
Introduction
Conditions affecting the liver, or influenced by liver-related pathophysiology such as cardiometabolic diseases, account for millions of deaths per year worldwide.1,2,3 In many liver diseases, treatment options are insufficient and there is an unmet need for new pharmacological therapies.4
Ribonucleic acid (RNA) therapeutics have emerged as a promising therapy for various genetic conditions, including those of the liver.5 A novel site-directed RNA editing method uses chemically modified editing oligonucleotides (EONs) to attract the endogenous enzyme adenosine deaminase acting on RNA (ADAR) to perform specific edits on messenger RNA (mRNA).6,7,8 ADAR is naturally present in all human cells, binds to double-stranded RNA and converts adenosine to inosine (A → I), which is interpreted as a guanosine (G) during translation.9,10,11,12 EONs (strands of nucleotides forming an ADAR-binding backbone and an editing enabling region) can guide ADAR to perform A → I nucleoside changes at specific sites on single-stranded mRNA.7,8,12,13,14 This technique avoids delivery barriers, immune responses, off-target editing associated with exogenous or bacterial-derived enzymes and poses no risk of permanent genome changes.6,13,15
EON-guided ADAR-mediated RNA editing has broad therapeutic potential: firstly, by correcting disease-causing RNA variants;7,13,15 secondly, by altering mRNA expression12; and thirdly, by modulation of amino acid sequences to alter protein function, for example by affecting post-translational modification sites16. ADAR-mediated RNA editing is currently being investigated in conditions affecting the liver and central nervous system.7,8,12,17.
Current development of the EONs used to recruit ADAR is based on decades of research into drug development, natural RNA editing, and biological pathways. As the field is developing rapidly, addressing challenges to develop EONs that guide efficient, precise, ADAR-mediated RNA editing is of importance.13,15 Historically, drug development for liver diseases has been impacted by limitations in the available in vitro and in vivo models, which has resulted in delays in development and drug failure across all stages of clinical development.4,18 While in vitro liver-modeling options, such as traditional cell culture methodologies (e.g., primary human hepatocytes [PHH] monolayer cultures), are evolving rapidly, they can lack the required complexity to accurately mimic disease pathophysiology.4,19 In vitro cultures quickly lose their specificity and hepatocyte-specific functionalities and behave as isolated cells as opposed to whole tissues.19,20,21,22 Furthermore, there is a shortage of liver tissue to form the primary material of in vitro cultures, with high-quality liver favored for transplantation over research use.20 For in vivo models, results are often insufficiently translatable due to the differences in biological and pathophysiological processes between animals and humans, resulting in unforeseen outcomes during clinical trials.4,18,21
To improve predictions of EON-guided ADAR-mediated RNA editing in humans and to increase the chances of successful drug development, in vitro models are required to provide conditions as close as possible to human liver tissue and maintain as much as possible the natural biochemical activities of liver cells. RNA expression varies across different in vitro liver models, and further complexity is introduced when evaluating RNA editing in vivo due to genomic differences between animals and humans.21,22 For in vitro assessments of ADAR RNA editing technology, it is important to confirm that models express sufficient levels of relevant genes and proteins. For example, one of the most important functions of hepatocytes is the synthesis of serum proteins, more specifically of serum albumin.23 Measurement of albumin secretion is used as a liver function test to indicate potential liver injury and liver pathology.23 To evaluate the presence and function of hepatocytes or hepatocyte-like cells both in vitro and in vivo, the constitutive production of albumin is commonly used as a biomarker.24 Because EONs rely on ADAR recruitment, successful models must maintain functional endogenous RNA editing machinery.25 Due to low levels of RNA editing, many cell line studies have relied on overexpression of ADAR to obtain quantifiable results, affecting the interpretation of the results.22,25.
One of the main hurdles for RNA therapeutics is target engagement caused by limited diffusion across lipid bilayers due to their large size and overall negative charge, their so-called non-productive uptake and the subcellular trafficking through the endo-lysosomal pathways hampering exposure to the target site.26 To facilitate the uptake of RNA therapeutics, several strategies have been developed promoting uptake (among others lipofection/chemical conjugates), preventing the degradation of RNA therapeutics, enhancing intracellular transport and facilitating endosomal escape as established by, for example, encapsulation in lipid nanoparticles (LNP).27 When administering RNA therapeutics, the delivery method (e.g., gymnosis, lipofection, LNPs) can have an impact on the level of editing observed.26 Therefore, it is important to understand how accurately in vitro models mirror the editing levels in vivo when using different delivery methods. Development of reliable methods for the screening and selection of lead candidates has been crucial for the testing of short-interfering RNA (siRNA) with LNP delivery, and assessment of serum components, endosomal environment and immunogenicity which are essential to investigate during formulation development.28,29 As such, establishing and validating in vitro models that accurately assess EONs and replicate the inherent editing capacity of ADAR-mediated RNA editing is crucial in the hit and lead selection phase of drug development.
Another major challenge for RNA therapies is their delivery to cells of interest.30 N-acetylgalactosamine (GalNAc) is used for the targeted delivery of siRNAs and antisense oligonucleotides (ASOs) to hepatocytes.31 GalNAc specifically binds to asialoglycoprotein receptors (ASGR1), which are readily expressed on hepatocytes, facilitating the uptake of RNA therapies. Multiple GalNAc-based RNA therapeutics have received global approval and advanced to clinical applications.32 This underscores the efficacy of targeted RNA therapeutics and the importance of developing efficient delivery mechanisms.
Here, we characterized the available in vitro models to assess their suitability for testing EON-guided ADAR-mediated RNA editing. In addition, we assessed the translatability of the EON editing capacity in vivo in the liver of non-human primates (NHPs).
Materials and methods
Part 1: characterization of the in vitro models
We compared gene expression across multiple liver models: hepatocellular carcinoma-derived cell lines HUH7, HepG2 and Hep3B; immortalized human hepatocytes (IHH); PHH from single or multiple donors; HepatoPac PHH cultures; induced pluripotent stem cell (iPSC)-derived hepatocytes; bipotent hepatic cells HepaRG; and tripartite liver microtissues (LMTs). Furthermore, HepG2, HepaRG and PHH cells were cultured in multiple formats, either as a monolayer (2D) culture or as spheroids (3D) culture. In this way, we investigated and characterized models of varying complexity and similarity to human liver. Detailed information on cell culture conditions, including cell model suppliers, seeding densities, and plate formats, is provided in the Supplemental Materials and Methods.
In humans there are three ADAR genes; ADAR1, ADAR2 and the catalytically inactive ADAR3.33 Here we used a digital polymerase chain reaction (dPCR) assay designed to detect ADAR1.
Part 2: translatability of in vitro to in vivo in NHP models
Following the characterization of the different in vitro models, we assessed the translatability of EON editing capacity in the selected in vitro liver models and in vivo in the liver of NHP. We added to the experiment the assessment of editing in primary NHP hepatocytes. We explored the intrinsic editing capabilities of EONs in vivo using LNP as the delivery method. This study was conducted according to the ethical guidelines, approved by the Institutional Animal Care and Use Committee (IACUC) and is reported in accordance with the ARRIVE guidelines. During this study animal welfare was closely monitored. NHPs (n = 4) were treated with intravenous injection of 2 mg/kg EONs directed toward the ACTB mRNA at Day (D) 1, D8 and D15. For this experiment, EONs were encapsulated in LNPs to support delivery into the liver cells. The LNP formulation was Dlin-MC3-DMA, distearoylphosphatidylcholine (DSPC), cholesterol, and DMG-PEG2000 lipids (ratio 50:10:38.5:1.5). To assess translatability between the in vitro models and the results observed in NHPs, PHH and NHP hepatocytes were treated with 250 nM ACTB-EON LNP and editing was assessed after 3 days of treatment.
EON treatment
EON treatment by gymnosis was performed by adding the EONs at 1 µM (non-cytotoxic) concentration into the cell culture media for either 2, 3, 7 or 14 days depending on the model used. EON transfection was carried out using Lipofectamine 2000 at a 1:3 Lipofectamine:EON ratio, with EONs at a final concentration of 100 nM. The media was refreshed 24 h post-transfection, and the cells were harvested after 48 h of treatment. LNP-mediated EON delivery in PHH cells was performed by adding the LNP-EON at non-cytotoxic 100 nM or 250 nM into the cell culture media. LNP formulation was the GenVoy-ILM, containing Precision NanoSystems Ionizable (PNI) lipid 50 mol%, DSPC 10 mol%, cholesterol 37.5 mol%, and PNI stabilizer 2.5 mol%. NHP hepatocytes were treated with 250 nM EON encapsulated in LNP for 3 days. Non-optimized EONs were utilized for ACTB, primarily serving as a model to explore editing within in vitro and in vivo experimental setups.
Transcript quantification and endogenous RNA editing assays
In vitro cultured models
For dPCR and Sanger sequencing analysis, cells were lysed with TRI Reagent (Zymo Research, R2050-1–200) and RNA was isolated with Direct-zol RNA Microprep kit (Zymo Research, R2062) according to manufacturer’s protocol. Complementary DNA (cDNA) synthesis was performed with Maxima reverse transcriptase (Thermo Fisher, EP0742), Oligo(dT)18 primers (Thermo Fisher, SO132), random hexamer primers (Thermo Fisher, SO142), 10 mM dNTP mix (Thermo Fisher, R0192), and ambion nuclease free water (Life Technologies, AM9937). For Sanger sequencing analysis, cDNA was used to amplify the transcript of interest using DNA Polymerase (SsoAdvanced™ Universal SYBR® Green Supermix, Bio-Rad, 172–5270) and primers specific for ACTB (Fw: 5’-CGAGCATCCCCCAAAGTTCAC-3’; Rv: 5’-CACTCCCAGGGAGACCAAAAGC-3’, product = 347 bp). PCR products were analyzed by Sanger sequencing (BaseClear B.V.). A-to-I edits were quantified using EditR software (https://moriaritylab.shinyapps.io/editr_v10/). Briefly, the area under the curve for each position across each base (A, T, G or C) was directly extracted from the sequencing file and determined by the makeBaseCalls function of sangerseqR. The percentage of editing was calculated using the following equation: 100 × (area A/sum area A + T + G + C). Human liver total RNA samples were purchased from Takara (636531), Thermo Fisher Scientific (AM7960), and Zenbio (RBAmi-LT10). RNA used for transcript quantification in Fig. 1 was isolated from cells at the start of EON treatment.

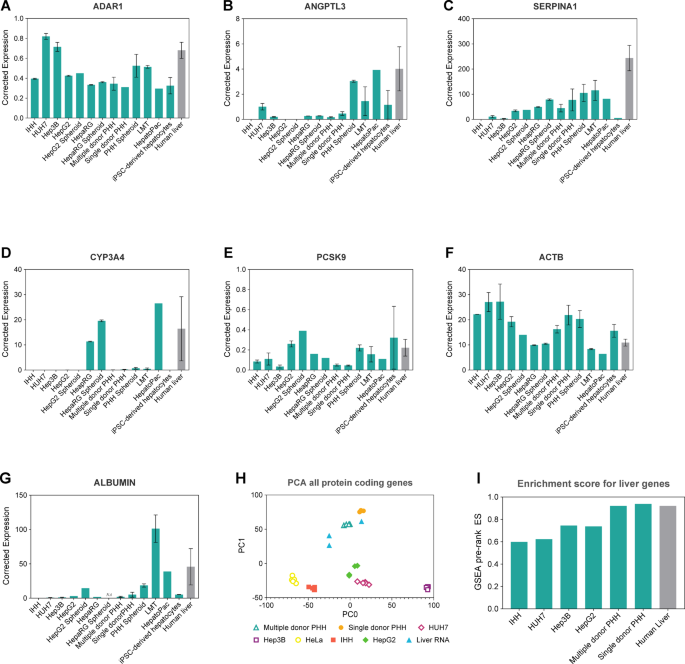
In vitro liver models expressing ADAR1 and liver-specific genes measured 24 h post-seeding (IHH, HUH7, Hep3B, HepG2, HepaRG, PHH models), 7 days post-seeding (HepG2 Spheroid, HepaRG Spheroid, PHH Spheroid, iPSC-derived hepatocytes) and 1 day after receiving the cells (LMT, HepatoPac). Expression of A ADAR1, B ANGPTL3, C SERPINA1, D CYP3A4, E PCSK9, F ACTB and G ALBUMIN were determined using dPCR assays. Expression was corrected to the geometric mean of four housekeeping genes (see Materials and Methods for details). Data shown is mean ± standard error of the mean (SEM), n = 1–3 biological replicates. H Principal component analysis of protein coding gene expression in in vitro liver models. I Enrichment score for in vitro model liver gene expression. NA: not available
For RNA sequencing, total RNA was extracted using RLT buffer (QIAGEN) and isolated using the RNeasy Mini Kit (QIAGEN, 74104) according to the manufacturer’s instructions. Samples were DNAse treated and purified prior to library preparation. Ribosomal RNA-depleted whole transcriptome RNA sequencing was performed by Genomescan B.V. using Illumina NovaSeq sequencing, Paired-End, 150 bp. Each sample generated approximately 60 Gb of Paired-End reads. Performance of sample preparations including unique dual indexing and unique molecular identifiers were done according to GenomeScan B.V. Standard Operating Procedures. Raw 150PE reads were trimmed for adaptors using BBDuk and mapped to the Human Genome (GRCh38) using HISAT2 (HISAT version 2.2.1, no mix, no discordant parameters). PCR duplicates were removed using samtools dedup and gene read counts were extracted using HTSeq. Differential gene expression was calculated using DESeq2.
For enrichment score for in vitro model liver gene expression, a Wald test stat (default parameter) was used to rank the genes as Gene Set Enrichment Analysis (GSEA) pre-rank algorithm. It was defined as log2 fold change/standard error (from differential gene expression analysis). The source of the liver-specific gene set employed was SU_LIVER.v2023.2.Hs.gmt (https://www.gsea-msigdb.org/gsea/msigdb/cards/SU_LIVER).
NHP tissue
The methods used to determine ACTB editing in NHP tissue are described in the Supplement (Table S2).
Albumin secretion
The secretion of albumin by the different models was determined by quantifying the albumin concentration in the cell culture medium using the Human Albumin ELISA Kit (Abcam, ab108788) according to the manufacturer’s instructions. Parameters used for calculating albumin secretion are described in the Supplement (Table S3).
In vivo NHP study
Four male naïve cynomolgus monkeys (2–4 years of age) were provided by Guangdong Chunsheng Biotechnology Development Co., LTD, Hainan New Source Biotech Co., Ltd and Fangchenggang Kanglu Biotechnology Co., Ltd. They were acclimatized for 10 days before being dosed intravenously with 2 mg/kg at D1, D8 and D15. This study was exploratory, aiming to evaluate efficacy of oligonucleotides for ACTB RNA editing in vivo; no control group was used, or randomization performed, and the study was performed under unblinded conditions. Each cynomolgus monkey was assessed in an individual experimental unit. The LNP formulation was administered as an intravenous infusion over a time period of 60 min via the cephalic vein via a percutaneously placed catheter. A new sterile indwelling catheter was placed prior to each dose. Before the LNP formulation infusion, animals were anesthetized by an intramuscular dose of 0.02 mg/kg of atropine, followed by an intramuscular dose of 5–6 mg/kg of Zoletil. Blood samples were collected at several timepoints post-dose to monitor clinical chemistry parameters and needle biopsies of about 10 mg were taken from the liver at D4 and D18. For ethical reasons and to allow sufficient recovery of the animals, the minimal time between biopsies was 14 days. The monkey liver samples were homogenized by the QIAGEN Tissue Lyser II with TRIzol. The RNA was extracted using chloroform and precipitated with an equal volume of isopropyl alcohol. After incubation at − 20 °C, the RNA pellets were washed twice with 70% ethanol and dissolved in RNase-free-water. The ACTB editing was determined by droplet digital PCR (ddPCR).
EON concentration determination by hybridization HPLC
Levels of the ACTB-EON in liver and cell suspensions were measured by peptide nucleic acid (PNA) hybridization assay (Axolabs, Germany). For this method, a complementary Atto-425 dye-labeled PNA probe was hybridized to the EON and quantified using AEX-HPLC analysis with fluorescence detection. Liver samples were lysed in Tissue & Cell lysis solution (containing 0.25 mg Proteinase K) using a Vibracell Ultrasonic processor to obtain a final concentration of 66.67 mg/mL of tissue lysate. The lysed samples were incubated for 60 min at 65 °C and 20 µL of each sample was transferred to a 96-well plate with 70 µL dilution buffer (10% MeCN + 0.01% Tween20 in PBS). The samples were precipitated using KCl (10 µL per sample) for 30 min on ice, and centrifuged for 15 min at 2000 g. The supernatant was transferred to a 96-well PCR plate and 140 µL of hybridization mixture with ~ 14 nM of the Atto-425 dye-labeled PNA was added. Annealing was carried out by incubation at 95 °C for 15 min using an Eppendorf Thermomixer C, after which the samples were snap-cooled on ice for an additional 15 min. The plate was loaded in a ThermoFisher UltiMate 3000 Quad-pump HPLC system, run through a DNAPac PA200 anion exchange column, and signal detected by an UltiMate 3000 RS Fluorescence Detector. The mobile phases consisted of buffer A (25 mM Tris–HCl, 1 mM EDTA, 3 M Urea, 30% MeCN) and buffer B (800 mM NaClO4 in buffer A). A gradient of buffer B (20–80% in 5 min) was used for the elution of the EON-PNA hybrids. The column temperature was kept at 60 °C. The Atto-425 signal (436 nm ex/484 nm em) was recorded, and detected peaks were integrated to obtain the area under the curve (AUC). The final concentrations from the experimental samples were ascertained by correlating the obtained AUCs with the corresponding calibration curve, which was generated by spiking known amounts of the respective EON reference formulation into tissue lysates derived from an untreated animal.
Histology
All cells were fixed with formalin. HepaRG spheroids, PHH spheroids, and LMTs were processed with tissue processor Excelsior AS (Epredia™), embedded in paraffin and sectioned (3 µm thickness). Antigen retrieval was performed with Citrate buffer pH 6.0. ASGR1 was stained after permeabilization with Triton X-100 (Merck, X100-100 ml) with an ASGR1 Rabbit Polyclonal antibody (Proteintech, 11,739–1-AP). The secondary antibody used was AF647 goat anti rabbit (Invitrogen, A32733). The nucleus was stained with Hoechst 33,342 (Invitrogen, H3570). Images were obtained with a Zeiss LSM800 confocal microscope.
Statistical analysis
For hybridization HPLC data collection was performed with the softwares Spectralis 6.8, Heidelberg Engineering, Germany and Diagnosys, Espion V6, Lowell, MA, USA. and data analysis was performed with the software SAS v9.4. No custom code or algorithms were used. All other data was analyzed in Microsoft Excel and GraphPad Prism unless stated otherwise. Statistical tests are noted in the figure legends when used.
Results
Part 1: characterization of in vitro liver models
For the characterization of the liver models used in this study, several factors were considered. Firstly, to ensure proper assessment of EON-guided ADAR-mediated RNA editing, it is important to confirm that models express the relevant genes and proteins at sufficient levels, as multiple parameters drive the understanding of the potential of the technology. Expression of ADAR enzymes should be present and consistent in all models assessing EON-guided ADAR-mediated RNA editing as it is the main driver of the editing process. Secondly, for good characterization, liver-specific genes should be expressed to levels consistent with the human liver. Therefore, we used commercial samples of human total liver RNA as control in this study. Similarly, the target of interest (RNA and protein) should also be expressed to a sufficient and consistent level to what would be expected in human liver to allow proper assessment of the targeted editing capacity. For this study, we have used the ubiquitously expressed gene ACTB as the main target for model assessment. Lastly, the amino sugar GalNAc is widely used in oligonucleotide therapeutics to facilitate uptake into hepatocytes via the ASGR1. In our study, we have assessed the models for presence of ASGR1 as a transporter for EONs via potential conjugation with GalNAc (5’-triantennary) delivery technology.
Characterization of the ASGR1 receptor
The GalNAc ligand has been widely used for liver-specific delivery of oligonucleotides as the GalNAc receptor ASGR1 is abundantly expressed in hepatocytes.31,32,35 We characterized the presence of the ASGR1 receptor at the RNA and protein levels in the same in vitro liver models using dPCR and immunofluorescence studies, respectively. In general, there was a trend between expression at the RNA level and protein level. For example, ASGR1 protein was undetectable in IHH cells, in line with the lack of ASGR1 RNA expression in this model (Fig. 2). Likewise, HUH7 cells and Hep3B cells showed little expression at the RNA and protein levels, while PHH spheroids showed high expression in both assays (Fig. 2). Importantly, the use of immunofluorescence studies to show localization of ASGR1 showed that high expression at the RNA level was not per se related to high protein expression of ASGR1. For example, in HepG2 cells ASGR1 RNA expression was high but the protein was mainly localized in the cytoplasm, suggesting that the receptor may not be trafficked to the membrane or fully functional in these cells.

In vitro liver models express the GalNAc receptor ASGR1 measured 24 h post-seeding (IHH, HUH7, Hep3B, HepG2, HepaRG, PHH models), 7 days post-seeding (HepG2 Spheroid, HepaRG Spheroid, PHH Spheroid, iPSC-derived hepatocytes) and 1 day after receiving the cells (LMT, HepatoPac). A Expression of ASGR1 was determined using dPCR assay. Expression was corrected to the geometric mean of four housekeeping genes. Data shown is mean ± SEM, n = 3 biological replicates (except n = 1 with 3 technical replicates for HepG2 spheroids and HepatoPac). B Localization of ASGR1 (red) expression in selected in vitro liver models. Scale bar = 10 µm
Part 2: oligonucleotide-mediated editing in in vitro liver models
Expression of ADAR1 and select liver-related genes across models
A total of thirteen liver cell models, cultured in 2D and 3D format, were tested and compared to human liver in this study. ADAR1 expression was relatively consistent across models, with PHH spheroids, LMTs, HUH7, and Hep3B showing the closest ADAR1 expression to human liver (Fig. 1A). ANGPTL3, a liver-produced angiopoietin-like protein involved in lipoprotein lipase inhibition34, exhibited varying expression levels across different liver models. Generally, it was expressed at low levels in cell lines and 2D models (except HUH7 and HepatoPac) but showed higher levels in 3D models such as PHH spheroids and LMTs (Fig. 1B). SERPINA1, encoding alpha-1-antitrypsin, a serine protease expressed in the liver, displayed increasing expression relative to model complexity. Cell lines exhibited lower levels compared to PHH models, which in turn were slightly lower than 3D models. Generally, all cell models tend to express lower levels of SERPINA1 in comparison to human liver (Fig. 1C). CYP3A4, which is involved in drug metabolism, exhibited relatively high expression in HepaRG (in both 2D and 3D formats) and HepatoPac (Fig. 1D). PCSK9, a proprotein convertase involved in low density lipoprotein receptor degradation, showed variable expression levels across models (Fig. 1E). The levels of the study target ACTB, encoding the structural protein β-actin, were also assessed, and showed comparable expression levels across models (Fig. 1F). ALBUMIN (ALB) is an important marker given its role in influencing drug binding and pharmacokinetics. ALB transcripts were detectable in most systems, showing a tendency towards higher levels of expression in LMTs, followed by HepatoPac, and PHH spheroids. In contrast, hepatoma cell lines (HepG2, Hep3B, and Huh7) showed low ALB transcript abundance, while transcripts were undetectable in IHH cells. Similar expression levels were observed for PHH (single donor) and iPSC-derived hepatocytes (Fig. 1G).
Comparison of human in vitro models vs human liver tissue
We used whole transcriptome sequencing to further analyze the following human liver cell models in comparison to human liver tissue: cancer-derived cell lines (HepG2, HUH7, Hep3B, IHH, HeLa), and PHH (multiple and single donor). We observed that in a principal component analysis focused on protein coding genes, the human liver samples clustered closer to the PHH cell lines than to the remaining cell line models, suggesting a closer gene expression profile between the liver samples and the PHH models (Fig. 1H). In order to further gauge the degree of liver identity of each model compared to the human liver tissue, we analyzed how enriched each model was for liver-specific genes using HeLa cells as a reference. We calculated the differential gene expression between each model (including human liver) and HeLa cells and, using the Gene Set Enrichment Analysis pre-rank algorithm, we determined the enrichment of each model for liver-specific genes. As expected, liver cells showed an enrichment score above 0.9. Both PHH models showed similar enrichment levels as human liver, reinforming the notion that their transcriptional profile is similar to human liver. The remaining models (HepG2, HUH7, Hep3B, IHH) showed less enrichment for the liver-specific gene set (enrichment score between 0.60 and 0.74) (Fig. 1I).
Comparison of albumin secretion
To further characterize in vitro liver models, we compared albumin secretion as a proxy of liver functionality in a subset of the models, especially paying attention to differences between 2D vs 3D culture models. To that end, we measured albumin secretion in cell lines IHH, HUH7, Hep3B and HepG2, in PHH (as 2D and 3D cultures), and LMTs (Fig. S2). We observed differences in the levels of albumin secretion among the different cell lines, with HepG2 having significantly the highest rate of albumin secretion of the 2D models. Interestingly, albumin secretion was quite low in the PHH 2D culture model; PHH cultured in 3D showed on average 17 times more secretion than when cultured in 2D. These results suggest that a 3D structure may be needed for proper albumin secretion; however, additional factors such as specific donor characteristics cannot be ruled out.
Editing capability of 2D in vitro liver models
We evaluated the editing efficiency in 2D in vitro liver models using an EON targeting ACTB, comparing the efficacy of naked EON versus GalNAc-conjugated EON to assess potential enhancements in hepatocyte uptake. Treatment was administered through gymnosis to test the cellular entry of the EONs without a delivery agent, at 1 µM concentration. Among cell lines IHH, HepG2, Hep3B, and HUH7 treated with 1 µM EON for 48 h, IHH exhibited a trend towards higher editing percentage (~ 12%), while HepG2 showed the lowest (1%), with HUH7 at 3% and Hep3B at 5.5%. GalNAc conjugation led to a two-fold increase in editing in HepG2 (~ 3%), with minimal effects in HUH7 and Hep3B and a decrease in IHH to 5% (Fig. 3A). HepaRG cultures allowed for extended treatment durations of up to 7 and 14 days, revealing similar editing levels between naked (unconjugated) and GalNAc-conjugated EONs at D7 (8.5% and 6.5%, respectively) but a non-significant increase at day 14, reaching 45% with naked EON and 17% with GalNAc-conjugated EON, indicating a potential time-dependent effect on editing efficiency with different delivery strategies, warranting further investigation into the long-term stability and efficacy of GalNAc-conjugated EONs in HepaRG cultures. (Fig. 3B). In PHH (Fig. 3C), we tested our EONs (naked and GalNAc-conjugated, 1 µM, 3 days treatment) coming from a single donor or a 10-donor pool. Both single donor and pooled samples exhibited comparable editing levels with naked EON (~ 30%). A modest improvement was observed with GalNAc-EON in multiple donor cells (~ 37%) and notably higher editing of 56% in single donor cells (Fig. 3C). HepatoPac, a liver model composed of hepatocytes and stromal cells, showed ACTB editing rates of 44% and 58% with naked and GalNAc-conjugated EONs, respectively, after 7 days of treatment at 1 µM (Fig. 3D). Additionally, iPSC-derived hepatocytes treated for 7 days with 1 µM naked or GalNAc-conjugated ACTB-EON reached 35% editing with the naked EON and 46% editing with the GalNAc-conjugated EON (Fig. 3E). An increased trend in ACTB editing was observed after 14 days of treatment, reaching up to 59% with the naked EON and 74% with the GalNAc-conjugated EON (Fig. 3E). These findings underscore the diverse editing capabilities of different liver models, demonstrating that the influence of GalNAc conjugation to EON on editing capability strongly depends on the model system used. We observed a positive correlation between ASGR1 receptor expression and editing efficiency in PHH (single and multiple donor), across both 2D and 3D formats (Pearson r = 0.859; P = 0.0283), as well as in HepatoPac, LMTs, and iPSC-derived hepatocytes. However, in cancer-derived cell lines (IHH, HepG2, Hep3B, HUH7, and HepaRG), where ACTB editing levels were generally low, there was no significant correlation between ASGR1 expression and editing efficiency. This suggests that ASGR1-mediated uptake may play a critical role in editing efficiency in primary and non-cancerous hepatocyte models, but other factors likely influence editing in cancer-derived lines.

Editing of ACTB in 2D in vitro liver models. ACTB editing percentage after treatment with either 1 µM unconjugated or GalNAc-conjugated ACTB-EON in A cell lines HUH7, Hep3B, HepG2 and IHH for 48 h, B HepaRG for 7 and 14 days, C single or multiple donor PHH for 3 days, D HepatoPac for 7 days, and E iPSC-hepatocytes for 7 and 14 days, compared to untreated cells. Data shown is mean ± SEM, n = 1–3 biological replicates. F Dose–response of unconjugated and GalNAc-conjugated EON in multiple donor PHH after 3 days.
Editing capability of 3D in vitro liver models
In PHH spheroids, treatment for 7 days with 1 µM ACTB-EON or ACTB-GalNAc EON showed 36% and 69% editing, respectively (Fig. 4A). In LMTs, an average of 32% editing was observed for the naked EON and 44% editing for the GalNAc-conjugated EON (Fig. 4B). The difference in editing level reported with the GalNAc-conjugated EON between the two models is in line with ASGR1 expression reported in Fig. 2A, suggesting that higher expression of ASGR1 receptor in PHH spheroids leads to higher EON uptake and ultimately increased editing level. Because of the longer life span of 3D models compared to 2D models, we were able to test RNA editing over time using longitudinal studies. In general, time course treatment up to 14 days with the naked ACTB-EON resulted in a treatment accumulative effect, where higher editing efficiency was observed at the end of treatment in all the 3D models assessed. Continuous treatment of HepG2 spheroids resulted in 7%, 12%, 15% and 25% editing at day 3, 5, 7 and 14, respectively (Fig. 4C). Continuous treatment of PHH spheroids resulted in 6%, 9%, 10% and 19.5% editing at day 3, 5, 7 and 14, respectively (Fig. 4D). Of note, PHH used for the time course experiment were of a different donor than the cells used in Fig. 4A, showing the influence on editing results depends on the cell origin. Continuous treatment of LMTs resulted in 12%, 24%, 30% and 37% editing at day 3, 5, 7 and 14, respectively (Fig. 4E).

Editing of ACTB in 3D in vitro liver models. A–B ACTB editing percentage after treatment with either 1 µM unconjugated or GalNAc-conjugated ACTB-EON in A PHH spheroids and B LMTs. C–E ACTB editing percentage after treatment with unconjugated ACTB-EON in C HepG2 spheroids, D PHH spheroids, and E LMTs, up to 14 days. Data shown is mean ± SEM, n = 1–3 biological replicates
EON delivery methods and editing measurement methods
We observed that in some of the models tested, ACTB was not successfully edited following gymnotic uptake of the EONs. To address this, we evaluated the editing potential of different delivery methods for ACTB-EONs at 100 nM, comparing gymnosis, transfection (Lipofectamine 2000), and LNP-mediated delivery (Genvoy-ILM). Editing efficiency in PHH varied significantly depending on the delivery method. Gymnotic uptake resulted in 18% editing, transfection yielded 30%, and LNP-mediated delivery achieved the highest efficiency at 44% (Fig. 5A).

Editing of ACTB in in vitro liver models depends on delivery method. A ACTB editing percentage in multiple donor PHH after treatment with 100 nM unconjugated ACTB-EON via gymnosis, transfection, or LNP. Data shown is mean ± SEM, n = 3 biological replicates. B ACTB editing percentage in HepG2 cells after treatment with 100 nM unconjugated ACTB-EON via gymnosis or transfection. The data shown is mean ± SEM, n = 1 biological replicate. C Comparison of editing percentage calculation method. ACTB editing percentage calculated using either Sanger sequencing or dPCR method (see Materials and Methods for details)
Interestingly, the differences in editing efficiency were more pronounced in HepG2 cells. Gymnotic uptake led to only 1% editing, while transfection resulted in 23% editing (Fig. 5B). No LNP data was available for this model. These results highlighted the importance of model and delivery method selection.
To ensure reliable and accurate measurement of A-to-G editing, we compared editing percentages calculated using Sanger sequencing and dPCR in PHH cells treated with ACTB-EONs at 0.01 µM and 1 µM. The results showed minor differences between the two methods (Fig. 5C), with Sanger sequencing consistently reporting slightly higher editing percentages. This suggests that while both methods are suitable for assessing editing efficiency, dPCR may offer slightly greater stringency and accuracy.
Part 3: translatability of in vitro to in vivo in NHP models
Naïve male cynomolgus monkeys (Macaca fascicularis) were intravenously infused at D1, D8, and D15 with 2 mg/kg unconjugated ACTB-EON using LNP to achieve efficient hepatocyte delivery in vivo. The animals well-tolerated the LNP formulation with ACTB-EON and no abnormalities were observed throughout the study. The ddPCR results of collected liver biopsies showed an editing efficiency of up to 70% and that the mean ACTB RNA editing efficiency on D4 and D18 was 62.97% and 62.26%, respectively (Fig. 6A). About 93% ACTB editing was observed in the PHH in vitro model and 76.6% in primary NHP hepatocytes, which correlated well with the high editing values observed in vivo (Fig. 6B).

In vitro—in vivo comparison of ACTB editing upon exposure of an LNP-formulated unconjugated ACTB-EON to PHH cells and to NHP cells (in vitro) and upon intravenous dosing of the unconjugated ACTB-EON to male cynomolgus monkeys at 2 mg/kg on D1, D8, and D15. A ACTB editing percentage in NHP liver biopsies 4 or 18 days following intravenous dosing of the unconjugated ACTB-EON formulated in LNP to male cynomolgus monkeys at 2 mg/kg on D1, D8, and D15. Data are presented as mean ± SEM, n = 4. B ACTB editing percentage in multiple donor PHH or NHP primary hepatocytes after treatment with 250 nM unconjugated ACTB-EON formulated in LNP for 72 h. Data are presented as mean ± SEM, n = 3 biological replicates
The liver concentration of unconjugated ACTB-EON in livers of the NHP was 1,010 ± 122 nM at D4 and 1,568 ± 140 nM at D11 which was about four–sixfold higher than the concentration of ACTB-EON in medium used for treatment of PHH and primary NHP hepatocytes with LNP EON (250 nM) (Fig. 6A).
Similar to the approach used for the in vitro models, where different quantification methods were evaluated, editing efficiency in liver-treated NHP samples was assessed using dPCR, Sanger sequencing, and next-generation sequencing (NGS). As shown in Supplementary Fig. S1, all three methods produced highly consistent results, reinforcing the reliability and robustness of our data.
Discussion
This study aimed to assess the potential of in vitro liver models to facilitate ADAR RNA editing using ACTB as a target and to evaluate the efficiency of GalNAc-conjugated oligonucleotides across various 2D and 3D culture systems.
In vitro liver models serve as crucial tools for optimizing EONs during the hit and lead optimization phases of drug development. They enable the ranking of EON candidates based on editing efficiency, testing of targeting strategies like GalNAc conjugation, and evaluation of lipid nanoparticle formulations for enhancing delivery.4,36 These models also allow for the assessment of hepatotoxicity, metabolic stability, and on- and off-target effects, providing essential data for early-stage drug development.37 Hepatotoxicity of EONs was assessed in vitro using lactate dehydrogenase (LDH) and caspase activity. Across multiple targets, including both conjugated and unconjugated EONs, as well as LNP-formulated EONs, no cytotoxicity was observed. In this study, of the in vitro liver models tested, ADAR expression was consistent across all tested models, indicating that each model has the capacity to effectively assess ADAR RNA editing activities and confirming that ADAR expression does not appear to be a limiting factor in these assessments. Prioritizing the use of PHH spheroids, LMTs, HUH7, and Hep3B models to assess ADAR editing capacity of EONs during preliminary evaluations appears to be a reasonable approach, as these models have demonstrated ADAR expression levels most similar to those observed in human liver tissue.
The editing efficiency of 2D-cultured liver models was evaluated using ACTB-targeting EON, with a comparison between naked and GalNAc-conjugated EONs. The results revealed distinct differences in editing efficiencies across cell lines, indicating model-specific responses to the delivery strategies. The varied effects of GalNAc conjugation emphasize the necessity of proper GalNAc receptor localization for optimal oligonucleotide delivery in these models. This highlights the importance of model-specific mechanisms in determining the potential of these models for RNA editing.
The cell lines responded differently to EON treatment. IHH cells displayed the highest editing efficiency with naked EON, despite their relatively low expression of liver markers. This suggests that a high editing percentage does not necessarily determine the model’s suitability for RNA editing studies. Conversely, HepG2 exhibited the lowest editing efficiency, with responses to GalNAc conjugation varying among cell lines. Interestingly, HepaRG treatment with the GalNAc-conjugated EON did not lead to an increase in editing efficiency compared to the naked oligonucleotide. This observation may reflect incomplete functional activity of the ASGR1 receptor in this in vitro system. While ASGR1 mRNA levels remained stable between day 1 and day 7 of culture, immunolocalization studies revealed predominantly intracellular receptor distribution, suggesting limited recycling of ASGR1 back to the plasma membrane. This intracellular retention may hinder receptor-mediated uptake of GalNAc-conjugated compounds. Alternatively, receptor saturation or competitive binding may also account for the absence of a differential effect. These findings underscore the critical role of receptor expression, uptake mechanisms, and intracellular processing in determining the editing capabilities of these models.
PHH and HepatoPac models showed strong editing efficiencies with both naked and GalNAc-conjugated EONs, demonstrating their potential for in vitro editing studies. Single-donor PHH displayed comparable editing levels to those observed in the 10-donor pool, suggesting that PHH is a reliable model despite inherent donor variability. HepatoPac, composed of hepatocytes and stromal cells, exhibited substantial editing efficiencies with both EON formulations. PHH spheroids demonstrated the highest liver marker expression, albumin secretion, and editing capabilities, reinforcing the superiority of 3D over 2D formats in mimicking liver physiology.
The iPSC-derived hepatocytes exhibited moderate editing efficiencies with both naked and GalNAc-conjugated EONs, making them a viable option for RNA editing studies, particularly in applications that require scalability and accessibility. Although their editing capabilities were not as high as those observed in PHH models, iPSC-derived hepatocytes offer valuable potential for larger-scale or high-throughput applications.
In vitro hepatocyte models are essential for liver-targeting therapies during the drug development process. Initially developed for small molecule studies—focusing on absorption, distribution, metabolism, and excretion, PK, drug transport, and pharmacodynamics—their application to RNA editing technologies is increasingly relevant. This study provides insight into key factors affecting hepatocyte-targeting mRNA editing. Further research is necessary to fully establish the value of these models in the context of RNA editing technologies.
Investigating the EON editing capacity on a specific target, model selection will be largely guided by the expression levels of the target gene for ADAR RNA editing, given the observed variability in target expression across different models.
While PHH models are considered the gold standard, significant variability exists between donors and between 2D and 3D cultures.19,38,39 This variability may mirror the diverse liver conditions found in patients, offering both a challenge and an opportunity for clinically relevant insights.19,39 Testing EONs across multiple donor samples can provide valuable data on the potential efficacy of treatments in diverse patient populations. However, PHH models in 3D formats, while more physiologically relevant, face limitations such as high cost, limited availability, and donor variability.40 In contrast, immortalized liver cell lines like HepG2, HUH7, and Hep3B offer scalability and reproducibility but may not fully replicate the complexity of primary hepatocytes.39,40
In vitro models provide critical insights into RNA editing, while in vivo systems introduce additional complexities such as systemic distribution, extrahepatic degradation, and kinetic behavior, all of which influence editing efficacy.41 Immunogenicity is another factor to consider; although antisense oligonucleotides have shown low immunogenic potential, the slow onset of antibody development could affect long-term efficacy.42 However, in general, the immunogenic response to phosphorothioate-modified oligonucleotides appears minimal, with anti-drug antibodies having little impact on circulating drug concentrations and pharmacokinetics.43,44
These findings emphasize the importance of considering model-specific responses and delivery strategies when assessing RNA editing capabilities in liver models. PHH models, particularly in 3D culture formats, closely mimic human liver physiology, but iPSC-derived hepatocytes offer scalability and accessibility. Further research is warranted to understand the mechanisms driving the observed variations and to optimize editing protocols across different liver models. Ultimately, the choice of an in vitro model should align with the specific objectives of the study, balancing physiological relevance, scalability, and feasibility for the most effective outcomes. A multimodel assessment utilizing composite scoring to comprehensively assess the key parameters could represent an approach of choice to maximize translatability across preclinical models and increase the likelihood of success in clinical applications.
Conclusions
Conditions affecting the liver or those influenced by pathophysiological processes in the liver still represent a significant and growing global health burden, with limited treatment options available for many of them. The emergence of RNA therapeutics, particularly ADAR-mediated RNA editing, has introduced new possibilities to address liver disorders. Leveraging EONs to guide endogenous ADAR enzymes to perform site-specific mRNA edits holds promise to address unmet therapeutic needs. Overall, these findings underscore the importance of considering model-specific responses and delivery strategies when assessing the editing capabilities in in vitro liver models. Further investigations are warranted to elucidate the underlying mechanisms driving the observed variations and to optimize editing protocols for different liver model systems.
Data availability
The data from this study are available in the Gene Expression Omnibus repository (accession number: GSE304242 [https://www.ncbi.nlm.nih.gov/geo/]).
References
-
Wang, T. Y. et al. Association of metabolic dysfunction-associated fatty liver disease with kidney disease. Nat. Rev. Nephrol. 18, 259–268. https://doi.org/10.1038/s41581-021-00519-y (2022).
-
Zhou, X. D. et al. An international multidisciplinary consensus statement on MAFLD and the risk of CVD. Hepatol. Int. 17, 773–791. https://doi.org/10.1007/s12072-023-10543-8 (2023).
-
Devarbhavi, H. et al. Global burden of liver disease: 2023 update. J. Hepatol. 79, 516–537. https://doi.org/10.1016/j.jhep.2023.03.017 (2023).
-
Kaur, S. et al. In vitro models for the study of liver biology and diseases: Advances and limitations. Cell Mol. Gastroenterol. Hepatol. 15, 559–571. https://doi.org/10.1016/j.jcmgh.2022.11.008 (2023).
-
Baruteau, J., Brunetti-Pierri, N. & Gissen, P. Liver-directed gene therapy for inherited metabolic diseases. J. Inherit. Metab. Dis. 47, 9–21. https://doi.org/10.1002/jimd.12709 (2024).
-
Montiel-Gonzalez, M. F., Diaz Quiroz, J. F. & Rosenthal, J. J. C. Current strategies for site-directed RNA editing using ADARs. Methods 156, 16–24. https://doi.org/10.1016/j.ymeth.2018.11.016 (2019).
-
ASGCT 27th Annual Meeting Abstracts. Molecular Therapy 32, 1–889 (2024). https://doi.org/10.1016/j.ymthe.2024.04.020
-
Cnubben NHP, e. a. ASGCT 27th Annual meeting abstracts. Molecular Therapy 32, 1–889 (2024).
-
Tan, M. H. et al. Dynamic landscape and regulation of RNA editing in mammals. Nature 550, 249–254. https://doi.org/10.1038/nature24041 (2017).
-
Nishikura, K. Functions and regulation of RNA editing by ADAR deaminases. Annu. Rev. Biochem. 79, 321–349. https://doi.org/10.1146/annurev-biochem-060208-105251 (2010).
-
Nishikura, K. A-to-I editing of coding and non-coding RNAs by ADARs. Nat. Rev. Mol. Cell Biol. 17, 83–96. https://doi.org/10.1038/nrm.2015.4 (2016).
-
Booth, B. J. et al. RNA editing: Expanding the potential of RNA therapeutics. Mol Ther 31, 1533–1549. https://doi.org/10.1016/j.ymthe.2023.01.005 (2023).
-
Doherty, E. E. et al. ADAR activation by inducing a syn conformation at guanosine adjacent to an editing site. Nucleic Acids Res. 50, 10857–10868. https://doi.org/10.1093/nar/gkac897 (2022).
-
Monian, P. et al. Endogenous ADAR-mediated RNA editing in non-human primates using stereopure chemically modified oligonucleotides. Nat Biotechnol 40, 1093–1102. https://doi.org/10.1038/s41587-022-01225-1 (2022).
-
Doherty, E. E. et al. Rational design of RNA editing guide strands: cytidine analogs at the orphan position. J. Am. Chem. Soc. 143(18), 6865–6876 (2021).
-
Slotkin, W. & Nishikura, K. Adenosine-to-inosine RNA editing and human disease. Genome Med. 5, 105. https://doi.org/10.1186/gm508 (2013).
-
Zabaleta, N., Torella, L., Weber, N. D. & Gonzalez-Aseguinolaza, G. mRNA and gene editing: Late breaking therapies in liver diseases. Hepatology 76, 869–887. https://doi.org/10.1002/hep.32441 (2022).
-
Serras, A. S. et al. A critical perspective on 3D liver models for drug metabolism and toxicology studies. Front Cell Dev. Biol. 9, 626805. https://doi.org/10.3389/fcell.2021.626805 (2021).
-
Bell, C. C. et al. Characterization of primary human hepatocyte spheroids as a model system for drug-induced liver injury, liver function and disease. Sci. Rep. 6, 25187. https://doi.org/10.1038/srep25187 (2016).
-
Harrison, S. P. et al. Liver organoids: Recent developments limitations and potential. Front Med (Lausanne) 8, 574047. https://doi.org/10.3389/fmed.2021.574047 (2021).
-
Gupta, R. et al. Comparing in vitro human liver models to in vivo human liver using RNA-Seq. Arch Toxicol 95, 573–589. https://doi.org/10.1007/s00204-020-02937-6 (2021).
-
Schaffer, A. A. et al. The cell line A-to-I RNA editing catalogue. Nucleic Acids Res. 48, 5849–5858. https://doi.org/10.1093/nar/gkaa305 (2020).
-
Buyl, K., De Kock, J., Bolleyn, J., Rogiers, V. & Vanhaecke, T. Measurement of albumin secretion as functionality test in primary hepatocyte cultures. Methods Mol. Biol. 1250, 303–308. https://doi.org/10.1007/978-1-4939-2074-7_22 (2015).
-
Roll, G. R. & Willenbring, H. Transplanted nonviable human hepatocytes produce appreciable serum albumin levels in mice. Stem Cell Res. 5, 267–270. https://doi.org/10.1016/j.scr.2010.07.004 (2010).
-
Bellingrath, J. S., McClements, M. E., Fischer, M. D. & MacLaren, R. E. Programmable RNA editing with endogenous ADAR enzymes—a feasible option for the treatment of inherited retinal disease?. Front Mol Neurosci 16, 1092913. https://doi.org/10.3389/fnmol.2023.1092913 (2023).
-
Zhu, Y., Zhu, L., Wang, X. & Jin, H. RNA-based therapeutics: An overview and prospectus. Cell Death Dis. 13, 644. https://doi.org/10.1038/s41419-022-05075-2 (2022).
-
Wei, P. S. et al. Enhancing RNA-lipid nanoparticle delivery: Organ- and cell-specificity and barcoding strategies. J. Control Release 375, 366–388. https://doi.org/10.1016/j.jconrel.2024.08.030 (2024).
-
Zhang, Y. et al. The development of an in vitro assay to screen lipid based nanoparticles for siRNA delivery. J Control Release 174, 7–14. https://doi.org/10.1016/j.jconrel.2013.11.006 (2014).
-
de Groot, A. M. et al. Immunogenicity testing of lipidoids in vitro and in silico: Modulating lipidoid-mediated TLR4 activation by nanoparticle design. Mol. Ther. Nucleic. Acids 11, 159–169. https://doi.org/10.1016/j.omtn.2018.02.003 (2018).
-
Paunovska, K., Loughrey, D. & Dahlman, J. E. Drug delivery systems for RNA therapeutics. Nat. Rev. Genet. 23, 265–280. https://doi.org/10.1038/s41576-021-00439-4 (2022).
-
Debacker, A. J., Voutila, J., Catley, M., Blakey, D. & Habib, N. Delivery of oligonucleotides to the liver with GalNAc: From research to registered therapeutic drug. Mol. Ther. 28, 1759–1771. https://doi.org/10.1016/j.ymthe.2020.06.015 (2020).
-
Cui, H., Zhu, X., Li, S., Wang, P. & Fang, J. Liver-targeted delivery of oligonucleotides with N-acetylgalactosamine conjugation. ACS Omega 6, 16259–16265. https://doi.org/10.1021/acsomega.1c01755 (2021).
-
Savva, Y. A., Rieder, L. E. & Reenan, R. A. The ADAR protein family. Genome Biol. 13, 252. https://doi.org/10.1186/gb-2012-13-12-252 (2012).
-
Liu, J., Afroza, H., Rader, D. J. & Jin, W. Angiopoietin-like protein 3 inhibits lipoprotein lipase activity through enhancing its cleavage by proprotein convertases. J Biol Chem 285, 27561–27570. https://doi.org/10.1074/jbc.M110.144279 (2010).
-
D’Souza, A. A. & Devarajan, P. V. Asialoglycoprotein receptor mediated hepatocyte targeting—strategies and applications. J. Control Release 203, 126–139. https://doi.org/10.1016/j.jconrel.2015.02.022 (2015).
-
Sago, C. D. et al. High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proc. Natl. Acad. Sci. USA 115, E9944-e9952. https://doi.org/10.1073/pnas.1811276115 (2018).
-
Godoy, P. et al. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch Toxicol 87, 1315–1530. https://doi.org/10.1007/s00204-013-1078-5 (2013).
-
Arez, F. et al. Heterotypic spheroids as a strategy for 3D culture of cryopreserved primary human hepatocytes in stirred-tank systems. SLAS Discov. https://doi.org/10.1016/j.slasd.2025.100210 (2025).
-
Caddeo, A., Maurotti, S., Kovooru, L. & Romeo, S. 3D culture models to study pathophysiology of steatotic liver disease. Atherosclerosis 393, 117544. https://doi.org/10.1016/j.atherosclerosis.2024.117544 (2024).
-
Tutty, M. A., Movia, D. & Prina-Mello, A. Three-dimensional (3D) liver cell models—a tool for bridging the gap between animal studies and clinical trials when screening liver accumulation and toxicity of nanobiomaterials. Drug. Deliv. Transl. Res. 12, 2048–2074. https://doi.org/10.1007/s13346-022-01147-0 (2022).
-
Frazier, K. S. Antisense oligonucleotide therapies: The promise and the challenges from a toxicologic pathologist’s perspective. Toxicol. Pathol. 43, 78–89. https://doi.org/10.1177/0192623314551840 (2015).
-
Dhuri, K. et al. Antisense oligonucleotides: An emerging area in drug discovery and development. J. Clin. Med. https://doi.org/10.3390/jcm9062004 (2020).
-
Gagliardi, M. & Ashizawa, A. T. The challenges and strategies of antisense oligonucleotide drug delivery. Biomedicines https://doi.org/10.3390/biomedicines9040433 (2021).
-
Hammond, S. M. et al. Delivery of oligonucleotide-based therapeutics: Challenges and opportunities. EMBO Mol. Med. 13, 13243. https://doi.org/10.15252/emmm.202013243 (2021).
Acknowledgements
Editorial support for the preparation of this manuscript (under the guidance of the authors) was provided by Becky Vickers and Suzanne Flowers, Apothecom (UK) and funded by ProQR Therapeutics.
Funding
This work was supported by ProQR Therapeutics, Leiden, The Netherlands. Editorial support was funded by the study sponsor, ProQR Therapeutics.
Ethics declarations
Competing interests
MA, NHPC, DAP, BA, PCdV, MM, LM, MvB and GJP are employees of ProQR.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Aguila, M., Cnubben, N.H.P., Parfitt, D.A. et al. Evaluation of in vitro liver models and non-human primates for predicting ADAR RNA editing efficiency in human liver. Sci Rep 15, 41494 (2025). https://doi.org/10.1038/s41598-025-25280-1
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-25280-1