate climatic conditions. In case of severe infection, this soil-residing fungi reduces up to 50% of the crop yield in rice-growing countries of southeastern Asia18. Unjudicial uses of chemical pesticides not only result in the development of resistant strains of the pathogen but also cause serious environmental hazards. Recent findings have shown that metabolic resistance in R. solani may develop due to mutations in fungicide target sites and changes in gene expression of Glutathione S-transferase and Cytochrome p45019,20. Low-biodegradable synthetic fungicides have raised concerns due to their detrimental effects on crops and biodiversity. Sustainable agricultural practices require the development of eco-friendly plant-derived broad-spectrum biodegradable botanical antifungal formulations21,22. Different ratio of phytochemical compounds in solvents with definite physicochemical properties (pH, viscosity, and stability) as formulation attributed with improved quality and optimized application doses as compared to raw plant parts. Due to the unavailability of resistant genes against this devastating fungus, synthetic fungicides are the only effective strategy for managing R. solani23. Combinations of diverse secondary metabolites like phenolics, terpenoids, and alkaloids in plant-derived fungicides exhibit antifungal activity by suppressing fungal spore germination, sporulation, and hyphal development24.
The biologically synthesized quinoline alkaloids had been replaced by synthetic alternatives due to the development of resistance by Plasmodium vivax towards quinine. Therefore, it is necessary to explore other biological applications of quinoline alkaloids for commercial purposes, utilizing natural sources of these alkaloids. In the present investigation, Cinchona based formulation was tested against Rhizoctonia solani. Furthermore, the research examines the underlying molecular mechanism of the fungal inhibition triggered by effective Cinchona based formulation. The result indicates dual functionality of the extract on the fungal metabolism. This multiple mode of inhibition is indicative of the robustness of Cinchona-based formulation as botanical fungicide. The findings provide fresh insights into development of resistance-manageable biopesticides.
Results
Evaluation of antifungal efficacy of Cinchona bark extract in different solvent
EC50 value of extracts
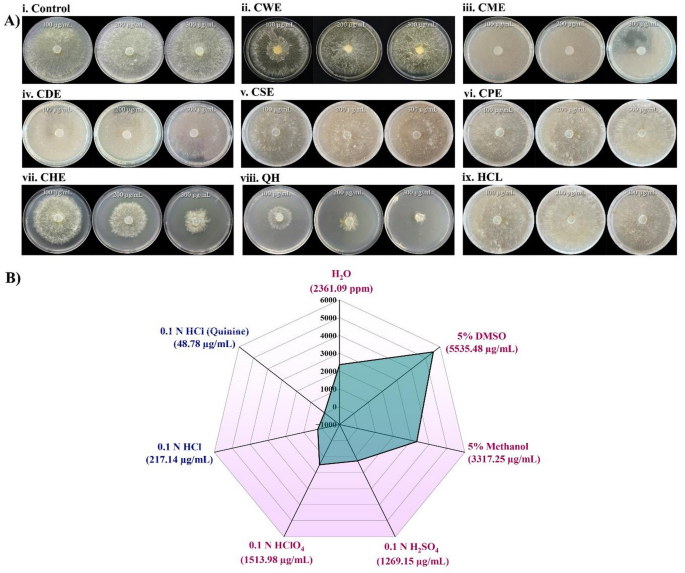
Different acidic extracts were tested against rice pathogen Rhizoctonia solani AG1-IA in poison food assay at different concentrations (10 to 500 µg/mL) to determine their antifungal activity. CHE showed the highest antifungal activity (EC50 value of 217.14 ± 0.40 µg/mL) effectively suppressing Rhizoctonia solani mycelia development (Fig. 1A,B). CPE and CSE showed considerable antifungal activity with EC50 values of 1513.98 µg/mL and 1269.15 ± 0.70 µg/mL, respectively. CWE showed a moderate EC50 compared to other selected solvents (2361.09 ± 0.56 µg/mL). CME and CDE showed lowest antifungal activity with higher EC50 value of 3317.25 ± 0.42 µg/mL and 5535.48 ± 0.23 µg/mL (Fig. 1B). These findings indicated the importance of solvent selection in enhancing the extraction of bioactives from Cinchona bark. Specially, using 0.1 N HCl as solvent significantly improves the extraction of active antifungal compounds from Cinchona bark. As the major alkaloid of Cinchona bark is quinine, the experiment was repeated using QCl which showed EC50 value of 48.78 ± 0.27 µg/mL (Fig. 1A,B). The descriptive statistics with regression analysis of EC50 values for different prepared solvents have been incorporated in Table 1. The highest (4.61%) total quinine content was found in CBF, and the 95% CI (confidence interval) range was lowest in the case of CBF among the extracts.

(A) Antifungal activity of different Cinchona bark extracts against Rhizoctonia solani strain AG1-IA by poison food assay. Mycelial growth after 3 days on PDA plates containing different Cinchona extracts viz. Cinchona Water extract – CWE, Cinchona Methanol Extract – CME (5% methanol), Cinchona DMSO Extract – CDE (5% dimethyl sulfoxide), Cinchona H2SO4 Extract – CSE (0.1 N sulfuric acid), Cinchona HClO4 Extract – CPE (0.1 N perchloric acid), Cinchona HCl Extract – CHE (0.1 N hydrochloric acid) and Quinine in HCl (0.1 N Hydrochloric acid) at a concentration of 100 µg/mL, 200 µg/mL and 300 µg/mL. Plates containing only 0.1 N hydrochloric acid were used as solvent control while untreated plates served as negative controls (ii) 2D spider diagram illustrating the EC50 values of Cinchona bark extracts against Rhizoctonia solani AG1-IA in various solvent systems. The antifungal efficacy varies depending on the solvent, with the highest activity occurring in 0.1 N HCl with quinine and the lowest in 5% DMSO.
The current study uses a formulation strategy in which active phytochemicals like quinine are extracted under mild acidic conditions (0.1 N HCl) without causing structural alterations. The hydrochloric acid-based extract (CHE) had the highest antifungal activity against Rhizoctonia solani and the lowest EC50 value. Based on its improved efficacy, CHE was designated as the Cinchona Bark Formulation (CBF) for further extensive analysis.
Bioactive compound profiling of CBF
CBF (200 µg/mL) was subjected to GC-MS analysis to explore its bioactive components. GC-MS analysis revealed that cinchona bark extract contains diverse bioactive compounds across several chemical classes. Quinoline alkaloids including quinine, 2-hydroxyquinine, cinchonine, cinchonidine and cinchonine-p-bromobenzenesulfonate were found to be predominant compounds accounting for 82.93% of total peak area (Fig. 2A). Other detected classes included imidic acid derivatives (13.14%), tryptophan derivatives (1.68%), coumarins (1.08%) and minor constituents such as complex terpenoids (0.51%), saturated hydrocarbons (0.41%), aniline/Schiff bases (0.19%) and nitro-cycloalkane derivatives (0.06%) (Table 2).

Bioactive compound profile of Cinchona bark extract in 0.1 N HCl (CBF) using: (A) GC-MS chromatogram showing four major quinoline alkaloids—cinchonine, cinchonidine, hydroxyquinine, and quinine—represented as peaks 1 to 4, respectively; (B) RP-HPLC overlay of standard quinine (50 µg/mL) and Cinchona bark extract (CBF, 200 µg/mL). Sharp peak at retention time 2.7 min identified by arrow detects the presence of quinine; (C) FTIR spectrum displaying characteristic functional group vibrations, supporting the presence of alkaloid compounds and N-H bond stretching.
Detection and quantification of quinine and other major alkaloids like quinidine, cinchonine, cinchonidine was performed by RP-HPLC using C18 column (Supplementary information Fig. 1). Total quinine (13.67 ± 0.12 µg/mL) was detected in CBF (200 µg/mL) at 2.61 min over a run time of 15 min (5.43 ± 0.23% in Cinchona bark powder) (Fig. 2B) (y = 2302.8x + 1747.4, R² = 0.985). Other alkaloids were found in negligible quantity. Among all the solvents examined 0.1 N HCl provided highest total quinine content (4.61 ± 0.03%) followed by 0.1 N H2SO4 (1.61 ± 0.06%), 5% methanol (1.36 ± 0.02%), 0.1 N HClO4 (1.34 ± 0.072%), 5% DMSO (0.89 ± 0.018%) and aqueous extraction (0.029 ± 0.02%) (Supplementary information Fig. 2).
Fourier Transform Infrared (FTIR) spectrum analysis is a primary tool for observing the stretching of bonds. FTIR spectrum analysis was conducted to identify the fingerprint regions of quinine from CBF. These revealed the presence of Quinoline alkaloid-specific N-H, N-O, C-O and C-N stretching with variable intensities and specific functional groups with distinctive peak types between the wavenumbers of 400–1500 cm−1 (Fig. 2C; Table 3).The presence of this N-H bond can be found missing in quinine powder although QCl was found with a strong peak for N-H at wavenumber of 1513.02 cm−1 (Supplementary information Fig. 3).

In silico investigation of the amino acid permeases rhAAP-I and rhAAP-II. (A) Predicted 3D structures of rhAAP-I and rhAAP-II with overall folding patterns. (B) Transmembrane orientation and ligand binding visualization demonstrating the interaction of quinine (Qn) and tryptophan (Trp) with proteins. (C) Binding site analysis reveals important interaction residues for quinine (Qn) and tryptophan (Trp) inside each permease, indicating differences in binding affinities and orientations between rhAAP-I and rhAAP-II. (D,E) Site-directed mutations and their effect on Molecular Docking of Quinine with Amino Acid Permeases rhAAP-I (D) and rhAAP-II (E) predicting the effects of interacting amino acid mutations on ligand Stability.
In silico analysis suggests quinine acts as noncompetitive inhibitor of AAP of Rhizoctonia
Structural modeling of Rhizoctonia Amino acid permease was performed with available templates in the Uniprot database. The Rhizoctonia amino acid permeases I (rhAAP-I) containing 529 amino acids revealed 91.12% sequence identity with the template Rhizoctonia solani AG-8 (AlphaFold DB model template: A0A066W7G8). Rhizoctonia amino acid permease -II (rhAAP-II) consisting of 307 amino acids showed 70.78% similarity with the template from Thanatephorus cucumeris strain AG1-IA (AlphaFold DB model template: L8WMA4). Ramachandran favored regions for both the models were found as 95.64% (rhAAP-I) and 94.27% (rhAAP-II) (Fig. 3A, Supplementary information Fig. 4). The models are available in Model Archive (modelarchive.org) with the accession codes ma-lwg2r (https://modelarchive.org/doi/https://doi.org/10.5452/ma-lwg2r) and ma-5kjcj (https://modelarchive.org/doi/https://doi.org/10.5452/ma-5kjcj) under Swiss Institute of Bioinformatics (Fig. 3A). The Boiled Egg Model (SwissADME) analysis indicated that quinine exhibits high lipophilicity (LogP: 3.36) and moderate solubility (LogS: −3.71), suggesting efficient membrane permeability and passive diffusion. Both rhAAP-I and rhAAP-II were predicted to be localized in plasma membrane, implicating their roles in the intracellular transport or trafficking of amino acids (Fig. 3B, Supplementary information Fig. 4B). Secondary structure prediction using PSIPRED and transmembrane topology analysis with TMHMM identified multiple transmembrane helices (12 for rhAAP-I and 3 for rhAAP-II), reinforcing their transport function. Membrane orientation via (PPM 3.0 server) provided spatial orientation insights of both the selected permease (Supplementary information Fig. 4C and D). Ligand Binding studies using Discovery Studio and PyRx demonstrated rhAAP-I and rhAAP-II binds with tryptophan with − 6.6 kcal/mol and − 5.4 kcal/mol, respectively, suggesting Trp transport as its primary role (Fig. 3B). The quinine showed a stronger binding affinity compared to tryptophan in both permeases, with binding energies of −7.8 kcal/mol for rhAAP-I and − 6.5 kcal/mol for rhAAP-II (Table 4). AutoDock simulation revealed different active site binding for tryptophan and quinine. Interaction analysis revealed that tryptophan and quinine bind to the distinct amino acid residues in the enzyme: tryptophan in rhAAP-I interacts with ARG:77, GLY:244, ASN:410, ASP:461, PHE:462, LEU:240 and LEU:243, whereas quinine binds HIS:36, VAL:362, LEU:42, TYR:35, ARG:363, PHE:37, 37 and ALA:119. Similarly, in rhAAP-II, tryptophan interacts with HIS:96, GLY:119, ALA:121, and PRO:113, while quinine engages ARG:207, PHE:196, CYS:193, PRO:218, PRO:190, HIS:217, ILE:189, and LEU:210 forming stable hydrogen bonding and π-alkyl interactions (Fig. 3C). The ligand interaction was verified using site-directed changes in interactive amino acids. In rhAAP-I, ARG363, HIS36, and TYR35 interacted strongly with the ligand via Pi-Alkyl, Alkyl, and hydrogen bonds (Table 4). Mutations in ARG363 to ALA363, HIS36 to ALA36, and TYR35 to PHE35 reduced structural stability (RMSD: 1.8, 1.6, and 1.4 Å, respectively). Parallelly, these mutations were found to disrupt ligand interactions and decrease binding strength (−6.1, −6.3 and − 6.5 kcal/mol) (Fig. 3D). Similarly, in rhAAP-II, HIS217 and CYS193 stabilized quinine binding with hydrogen bonds and hydrophobic interactions (Table 4). Mutations in HIS217 to ALA217 and CYS193 to ALA193 reduced structural stability (RMSD:1.7 and 1.5Å) as well as binding strength (−5.8 and 5.7 kcal/mol) (Fig. 3E). These data demonstrated that the amino acid pocket made up of ARG363, HIS36, TYR35, HIS217, and CYS193 is highly specific to quinine. Site-directed alterations drastically reduce binding efficiency and structural alignment, highlighting the pocket’s importance in quinine recognition (Table 4). The molecular docking results confirmed that the original residues at the quinine binding site are required for stable and specific binding.

The effect of tryptophan supplementation (100 to 400 µg/mL) on the antifungal activity of CBF and QCl: (A) Mycelial growth inhibition on agar plates at different concentrations of CBF and QCl. PDA plates with CBF (200 µg/mL) and QCl (50 µg/mL) were used as treatment control, whereas PDA plates without any treatments were considered as negative control; (B) Graphical representation of percentage inhibition in fungal growth. (C) Graphical representation of fungal dry weight (mg) and sclerotia weight (mg) taken after 7 days of fungal growth in PDB (data represent mean ± SE, n = 3, *p < 0.001). (D) Micromorphological changes in fungal hyphae (red arrows indicate structural abnormalities and black arrows indicate normal hyphal structure); Untreated plates served as negative controls.
Cinchona based formulation (CBF) causes trp starvation in rhizoctonia
Fungal growth parameters
The bioinformatic analysis pointed out the inhibitory effect of quinine on the Trp transporters in Rhizoctonia. To investigate the possible mechanism of QCl and CBF, a tryptophan supplementation experiment was performed to demonstrate the effects of Trp on the fungal growth recovery. The fungal growth was found to be lowest in CBF compared to all other acidic extracts and the negative control. When an increasing concentration of tryptophan (50 to 500 µg/mL) was supplied in the CBF media, the mycelial growth and sclerotia development were recovered by the fungi (Fig. 4A). Trp supplemented CBF and QCl media at a concentration of 400 µg/mL resulted in complete recovery of fungal mycelium in media (Fig. 4A). The percentage inhibition, average fungal dry weight, and sclerotial weight were measured in the treatments with or without Trp supplementation. The CBF and QCl showed 58.76 ± 4.67 and 86.59 ± 6.13% inhibitions in fungal growth (Fig. 4B). The additional supplementation of 400 µg/mL Trp reduced the growth inhibition by 70.39% and 74.51% for CBF and QCl respectively (Table 5). Fungal dry weight and the sclerotia weight for QCl were 5 ± 0.82 mg and 12 ± 0.92 mg, respectively (Fig. 4C). For CBF treatment, fungal dry weight and sclerotia weight were measured at 14 ± 1.01 mg and 22 ± 1.21 mg, respectively (Fig. 4C). QCl + Trp-treated (400 µg/mL) samples showed partial recovery with fungal dry weight of 34.00 ± 1.10 mg and sclerotia weight of 54.67 ± 0.91 mg. CBF + Trp-treated samples had a fungal dry weight of 33.33 ± 1.53 mg and sclerotia weight of 67.67 ± 1.06 mg, suggesting that the Trp-supplemented media fungal growth is lower than control but greater than QCl and CBF. The Coefficient of Variation (CV) results indicated that QCl (7.08%) and CBF (5.5%) mediated inhibitions are consistent, whereas QCl + Trp (13.68%) and CBF + Trp (9.94%) were more variable. In terms of fungal dry weight, Control exhibited a consistent growth (2.25%), whereas in CBF, growth varied (16.40%) significantly. For sclerotia weight, QCl (1.66%) and QCl+ Trp (1.57%) experiments, the growth was constant, whereas for CBF (7.67%) and CBF + Trp (5.50%) experiments, the growth was more variable. The figure shows statistically significant differences (p < 0.001) between QCl vs. QCl + Trp and CBF vs. CBF + Trp, indicating that tryptophan significantly reduces the antifungal effectiveness of both treatments (Table 5). Micromorphological studies of Rhizoctonia mycelia under various treatments (CME, CDE, CSE, and CPE) revealed normal hyphal development (Fig. 4D). The hyphae showed smooth expansion, homogeneous branching, and structural integrity without abnormalities or inhibition. Micro-morphological observations of CBF-treated mycelia revealed variation in hyphal characteristics of the Rhizoctonia. The deformities were observed in the angle of hyphal emergence from fusion cells. In the treated cells, the angle is reduced with distorted constriction in the emerging hyphal structure. The abnormalities in hyphal structure were recovered to their original uninucleate septate structure when the media was supplemented with Trp (Fig. 4D). In vivo application of the CBF and QCl projected similar results. Soil-inoculated Rhizoctonia produced characteristic symptoms like water-soaked lesions, irregular necrotic spots, and grayish-white mycelial growth in the control, whereas foliar spray with 50 µg/mL QCl and 200 µg/mL CBF in every 5 days exhibited significant reduction in lesion formation with infection severity of 19.645% (QCl) and 8.54% (CBF) (Fig. 5). On the other hand, seedlings sprayed with QCl + Trp and CBF + Trp displayed a resurgence of fungal infection (28.98% and 38.78%, respectively). These findings indicated that the effects of CBF and QCl treatments can be reversed with Trp supplementation.

In vivo antifungal activity of various treatments against Rhizoctonia solani in Rice (21-days old) (A) Visual assessment of infection severity in 14 days old rice seedlings grown in R. solani pre-inoculated soil media; (B) ImageJ (version 1.8)-processed grayscale images highlighting infection severity in rice seedlings treated with CBF, CBF + Trp, QCl, and QCl + Trp. All the treatments were compared with control trays without the application of any treatments.
Intracellular level of Tryptophan and quinine
A time course study of Rhizoctonia solani growth under various treatments was conducted over 120 h (Fig. 6A). The Control with PDA media only showed a gradual increase, with a final OD of 0.63 ± 0.04, indicating unconstrained fungal development. The QCl treatment effectively inhibited fungal growth, with a low OD (0.19 ± 0.02) at 120 h after incubation. The CBF treatment demonstrated excellent antifungal efficacy, with a final OD of 0.18 ± 0.02. QCl + Trp had an OD of 0.55 ± 0.05, while CBF + Trp successfully suppressed growth with a final OD of 0.45 ± 0.04, clearly showing the effect of Trp on the recovery of fungal growth. The quantification of Trp and Quinine before and after 400 µg/mL Trp supplementation in fungal media was conducted by HPLC. External supplementation of Trp in the CBF and QCl treated media elevated tryptophan levels (y = 9832x – 35426, R² = 0.99) in the mycelia significantly (120.39 ± 0.96 mg/100 g and 98.39 ± 0.85 mg/100 g, respectively) (Fig. 6B). In CBF and QCl-treated mycelia, 8.98 ± 0.18 mg/100 g and 17.65 ± 0.14 mg/100 g quinine was detected, respectively, which remained unaltered when Trp was supplemented (Fig. 6C). CV results demonstrated that all treatments were extremely consistent, with Control (0.24%) and QCl + Trp (0.06%) being the most stable for tryptophan content, while QCl (0.85%) and CBF + Trp (1.03%) had low variability for quinine (Table 5). These results confirmed that both CBF and QCl affected Trp accumulation negatively and cause Trp starvation in the mycelia however Trp supplementation did not influence the quinine uptake.

(A) Growth curve analysis of R. solani under different treatments viz. QCl, CBF, QCl + Trp, CBF + Trp up to 120 h after inoculation through expressed in OD405. Spectrophotometric observations were recorded at every 5 h intervals to monitor fungal growth dynamics in response to the treatments. All the treatments were compared with PDB culture without any treatment i.e. control. Results were expressed as mean of three replicates with SD values. (B) Quantification of tryptophan content and quinine content from fungal mycelia using RP-HPLC: (i) Total tryptophan concentration in fungal biomass after various treatments (mean ± SE, n = 3, *p < 0.001); (ii) RP-HPLC chromatogram with tryptophan peak in QCl + Trp (50 µg/mL of QCl + 400 µg/mL of Trp) treatment. (C) (i) Fungal quinine concentration across treatments (mean ± SE, n = 3); (ii) RP-HPLC chromatogram reveals a quinine peak in QCl + Trp (50 µg/mL of QCl + 400 µg/mL of Trp) treatment.
Quinine alters the expression of amino acid permeases and Cyt.p450
Transcript of rhAAP-I and AAP-II were measured across all the conditions. CWE, QCl, QCl + Trp, CBF and CBF + Trp. rhAAP-I exhibited a 9.29-fold and 4.76-fold increase of expression in QCl and CBF-treated hyphae, respectively. The addition of tryptophan exhibited a reduced expression of the genes (1.61-fold and 4.6-fold) compared to CBF and QCl. rhAAP-II expression also showed a similar trend of increased expression under QCl (6.19-fold) and CBF (4.23-fold), and comparatively low expression (1.72 and 1.39-fold, respectively) under tryptophan supplementation (Fig. 7). Expression of Cytochrome P450 (Cyt.P450) has been linked to fungal resistance against fungicides. The Cinchona water extract (CWE) showed a very high expression of Cyt.P450, implying that the fungus effectively blocks the CWE, and it does not have any significant fungicidal activity on the Rhizoctonia solani. The expression of Cyt.P450 was found to be decreased in QCl and CBF (1.58 and 2.73 -and 2.73-fold, respectively). Interestingly, Cyt.P450 expression increased 5 to 6-fold when the QCl and CBF-treated media were supplemented with Trp (6.3 and 5.75-fold, respectively) (Fig. 7). These results highlighted that CBF and QCl treatments strongly modulate rhAAP and Cyt.P450 gene expression, potentially linking them to the tryptophan starvation mechanism.

Relative expression of amino acid permeases rhAAP-I (magenta) and rhAAP-II (Blue) and Cytochrome P450 (purple) in Rhizoctonia solani AG1-IA treated with Cinchona-derived extracts. qPCR analysis demonstrated a fold change in expression levels under several treatments (CWE, CBF, CBF + Trp, QCl, QCl + Trp) compared to the control. CWE is considered as solvent control while culture without any treatments were termed as negative control. (Data are mean ± SE, n = 3; p < 0.05, p < 0.01, and p < 0.001).
CBF affects the mitochondrial membrane and ETC of the fungus
Treatment with QCl and CBF resulted in a significant reduction in mitochondrial membrane potential (MMP) and increased Reactive Oxygen Species (ROS) levels in fungal mycelia (Fig. 8A & B). DCFH-DA staining exhibited that Reactive Oxygen Species (ROS) levels were significantly elevated in QCl (21.38 ± 0.63 RFU) and CBF (17.69 ± 0.62 RFU) treated mycelia (Fig. 8C). The ROS accumulation adversely affects mitochondrial membrane permeability (MMP), which was evident by reduced fluorescence when the mycelia were stained with Rhodamine123 (7.79 ± 0.60 and 11.03 ± 0.6 RFU, respectively). Trp supplemented mycelia exhibited reduction of ROS accumulation but failed to significantly restore the MMP and respiratory function of the fungus as the control experiment (Fig. 8C). To confirm further, the impact of different treatments on mitochondrial succinate dehydrogenase (SDH) activity of Rhizoctonia solani was measured (Fig. 8D). In the control experiment with PDA media, SDH activity showed a natural time-dependent drop, progressively declining from 199.34 ± 0.92 µg INT-formazan g−1 min−1 at 0 h to 136.97 ± 0.65 µg INT-formazan g−1 min−1 at 120 h. SDH activity in QCl-treated samples showed significant suppression, dropping from 193.13 ± 0.81 µg INT-formazan g−1 min−1 at 0 h to 9.93 ± 0.93 µg INT-formazan g−1 min−1 after 120 h. The activity of QCl in combination with tryptophan (QCl + Trp) decreased SDH activity from 189.21 ± 0.84 µg INT-formazan g−1 min−1 at 0 h to 78.87 ± 0.94 µg INT-formazan g−1 min−1 after 120 h, greater than only QCl treatment. CBF treatment also decreased SDH activity from 195.21 ± 0.91 µg INT-formazan g−1 min−1 at 0 h to 36.60 ± 0.84 µg INT-formazan g−1 min−1 at 120 h. This inhibition was mitigated by CBF + Trp, as levels dropped from 190.03 ± 0.94 µg INT-formazan g−1 min−1 at 0 h to 100.13 ± 0.71 µg INT-formazan g−1 min−1 at 120 h. QCl and CBF had the most potent inhibitory effects on SDH activity.

Evaluation of reactive oxygen species (ROS) accumulation and mitochondrial membrane potential (MMP) expression of electron transport chain genes in Rhizoctonia solani exposed to Cinchona-derived extracts. (A,B) Representative fluorescence micrographs of ROS stained with 2′,7′-dichlorodihydrofluorescein diacetate(DCFH-DA) and MMP stained with Rhodamine 123; (C) Quantitative assessment of relative fluorescence intensity (RFU) for ROS and MMP determined by ImageJ (version 1.8); (D) Time-course enzyme assay of succinate dehydrogenase (SDH) activity in Rhizoctonia solani expressed in µg INT-formazan g−1min−1.SDH activity was determined spectrophotometrically over a 120 h period at every 20 h intervals under different treatments (QCl, QCl + Trp, CBF, and CBF + Trp). Plates where fungal growth was observed without any treatments was taken as control. Values are presented as mean ± SD of replicate experiments.; (E) qPCR analysis of differential gene expression for QCR1 and QCR9 (components of ubiquinol-cytochrome C reductase, Complex III) as well as COX2 and COX3 (components of Cytochrome c oxidase, Complex IV) under different treatments. The data represents the mean ± SE (n = 3); p < 0.05, p < 0.01, and p < 0.001.
Expression of electron transport chain (ETC) genes
Given the observed ROS accumulation and mitochondrial membrane depolarization, it was hypothesized that these effects may be linked to altered expression of electron transport chain (ETC) genes. qRT-PCR analysis was performed for QCR1 and QCR9 (components of ubiquinol-cytochrome C reductase, Complex III) as well as COX2 and COX3 (components of Cytochrome c oxidase, Complex IV). COX3 showed an increased expression (fold change of 10.08) compared to control (Fig. 8E). Other genes exhibited a 3-to-6-fold increase in their expression under QCl treatment. CBF also exhibited moderate upregulation of the genes (2 to 6-fold). External tryptophan supplementation in QCl + Trp and CBF + Trp treatments slightly altered ETC gene expression but failed to show significant recovery in the compensatory transcription of the genes.
Discussion
Sheath blight disease caused by Rhizoctonia solani AG1-IA is a serious constraint on rice production in India and other countries of Southeast Asia. Surveys conducted between 2016 and 2018 identified 21 isolates from various hosts, indicating widespread distribution of Rhizoctonia and severe output losses ranging from 14.3% to 39.7% depending on agro-climatic conditions25,26. R. solani AG-1 IA causes sheath blight in rice, maize, and other Gramineous plants27. Rhizoctonia AG1-IA isolate shares > 95% genomic similarity with other strains in core genes with minor variation in accessory genes that regulate pathogenicity28,29. The selection of AG1-IA isolate for the current study was based on its dominance and severity in the eastern region of India.
While synthetic quinine derivatives have been tested against plant pathogenic fungi, the antifungal properties of natural Cinchona bark extracts have not been studied30,31. The current work indicates the efficacy of a simple and environmentally friendly extraction method against R. solani. Quinine, the main alkaloid, is slightly basic, and its solubility is affected by protonation with acidic solvents (0.1 N HCl), providing higher concentrations than neutral or less polar solvents like water, DMSO, or methanol32. Quinine has antifungal properties as evidenced by its EC50 of 48.78 µg/mL against Rhizoctonia solani. Commercial synthetic fungicides with a range of EC50 values varies greatly in efficacy, has diverse mode of action and often impose adverse effect on environment: tebuconazole (EC50 = 3.10 µg/mL) inhibits ergosterol biosynthesis but is moderately persistent, hymexazol (EC50 = 10.49 µg/mL) disrupts RNA and protein synthesis with low toxicity but have poor degradability, azoxystrobin and trifloxystrobin have higher EC50 values (49.7 and 97.1 µg/mL) and pose ecological risks, similar to carbendazim (EC50 = 26.96 µg/mL)33,34,35,36. Quinine, a natural alkaloid, offers an eco-friendly alternative that has a lower environmental impact while maintaining sustainable efficacy. Among the different Cinchona extract CBF, prepared in 0.1 N HCl showed significant antifungal activity with an EC50 of 217.14 µg/mL. This is similar to the previous reports with Moutan cortex and Ageratum conyzoides plant extracts that demonstrated EC50 values ranging from 145.33 to 450 µg/mL against R. solani17,37. Quinine hydrochloride has also been shown to have antifungal properties against Candida albicans under blue light38. Cinchona bark contains bioactive compounds other than quinoline alkaloids, such as coumarins that help in cell wall inhibition of the fungal pathogen39, nitro-containing cycloalkanes that reduce reactive nitrogen species generation, Schiff bases and aniline derivatives with broad-spectrum antimicrobial activity via metal chelation and enzyme inhibition40, and imidic acid derivatives with antimicrobial potential41. Terpenoid-like chemicals present in cinchona bark contribute by affecting microbial membranes and metabolism42. The combined action of quinine and these metabolites is likely responsible for CBF’s inhibitory effect on Rhizoctonia hyphae.
Quinine has two basic nitrogen atoms, one in its quinuclidine tertiary amine (pKa:8.5) and another in its quinoline ring (pKa:4.1). At neutral pH, the N-atom in the tertiary amine gets protonated. With further acidification, the N-atom in the quinoline ring also gets protonated, forming an overall deprotonated (+ 2) form of quinine. The FTIR of both QCl and CBF showed a characteristic N+-H bond, which is generally absent in quinine. The N+-H bond may be formed due to the reaction of HCl and the formation of quinine hydrochloride, where the quinoline N is attached with the hydrogen of HCl, and the Cl− balances the positive charge of hydrogen with an electrostatic bond43. This leads to more solubility of quinine in aqueous solution (Fig. 9). However, inside the fungal cells at physiological pH (~ 7.4), partial deprotonation to neutral (Q) or monocationic (QH⁺) forms of quinine is more prevalent. In both forms, quinine is reported to be uptaken by the fungal cells by passive diffusion44. Inside acidic intracellular compartments of the fungi, quinine is re-protonated and dissociated from the chloride ion and trapped by a process known as ion trapping, which leads to intracellular accumulation and antifungal effects45.

Chemistry of formation of Hydrochloride derivatives of quinoline alkaloids (quinine, quinidine, cinchonine, and cinchonidine) in 0.1 N HCl (pH 1–3, 25–30 °C). Protonation at nitrogen centers resulted in mono- and diprotonated species. Basicity is indicated by pKa values, while Cl⁻ counterions balance created the positive charges.
Quinine-derived fungicides have been shown to damage hyphae, organelles, membranes, and induce leakage in fungi46,47. In S. cerevisiae, quinine inhibits tryptophan uptake via Tat2p48. Molecular docking in the present investigation verified quinine binding to amino acid permeases (rhAAPs) through residues ARG363, HIS36, TYR35, HIS217, and CYS193 and site directed mutations at these amino acids reducing the binding efficacy by disrupting conventional H-bonds49. QCl and CBF treatments significantly suppressed Rhizoctonia solani growth studied over 120 h, consistent with previous research demonstrating the efficacy of fungicides in managing plant diseases50. The partial recovery of fungal growth in combinatorial treatments with tryptophan shows recovery of the fungi, probably in response to environmental stresses51. The time-course study and quantification of Trp in the CBF and QCL-treated mycelia clearly indicate Trp starvation in the treated fungi. However, more direct evidence using in vitro experiments is required for further confirmation. Tryptophan helps fungal stress tolerance by acting as a precursor of melatonin/serotonin, tryptophol and a CytP450 enzyme stabilizer52,53,54. CytP450 downregulation in CBF-treated fungus with partial recovery under tryptophan suggests a connection between tryptophan starvation and xenobiotic stress. Fungi tend to compensate for the reduced amino acid transport by overexpressing TAT1, GAP1, and AGP1 permeases via the GAAC-Gcn4p pathway55,56,57. Similarly, the upregulation of rhAAPI and rhAAPII in the treated fungi supports the in-silico results, suggesting that quinine blocks rhAAPs and restricts the cell-to-cell movement of tryptophan in Rhizoctonia, causing tryptophan starvation.
CBF and QCl also lead ROS-mediated oxidative stress and disruption of MMP in Rhizoctonia. Quinoline and other bioactive compounds and their derivatives have demonstrated similar accumulation of ROS and alteration in MMP in other filamentous fungi16. Both QCl and CBF treatments significantly reduced the activity of succinate dehydrogenase (SDH), a mitochondrial membrane-bound enzyme, as evident in the time course enzyme assay. SDH is a target to estimate mitochondrial dysfunction and exogenous chemicals or fungicides are known to block SDH activity by producing oxidative stress and mitochondrial membrane damage58. Cumulative upregulation of different components of the electron transport chain, like QCR1, QCR9 (Complex III), and COX2, COX3 (Complex IV) indicated compensatory stress responses by Rhizoctonia, as shown in previous studies with Aspergillus fumigatus and Candida albicans59,60.
The application of natural bioactive formulations in the management of plant pathogens has tremendous benefits for the environment and the farming community. Quinine is one of the most valuable natural compounds that have a proven role in mitigating diseases in humans. The present study focuses on the development of chinchona-based easy and affordable formulation to manage the deadly fungus Rhizoctonia. The results of the present investigation suggest that quinine works against the fungus in a multifaceted way by tryptophan starvation and mitochondrial respiratory dysfunction. It affects tryptophan permeases, causing tryptophan scarcity in the fungal cell. In addition, application of quinine-based formulations results in the accumulation of reactive oxygen species (ROS) in the fungal cells, which depolarizes mitochondrial membranes and impairs the fungal respiratory system. These metabolic disruptions of the fungus by quinine result in fewer sclerotia and hyphae structural abnormalities. All of these findings point to the dual mode of action of quinine-rich Cinchona bark-based formulations that exert a deleterious effect on Rhizoctonia solani.
Materials and methods
Chemicals and instruments
All the chemicals and reagents utilized in this investigation were analytical or molecular biology grade. Hydrochloric acid (37%, 320331), sulfuric acid (98%, 258105), perchloric acid (70%, 244252), methanol (HPLC grade, 34860), dimethyl sulfoxide (≥ 99.9%, D8418), acetic anhydride (≥ 99%, 320102), and trifluoroacetic acid (≥ 99%, 302031) were purchased from Sigma-Aldrich as well as acetic acid glacial (Merck, 100063), were used for extraction and HPLC analyses.
For tryptophan estimation, papain (P3125), sodium acetate anhydrous (S2889), ferric chloride hexahydrate (F2877), L-tryptophan (T0254), and sodium hydroxide (≥ 98%, S5881) were purchased from Sigma-Aldrich, and nylon membrane filters 0.2 μm (NY2002500) from Millipore. For ROS and mitochondrial membrane potential assays, rhodamine 123 (Sigma-Aldrich, R8004), DCFH-DA (Sigma-Aldrich, D6883), and phosphate-buffered saline (Gibco, 10010023) were employed. The succinate dehydrogenase (SDH) activity assay utilized sodium succinate dibasic hexahydrate (Sigma-Aldrich, S2378), INT tetrazolium (Sigma-Aldrich, I8377), phenazine methosulfate (Sigma-Aldrich, P9625), sodium azide (HiMedia Laboratories, MB075), formaldehyde 37% (Merck, 104003), and dimethyl sulfoxide (Sigma-Aldrich, D8418).
For genomic DNA isolation, the following reagents were used: Tris-HCl pH 8.0 (Sigma-Aldrich, T5941), EDTA disodium salt dihydrate pH 8.0 (Sigma-Aldrich, E5134), NaCl (≥ 99.5%, Sigma-Aldrich, S9888), CTAB (≥ 99%, Sigma-Aldrich, H6269), Proteinase K (Thermo Fisher Scientific, EO0491), RNase A (Thermo Fisher Scientific, EN0531), phenol: chloroform: isoamyl alcohol 25:24:1 (Sigma-Aldrich, P2069), chloroform: isoamyl alcohol 24:1 (Sigma-Aldrich, C0549), isopropanol (≥ 99.5%, Sigma-Aldrich, I9516), ethanol absolute (≥ 99.8%, Merck, 100983), TE buffer (Thermo Fisher Scientific, AM9849), and DNA LoBind microcentrifuge tubes (Eppendorf, 022431021). Potato dextrose agar (HiMedia Laboratories, M403) and potato dextrose broth (HiMedia Laboratories, M402) were used for fungal culture. For gene expression analysis, TRIzol™ reagent (Invitrogen, 15596026), M-MLV reverse transcriptase (New England Biolabs, M0253), dNTP mix (New England Biolabs, N0447), RNaseOUT™ inhibitor (Invitrogen, 10777019), SYBR™ Green Master Mix (Applied Biosystems, 4309155), and diethylpyrocarbonate (Sigma-Aldrich, D5758) were used.
HPLC-grade chromatographic reagents and molecular-grade reagents were procured from Thermo Fisher Scientific Inc. (USA). The GC analyses were performed using a Thermo Scientific-GC-TRACE-1300 coupled with a MS analyser (Thermo MS ISQ 7000). HPLC analyses were conducted on Waters alliance-e2695 system equipped with Waters-2489 UV-VIS detector and 2475 Fluorescence (FLR) Detector. Fourier-transform infrared (FTIR) spectroscopy was carried out using Bruker Alpha FTIR spectrophotometer (Ettlingen, Germany). Microscopic analysis was performed using ZEISS Primostar 3 compound microscope (Carl Zeiss Jena, Oberkochen, Germany) and Axioscope-5 fluorescence microscope (Carl Zeiss in Germany). Gene expression analysis was performed with nanodrop spectrophotometer (biolab nd-1000) (Thermo Fisher Scientific, Scoresby, Vic, Australia) and qRT-PCR (Quant studio-5, Applied Biosystems, USA).
Sample collection
Mature bark samples were taken from Cinchona plants aged 14–16 years from a plantation in the Darjeeling district, West Bengal, India (26° 58′ 17″ N, 88° 22′ 16″ E). Following collection, the materials were completely cleaned, dried at 45 °C for approximately 48 h, and finely pulverized. The powdered samples were then stored in airtight containers, appropriately labeled, and kept dry until further investigation.
Preparation of Cinchona bark extract
In order to examine how pH affects extraction, effectiveness and phytochemical composition, cinchona bark extracts were made using a variety of solvent systems. Cinchona extract was made by combining 200 mg of finely powdered Cinchona bark with 100 mL of the solvents i.e. 0.1 N hydrochloric acid (Cinchona HCl Extract – CHE), 0.1 N sulfuric acid (Cinchona H2SO4 Extract – CSE), 0.1 N perchloric acid (Cinchona HClO4 Extract – CPE), 5% methanol (Cinchona Methanol Extract – CME), and 5% dimethyl sulfoxide (Cinchona DMSO Extract – CDE). The mixtures were incubated at 40 °C for 10 min to aid in the process, and the extracts were centrifuged at 4000 rpm for 10 min. The supernatants (2000 µg/mL) were collected for further investigation.
To prepare the acidic solvents, 0.83 mL of concentrated hydrochloric acid (about 37%, density = 1.18 g/mL) was diluted with 100 mL of double-distilled water to make 0.1 N HCl. Similarly, 0.1 N H2SO4 was created by adding 0.28 mL of concentrated sulfuric acid (∼98%, density ≈ 1.84 g/mL) to 100 mL of distilled water, and 0.1 N HClO4 was prepared by diluting 0.81 mL of perchloric acid (≈ 70%, density ≈ 1.67 g/mL) to 100 mL with distilled water. The 5% methanol and 5% DMSO solutions were made by combining 5 mL of methanol or DMSO with 95 mL of distilled water.
The natural aqueous extract of Cinchona bark has a mildly acidic to near-neutral pH, ranging from 5.5 to 6.5. The addition of 0.1 N hydrochloric acid (CHE) lowers the extract pH to around 0.8–1.5, boosting the protonation of alkaloids like quinine and increasing their solubility in aqueous environments through salt formation. Extracts made using sulfuric acid (CSE, pH ~ 1.0–1.3.0.3) and perchloric acid (CPE, pH ~ 0.8–1.2) exhibit comparable pH ranges. In contrast, methanol (CME) and DMSO (CDE) extracts have larger pH ranges, typically 5.5–6.5 and 6.5–7.2, respectively.
Plant pathogenic fungi
Infected parts of rice plants were collected from standing crops in Alipurduar district of West Bengal, India (26⁰ 50՛96՛՛N and 89⁰ 4՛22՛՛E), an area known for its prevalence of sheath blight disease. The samples were carefully transported to the laboratory, where they were sub-cultured on potato dextrose agar (PDA) and incubated at 28 °C to obtain pure fungal cultures.
Genomic DNA extraction methods
To isolate genomic DNA, 200 mg of lyophilized Rhizoctonia solani mycelial mat was ground into a fine powder using a sterile pestle and mortar and 500 µL of extraction buffer based on the CTAB-phenol-chloroform-isoamyl alcohol (CTABPCI) method61. The extraction buffer consisted of 200 mM Tris-HCl (pH 8.0), 25 mM EDTA (pH 8.0), 250 mM NaCl, and 10% CTAB. After homogenizing the sample, the extract was transferred to microcentrifuge tube with addition of 3 µL of proteinase K and 3 µL of RNase. The contents were briefly vortexed before being incubated at 37 °C for 1 h to allow for enzymatic digestion of proteins and RNA. After incubation, the tubes were heated in a water bath at 65⁰ C for 10 min to increase cell lysis and DNA release. After lysis, one volume of phenol, chloroform, and isoamyl alcohol (25:24:1) was added to the lysate, well mixed for 5 min, then centrifuged at 12,000 rpm for 5 min. The aqueous phase was carefully extracted and combined with an equal volume of chloroform: isoamyl alcohol (24:1), then centrifuged under the same circumstances. The aqueous layer was collected and combined with one volume of ice-cold isopropanol. It was then refrigerated at −20 °C overnight for DNA precipitation. The precipitated DNA was centrifuged at 10,000 rpm for 5 min before being rinsed with 100% ethanol to eliminate any leftover salts or contaminants. The DNA pellet was rinsed twice with 1 mL of 70% ethanol, air-dried quickly, and then resuspended in 200 µL of either deionized water or 1X TE buffer [200 mM Tris-HCl (pH 8.0), 20 mM EDTA (pH 8.0)] for downstream applications like PCR and sequencing.
In vitro antifungal bioassay and calculation of EC50
Poison food assay
The poisoned food technique was used to assess the antifungal activity of various Cinchona bark extracts on Rhizoctonia solani62. The extracts were combined with potato dextrose agar (PDA), and 20 mL of the mixture was placed into sterile Petri dishes (60 × 15 mm) and allowed to harden under a laminar airflow hood. A 5 mm diameter mycelial disc of actively growing R. solani was inserted in the center of each plate. The extracts were evaluated at doses of 10 to 500 µg/mL. Plates containing quinine dissolved in 0.1 N hydrochloric acid (QCl: Quinine HCl solution) were used as positive control to assess its antifungal efficacy, whereas plates containing only 0.1 N hydrochloric acid (HCl) without quinine were used as solvent control to evaluate the solvent’s effect alone. All plates were incubated at 28 °C, and fungal growth was assessed following complete radial expansion on control plates. The extent of mycelial growth on the treated plates was recorded once the radial expansion of the fungal mycelium completely covered the control plates. The antifungal efficacy of the treatments was quantified using a specific formula.
% of inhibition = C-T/C ×100.
Where, C = The diameter of the fungus colony formed on the medium without any antifungal or experimental treatment. (cm), T = diameter of fungal disc in treated plates.
EC50 value
The EC50 of various treatments against Rhizoctonia solani was obtained using the poison food technique63. Cinchona bark extracts (QCl, CSE, CPE, CWE, CDE, and CME) were mixed onto potato dextrose agar (PDA) plates at different concentrations (10, 50, 100, 250, and 500 µg/mL). In each treatment, 20 mL of the mixture was put into sterile Petri dishes (60 × 15 mm), solidified in a laminar flow hood, and then inoculated at the center with a 5 mm mycelial disc froming an actively growing R. solani culture. Plates containing only 0.1 N hydrochloric acid were used as solvent controls, while untreated plates served as negative controls. All treatments were performed in duplicate and repeated three times to ensure accuracy. The plates were incubated at 28 °C until the mycelium had completely covered the control plates. The radial growth of fungal cells was recorded, and the % inhibition was calculated as follows:
% Inhibition = [(Control growth − Treated growth)/Control growth] × 100.
The inhibitory data were translated into a linear dose-response relationship using probit analysis. These modified values were then regressed against the log10-transformed concentrations of the Cinchona extract with IBM SPSS Statistics version 21.0. The EC50 values for each solvent treatment were calculated using the regression curve at the probit zero point, which represents the concentration that inhibited fungal growth by 50%. This method provided a consistent means to compare the effectiveness of the Cinchona extract across different solvent systems and concentration ranges.
Bioactive compound profiling of CBF
GC-MS
Cinchona bark was extracted by dissolving 1 g dried powder in 20 mL of 0.1 N HCl (CBF), shaken at 200 rpm for 12 h. The mixture was centrifuged at 4000 rpm for 10 min and 200 µL of the supernatant was diluted with 4 mL methanol. This solution was injected into GC-MS coupled with TR-1MS capillary column (30 m × 0.25 mm ID × 0.25 μm film thickness) with helium as carrier gas (1mL/min). Oven temperature was programmed from 50 °C to 250 °C at 8 °C/min. Data acquisition and analysis were performed using Chromalion 7 software. Spectra (m/z 35–700) were compared against the NIST 20 Mass spectral Library64.
RP-HPLC
RP-HPLC analysis for detection and quantification of quinine in CBF was performed by the method given by Jeadkong et al., 201765. 1 mL of CBF (200 µg/mL) was diluted with 5 mL of methanol, filtered with 0.22 μm syringe filter and 10 µL of the filtrate was injected into HPLC system. Sepeartion of analyte was performed on a C18 column (4.6 × 150 mm, Symmetry-Waters) using an isocratic mobile phase of ammonium acetate (pH 7), acetonitrile and methanol in a ratio of 40:25:35. Instrument settings included a flow rate of 1 mL/min, pressure between 1560 and 1590 psi, column temperature at 25 °C and detection wavelength of 330 nm. A calibration curve was prepared from standard quinine solutions (1 to 100 µg/mL).
FTIR
Cinchona bark powder was first made as a 5000 µg/mL extract by soaking 500 mg of finely powdered bark in 100 mL of 0.1 N hydrochloric acid (HCl). This mixture was incubated at 40 °C for 10 min, cooled to room temperature, and centrifuged at 4000 rpm for 10 min to produce a clear supernatant. To do FTIR analysis, gently apply 5–10 µL of the solution to the Platinum ATR crystal surface of a Fourier-transform infrared spectrophotometer. Kumar et al. (2015) detailed the acquisition of spectral data under inert atmospheric conditions spanning a wavenumber range of 5000–400 cm−1, which allowed for comprehensive identification of distinctive stretching and bending vibrations66.
Tryptophan supplementation
R. solani inhibition by QCl and CBF were studied for retrieval in fungal media through external supply of L-tryptophan. Potato Dextrose Agar and Potato Dextrose Broth were prepared separately at the chosen EC50 values of QCl and CBF. L-tryptophan concentration of 10 to 500 µg/mL was added during the preparation of respective extracts and added into the media. These extracts enriched with tryptophan were named as CBF + Trp (Cinchona Extract in 0.1 N HCl with tryptophan) and QCl + Trp (Quinine in 0.1 N HCl with tryptophan). Plates with only QCl (50 µg/mL) and CBF (200 µg/mL) were used as treatment controls, with untreated plates serving as negative controls.
Morphological parameters of fungal growth
R. solani inoculated in PDA and PDB were subjected to measure total dry weight (mg) and sclerotia weight (mg) incubated at 28 ± 1 °C for 7 days. Observations on fungal dry weight for Control, QCl, CBF, CBF + Trp and QCl + Trp treated media were recorded after complete removal of water from total fungal biomass at 40 °C for 12 h. Sclerotia from each petri dishes were collected and surface water were removed by blotting paper.
Micro-Morphological characterization
For the micro-morphological examination, the mycelia were collected, repeatedly washed with PBS buffer, spread over a glass slide, and covered with a coverslip. The slides were mounted on a compound microscope for the detection of mycelial abnormalities. The samples were observed under a magnification of 10X.
In vivo antifungal assay
R. solani inoculum was prepared in PDB (Potato Dextrose Broth) and incubated at 28 °C for 10 days until a mycelial mat with sclerotia was formed. The mat was mixed with autoclaved vermicompost and incubated at 25 ± 1 °C for 5 to 7 days. Inoculated vermicompost was then mixed with sieved soil in 1:4 ratio to prepare the soil medium. Seeds of the sheath blight susceptible rice variety “Swarna” were sown in seed germination trays (50 cm × 30 cm ×10 cm) filled with prepared inoculated soil media. Different prepared extracts viz. CBF (200 µg/mL), QH (50 µg/mL), CBF + Trp (400 µg/mL tryptophan) and QCl + Trp (400 µg/mL tryptophan) were sprayed every 5 days intervals up to 21 days. Trays were photographed on 10- and 18-days post sowing and infection was analyzed using ImageJ (v1.8). The photos were converted to binary (black and white) format to distinguish between infected (black) and healthy (white) areas. The percentage of infected area (infection severity) was estimated using the following formula:
$$:Infection:severity=frac{Infected:left(blackright)pixels:for:area:of:interest}{Total:pixels:in:area:of:interest}times:100$$
In silico molecular docking of quinine and Tryptophan with amino acid permease (AAP)
BOILED-Egg model in SwissADME was used to predict the membrane permeability of quinine for lipophilicity and membrane diffusion analysis67. Homology modeling of both the Rhizoctonia amino permease I and II proteins (rhAAP-I and rhAAP-II) were conducted using SWISS-MODEL68 and their secondary structure were predicted with PSIPRED (Data Partition and Semi-Random Subspace Method)69. Membrane localization of rhAAP-I and rhAAP-II was analyzed using DeepLoc 2.070 multi-label subcellular localization prediction using protein language models) while transmembrane helix prediction was performed using TMHMM (Hidden Markov model)71. Molecular docking studies were carried out in Biovia Discovery Studio client version, 2023 and PyRx (AutoDock vina) to access the binding interactions of quinine and tryptophan with permeases for analysis of affinities and interaction72. Molecular docking was also used to explore the interactions between quinine and two proteins, rhAAP-I and rhAAP-II, using AutoDock Vina. Key residues in the binding locations were chosen for docking. Mutations (ARG363 to ALA363, HIS36 to ALA36, TYR35 to PHE35 in rhAAP-I; HIS217 to ALA217, CYS193 to ALA193 in rhAAP-II) were introduced using Discovery Studio, and energy minimization was performed before docking. Mutations’ effects on ligand binding stability and affinity were assessed using binding efficiency, RMSD, and interaction types. The results were shown using Discovery Studio and PyRx73.
Time course study for fungal growth
The mycelial growth of Rhizoctonia solani was evaluated using potato dextrose broth (PDB; HiMedia Laboratories, India). Actively growing mycelial plugs (5 mm in diameter) were inoculated into 50 mL of sterile PDB in 100 mL conical flasks with several treatments: QCL, CBF, QCL + Trp, CBF + Trp, and an untreated Control. All cultures were kept at 28 ± 2 °C under static conditions. Fungal growth was tracked by measuring the optical density (OD) at 405 nm with a UV-Visible spectrophotometer (Shimadzu, Japan) at every 5-hour intervals over 120 h, to monirtor fungal growth and inhibition in the treated and control samples throughout the growth cycle74.
Determination of Tryptophan and quinine from R. solani
Fungal mycelium was grown for five days in liquid broth containing treatments (QCl, QCl + Trp, CBF and CBF + Trp) were collected, filtered twice through muslin cloth and dried at 40 °C.The resultant powder was subjected to total tryptophan estimation.
Colorimetric estimation of Tryptophan
100 mg of defatted fungal powder was digested in 5 mL papain solution (4000 µg/mL in 0.1 N sodium acetate buffer, pH 7) at 65 °C for 12 h. After centrifugation (10,000 rpm for 10 min), 1 mL supernatant was mixed with reagent C prepared freshly by mixing equal volumes of Reagent A (135 mg FeCl₃·6 H2O in 0.25 mL water diluted to 500 mL with glacial acetic acid + 2% acetic anhydride) and Reagent B (30 N H2SO4). The mixture was incubated at 65 °C for 15 min producing an orange-red color appearance. Absorbance was measured at 545 nm using papain solution as blank. Tryptophan standards (1–100 µg/mL) were used for calibration75.
HPLC quantification of Tryptophan
100 mg dried fungal powder was hydrolyzed with 3 mL alkaline solution of 4 M NaOH and incubated at 110 °C for 20 h. After cooling, the hydrolysate was washed thrice with 5 mL 0.1 N HCl. pH was adjusted to 6.3 with 3 N HCl and diluted to 50 mL with acidic water. The solution was centrifuged for 2 min (1000 rpm, 4 °C) and filtered through a 0.2 μm nylon membrane. A 200 µL aliquot was diluted in 1:6 ratio with double distilled water for analysis. Separation was done with C18 column (4.6 × 250 mm, 5 μm) and fluorescence detector (excitation 280 nm, emission 340 nm). The mobile phase was 0.1% trifluoroacetic acid and methanol (75:25 v/v), isocratic flow at 1 mL/min, column temperature 30 °C with total run time of 10 min. The injection volume was 10 µL. tryptophan standard concentration of 10 to 100 µg/mL was prepared and all the results were procured from Empower software76.
Estimation of quinine in fungus cell by HPLC
QCl and CBF were added to Potato Dextrose Broth (PDB) at 50 and 200 µg/mL concentrations, respectively. R. solani cultures were added to the medium and incubated at 28 °C for 5 days. The fungal biomass was then removed from media washed with double distilled water and recovered by filtering through autoclaved muslin fabric and drying at 45 °C for 6 h to extract moisture. The dried samples were powdered using mortar and pestle and analyzed for total quinine content using RP-HPLC.
Estimation of reactive oxygen species and mitochondrial membrane potential in fungal mycelia
Mycelium from 3-day-old Rhizoctonia solani cultures, both control and treatment, was stained with fluorescent dyes to determine the buildup of reactive oxygen species (ROS) and mitochondrial membrane potential (MMP). Rhodamine 123 at a concentration of 1 µM was employed to measure mitochondrial membrane potential. Rhodamine 123 is a cationic fluorescent dye that accumulates specifically in mitochondria due to their negative membrane potential. Healthy mitochondria with intact membrane potential have strong fluorescence due to considerable dye accumulation, whereas depolarized or injured mitochondria have lower fluorescence intensity. Zhang et al. (2018) outlined the process of incubating samples with 500 µL of rhodamine 123 and washing with PBS77.
To detect ROS, 10 µM DCFH-DA (2′,7′-dichlorodihydrofluorescein diacetate) was utilized as a probe. DCFH-DA is a cell-permeable, nonfluorescent molecule that, once inside the fungal mycelium, is deacetylated by cellular esterases to produce DCFH. In the presence of ROS, DCFH oxidizes fast to highly fluorescent DCF (Dichlorofluorescein), allowing ROS to be visualized and quantified using green fluorescence intensity. Kim and Xue (2020) reported that samples were treated with 500 µL of DCFH-DA at 37 °C for 30 min before being rinsed with PBS78.
Finally, stained mycelia were examined using an Axioscope-5 fluorescent microscope (Carl Zeiss, Germany), and the resulting pictures were processed using ImageJ (version 1.8) to determine the Relative fluorescent Unit (RFU) for each treatment. The normalized Relative Fluorescence Unit (RFU) is the mean green value of the Region of Interest (ROI) from imageJ.
Succinate dehydrogenase (SDH) activity
The activity of succinate dehydrogenase (SDH) in Rhizoctonia solani was measured using a spectrophotometric assay that involves the reduction of 2-(p-iodophenyl)−3-(p-nitrophenyl)−5-phenyltetrazolium chloride (INT) to formazan79. Fungal biomass was measured at 0, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, and 120 h in cultures treated with Control, QCl, CBF, QCl + Trp, and CBF + Trp. Approximately 1 mg of dried mycelial biomass was mixed with 1 mL of a reaction solution containing 0.5 M sodium succinate and 0.68 mM INT. Phenazine methosulfate (PMS) served as an electron carrier, whereas sodium azide (0.1% w/v) prevented cytochrome oxidase interference. The reaction mixture was incubated at 37 °C for 60 min before being stopped by the addition of 0.4 mL of 37% formaldehyde.
Formazan crystals formed during the reaction were extracted with 1 mL of dimethyl sulfoxide (DMSO) and vortexed until fully dissolved. The absorbance of the solution was measured at 570 nm with a UV-visible spectrophotometer. The concentration of INT-formazan (C) was estimated using Beer-Lambert’s equation, preserving absorbance at 570 nm, extinction coefficient of INT-formazan (15,000 M−1cm−1), and 1 cm path length.
The concentration was converted to mass using the equation: Mass (µg) = C×V×MW.
Where: V is the assay volume (0.001 L), MW is the molecular weight of INT-formazan (466 g/mol).
Finally, SDH activity was expressed as:(:frac{Mass:of:formazan:left(mu:gright)}{mg:of:dry:mass:left(mgright)times:time:left(text{m}text{i}text{n}right)})
SDH activity was expressed as micrograms of INT-formazan per gram of dry biomass per minute (µg INT-formazan g−1 min−1).
Differential gene expression analysis
Rhizoctonia solani AG1-IA was grown in cultures on a stationary liquid medium and collected using forceps. This harvested biomass of fungal cells was centrifuged at 16,000 g for 5 minutes80.The tubes and bottles were treated with 0.1% DEPC solution at 37 °C overnight, autoclaved twice at 121 °C for 20 min and then dried at 100 °C before use. The harvested fungal material from control and treatments was ground to a fine powder in liquid N2. The RNA was isolated using TRIZOL regent following Schumann et al., 201381 The isolated RNA was reverse transcribed into cDNA by M-MLV reverse transcriptase enzyme (New England Biolabs). Relative expression of rhAAP-I and rhAAP-II (Amino Acid Permeases), QCR1 and QCR9 (components of ubiquinol-cytochrome C reductase, Complex III) as well as COX2 and COX3 (components of Cytochrome c oxidase, Complex IV) genes was estimated using Realtime PCR (Quant studio-5, Applied Biosystems, USA). To normalize gene expression data in Rhizoctonia solani, we employed β-actin, a stable housekeeping gene under many experimental circumstances82. This normalization procedure is required to account for differences in RNA quantity, quality, and reverse transcription efficiency among samples. The relative expression levels were estimated with the 2^(-ΔΔCt) technique. To determine ΔCt for each sample, the Ct values for each gene are normalized to the Ct value of the housekeeping gene (β-actin) The ΔCt of treated and control samples is compared to calculate ΔΔCt. The fold change is calculated by raising 2 to the power of -ΔΔCt, indicating the proportionate increase or decrease in gene expression compared to the control. This method provides a stable and reproducible way for quantifying gene expression changes across multiple treatments. A list of primer sequences for different genes used in the qPCR study is provided in Supplementary information (Table S1).
Statistical analysis
All experiments were conducted with a minimum of three replicates unless specified. Error bars indicated the Standard Deviation (SD) of the mean and presented as Mean ± SEM. Inhibition percentage, sclerotia weight and gene expression analysis was performed using one-way ANOVA followed by t-test. Statistical significance is denoted by asterisks, with three stars indicating p < 0. 001. Statistical analysis of HPLC and colorimetric data were analyzed by two-way ANOVA followed by Tukey’s test with error bars representing significant differences (p < 0.001). All graphs were generated by using OriginPro 2024 (https://www.originlab.com). Using the “Statistics on Columns” tool, descriptive statistical analysis was carried out to derive the mean, standard deviation (SD), standard error (SE), and 95% confidence intervals (CI). The standard error was obtained by dividing the SD by the square root of the number of replicates (n = 3). The 95% confidence interval was calculated as CI = Mean ± (t × SE), with the t-value derived using the Student’s t-distribution with two degrees of freedom. The data were displayed graphically, with error bars denoting confidence ranges to highlight measurement variability and dependability.
Data availability
Data supporting the findings of this work are available within this paper and its Supplementary Information files. Additional datasets used during the current study available from the corresponding author on reasonable request. *Rhizoctonia solani* strain AG-1-IA internal transcribed spacer (ITS) region and partial LSU rRNA gene sequence, amplified using ITS1 and ITS4 primers were submitted to the NCBI GenBank with accession no. PV220927.1 (https://www.ncbi.nlm.nih.gov/nuccore/PV220927.1/). The models generated through docking are available in Model Archive (modelarchive.org) with the accession codes *ma-lwg2r* (https://modelarchive.org/doi/10.5452/ma-lwg2r) and *ma-5kjcj* (https://modelarchive.org/doi/10.5452/ma-5kjcj) under Swiss Institute of Bioinformatics.
References
-
Rai, S. & Rai, A. Wartime history of Cinchona and its cultivation in Darjeeling. Esmeralda, The Orchid Society of Eastern Himalaya & The Directorate of Cinchona & Other Medicinal Plants 1, 72–80 (2015).
-
Dubey, A. & Singh, Y. Medicinal properties of Cinchona alkaloids—A brief review. Asian J. Pharm. Sci. 11, 2231–5659. https://doi.org/10.52711/2231-5659.2021.00036 (2021).
-
Al-Harbi, L. N. et al. Cinchona officinalis phytochemicals-loaded iron oxide nanoparticles induce cytotoxicity and stimulate apoptosis in MCF-7 human breast cancer cells. Nanomater 12, 3393. https://doi.org/10.3390/nano12193393 (2022).
-
Hariyanti, R. et al. A review: Pharmacological activities of Quinoline alkaloid of Cinchona Sp. Biointerface Res. Appl. Chem. 13, 1–13. https://doi.org/10.33263/briac134.319 (2023).
-
de Villiers, K. A. & Egan, T. J. Heme detoxification in the malaria parasite: a target for antimalarial drug development. Acc. Chem. Res. 54, 2649–2659. https://doi.org/10.1021/acs.accounts.1c00207 (2021).
-
Khozoie, C., Pleass, R. J. & Avery, S. V. The antimalarial drug quinine disrupts Tat2p-mediated Tryptophan transport and causes Tryptophan starvation. J. Biol. Chem. 284, 17968–17974. https://doi.org/10.1074/jbc.M109.005843 (2009).
-
Islahudin, F., Khozoie, C., Bates, S., Ting, K. N. & Avery, S. V. Cell wall perturbation sensitizes fungi to the antimalarial drug chloroquine. Antimicrob. Agents Chemother. 57, 3889–3896. https://doi.org/10.1128/AAC.00478-13 (2013).
-
Rosenthal, P. J. & Miller, L. H. Antimalarial chemotherapy: mechanisms of action, resistance, and new directions in drug discovery. Infect. Dis., Springer, New Jersey. (2001). https://doi.org/10.1007/978-1-59259-111-4
-
Eyal, S. The fever tree: from malaria to neurological diseases. Toxins 10, 491. https://doi.org/10.3390/toxins10120491 (2018).
-
Xiao, H. et al. Cinchona alkaloid derivative-catalyzed enantioselective synthesis via a Mannich-type reaction and antifungal activity of β-amino esters bearing benzoheterocycle moieties. Mol 19, 3955–3972. https://doi.org/10.3390/molecules19043955 (2014).
-
Chen, Y. J. et al. Antifungal exploration of Quinoline derivatives against phytopathogenic fungi inspired by quinine alkaloids. J. Agric. Food Chem. 69, 12156–12170. https://doi.org/10.1021/acs.jafc.1c05677 (2021).
-
Završnik, D., Muratović, S., Špirtović, S., Softić, D. & Medić-Šarić, M. Evaluation of antimicrobial activity of some Quinoline derivatives. Bosn J. Basic. Med. Sci. 8, 277–281 (2008).
-
Vallieres, C., Raulo, R., Dickinson, M. & Avery, S. Novel combinations of agents targeting translation that synergistically inhibit fungal pathogens. Front. Microbiol. 9, 2355. https://doi.org/10.3389/fmicb.2018.02355 (2018).
-
Dawidowicz, L. et al. Possibility of quinine transformation in food products: LC–MS and NMR techniques in analysis of quinine derivatives. Eur. Food Res. Technol. 244, 105–116. https://doi.org/10.1007/s00217-017-2940-0 (2018).
-
Shang, X. F. et al. Integrated proteomics and transcriptomics analyses reveals the possible antifungal mechanism of an indoloquinoline alkaloid neocryptolepine against Rhizoctonia Solani. J. Agric. Food Chem. 69, 6455–6464. https://doi.org/10.1021/acs.jafc.1c01385 (2021).
-
Chintala, S. et al. Turmeric (Curcuma longa L.) rhizome extract mediated silver nanoformulation exhibits enhanced antifungal property against Rhizoctonia Solani and boosts innate immunity of rice. Ind. Crops Prod. 206, 117616. https://doi.org/10.1016/j.indcrop.2023.117616 (2023).
-
Zhao, Y. et al. Inhibitory activity against Rhizoctonia Solani and chemical composition of extract from Moutan cortex. Chem. Biodivers. 21, e202400337. https://doi.org/10.1002/cbdv.202400337 (2024).
-
Prakasam, V. et al. Sheath blight of rice and its management. Tech. Bull. No. 72, Directorate of Rice Research (ICAR), Rajendra Nagar (2013).
-
Cheng, X. et al. Cytochrome P450 and glutathione S-transferase confer metabolic resistance to SYP-14288 and multi-drug resistance in Rhizoctonia Solani. Front. Microbiol. 13, 806339. https://doi.org/10.3389/fmicb.2022.806339 (2022).
-
Sierotzki, H. & Scalliet, G. A review of current knowledge of resistance aspects for the next-generation succinate dehydrogenase inhibitor fungicides. Phytopathol 103, 880–887. https://doi.org/10.1094/PHYTO-01-13-0009-RVW (2013).
-
Bhandari, S., Yadav, P. K. & Sarhan, A. T. Botanical fungicides; current status, fungicidal properties, and challenges for wide-scale adoption: a review. Rev. Food Agric. 2, 63–68. https://doi.org/10.26480/rfna.02.2021.63.68 (2021).
-
Yoon, M. Y., Cha, B. & Kim, J. C. Recent trends in studies on botanical fungicides in agriculture. Plant. Pathol. J. 29, 1–9. https://doi.org/10.5423/PPJ.RW.05.2012.0072 (2013).
-
Senapati, M. et al. Rhizoctonia Solani Kühn pathophysiology: status and prospects of sheath blight disease management in rice. Front. Plant. Sci. 13, 881116. https://doi.org/10.3389/fpls.2022.881116 (2022).
-
Kursa, W., Jamiołkowska, A., Wyrostek, J. & Kowalski, R. Antifungal effect of plant extracts on the growth of the cereal pathogen Fusarium spp.—an in vitro study. Agron 12, 3204. https://doi.org/10.3390/agronomy12123204 (2022).
-
Ray, K., Roy Barman, A., Sen, K., Dutta, S. & Ray, S. Molecular variability and host range of Rhizoctonia Solani AG1-IA in rice–vegetable-based cropping systems in West Bengal, India. J. Plant. Pathol. https://doi.org/10.1007/s42161-022-01283-z (2022).
-
Ahamad, F. & Khan, M. R. Incidence of sheath blight in irrigated rice and associated yield losses in Northern India. Plant. Dis. 107, 2907–2915 (2023).
-
Choudhary, P. et al. A rapid colorimetric LAMP assay for detection of Rhizoctonia Solani AG-1 IA causing sheath blight of rice. Sci. Rep. 16(1), 22022. https://doi.org/10.1038/s41598-020-79117-0 (2020).
-
Lee, D. Y. et al. Comparative genome analyses of four rice-infecting Rhizoctonia Solani isolates reveal extensive enrichment of homogalacturonan modification genes. BMC Genom. 22, 242. https://doi.org/10.1186/s12864-021-07549-7 (2021).
-
Kaushik, A. et al. Pangenome analysis of the soilborne fungal phytopathogen Rhizoctonia Solani and development of a comprehensive web resource: RsolaniDB. Front. Microbiol. 13, 839524. https://doi.org/10.3389/fmicb.2022.839524 (2022).
-
Peng, J. W. et al. Design, synthesis, and structure–activity relationship of Quinazolinone derivatives as potential fungicides. J. Agric. Food Chem. 69, 4604–4614. https://doi.org/10.1021/acs.jafc.0c05475 (2021).
-
Yang, G. Z. et al. Design, synthesis, and antifungal evaluation of novel Quinoline derivatives inspired from natural quinine alkaloids. J. Agric. Food Chem. 67, 11340–11353. https://doi.org/10.1021/acs.jafc.9b04224 (2019).
-
Warhurst, C., Craig, J. C., Adagu, I. S., Meyer, D. J. & Lee, S. Y. The relationship of physico-chemical properties and structure to the differential antiplasmodial activity of the Cinchona alkaloids. Malar. J. 2, 26. https://doi.org/10.1186/1475-2875-2-26 (2003).
-
Gong, C. et al. Status of fungicide resistance and physiological characterization of Tebuconazole resistance in Rhizoctonia Solani in Sichuan Province, China. Curr. Issues Mol. Biol. 44, 4859–4876. https://doi.org/10.3390/cimb44100330 (2022).
-
Arabiat, S. & Khan, M. F. R. Sensitivity of Rhizoctonia Solani AG-2-2 from sugar beet to fungicides. Plant. Dis. 100, 2427–2434. https://doi.org/10.1094/PDIS-01-16-0113-RE (2016).
-
Lei, Z. W., Yao, J., Liu, H., Ma, C. & Yang, W. Synthesis and bioactivity of novel sulfonate scaffold-containing Pyrazolecarbamide derivatives as antifungal and antiviral agents. Front. Chem. 10, 928842. https://doi.org/10.3389/fchem.2022.928842 (2022).
-
Sun, S. et al. Use of lentinan and fluopimomide to control cotton seedling damping-off disease caused by Rhizoctonia solani. Agriculture 12, 75. https://doi.org/10.3390/agriculture12010075 (2022).
-
Nguyen, C. et al. Antifungal activities of ageratum conyzoides L. extract against rice pathogens pyricularia oryzae and rhizoctonia Solani. Agriculture 11, 1169. https://doi.org/10.3390/agriculture11111169 (2021).
-
Leanse, L. L., Goh, X. S. & Dai, T. Quinine improves the fungicidal effects of antimicrobial blue light: implications for the treatment of cutaneous candidiasis. Lasers Surg. Med. 51, 745–753. https://doi.org/10.1002/lsm.23180 (2019).
-
Koenig, H. N. et al. Novel nitro-heteroaromatic antimicrobial agents for the control and eradication of biofilm-forming bacteria. Antibiotics 10, 855. https://doi.org/10.3390/antibiotics10070855 (2021).
-
Noriega Medellín, S. et al. The diverse biological activity of recently synthesized nitro compounds. Pharmaceuticals 15, 717. https://doi.org/10.3390/ph15060717 (2022).
-
Patel, S. & Shah, M. Schiff bases and their metal complexes: antimicrobial applications and mechanisms. Chem. Biol. Drug Des. 97, 523–536. https://doi.org/10.1111/cbdd.13762 (2021).
-
Al-Ghamdi, H. A. et al. Synthesis and biological evaluation of novel imidazole derivatives as antimicrobial agents. Biomolecules 14, 1198. https://doi.org/10.3390/biom14091198 (2024).
-
Mahizan, N. A. et al. Terpene derivatives as a potential agent against antimicrobial resistance (AMR) pathogens. Mol 24, 2631. https://doi.org/10.3390/molecules24142631 (2019).
-
Muhtadi, J., Loutfy, M. A. & Hassan, M. M. A. Quinine hydrochloride. In: Florey, K. (Ed.), Analytical Profiles of Drug Substances, vol. 12, 547–621. Academic Press, New York. https://doi.org/10.1016/S0099-5428(08)60176-8 (1983).
-
Kitagawa, T., Matsumoto, A., Terashima, I. & Uesono, Y. Antimalarial quinacrine and chloroquine lose their activity by decreasing cationic amphiphilic structure with a slight decrease in pH. J. Med. Chem. 64, 3885–3896. https://doi.org/10.1021/acs.jmedchem.0c02056 (2021).
-
Crawford, J. H. The solubility of quinine hydrochloride. Br. Med. J. 1, 874. https://doi.org/10.1136/bmj.1.3205.874 (1922).
-
El Shafey, R. A. S. et al. Morphological, pathological, and molecular characterization of rice sheath blight disease causal organism Rhizoctonia Solani AG-1 IA in Egypt. Arch. Phytopathol. Plant. Prot. 52, 1–23. https://doi.org/10.1080/03235408.2019.1650544 (2019).
-
Amano, K. et al. Hyperactive mutation occurs adjacent to the essential glutamate 286 for transport in the yeast Tryptophan permease Tat2. Biochem. Biophys. Res. Commun. 509, 1047–1052. https://doi.org/10.1016/j.bbrc.2019.01.038 (2019).
-
Chen, D. et al. Regulation of protein–ligand binding affinity by hydrogen bond pairing. Sci. Adv. 2, e1501240. https://doi.org/10.1126/sciadv.1501240 (2016).
-
Janeczko, M., Kochanowicz, E., Górka, K. & Skrzypek, T. Quinalizarin as a potential antifungal drug for the treatment of Candida albicans fungal infection in cancer patients. Microbiol. Spectr. 12, e0365223. https://doi.org/10.1128/spectrum.03652-23 (2024).
-
Aruanno, M., Bachmann, D., Sanglard, D. & Lamoth, F. Link between heat shock protein 90 and the mitochondrial respiratory chain in the caspofungin stress response of Aspergillus fumigatus. Antimicrob. Agents Chemother. 63, e00208-19. https://doi.org/10.1128/AAC.00208-19 (2019).
-
Tan, D.-X.; Manchester, L. C.; Liu, X.; Rosales-Corral, S. A.; Acuna-Castroviejo, D.; Reiter, R. J. Mitochondria and Chloroplasts as the Original Sites of Melatonin Synthesis: A Hypothesis Related to Melatonin’s Primary Function and Evolution in Eukaryotes. J. Pineal Res. 2013, 54 (2), 127–138. https://doi.org/10.1111/jpi.12026.
-
Han, Z.; Wang, J.; Ding, Y.; Sun, Z.; Wang, Y.; Wang, Y.; Yang, L.; Wang, Y. New Observation in Biocontrol of Penicillium caperatum against Fusarium oxysporum on Saposhnikovia divaricata and as a Plant Growth Promoter. Fermentation 2023, 9 (4), 361. https://doi.org/10.3390/fermentation9040361.
-
Ravanfar, R.; Sheng, Y.; Gray, H. B.; Winkler, J. R. Tryptophan Extends the Life of Cytochrome P450. Proc. Natl. Acad. Sci. U.S.A. 2023, 120 (50), e2317372120. https://doi.org/10.1073/pnas.2317372120 (52-54)
-
Braus, G. H. Aromatic amino acid biosynthesis in the yeast Saccharomyces cerevisiae: A model system for the regulation of a eukaryotic biosynthetic pathway. Microbiol. Rev. 55, 349–370. https://doi.org/10.1128/mr.55.3.349-370.1991 (1991).
-
Iraqui, I., Vissers, S., André, B. & Urrestarazu, A. Transcriptional induction by aromatic amino acids in Saccharomyces cerevisiae. Mol. Cell. Biol. 19, 3360–3371. https://doi.org/10.1128/MCB.19.5.3360 (1999).
-
Albrecht, G., Mösch, H. U., Hoffmann, B., Reusser, U. & Braus, G. H. Monitoring the Gcn4 protein-mediated response in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 273, 12696–12702. https://doi.org/10.1074/jbc.273.21.12696 (1998).
-
Zhang, S.-W., Wu, Z.-M., Yang, Y. & Li, K.-T. Antifungal action of antifungalmycin N2 against Rhizoctonia solani by disrupting cell membrane and inhibiting succinate dehydrogenase. Curr. Microbiol. 77, 254–260. https://doi.org/10.1007/s00284-019-01837-y (2020).
-
Mavrianos, E. et al. Mitochondrial two-component signaling systems in Candida albicans. Eukaryot. Cell 12, 913–922. https://doi.org/10.1128/EC.00048-13 (2013).
-
Qin, Y., Wang, J., Lv, Q. & Han, B. Recent progress in research on mitochondrion-targeted antifungal drugs: a review. Antimicrob. Agents Chemother. 67, e00003-23. https://doi.org/10.1128/aac.00003-23 (2023)
-
Li, X.-L. & Yao, Y.-J. Revision of the taxonomic position of the Phoenix 9 mushroom. Mycotaxon 91, 61–73 (2005).
-
Chaudhary, D., Khan, M.S., Prabhat, S.A. & Prasad, Y.A. Antifungal activity of three different ethanolic extracts against isolates from diseased rice plant. J. Anal. Tech. Res. 1, 47–63 https://doi.org/10.26502/jatri.007 (2019).
-
Lehner, M. S. et al. Fungicide sensitivity of Sclerotinia sclerotiorum: a thorough assessment using discriminatory dose, EC₅₀, high-resolution melting analysis, and description of new point mutation associated with thiophanate-methyl resistance. Plant Dis. 99, 1537–1543 https://doi.org/10.1094/PDIS-11-14-1231-RE (2015).
-
Jamal, R. K. & Jose, V. Determination of phytochemicals by GC–MS in methanol extract of Elephantopus scaber L. J. Pharmacogn. Phytochem. 6, 807–813 (2017).
-
Jeadkong, S., Sawatdee, S. & Atipairin, A. Determination of quinine sulfate in extemporaneous suspension by stability indicating high performance liquid chromatography. Res. J. Pharm. Technol. 10, 1603–1610. https://doi.org/10.5958/0974-360X.2017.00282.7 (2017).
-
Kumar, S., Manoj, P. & Giridhar, P. Fourier transform infrared spectroscopy (FTIR) analysis, chlorophyll content and antioxidant properties of native and defatted foliage of green leafy vegetables. J. Food Sci. Technol. 52, 8131–8139. https://doi.org/10.1007/s13197-015-1959-0 (2015).
-
Daina, A., Michielin, O. & Zoete, V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 7, 42717. https://doi.org/10.1038/srep42717 (2017).
-
Waterhouse, M. et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46 (W1), W296–W303. https://doi.org/10.1093/nar/gky427 (2018).
-
Ma, Y., Liu, Y. & Cheng, J. Protein secondary structure prediction based on data partition and semi-random subspace method. Sci. Rep. 8, 9856. https://doi.org/10.1038/s41598-018-28084-8 (2018).
-
Thumuluri, V., Almagro Armenteros, J. J., Johansen, A. R., Nielsen, H. & Winther, O. DeepLoc 2.0: multi-label subcellular localization prediction using protein language models. Nucleic Acids Res. 50 (W1), W228–W234. https://doi.org/10.1093/nar/gkac278 (2022).
-
Krogh, A., Larsson, B., von Heijne, G. & Sonnhammer, E. L. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305, 567–580. https://doi.org/10.1006/jmbi.2000.4315 (2001).
-
Dallakyan, S. & Olson, A. J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 1263, 243–250. https://doi.org/10.1007/978-1-4939-2269-7_19 (2015).
-
Ejaz, S. A. et al. In-silico investigations of quinine and quinidine as potential inhibitors of AKR1B1 and AKR1B10: functional and structural characterization. PLoS One 17, e0271602. https://doi.org/10.1371/journal.pone.0271602 (2022).
-
Meletiadis, J., Meis, J. F., Mouton, J. W. & Verweij, P. E. Analysis of growth characteristics of filamentous fungi in different nutrient media. J. Clin. Microbiol. 39, 478–484. https://doi.org/10.1128/JCM.39.2.478-484.2001 (2001).
-
Kaur, A. et al. Optimization of protein quality assay in normal, opaque-2, and quality protein maize. Front. Sustain. Food Syst. 6, 743019. https://doi.org/10.3389/fsufs.2022.743019 (2022).
-
Akkus Cevikkalp, S., Biringen Löker, G., Yaman, M. & Amoutzopoulos, B. A simplified HPLC method for determination of tryptophan in some cereals and legumes. Food Chem. 193, 45–49. https://doi.org/10.1016/j.foodchem.2015.02.108 (2015).
-
Zhang, Z.-J., Jiang, Z.-Y., Zhu, Q. & Zhong, G.-H. Discovery of β-carboline oxadiazole derivatives as fungicidal agents against rice sheath blight. J. Agric. Food Chem. 66, 9598–9607.
-
Kim, H. & Xue, X. Detection of total reactive oxygen species in adherent cells by 2′,7′-dichlorodihydrofluorescein diacetate staining. J. Vis. Exp. (2020) e60682. https://doi.org/10.3791/60682
-
Kręgiel, D., Berlowska, J., & Ambroziak, W. Succinate dehydrogenase activity assay in situ with blue tetrazolium salt in Crabtree-positive Saccharomyces cerevisiae strain. Folia Microbiologica, 57(1), 1–4 (2012).
-
Argumedo-Delira, R., Gonzalez-Mendoza, D. & Alarcon, A. A rapid and versatile method for the isolation of total RNA from the filamentous fungus Trichoderma sp. Ann. Microbiol. 58, 761–763. https://doi.org/10.1007/BF03175587 (2008).
-
Schumann, N. A., Smith, M. B. & Wang, M. A fast and efficient method for preparation of high-quality RNA from fungal mycelia. BMC Res. Notes. 6, 71. https://doi.org/10.1186/1756-0500-6-71 (2013).
-
Prabhukarthikeyan, S. R. et al. Comparative proteomic analysis of Rhizoctonia solani isolates identifies the differentially expressed proteins with roles in virulence. J. Fungi 8, 370. https://doi.org/10.3390/jof8040370 (2022).
Acknowledgements
The authors would like to acknowledge the Virus testing laboratory facility under RKVY, STI HUB laboratory facility, Niche Area of Excellence (NAE) facility and Department of Biochemistry of Uttar Banga Krishi Viswavidyalaya, Pundibari, Cooch Behar, West Bengal, India for necessary instrumentation support. The authors would also like to acknowledge Dr. Amitabha Bandyopadhyay for his supervision and critical input during the entire course of study.
Funding
The authors gratefully recognize the financial support received under the project “Study to strengthen the value chains based on Cinchona and other medicinal plants in West Bengal from UBKV” sanctioned vide ref. no. 485/DCOMP/2022 of Directorate of Cinchona and other Medicinal Plants by Department of Food processing Industries and Horticulture, Government of West Bengal, India.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Das, S., Ghosh, S., Pal, K. et al. Cinchona-based liquid formulation exhibits antifungal activity through Tryptophan starvation and disruption of mitochondrial respiration in Rhizoctonia Solani. Sci Rep 15, 39034 (2025). https://doi.org/10.1038/s41598-025-23347-7
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-23347-7