
Introduction
The ECM is vital to tissue development as a source of structural support for tissues and a source of key developmental signals through transduction of mechanical cues, transduction of biochemical cues, and sequestration of growth factors. These cues are essential to development as knockouts of many ECM proteins are embryonic lethal1,2,3 and the ECM changes with development4,5,6. For this reason, many groups have used the ECM to guide stem cell behavior. Decellularized cardiovascular7,8,9,10,11 kidney12,13 and liver14 ECM have been shown to direct stem cell behavior, as have other tissue-derived ECM15,16. However, the complex composition of decellularized ECM limits our ability to identify particular matrix component(s) essential to elicit the desired biological response and could pose roadblocks to clinical applications. An alternative strategy is to utilize individual, defined ECM proteins, alone or in combination, in a bottom-up approach. The advantage of this method is that the matrix is defined and consistent, allowing study of the effects of individual proteins. A number of studies have used this method to direct stem cell behavior towards a variety of cell types17,18,19,20,21,22,23 and here we use a similar approach to drive endothelial specification.
The vascular basement membrane is a specialized sheet of ECM which is found at the basal surface of endothelial cells which line blood vessels. It is primarily composed of type IV Collagen (CIV), Nidogen, and Laminins. CIV is essential for basement membrane stability in vivo24 with aneurysm being a common feature of CIV deficiency syndromes25. In vitro, CIV has been used as a substrate for endothelial differentiation26; however, without additional biochemical stimuli it does not induce significant endothelial differentiation, though it may improve differentiation when used in combination with other ECM components27,28. Nidogens serve a primarily structural role in the basement membrane29 leading to vascular instability in nidogen knockout mice30. Due to their structural role, and the fact that endothelial function in nidogen deficient mice is normal, nidogens have not been used in endothelial differentiation, though nidogen is present in many laminin preparations31,32. Laminins are cross shaped proteins composed of three distinct chains, which confer subtype identity. For example, Laminin 421 contains the α4, β2, and γ1 chains. Laminin 111 (LN111) remains the most studied laminin due to easy access and its presence in Matrigel33. However, Laminin 111 is not found in the vascular basement membrane34. During development, Laminin 411 (LN411) is the primary laminin in the vascular basement membrane. After birth, Laminin 511 (LN511) begins to be expressed, promoting tight junction formation. Laminin 111, prevalent in Matrigel, is commonly used as a substrate for endothelial differentiation35,36. In our previous work, we found that Laminin 411 coating resulted in significantly more endothelial differentiation than Matrigel or Laminin 11128. However, despite the increase in endothelial differentiation, efficiency was still relatively low, necessitating further improvements.
Here we utilize a Design of Experiments approach to optimize an ECM formulation for endothelial differentiation and identify key signaling pathways that are triggered during this process and which are responsible for the differential effect of individual ECM proteins on stem cell differentiation. The optimized ECM formulation was further adapted as a bioink for 3D bioprinting. In this context, we demonstrate for the first time the ability to spatially define stem cell differentiation in a bioprinted scaffold using specified ECM formulations.
Results
Optimization of an extracellular matrix formulation to support endothelial differentiation using DoE
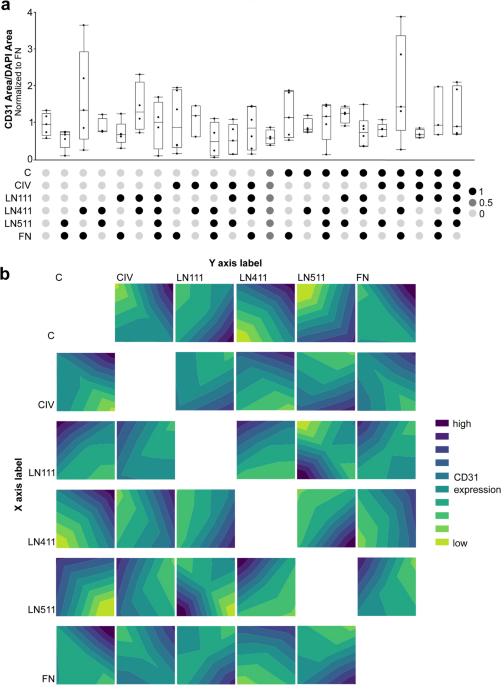
The first step of the Design of Experiments (DoE) approach was a set of factorial experiments. In these experiments, each factor, in this case protein coating concentration, is set to two different levels, low (-) and high (+). For Collagen I (C), CIV, LN111, LN411, and LN511 the low concentration was set to 0. The low Fibronectin (FN) concentration was set to 22 µg/mL, which was the lowest concentration on which human induced pluripotent stem cells (hiPSCs) showed good attachment and survival. Without the added FN, attachment was very poor on some proteins of interest, which would have made analyzing their effects on differentiation impossible. The high concentrations were set based on literature values for ECM coating for cell culture applications to 35.5 µg/mL, 28 µg/mL, 15.8 µg/mL, 0.8 µg/mL, 0.8 µg/mL, and 75 µg/mL for C, CIV, LN111, LN411, LN511, and FN respectively. One center point (000000 in Table 1) was added to the factorial experiments to account for nonlinear responses to ECM concentration. The response was defined as expression of the endothelial marker cluster of differentiation 31 (CD31) as detected by immunofluorescence.
The factorial experiments were analyzed up to 2nd order interactions which were graphed in contour plots (Fig. 1). Parameter estimates associated with the Design of experiments (DoE) analysis were also evaluated. These estimates showed that C, CIV, and LN411 had positive and significant associations with CD31 expression with the highest levels of CD31 expression corresponding to the highest concentrations of these proteins. Thus, the high concentration of these proteins was increased by a factor of 2 for the subsequent response surface regression. LN111 and LN511 also showed significant associations with CD31 expression. However, the maximum expression occurred at the lowest concentrations of these proteins, which was set to 0, so these proteins were eliminated. The trend for FN was similar to the first group of proteins, with the highest differentiation seen at the highest FN concentrations. However, because this association was not significant the high concentration of FN was kept the same.

Stage 1 of the Design of Experiments Approach for Optimizing ECM for Endothelial Differentiation. Abbreviations used: Collagen I (C), Collagen IV (CIV), Laminin 111 (LN111), Laminin 411 (LN411), Laminin 511 (LN511), Fibronectin (FN). (a) Graphical representation of CD31 area per DAPI area normalized to the FN control for the conditions indicated by the first stage of the DOE approach. The relative concentration of each protein is indicated by the heatmap below the graph for each condition. Concentrations vary between 0 (the lowest concentration for each protein) and 1 (the highest concentration for each protein). (b) Contour plots showing the relationship between CD31 expression and concentration of individual proteins. The lowest level of CD31 expression is indicated by the bright green and the highest by the dark blue.The resolution of all figures is low, but figure #1 is especially low. We have attached the high resolution figure 1 here.
Following the factorial experiments, a response surface regression was used to identify the theoretical optimum ECM formulation. An on face central composite design was used to add levels of ECM concentration for the four proteins brought through to this stage: C, CIV, LN411, and FN as shown in Table 2. Following this set of experiments, a regression analysis was performed to determine the coefficients of the response surface relating ECM exposure to CD31 expression based only on significant terms up to the 3rd order (Fig. 2). The resulting parameter estimates shown in Fig. 2b and the values at which a maximum CD31 expression was predicted were identified. This prediction solution of 35.6 µg/mL C, 67.2 µg/mL CIV, 0.9 µg/mL LN411, and 22 µg/mL FN is the Theoretical Optimized formulation (TheO), with the concentrations of CIV and LN411 having the largest effect sizes and FN having a medium effect size. While the concentration of C was predicted to be significant, the effect size was predicted to be small.

Stage 2 of the Design of Experiments Approach for Optimizing ECM for Endothelial Differentiation. Abbreviations used: Collagen I (C), Collagen IV (CIV), Laminin 411 (LN411), Fibronectin (FN). (a) Graphical representation of CD31 staining per DAPI area normalized to the FN control for the conditions indicated by the second stage of the DOE approach. The relative concentration of each protein is indicated by the heatmap below the graph for each condition. Concentrations vary between 0 (the lowest concentration for each protein) and 1 (the highest concentration for each protein). (b) A table showing parameter estimates for significant terms in the model produced by stage 2 DOE results. Terms up to the third order were included. (c) Graphical representation of the main effect size for the concentration of each ECM protein. (d) Graphical representation of the total effect size for the concentration of each ECM protein.
Validation of TheO
The TheO formulation was compared to the ++++ formulation (i.e., maximum concentrations of all four proteins) to determine whether optimal concentration of ECM proteins was essential to drive endothelial specification. TheO was also compared to LN411 + FN, our previously published formulation known to support endothelial specification and FN alone, the condition representing the minimum amount of ECM needed to support hiPSC attachment. Additionally, TheO without FN (TheO-FN) was evaluated as the model used did not allow for concentrations of FN below 22 µg/mL to be evaluated, as again that was the lowest concentration of FN on which consistent cell attachment was seen. Since the TheO formulation indicated the lowest FN concentration would lead to the highest CD31 expression and the relatively high concentrations of other proteins could allow for sufficient cell survival on the coating, TheO-FN was included to determine the necessity of FN inclusion in the formulation. Interestingly, the TheO formulation resulted in very little endothelial differentiation, around the level seen on FN alone (Fig. 3a, b). However, TheO-FN led to a high level of endothelial differentiation (Fig. 3a, b). The fact that the TheO condition did not produce the expected result of high levels of differentiation could be attributed to several factors including the complexity of cell-ECM interactions or high variability in levels of differentiation. However, the reason for the discrepancy is likely due to use of an on face central composite design which does not allow for accurate modeling outside of the original parameter space. Differentiation on TheO-FN was also significantly higher than on LN411 + FN, which was previously shown to be significantly better than Matrigel28. TheO-FN also showed significantly more differentiation than was seen with the maximum concentration of all proteins (++++) or the maximum concentration of C, CIV, and LN411 without FN (+++-). Due to the significant level of differentiation on TheO-FN compared to the other conditions, it will now be referred to as Endothelial Optimized or EO.
Additionally, since the effect of C concentration was predicted to be small, TheO-C and EO-C were also tested. As predicted by the model, there was a general trend of less CD31 expression when no C was included but no significant difference was seen between the condition with versus without the inclusion of Collagen I in the formulation (Fig. 3a, b).

Validation of the theoretical optimized ECM formulation for endothelial differentiation. Abbreviations used: Collagen I (C), Collagen IV (CIV), Laminin 411 (LN411), Fibronectin (FN). (a) Representative immunofluorescence images showing CD31 (red) and DAPI (blue) staining following differentiation on ECM formulations. The formulations are indicated with colored circles in the top left of the images. The key can be found below part b of the figure. (b) Graphical representation of CD31 area per DAPI area normalized to the FN control for conditions indicated by the heatmap below the graph. The key is below the heatmap and is the same as in part a. * p < 0.05 ***p < 0.001 The large bracket indicates that all conditions within the bracket were significantly different from the indicated condition.
Factors that further enhance endothelial specification on EO
Vascular Endothelial Growth Factor (VEGF) is a well-known inducer of endothelial differentiation. VEGF was added to differentiations on TheO and EO ECM formulations in two ways. First, to capitalize on the VEGF binding capacity of ECMs28,37,38 10 ng/mL VEGF was added to wells coated with ECM, incubated for 30 min at room temperature, and washed prior to adding hiPSCs. ECM-bound VEGF significantly improved differentiation on the TheO formulation to the level of differentiation on EO, which was unchanged with VEGF addition (Fig. 4a). Second, VEGF was added to the differentiation media starting on Day 0 when the glycogen synthase kinase 3 inhibitor CHIR99021 (CHIR), the initiator of the differentiation protocol, was first added. This condition showed similar results to the VEGF pre-incubation condition (Figure S1). Because the results were similar, all future VEGF experiments were conducted with VEGF pre-incubation as this method capitalizes on the ability of ECM to sequester proteins, therefore using significantly less protein, and reducing cost. The similarity of the two VEGF conditions could be the result of many different factors including timing of VEGF exposure or ECM presentation of VEGF signals, which, while outside the scope of this work, could present interesting insights into VEGF signaling and warrants further investigation.
In the protocol used for endothelial differentiation, cells are allowed to attach in mTeSR for 2 days prior to administering CHIR to initiate differentiation. This means that the timing of the ECM signal and the small molecule signal for differentiation is offset. ECM matching the original coating was added to the media on the first day of CHIR treatment to determine if synchronous additional ECM exposure would improve endothelial differentiation. Differentiation was significantly improved on TheO, but not to the level of EO or EO + ECM, which were not significantly different from each other (Fig. 4b). This pattern may indicate that FN, which is present in TheO but not EO, could have a timing-dependent effect on endothelial differentiation whereas the other ECM components promote endothelial differentiation regardless of timing.
The role of TGFβ signaling in EO-driven endothelial specification
Since the presence or absence of FN has a significant impact on endothelial differentiation, the mechanism responsible for this outcome was investigated. FN is known to activate TGFβ signaling39 which has been shown to be detrimental to endothelial differentiation in other contexts40. To determine if TGFβ signaling was involved in this effect, the TGFβ inhibitor SB431542 (SB) was added beginning on day 0 of differentiation, when CHIR was first added. The addition of SB had no significant effect on endothelial differentiation on EO, which does not contain FN. However, differentiation was significantly improved on TheO which does contain FN (Fig. 4c). TGFβ was then added to the differentiations on both ECMs starting on day 0. This addition had no significant effect on differentiation on TheO; however, differentiation on EO was significantly reduced (Fig. 4d). Together, these results indicate that the difference in endothelial differentiation on the TheO and EO ECM coatings is driven at least in part by FN activated TGFβ signaling, which inhibits ECM-guided endothelial specification.
Combinatorial conditions to improve ECM-guided endothelial differentiation
In order to further augment ECM-guided endothelial differentiation, the components that improved differentiation on either TheO or EO were added in combination. On the TheO formulation, the combinatorial addition of VEGF and ECM added on day 0 of differentiation led to the highest level of differentiation. Interestingly, though SB alone improved endothelial differentiation, it had a different effect when added in combination with either VEGF incubation or ECM addition, reducing the level of differentiation. This counterintuitive effect has been previously reported for VEGF and SB40 and is likely due to the highly context-dependent nature of TGFβ signaling. The complexity of these pathways is further explored in Fig. 4. On the EO formulation, all combinatorial additions increased the level of endothelial differentiation to a similar degree above the level of EO with any of the additives alone, which did not differ significantly from EO (Fig. 4e). As shown in Fig. 4f, conditions containing SB led to a more uniform layer of endothelial cells, whereas the condition containing just VEGF and added ECM shows a more vessel like structure. This is likely due to changes in the number of contaminating smooth muscle cells as SB (i.e., TGFβ inhibition) is known to inhibit smooth muscle differentiation41,42. While there is not a significant difference in the relative quantity of endothelial cells generated between differentiation conditions, the different 3D architectures could lend themselves to differing applications.

TGFβ and VEGF signaling promote ECM-driven endothelial specification. Abbreviations used: Collagen I (C), Collagen IV (CIV), Laminin 411 (LN411), Fibronectin (FN), Theoretical Optimized ECM (TheO), Endothelial Optimized ECM (EO). (a) Graphical representation of CD31 area per DAPI area normalized to the FN control for TheO and EO with and without the addition of VEGF incubation prior to cell seeding. **p < 0.01. (b) Graphical representation of CD31 area per DAPI area normalized to the FN control for TheO and EO with and without the addition of ECM proteins matching the original coating for the condition added on day 0 of differentiation. *p < 0.05 **p < 0.01. (c) Graphical representation of CD31 area per DAPI area normalized to the FN control for TheO and EO with and without the addition of the TGBβ inhibitor SB beginning on day 0 of differentiation. *p < 0.05 **p < 0.01. (d) Graphical representation of CD31 staining per DAPI area normalized to the FN control for TheO and EO with and without the addition of TGFβ beginning on day 0 of differentiation. **p < 0.01 ***p < 0.001. (e) Graphical representation of CD31 area per DAPI area normalized to the FN control for TheO and EO with combinatorial addition of SB, VEGF, and ECM matching the original coating as indicated by the heatmap below the x axis of the graph. The light circles (0) indicate that the represented component was not included for that condition and dark circles (1) indicate that the component was included. *p < 0.05 **p < 0.01***p < 0.001. (f) Representative images of EO conditions from part e showing the extent of CD31 staining (red) with DAPI (blue). Notably, the morphology of the cells in the VEGF ECM condition is significantly different from the conditions which include SB.
EO ECM validation in 3D: evaluation of endothelial differentiation in bioprinted constructs
While 2D culture systems are commonly used for stem cell differentiation, they lack the structural and biomechanical cues present in native tissues. In contrast, 3D environments provide a more physiologically relevant context, enabling improved cell-matrix interactions and promoting lineage-specific differentiation. Various strategies, including hydrogel encapsulation and self-assembled spheroids, have been developed to guide stem cells toward endothelial fates43. More recently, 3D bioprinting has emerged as a powerful tool to replicate the complexity of vascular networks due to its capacity for high-resolution spatial control and its compatibility with specialized bioinks for multicellular, perfusable constructs44. Among the bioprinting modalities, extrusion-based techniques, particularly the FRESH (Freeform Reversible Embedding of Suspended Hydrogels) method, allow direct printing of low-viscosity, ECM-based bioinks into a thermoreversible support bath, eliminating the need to print sacrificial materials. With this approach, vascularized scaffolds with fully biologic materials have been generated45. Integrating this approach with ECM formulations optimized for stem cell differentiation, such as the EO bioink, allows spatially defined stem cell patterning within bioprinted scaffolds.
Thus, to assess whether the optimized EO formulation maintained the capacity to drive endothelial specification in 3D environments, gelatin methacryloyl (GelMA) bioinks were combined with ECM-hiPSC mixtures and bioprinted using a support bath as previously described46,47. The specific ECM mixtures used were EO, a previously published cardiomyocyte-optimized ECM (CO)48 the cardiomyocyte-optimized ECM with doubled FN (CO-2FN), and no ECM (GelMA only) as a control. The structures printed were rectangular prisms of dimensions 5 mm x 5 mm with 1.8 mm thickness. The hiPSCs printed in the structure were subjected to the endothelial differentiation protocol used for the DoE experiments (no added VEGF or SB). Samples were then stained for CD31 to reveal vessel structures. Connected vascular networks were observed in the EO constructs (Fig. 5a); however, only segmented vessel-like structures were observed in the CO constructs, and there was very little vessel formation in the CO-2FN constructs. The enhanced endothelial differentiation induced by EO was confirmed by area quantification, where EO showed a significantly higher CD31 area ratio compared to all the other groups including the no ECM constructs (Fig. 5b, left). Further morphology analysis, including vessel length and branch points per area (Fig. 5b, middle and right) showed the same trends, indicating more developed vasculature in the EO group. Interestingly, while there is no significant difference, the no ECM control showed better endothelial differentiation compared to both the CO and the CO-2FN groups, which could be due to the inhibitory effect of FN on endothelial differentiation observed in the 2D experiments.
In addition to investigating the effects of individual ECM formulations on endothelial differentiation in 3D, bioprinted constructs with EO on one half of the structure and no ECM on the other were designed and fabricated (Fig. 5c, top row) to determine whether endothelial differentiation could be spatially controlled by defining ECM distribution. One of the EO components, CIV, was stained to show distribution of the ECM. While most of the area on the EO side was CIV positive as expected (81.29 ± 11.08%, Fig. 5c, bottom left; 5d, top row), the control side also showed substantial CIV deposition (30.26 ± 12.09%) at the end of the culture period. This likely reflects endogenous CIV secreted by cells during vasculogenesis as no CIV was observed on the control side just after printing or before initiating endothelial differentiation (Figure S2).
To focus on the effect of this exogenously secreted CIV, CD31 expression on the EO side was compared to the areas of the control side with and without CIV expression separately. Although the EO side of the constructs exhibited significantly higher CD31 expression compared to the control side without CIV expression (Fig. 5c, bottom right; 5d, bottom row), the CIV positive area of the control side showed even greater CD31 expression than the EO side overall (Figure S3). Notably, on the control side of constructs, the CIV positive area mostly arose from clumps of cells and ECM sprouting out from the printed structure. These structures were not observed in the constructs containing a single ECM formulation (Fig. 5d, top right), suggesting that these vasculogenesis-supporting niches were neither formed because of the GelMA bioink itself nor the endothelial differentiation protocol, and that a more complicated mechanism such as paracrine signaling or ECM remodeling may be involved in the formation of these sprouting structures. Overall, these results demonstrated that the EO ECM formulation could be applied to bioprinted constructs and maintain the capacity to enhance endothelial differentiation in 3D even at points remote from initial spatial deposition.

EO promotes endothelial differentiation in bioprinted constructs. Abbreviations used: Endothelial Optimized ECM (EO), Cardiomyocyte Optimized ECM (CO), Cardiomyocyte Optimized ECM with doubled Fibronectin concentration (CO-2FN), Collagen IV (CIV), Control area without collagen IV expression (Ctrl_CIV-). (a) Representative immunofluorescence images showing CD31 (red) and DAPI (blue) staining in bioprinted constructs with different ECM formulations. (b) Graphical representation of CD31 area ratio, total vessel length per area, and branch point per area for bioprinted constructs with different ECM formulations (n = 6 for EO; n = 3 for the other conditions). *p < 0.05 **p < 0.01 ***p < 0.001. (c) Schematic image showing the design of the dual component construct for spatially controlled EO distribution (upper left); a photograph of a bioprinted construct with red dye in the EO formulation for better visualization (upper right; scale = 5 mm), and graphical representation of CIV area ratio for the EO side and the no ECM control side of the dual component constructs (bottom, left, n = 4), as well as CD31 area ratio for the EO side and the no ECM control side without CIV expression of the dual component constructs (bottom right, n = 4). *p < 0.05 **p < 0.01. (d) Representative immunofluorescence images showing CIV (green) staining in the EO area and the control area of the dual component constructs (top row), as well as representative immunofluorescence images showing CIV (green), CD31 (red) and DAPI (blue) staining in the EO area and the control area without CIV of the dual component constructs (bottom row).
Discussion
In this study, we utilized a Design of Experiments framework to optimize ECM coatings to support endothelial differentiation of hiPSCs. This approach identified a formulation comprised of Collagen I, Collagen IV, and Laminin 411, termed Endothelial Optimum (EO), capable of promoting high levels of endothelial differentiation compared to the theoretical optimized formulation or Laminin 411 + FN. As we previously demonstrated that LN411 + FN outperformed Matrigel in inducing differentiation28 EO represents a significant advancement, providing a defined and efficient alternative for endothelial differentiation. These findings deepen our understanding of ECM-cell interactions in directing stem cell fate, build upon prior work leveraging ECM for vascular lineage commitment, and offer a powerful tissue engineering strategy to incorporate endothelial specification via 3D bioprinting of the EO formulation.
The EO formulation contains three ECM proteins, two of which (CIV and LN411) are found in the vascular basement membrane during development49. Interestingly, LN111, which is not expressed in the vascular basement membrane34 and LN511, which begins to be expressed postnatally34 had negative associations with endothelial differentiation indicating that there is a specificity to the response both in terms of the Laminin subtype, and timing of expression. This underscores the importance of understanding Laminin subtypes as separate entities rather than focusing on LN111, as is common due to its prevalence in tumor-derived ECMs like Matrigel. In future works, it will be important to evaluate laminin subtypes independently, given their diverse in vivo functions and varied effects on cell behavior in vitro.
ECM primarily interacts with cells through cell surface integrins clustered at focal adhesions. In previous work, we found that Laminin 411-integrin interactions converge on β-catenin, activating mesodermal genes or binding to the Notch intracellular domain to induce arterial gene expression50,51,52,53,54. Another way in which the ECM interacts with cells is through growth factor presentation or sequestration. Fibronectin, a key mediator of TGFβ signaling39 has been shown to inhibit endothelial differentiation40,55. To probe the role of TGFβ in the differential outcomes between EO (lacking Fibronectin) and TheO (containing Fibronectin), we employed the TGFβ inhibitor SB. While SB minimally affected differentiation on EO, it significantly enhanced differentiation on TheO, implicating TGFβ as a factor in the latter’s reduced differentiation efficiency. However, combining SB with other interventions, such as additional ECM proteins or VEGF, yielded distinct outcomes depending on the condition. This complexity, consistent with the TGFβ paradox, highlights the dual nature of TGFβ signaling, where its downstream effectors can either promote or inhibit endothelial differentiation depending on contextual cues56. For example, Activation of Smad2/3 by TGFβ increases expression of VEGF57,58 which increases endothelial differentiation. Smad2/3 also activates RSmad and Smad4 which, together with β-catenin, activate traditional TGFβ target genes, including the production of ECM such as collagens and fibronectin and TIMP3, which inhibits VEGFR2 and can therefore decrease endothelial differentiation59,60. Additionally, in the presence of BMP2/4 and Smad1/5/8 activation, Smad4, which is activated by TGFβ, can induce expression of Mesodermal and Endothelial genes61,62. We attempt to summarize our contribution and that of the literature to ECM-guided endothelial specification in a schematic (Fig. 6).
A major advantage of ECM-driven differentiation is that insoluble ECM can be spatially patterned using 3D printing, allowing for spatial control of differentiation. This is much more difficult to achieve with small, soluble molecules and growth factors, which easily diffuse away from their initial positions or must be tethered to other molecules, potentially inhibiting function. In the case of endothelial cells, tissue vascularization could be achieved by spatial patterning of cues for endothelial differentiation with those to support another cell type of interest. For cardiac tissue, a previously optimized ECM that supports cardiomyocyte specification48 could be patterned with the EO formulation to drive vascularized cardiac tissue. This could also be extended to additional cell types such as epicardial or endocardial cells to add further tissue complexity or to other complex systems such as the lungs, where patterning of vasculature and alveolar tissues is essential to tissue function. Here we demonstrate this possibility for the first time by 3D printing a bioink spiked with EO adjacent to a bioink lacking the EO formulation. Cell viability was maintained throughout both bioinks, but the EO-containing portion of the structure showed robust endothelial specification. Interestingly, over time, new CIV was deposited in spatial locations distant from the initial deposition of the EO formulation. This outcome could also be advantageous for tissue engineering applications, where infiltration of vessels into forming tissues is desired.
In conclusion, this study identifies EO, a defined ECM formulation composed of Collagen I, Collagen IV, and Laminin 411, as a highly effective platform for endothelial differentiation of hiPSCs. In the course of these studies, we made further strides in understanding the mechanisms by which ECM proteins can drive endothelial specification. We also made strides to eliminate reliance on Matrigel by harnessing the advantages of insoluble ECM for spatially controlled differentiation. In the end, our EO formulation lends itself to a broad range of potential applications including tissue vascularization, which has been a significant challenge for the field of tissue engineering.

Proposed Mechanism for ECM guided endothelial differentiation. A schematic showing the proposed mechanism of ECM guided endothelial differentiation in the context of EO and TheO. Signaling through Integrins, ILK, and FAK, Wnt, TGFβ, and Notch all converge at binding to active β-catenin28. TGFβ target genes can both activate VEGF signaling by inducing VEGF expression or inhibit VEGF expression by upregulating TIMP3 which inhibits VEGFR2 (VEGF receptor 2)57,60,63. Additionally, ECM production triggered by TGFβ activation can have complex effects on cell behavior39,60,62,64,65.
Methods
Human iPSC maintenance
hiPSCs (CCND2, human cardiac fibroblast-derived induced pluripotent stem cells expressing cyclin D2 under the myosin heavy chain promoter, authenticated by karyotyping, male) were kindly provided by the Zhang Laboratory who collected the human fibroblast source cells in accordance with the University of Minnesota Human Subjects Research Institutional Review Board66,67,68 and were maintained on Matrigel (Corning, cat# 354277) coated plates at 37 °C. mTeSR1 medium (STEMCELL, cat # 85850) was changed daily and cells were passaged at a ratio of 1:12 using ReLeSR (STEMCELL, cat# 05872) every 3–4 days at 80–90% confluency. hiPSCs were used between passages 40–70.
ECM coating
All coatings were prepared to the concentrations necessary in Dulbecco’s phosphate-buffered saline (DPBS) with calcium and magnesium. Solutions were kept on ice during preparation. 250 µL of solution was added to each well of a 48-well plate which was then incubated at 4 °C for 24 h. Wells were then washed twice with Phosphate buffered saline (PBS) and cells were seeded immediately to prevent drying of the protein coatings.
Endothelial differentiation in 2D culture
hiPSCs were singularized using Accutase (Millipore Sigma, cat # A6964) and seeded onto coated plates at a density of 263,000 cells/cm2 in mTeSR with 5 µM ROCK inhibitor (VWR, cat# 103538-728). mTeSR was replaced at Day − 1. On Day 0, media was replaced with LaSR (DMEM F12 Advanced (Thermo-fisher cat# 12634010), 100 µg/mL Ascorbic Acid (Sigma Aldrich, cat# 113170-55-1), 2.5 mM Glutamax (Thermo Fisher, cat# 35050061)) with 8µM CHIR99021 (CHIR)(Sigma-Aldrich, cat# SML1046). Media containing CHIR was replaced on day 1, and then LaSR medium was replaced every other day beginning on day 235,36. Cells were analyzed at day 10 of differentiation.
Design of experiments approach
The first stage of the Design of Experiments (DoE) approach was a set of factorial experiments in which the concentration of each ECM protein (C, CIV, LN111, LN411, LN511, FN) was set to either a low (0,0,0,0,0,22 µg/mL) or high (35.5, 56, 15.8, 0.8, 0.8, 75 µg/mL) level respectively. A center point was also included to allow for nonlinear associations between factors and the response. The response was the level of CD31 expression seen at day 10 of differentiation. Data were collected from at least 4 wells from at least 3 experimental replicates. JMP software was used to produce the experiment set, create contour plots, and identify statistically significant parameters and interactions. This information was used to inform the choice of proteins and concentrations for the second stage.
The second stage of the DoE approach used a Response Surface Methodology to determine the optimal ECM formulation for endothelial differentiation. In this stage, different levels of each protein were evaluated as shown in Table 2 using a central composite design. JMP software was used to fit the data, conduct ANOVA, and sort parameter estimates to identify the theoretical optimized ECM formulation (TheO).
The TheO formulation was then validated by immunostaining for CD31 as compared to the high value of all ECM components, the FN control, and LN411 + FN previously published by the Ogle lab28. Additionally, the TheO formulation was evaluated without FN as the DoE approach did not allow for the value to be set lower than the low FN concentration, and without C as its effect was predicted to be small.
Endothelial differentiation augmentation and cell signaling
In order to augment endothelial differentiation and investigate cell signaling pathways, SB431542 (SB) (Stemgent, cat# 040010), Vascular Endothelial Growth Factor (VEGF)(EGM-2 Bulletkit, Lonza, cat# CC-3156), and Transforming Growth Factor β (TGFβ)(PeproTech, cat# 100 − 21) were added to the media with every media change beginning on day 1 of differentiation at concentrations of 2µM, 50ng/mL, and 5µM respectively. Additionally, 50 ng/mL VEGF was incubated with the ECM coating for 30 min at room temperature (VEGF incubation). Wells were washed twice with PBS immediately prior to cell seeding28. Finally, additional ECM proteins matching the ECM coating were added to the media on Day 1 of differentiation.
3D Bioprinting with ECM-based Bioinks
To create 3D constructs with the endothelial optimized ECM (EO), EO was combined with Gelatin methacryloyl (GelMA, kindly provided by the Bioprinting Facility, UMN) for bioprinting. First, singularized hiPSCs were resuspended in mTeSR1 with the ECM proteins and ROCK inhibitor (Y-27632 2 HCl (cat#S1049, Selleckchem), and the mixture was incubated at room temperature for 15 min. The cell-ECM solution was then mixed with the precursor ink containing 20% (w/v) GelMA and 1% (w/v) lithium phenyl-2,4,6-trimethylbenzoylphosphinate (LAP, Allevi). The final cell-laden bioink optimized for endothelial differentiation (EO bioink) was composed of 15 million cells/mL with 10% GelMA, 64.37 µg/mL C, 121.5 µg/mL CIV, 1.63 µg/mL LN411, 0.5% LAP, and 5 µM ROCK inhibitor. The total ECM content was increased to match the previously published cardiomyocyte-optimized bioink48 while maintaining the same ratio as in the coating solution used for 2D experiments. The cardiomyocyte-optimized bioink, the cardiomyocyte-optimized bioink with doubled FN, and the bioink with no ECM addition were also prepared for control groups.
To create the bioprinted constructs, bioinks were extruded through 27-gauge needles into a support bath at 32 °C (FRESH printing v2.069). Microparticles used for the support bath were prepared by gelatin-gum arabic coacervation as previously described47. After printing, 405 nm blue light was immediately applied to both top and bottom sides of the support bath for 20 s to crosslink the printed constructs, and the support bath was then removed via incubation at 37 °C for 30 min followed by 3 rinses with PBS. The bioinks were printed as 20 mm long, 5 mm wide, and 1.8 mm high constructs and then cut into 5 mm x 5 mm x 1.8 mm squares for experiments. Bioprinted constructs containing only EO, cardiomyocyte-optimized ECM, cardiomyocyte-optimized ECM with doubled FN, or no additional ECM were printed separately to compare their capacity to induce endothelial differentiation. Bioprinted constructs with dual components (EO and no ECM control, Fig. 5c) were also created to investigate the effects of EO in a spatial manner.
Endothelial differentiation in 3D bioprinted constructs
Bioprinted constructs were cultured in mTeSR1 with 5 µM ROCK inhibitor for the first day, followed by daily media changes with mTeSR1 for 12 days. On day 0 and day 1 of differentiation, the media was replaced with LaSR supplemented with 8 µM CHIR. Starting from day 2 of differentiation, media was removed and replenished with fresh LaSR every other day. Differentiated constructs were harvested on day 10 for analysis.
Immunofluorescence and image analysis
CD31 expression was determined by immunofluorescence (primary: mouse anti-human CD31, 1:40 dilution, Invitrogen, Cat# 14-0311-82, secondary: goat anti mouse Alexaflour 647, 1:500 dilution, Invitrogen, Cat# A21236). Cells were imaged on the Leica DMi8 microscope with a 10x objective. For all experiments, 3 × 3 tilescans were collected in the approximate center of each well. CD31 area and DAPI area were determined for each image using FIJI (2.0.0-rc-68/1.52e). All data was normalized to the FN control for each experiment in order to account for interexperimental variability associated with stem cell differentiation.
For bioprinted constructs, samples were sequentially incubated in 4% PFA for 24 h and 30% sucrose solution for 24 h prior to OCT embedding. Cryosectioning was then performed as previously described48. Samples were sectioned from top to bottom in 5–10 μm sections. Sections from the first and the last 200 μm of tissue depth were not used to avoid edge effects. For immunofluorescence, samples were first incubated with a buffer composed of 5% BSA, 0.1% Triton X-100, 2% goat serum, and 1% glycine for 2 h to block non-specific binding sites. CD31 was stained as described above for 2D cell culture. The EO/control dual constructs were also stained for CIV (primary: rabbit anti-human CIV, 1:100 dilution, Chemicon, cat#AB748; secondary: goat anti-rabbit Alexaflour 488, 1:500 dilution, Invitrogen, cat#A32731) to reveal EO distribution. Samples were mounted with a DAPI/1,4-diazabicyclo[2,2,2]octane (DABCO; Sigma-Aldrich, cat#D27802) mounting solution. Tilescan imaging of one whole section from each sample was performed using the Leica DMi8 microscope with a 10x objective. CD31, CIV, and DAPI areas were determined in FIJI (2.14.0/1.54f), and the vessel length and the branch point count of vascular networks were measured using the Anamorf plugin46,70. CIV staining was also imaged with a 4x objective to provide better visualization of ECM distribution (Fig. 4d, top row).
Statistics
Statistical Analysis was performed using JMP software. For all experiments, the assumption of equal variance was tested using a Bartlett test. If p > 0.05, ANOVA followed by a Tukey’s post hoc test were used to determine significance. If p < 0.05, Welch’s test followed by the Games-Howell test (Tukey HSD with Welch’s correction for unequal variance) were used. Significance was defined as p < 0.05 for all experiments.
Data availability
Data for the current manuscript will be provided on reasonable request and requests can be made to the corresponding author, Brenda Ogle, ogle@umn.edu.
References
-
Miner, J. H., Cunningham, J. & Sanes, J. R. Roles for laminin in embryogenesis: exencephaly, syndactyly, and placentopathy in mice lacking the laminin alpha5 chain. J. Cell. Biol. 143, 1713–1723. https://doi.org/10.1083/jcb.143.6.1713 (1998).
-
Rozario, T. & DeSimone, D. W. The extracellular matrix in development and morphogenesis: a dynamic view. Dev. Biol. 341, 126–140. https://doi.org/10.1016/j.ydbio.2009.10.026 (2010).
-
Yang, J. T. et al. Overlapping and independent functions of fibronectin receptor integrins in early mesodermal development. Dev. Biol. 215, 264–277. https://doi.org/10.1006/dbio.1999.9451 (1999).
-
Hanson, K. P. et al. Spatial and Temporal analysis of extracellular matrix proteins in the developing murine heart: a blueprint for regeneration. Tissue Eng. Part. A. 19, 1132–1143. https://doi.org/10.1089/ten.TEA.2012.0316 (2013).
-
Li, Z. et al. Proteome-wide and matrisome-specific alterations during human pancreas development and maturation. Nat. Commun. 12, 1020. https://doi.org/10.1038/s41467-021-21261-w (2021).
-
Jacobson, K. R. et al. Extracellular matrix protein composition dynamically changes during murine forelimb development. iScience 27, 108838. https://doi.org/10.1016/j.isci.2024.108838 (2024).
-
Ott, H. C. et al. Perfusion-decellularized matrix: using nature’s platform to engineer a bioartificial heart. Nat. Med. 14, 213–221. https://doi.org/10.1038/nm1684 (2008).
-
Chen, W. C. et al. Decellularized zebrafish cardiac extracellular matrix induces mammalian heart regeneration. Sci. Adv. 2, e1600844. https://doi.org/10.1126/sciadv.1600844 (2016).
-
Goldfracht, I. et al. Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta Biomater. 92, 145–159. https://doi.org/10.1016/j.actbio.2019.05.016 (2019).
-
Gaggi, G. et al. Decellularized extracellular matrices and cardiac differentiation: study on human amniotic Fluid-Stem cells. Int. J. Mol. Sci. 21 https://doi.org/10.3390/ijms21176317 (2020).
-
Kurokawa, S. et al. In vivo recellularization of xenogeneic vascular grafts decellularized with high hydrostatic pressure method in a Porcine carotid arterial interpose model. PLoS One. 16, e0254160. https://doi.org/10.1371/journal.pone.0254160 (2021).
-
Figliuzzi, M., Bonandrini, B. & Remuzzi, A. Decellularized kidney matrix as functional material for whole organ tissue engineering. J. Appl. Biomater. Funct. Mater. 15, e326–e333. https://doi.org/10.5301/jabfm.5000393 (2017).
-
Sobreiro-Almeida, R. et al. Decellularized kidney extracellular matrix Bioinks recapitulate renal 3D microenvironmentin vitro. Biofabrication 13 https://doi.org/10.1088/1758-5090/ac0fca (2021).
-
Jaramillo, M., Yeh, H., Yarmush, M. L. & Uygun, B. E. Decellularized human liver extracellular matrix (hDLM)-mediated hepatic differentiation of human induced pluripotent stem cells (hIPSCs). J. Tissue Eng. Regen Med. 12, e1962–e1973. https://doi.org/10.1002/term.2627 (2018).
-
del Alió, J. L. et al. Corneal stroma enhancement with decellularized stromal laminas with or without stem cell recellularization for advanced keratoconus. Am. J. Ophthalmol. 186, 47–58. https://doi.org/10.1016/j.ajo.2017.10.026 (2018).
-
Giobbe, G. G. et al. Extracellular matrix hydrogel derived from decellularized tissues enables endodermal organoid culture. Nat. Commun. 10, 5658. https://doi.org/10.1038/s41467-019-13605-4 (2019).
-
Pimton, P. et al. Fibronectin-mediated upregulation of α5β1 integrin and cell adhesion during differentiation of mouse embryonic stem cells. Cell. Adh Migr. 5, 73–82. https://doi.org/10.4161/cam.5.1.13704 (2011).
-
Singh, P. & Schwarzbauer, J. E. Fibronectin and stem cell differentiation – lessons from chondrogenesis. J. Cell. Sci. 125, 3703–3712. https://doi.org/10.1242/jcs.095786 (2012).
-
Jung, J. P., Hu, D., Domian, I. J. & Ogle, B. M. An integrated statistical model for enhanced murine cardiomyocyte differentiation via optimized engagement of 3D extracellular matrices. Sci. Rep. 5, 18705. https://doi.org/10.1038/srep18705 (2015).
-
Mohammadpour, A., Arjmand, S., Lotfi, A. S., Tavana, H. & Kabir-Salmani, M. Promoting hepatogenic differentiation of human mesenchymal stem cells using a novel laminin-containing gelatin cryogel scaffold. Biochem. Biophys. Res. Commun. 507, 15–21. https://doi.org/10.1016/j.bbrc.2018.10.121 (2018).
-
Shibata, S. et al. Selective Laminin-Directed Differentiation of Human Induced Pluripotent Stem Cells into Distinct Ocular Lineages. Cell Rep 25, 1668–1679 e1665, (2018). https://doi.org/10.1016/j.celrep.2018.10.032
-
Chan, A., Ma, S., Pearson, B. J. & Chan, D. Collagen IV differentially regulates planarian stem cell potency and lineage progression. Proc. Natl. Acad. Sci. U S A. 118 https://doi.org/10.1073/pnas.2021251118 (2021).
-
He, J., Zhang, N., Zhu, Y., Jin, R. & Wu, F. MSC spheroids-loaded collagen hydrogels simultaneously promote neuronal differentiation and suppress inflammatory reaction through PI3K-Akt signaling pathway. Biomaterials 265, 120448. https://doi.org/10.1016/j.biomaterials.2020.120448 (2021).
-
Pöschl, E. et al. Collagen IV is essential for basement membrane stability but dispensable for initiation of its assembly during early development. Development 131, 1619–1628. https://doi.org/10.1242/dev.01037 (2004).
-
Steffensen, L. B. et al. Basement membrane collagen IV deficiency promotes abdominal aortic aneurysm formation. Sci. Rep. 11, 12903. https://doi.org/10.1038/s41598-021-92303-y (2021).
-
Di Bernardini, E. et al. Endothelial lineage differentiation from induced pluripotent stem cells is regulated by microRNA-21 and transforming growth factor β2 (TGF-β2) pathways. J. Biol. Chem. 289, 3383–3393. https://doi.org/10.1074/jbc.M113.495531 (2014).
-
Hou, L. et al. Combinatorial extracellular matrix microenvironments for probing endothelial differentiation of human pluripotent stem cells. Sci. Rep. 7, 6551. https://doi.org/10.1038/s41598-017-06986-3 (2017).
-
Hall, M. L. et al. Laminin 411 mediates endothelial specification via multiple signaling axes that converge on β-catenin. Stem Cell. Rep. 17, 569–583. https://doi.org/10.1016/j.stemcr.2022.01.005 (2022).
-
Reinhardt, D. et al. Mapping of Nidogen binding sites for collagen type IV, Heparan sulfate proteoglycan, and zinc. J. Biol. Chem. 268, 10881–10887 (1993).
-
Bader, B. L. et al. Compound genetic ablation of Nidogen 1 and 2 causes basement membrane defects and perinatal lethality in mice. Mol. Cell. Biol. 25, 6846–6856. https://doi.org/10.1128/mcb.25.15.6846-6856.2005 (2005).
-
Baatout, S. & Cheţa, N. Matrigel: a useful tool to study endothelial differentiation. Rom J. Intern. Med. 34, 263–269 (1996).
-
Adelfio, M., Szymkowiak, S. & Kaplan, D. L. Matrigel-Free Laminin-Entactin matrix to induce human renal proximal tubule structure formation in vitro. ACS Biomater. Sci. Eng. 6, 6618–6625. https://doi.org/10.1021/acsbiomaterials.0c01385 (2020).
-
Kleinman, H. K. & Martin, G. R. Matrigel: basement membrane matrix with biological activity. Semin Cancer Biol. 15, 378–386. https://doi.org/10.1016/j.semcancer.2005.05.004 (2005).
-
Hallmann, R. et al. Expression and function of laminins in the embryonic and mature vasculature. Physiol. Rev. 85, 979–1000. https://doi.org/10.1152/physrev.00014.2004 (2005).
-
Lian, X. et al. Efficient differentiation of human pluripotent stem cells to endothelial progenitors via small-molecule activation of WNT signaling. Stem Cell. Rep. 3, 804–816. https://doi.org/10.1016/j.stemcr.2014.09.005 (2014).
-
Bao, X. et al. Chemically-defined albumin-free differentiation of human pluripotent stem cells to endothelial progenitor cells. Stem Cell. Res. 15, 122–129. https://doi.org/10.1016/j.scr.2015.05.004 (2015).
-
Wijelath, E. S. et al. Novel vascular endothelial growth factor binding domains of fibronectin enhance vascular endothelial growth factor biological activity. Circ. Res. 91, 25–31. https://doi.org/10.1161/01.res.0000026420.22406.79 (2002).
-
Usuelli, M., Meyer, T., Mezzenga, R., Mitsi, M. & VEGF VEGFR2 bind to similar pH-sensitive sites on fibronectin, exposed by heparin-mediated conformational changes. J. Biol. Chem. 296, 100584. https://doi.org/10.1016/j.jbc.2021.100584 (2021).
-
Dallas, S. L. et al. Fibronectin regulates latent transforming growth factor-beta (TGF beta) by controlling matrix assembly of latent TGF beta-binding protein-1. J. Biol. Chem. 280, 18871–18880. https://doi.org/10.1074/jbc.M410762200 (2005).
-
Xu, J. G. et al. Inhibition of TGF-β signaling in SHED enhances endothelial differentiation. J. Dent. Res. 97, 218–225. https://doi.org/10.1177/0022034517733741 (2018).
-
Tang, Y., Yang, X., Friesel, R. E., Vary, C. P. & Liaw, L. Mechanisms of TGF-β-induced differentiation in human vascular smooth muscle cells. J. Vasc Res. 48, 485–494. https://doi.org/10.1159/000327776 (2011).
-
Xu, J. G., Zhu, S. Y., Heng, B. C., Dissanayaka, W. L. & Zhang, C. F. TGF-β1-induced differentiation of SHED into functional smooth muscle cells. Stem Cell. Res. Ther. 8, 10. https://doi.org/10.1186/s13287-016-0459-0 (2017).
-
Rofaani, E., Mardani, M. W., Yutiana, P. N., Amanda, O. & Darmawan, N. Differentiation of mesenchymal stem cells into vascular endothelial cells in 3D culture: a mini review. Mol. Biol. Rep. 51, 781. https://doi.org/10.1007/s11033-024-09743-8 (2024).
-
Kim, J. J., Cho, D. W. & and Advanced strategies in 3D Bioprinting for vascular tissue engineering and disease modelling using smart Bioinks. Virtual Phys. Prototyp. 19, e2395470. https://doi.org/10.1080/17452759.2024.2395470 (2024).
-
Shiwarski, D. J. et al. 3D Bioprinting of collagen-based high-resolution internally perfusable scaffolds for engineering fully biologic tissue systems. Sci. Adv. 11, eadu5905. https://doi.org/10.1126/sciadv.adu5905 (2025).
-
Lin, W. H. et al. A bionic testbed for cardiac ablation tools. Int. J. Mol. Sci. 23 https://doi.org/10.3390/ijms232214444 (2022).
-
Komosa, E. R., Lin, W. H. & Ogle, B. M. Toward robust and reproducible pluripotent stem cell expansion in bioprinted GelMA constructs. IJB 11, (2024). https://doi.org/10.36922/ijb.4633
-
Kupfer, M. E. et al. Situ expansion, differentiation, and electromechanical coupling of human cardiac muscle in a 3D bioprinted, chambered organoid. Circ. Res. 127, 207–224. https://doi.org/10.1161/circresaha.119.316155 (2020).
-
Thyboll, J. et al. Deletion of the laminin alpha4 chain leads to impaired microvessel maturation. Mol. Cell. Biol. 22, 1194–1202. https://doi.org/10.1128/mcb.22.4.1194-1202.2002 (2002).
-
Hayward, P. et al. Notch modulates Wnt signalling by associating with Armadillo/beta-catenin and regulating its transcriptional activity. Development 132, 1819–1830. https://doi.org/10.1242/dev.01724 (2005).
-
Yamamizu, K. et al. Convergence of Notch and beta-catenin signaling induces arterial fate in vascular progenitors. J. Cell. Biol. 189, 325–338. https://doi.org/10.1083/jcb.200904114 (2010).
-
de la Puente, P. et al. Identification of ILK as a novel therapeutic target for acute and chronic myeloid leukemia. Leuk. Res. https://doi.org/10.1016/j.leukres.2015.09.005 (2015).
-
LaFoya, B. et al. Notch: A multi-functional integrating system of microenvironmental signals. Dev. Biol. 418, 227–241. https://doi.org/10.1016/j.ydbio.2016.08.023 (2016).
-
Park, J. K., Lee, T. W., Do, E. K., Moon, H. J. & Kim, J. H. Role of Notch1 in the arterial specification and angiogenic potential of mouse embryonic stem cell-derived endothelial cells. Stem Cell Res. Ther. 9, 197. https://doi.org/10.1186/s13287-018-0945-7 (2018).
-
Gong, H. et al. PAR1 scaffolds TGFβRII to downregulate TGF-β signaling and activate ESC differentiation to endothelial cells. Stem Cell. Rep. 7, 1050–1058. https://doi.org/10.1016/j.stemcr.2016.10.006 (2016).
-
Vignais, M. L. [Cellular signaling in response to tgfbeta: the paradox of a factor that blocks cell proliferation and enhances metastasis]. Bull. Cancer. 86, 903–910 (1999).
-
Pertovaara, L. et al. Vascular endothelial growth factor is induced in response to transforming growth factor-beta in fibroblastic and epithelial cells. J. Biol. Chem. 269, 6271–6274 (1994).
-
Sánchez-Elsner, T. et al. Synergistic Cooperation between hypoxia and transforming growth factor-beta pathways on human vascular endothelial growth factor gene expression. J. Biol. Chem. 276, 38527–38535. https://doi.org/10.1074/jbc.M104536200 (2001).
-
Qi, J. H. et al. A novel function for tissue inhibitor of metalloproteinases-3 (TIMP3): Inhibition of angiogenesis by blockage of VEGF binding to VEGF receptor-2. Nat. Med. 9, 407–415. https://doi.org/10.1038/nm846 (2003).
-
Leivonen, S. K. et al. TGF-β-elicited induction of tissue inhibitor of metalloproteinases (TIMP)-3 expression in fibroblasts involves complex interplay between Smad3, p38α, and ERK1/2. PLoS One. 8, e57474. https://doi.org/10.1371/journal.pone.0057474 (2013).
-
Bai, H. et al. BMP4 regulates vascular progenitor development in human embryonic stem cells through a Smad-dependent pathway. J. Cell. Biochem. 109, 363–374. https://doi.org/10.1002/jcb.22410 (2010).
-
Zhao, M., Mishra, L. & Deng, C. X. The role of TGF-β/SMAD4 signaling in cancer. Int. J. Biol. Sci. 14, 111–123. https://doi.org/10.7150/ijbs.23230 (2018).
-
Mandriota, S. J., Menoud, P. A. & Pepper, M. S. Transforming growth factor beta 1 down-regulates vascular endothelial growth factor receptor 2/flk-1 expression in vascular endothelial cells. J. Biol. Chem. 271, 11500–11505. https://doi.org/10.1074/jbc.271.19.11500 (1996).
-
Ai, W. J. et al. R-Smad signaling-mediated VEGF expression coordinately regulates endothelial cell differentiation of rat mesenchymal stem cells. Stem Cells Dev. 24, 1320–1331. https://doi.org/10.1089/scd.2014.0253 (2015).
-
Luo, K. Signaling cross talk between TGF-β/Smad and other signaling pathways. Cold Spring Harb Perspect. Biol. 9 https://doi.org/10.1101/cshperspect.a022137 (2017).
-
Zhang, L. et al. Derivation and high engraftment of patient-specific cardiomyocyte sheet using induced pluripotent stem cells generated from adult cardiac fibroblast. Circ. Heart Fail. 8, 156–166. https://doi.org/10.1161/circheartfailure.114.001317 (2015).
-
Zhu, W., Zhao, M., Mattapally, S., Chen, S. & Zhang, J. CCND2 overexpression enhances the regenerative potency of human induced pluripotent stem Cell-Derived cardiomyocytes: remuscularization of injured ventricle. Circ. Res. 122, 88–96. https://doi.org/10.1161/circresaha.117.311504 (2018).
-
Zhao, M. et al. Cyclin D2 overexpression enhances the efficacy of human induced pluripotent stem Cell-Derived cardiomyocytes for myocardial repair in a swine model of myocardial infarction. Circulation 144, 210–228. https://doi.org/10.1161/circulationaha.120.049497 (2021).
-
Lee, A. et al. 3D Bioprinting of collagen to rebuild components of the human heart. Science 365, 482–487. https://doi.org/10.1126/science.aav9051 (2019).
-
Barry, D. J., Williams, G. A. & Chan, C. Automated analysis of filamentous microbial morphology with AnaMorf. Biotechnol. Prog. 31, 849–852. https://doi.org/10.1002/btpr.2087 (2015).
Funding
We acknowledge our funding sources National Heart Lung and Blood Institute R01 HL137204, R01 HL160779, and T32 HL007741.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hall, M.L., Lin, WH. & Ogle, B.M. Optimizing extracellular matrix for endothelial differentiation using a design of experiments approach. Sci Rep 15, 24479 (2025). https://doi.org/10.1038/s41598-025-09256-9
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-09256-9
