
Introduction
Lignin is considered one of the most prevalent polymers on earth. It naturally exists in plants as an integral part of lignocellulose to give them structure and rigidity1. Most technical lignins available today are byproducts of pulping processes, primarily kraft, soda, and sulfite methods2. Among them, the kraft process is the industry standard for paper and pulp production on a global scale. Each year, 130 million tons of kraft pulp are produced which may cause serious pollution to aquatic ecosystems upon direct disposal3. This abundant polymer can be broken down and transformed into molecules with higher value, allowing us to extract even more valuable products from it4.
Lignin degradation can be accomplished by biological processes using entire cells (bacterial or fungal biomass) or ligninolytic enzymes5, offering an energy efficient, environmentally safe, and selective method6. While white-rot fungi produce powerful lignin degrading enzymes, their industrial applicability is limited7. In contrast, bacteria tolerate broader conditions and are easier for genetic modifications8,9.
Laccase, lignin peroxidase (LiP), manganese peroxidase (MnP), versatile peroxidase (VP), and dye-decolorizing peroxidase (DyP) are examples of ligninolytic enzymes that can be produced by fungi and bacteria found in plant biomass and soil10,11. In biochemical research on ligninolytic bacteria, LiPs, MnPs, and VPs have not been observed. No homologs are discovered even when sequencing ligninolytic bacteria’s genomes or proteomes, implying fungi as their only known source. It has lately become obvious that DyPs, a different class of peroxidase, are quite abundant in bacteria. Although DyPs are structurally unrelated to the common fungal peroxidases, they do share several catalytic characteristics as well as redox potentials and reactivities12.
The use of bacterial ligninolytic enzyme systems has been expanding rapidly over the last few decades due to their potential in industrial processes and environmental applications7,13. Recent research has focused on separating different ligninolytic bacteria for usage in a range of biotechnological processes, including bioremediation14, biofuel production15, and industrial enzyme development16. The ligninolytic enzymes hold great promise for industrial use in a variety of fields, such as food processing, cosmetics, aromatic compounds degradation, lignin valorization, and dye effluent decolorization17,18.
The textile industry uses thousands of synthetic dyes to make fabrics. Over one-third of these are not absorbed by the fabric and end up in wastewater, threatening aquatic biodiversity and human health due to their toxicity and carcinogenicity19. Although physicochemical treatment techniques have been employed to eliminate dyes, they are not ecologically friendly nor commercially practical. On the other hand, biological methods are cheaper, effective, and sustainable for the environment20.
Laccases and peroxidases, two ligninolytic enzymes, efficiently remove dyes, helping reduce textile-related water pollution21. Dye biodegradation entails intricate biochemical reactions that break down dye molecules into simpler, more biodegradable, and non-toxic byproducts20,22. Decolorization occurs when the dyes lose their chromophores as a result of repeated oxidation processes 23.
Recent approaches emphasize the use of microbes or their enzymes to effectively treat textile dye effluents, incorporating different immobilization methods and bioreactor designs tailored for systems utilizing immobilized agents24. There are many reports on ligninolytic bacteria and their applications in dye degradation such as decolorization of methylene blue dye by a ligninolytic enzyme-producing Bacillus thuringiensis 25, multiple dye degradation by a newly isolated ligninolytic bacteria Bacillus paramycoides21, and decolorization of textile azo dyes by Streptomyces albidoflavus 3MGH26.
Several studies have focused on optimizing the decolorization of various dyes, including Azure B (AB), methylene blue (MB), and Congo red (CR). Serratia liquefaciens, a lignin peroxidase-producing bacterium, demonstrated rapid decolorization of AB dye (100 mg/L) in mineral salt medium supplemented with glucose and yeast extract, achieving over 90% decolorization within 48 h27. Similarly, Bacillus sp. MZS10 achieved 93.55% decolorization of AB dye within 14 h in a stirred-tank fermenter, with the process being dependent on cell density and optimized medium conditions28. Regarding MB, thermophilic Bacillus licheniformis B3-15 and Bacillus sp. s7s-1 showed the ability to decolorize and detoxify MB dye in aqueous solution using preformed biofilms on polypropylene perforated balls (BBs). At optimized initial conditions (10 mg L−1 MB, pH 5.2 for B3-15 or pH 4 for s7s-1), the two strains enhanced their decolorization potential, reaching 96 and 67%, respectively, showing significant detoxification potential29. In case of CR, Alcaligenes faecalis H77, bacterium isolated from textile wastewater, proved to be highly effective in biodegrading CR. Through optimization process, it decolorized 92.5% of CR (25 mg/L) in 72 h at pH 6.7 and 35 °C30.
Additionally, some studies have employed genomic and transcriptomic approaches to identify ligninolytic enzymes in diverse bacterial strains. In this context, whole genome sequencing (WGS) has become a critical tool for uncovering bacterial ligninolytic genes, such as those encoding DyP-type peroxidases, multicopper oxidases (laccases), and accessory enzymes involved in aromatic compound metabolism31. This technique enhances the understanding of microbial degradation mechanisms and supports the development of effective bioremediation strategies32. For instance, the genome of Pseudomonas Hu109A revealed key lignin-degrading genes, including DyP-type peroxidases, laccase, and O-methyltransferases. It also carries gene clusters for both medium- and short-chain polyhydroxyalkanoates (PHA) synthesis, showing its ability to convert lignin into valuable products and its potential role for industrial applications33. In another study,
Klebsiella spp. strains isolated from lignocellulose-rich environments exhibited over 20 genes linked to lignin degradation pathways (including catalases, peroxidases, glutathione S-transferases and thiol peroxidases), with strain P3TM.1 achieving 98% decolorization of MB within 48 h, indicating robust ligninolytic activity34.
Ongoing research aims to discover novel dye-degrading strains, optimize degradation conditions, and create bacterial consortia to increase the efficiency of degradation35. Yet few studies link bacterial screening with genomic analysis to correlate genetic potential with actual bioremediation process.
The aim of this study is detection and characterization of soil bacteria for a candidate with potential ligninolytic activity in industrial applications, especially in dye effluent decolorization. The study also aims to apply WGS as a powerful tool to identify lignin-metabolizing enzymes and gain deeper insights into their genetic and functional potential.
Materials and methods
Source of bacterial isolates
The bacterial isolates were recovered from 55 soil samples collected from various agricultural sites throughout Egypt including the Rakta Pulp and Paper Company’s effluent treatment facility in Eltalbia, Alexandria, Egypt, these sites are supposed to be rich in ligninolytic bacteria. The soil samples were gathered in sterile plastic bottles of 100 mL volumes. The samples were obtained from the higher layers of the soil, which contain a substantial amount of the microbial community. After collection, samples were kept at 4 °C for subsequent use afterward36.
Isolation of bacteria with potential ligninolytic activities
Enrichment and isolates’ recovery
Firstly, the enrichment method was used to allow the bacteria present in low numbers to multiply and reach detectable levels. This was done by the addition of 5 g of soil sample aseptically in a previously autoclaved 250 mL Erlenmeyer flask containing 95 mL minimal salt medium enriched with kraft lignin (KL). The enrichment culture was composed of 1 g/L of commercially available KL (Sigma-Aldrich, USA) and 0.1 g/L yeast extract in phosphate buffered (pH 7) minimal salt medium of the following ingredients (g/L): 4.55 K2HPO4, 0.53 KH2PO4, 0.5 MgSO4, and 5 NH4NO3. After soil sample addition, the flasks were placed in an orbital shaking incubator at 30 °C and 200 rpm for one week. Secondly, for isolation of potential ligninolytic bacteria, a series of tenfold serial dilutions were made by adding 1 ml of enriched soil suspension to 9 ml of sterile normal saline and vortexing the mixture followed by transferring 100 μL aliquots of the produced dilutions to surface inoculation of minimal salt medium agar plates containing 1 g/L KL and 0.05 g/L nystatin (to inhibit fungal growth). The plates were then incubated at 30 °C until appearance of colonies, up to one week37. The grown colonies were suspected to have potential ligninolytic activity since KL was the sole carbon source in the medium.
Purification and storage of bacterial isolates
Based on the variation of colony morphology, individual colonies were picked up and purified using the streak plate technique on minimal salt medium agar plates containing KL. Multiple subcultures were performed to obtain colonies that are pure and distinct38. The recovered isolates were kept in 25% glycerol at −80 °C for further investigation.
Assessment of the recovered isolates for their ligninolytic activities using dye decolorization assays
Solid phase dye decolorization assay
This assay was performed by using some indicator dyes to evaluate the ability of the bacterial isolate to decolorize the dye. Luria–Bertani (LB) agar plates containing dye to final concentrations of 25 mg/L for each of Azure B (AB) and methylene blue (MB) or 50 mg/L for congo red (CR) were prepared. All of the indicator dyes were filter sterilized before being aseptically added to the autoclaved media39. Pure colonies of each tested isolate were streaked onto the LB agar plates containing dye. Following that, the plates were incubated for five days while being checked each day for the appearance of decolorization zones and/or stained growth (dye adsorption by the growth).
Liquid phase dye decolorization assay
The bacterial isolates which had the ability to decolorize all the indicator dyes used in the previous assay, were subjected to the liquid phase dye decolorization assay. For each test isolate, single colonies from the growth obtained on LB plate were used for separate inoculation of 250 mL conical flasks containing 25 mL aliquots of LB broth. The flasks were placed in an orbital shaking incubator at 30 °C and 200 rpm until reaching the exponential phase, then pre-filtered sterilized dyes were aseptically added separately to the flasks at final concentrations of 6 mg/L for each of AB and MB or 40 mg/L for CR. Incubation was resumed for an additional 2 days under the same conditions. Control flasks without bacterial inoculation were included to account for spontaneous dye decolorization. After the completion of the experiment, samples were collected and subjected to centrifugation at 15,000 rpm for 10 min. The absorbance of each sample was estimated spectrophotometrically (Jenway 6800 UV/Vis spectrophotometer, UK) and the produced decolorization of the test dye was calculated as a percentage of its λmax absorbance using the equation shown below (Eq. 1). AB was measured at 650 nm and MB at 665 nm while CR at 470 nm.
$$mathbf{D}mathbf{e}mathbf{c}mathbf{o}mathbf{l}mathbf{o}mathbf{r}mathbf{i}mathbf{z}mathbf{a}mathbf{t}mathbf{i}mathbf{o}mathbf{n}(mathbf{%})=frac{{varvec{A}}{varvec{b}}{varvec{s}}{varvec{o}}{varvec{r}}{varvec{b}}{varvec{a}}{varvec{n}}{varvec{c}}{varvec{e}}left({varvec{c}}{varvec{o}}{varvec{n}}{varvec{t}}{varvec{r}}{varvec{o}}{varvec{l}}right)-mathbf{A}mathbf{b}mathbf{s}mathbf{o}mathbf{r}mathbf{b}mathbf{a}mathbf{n}mathbf{c}mathbf{e}(mathbf{t}mathbf{e}mathbf{s}mathbf{t})}{{varvec{A}}{varvec{b}}{varvec{s}}{varvec{o}}{varvec{r}}{varvec{b}}{varvec{a}}{varvec{n}}{varvec{c}}{varvec{e}}({varvec{c}}{varvec{o}}{varvec{n}}{varvec{t}}{varvec{r}}{varvec{o}}{varvec{l}})}times 100$$
Additionally, the visual examination of the pellet’s color after centrifugation was checked to determine whether the dye had been adsorbed to the cells rather than being degraded. The experiment was carried out in duplicate40.
Bacterial identification by 16S rRNA
This was carried out for the bacterial isolates which exhibited the highest percentage of dye decolorization in liquid phase assay. The purified cultures of the tested isolates were sent to Biotech Serve Co. (Giza, Egypt), for DNA extraction, amplification of the 16S rRNA gene, and sequencing using the Sanger dideoxy sequencing method. The two primers used for PCR amplification of the 16S rRNA gene were universal bacterial primers 27F (5′-AGA GTT TGA TCM TGG CTC AG-3′) and 1492R (5′-TAC GGY TAC CTT GTT ACG ACT T-3′). The NCBI BLAST (BASIC LOCAL ALIGNMENT SEARCH TOOL; http://blast.ncbi.nlm.nih.gov/Blast.cgi) was used to assess DNA similarity and sequence alignment. The sequences with the highest identity score were then retrieved and aligned using multiple sequence alignment tool with the CLUSTAL W of MEGA software (Version 11.0.13). Then, the aligned sequences were used for the preparation of a phylogenetic tree using the same software (MEGA) following the maximum likelihood method41.
Estimation of extracellular and intracellular ligninolytic activity for the test isolate
The dye was incubated with either the culture supernatant or crude cell lysate of the isolate that exhibited the highest activity in the previous assay to assess extracellular and intracellular activity, respectively. The experiment was carried out by culturing the isolate in both LB broth and lignin containing medium to induce the lignin degrading/dye-decolorizing enzymes40. Two pure colonies of the bacterial isolate were used for separate inoculation of 250 mL flasks containing 25 mL of either LB broth or Mineral Salt Medium supplied with 600 mg/L KL (MSM-KL), 10 g/L glucose, and 3 g/L peptone. The MSM was composed of the following components in g/L: K2HPO4, 2.0; Na2HPO4, 2.4; MgSO4.7H2O, 0.01; CaCl2, 0.01 and a trace element solution of 1.0 mL/L36. After that, the flasks were placed in an orbital shaking incubator at 30 °C and 200 rpm for 3–4 days. At the end of the incubation period, the contents of each flask were centrifuged at 4 °C and 7000 rpm in a cooling centrifuge for separation of culture biomass from culture supernatant. The biomass was subjected to sonication for the preparation of crude cell lysate. Both cell-free culture supernatant and crude cell lysate were used for determination of extracellular and intracellular ligninolytic activities, respectively as shown below.
Testing extracellular decolorizing activity of the test isolate
The decolorizing activity was carried out using the cell-free culture supernatant42. The reaction mixture was composed of 0.5 mL of the culture supernatant, 0.5 mL of 0.160 mM AB, and 1.5 mL of 125 mM sodium tartrate buffer (pH 3.0) and the assay was carried out in test tubes. To start the reaction, 0.5 mL of 2 mM hydrogen peroxide was added to each test tube43. Static and shaking conditions were both carried out for the reaction over a period of 2 days. Control flasks were included to account for spontaneous dye decolorization. The absorbance of each sample was estimated spectrophotometrically at λmax (650 nm) and the produced decolorization activity was calculated as a percentage relative to control using Eq. (1).
Testing the intracellular decolorizing activity of the test isolate using the crude cell lysate
After harvesting the cell biomass from each flask, the cells were washed using phosphate buffered saline (PBS) and resuspended in 5 mL of PBS, sonicated by using an ultrasonic processor with an amplitude of 50% for 5–10 min, in cycles of 30 s on and 30 s off, while maintaining the sample on ice to prevent overheating. After that, the sample was centrifuged at 15,000 rpm for 5 min. The supernatant was collected, recentrifuged under the same condition and this process was repeated several times to remove any cell debris. The obtained clear supernatant representing the crude cell lysate was used for measuring the decolorization activity. The assay was carried out in test tubes and the reaction mixtures were composed of certain volumes of crude cell lysate (0.1, 0.5, 1, and 2 mL). To each reaction mixture, 0.5 mL of 0.160 mM AB was added, and the volume was completed with PBS to 2.5 mL. To start the reaction, 0.5 mL of 2 mM hydrogen peroxide was added to each reaction mixture. Static and shaking conditions were both carried out for the reaction over a period of 2 days. Control flasks were included to account for spontaneous dye decolorization. The assay was completed as mentioned earlier.
Estimation of decolorizing activity of the test isolate under growth conditions
Single colonies were used for separate inoculation of 250 mL conical flasks containing 25 mL aliquots of either LB broth or MSM-KL supplied with 10 g/L of glucose, and 3 g/L of peptone36. The flasks were placed in a shaking incubator for 3 days, and then pre-filtered sterilized AB dye was aseptically added to the flasks at final concentrations of 6 mg/L. Incubation was resumed for an additional one day. Control flasks without bacterial inoculation were included to account for spontaneous dye decolorization40. Samples were withdrawn at various time intervals (0, 4, and 24 h) following dye addition and subjected to centrifugation at 15,000 rpm for 10 min. The absorbance of each sample was estimated spectrophotometrically at λmax (650 nm) and the produced decolorization activity was calculated as a percentage relative to control using Eq. (1).
Effect of different factors on growth and decolorizing activity of the test isolate using one-factor-at-a-time approach (OFAT)
Preparation of seed and dye biodegradation flasks
Fresh seed culture (1–3 × 106 CFU/mL) was prepared for each parameter under investigation and used to inoculate dye biodegradation flasks containing 25 mL MSM-KL per 250 mL flask. In each experiment, after the predetermined incubation period, prefiltered AB dye was added at 6 mg/L and the flasks were re-incubated under the same growth conditions for 1 additional day. Samples were withdrawn at different time intervals (0, 4, and 24 h) following AB addition for determination of dye decolorization activity and microbial growth by dry cell weight (DCW) method. The decolorizing activity was performed by the quantitative assay and expressed as a percentage following the previously mentioned Eq. (1). The uninoculated medium was used as a control in all cases. All experiments were done in duplicate.
Bacterial growth estimation by DCW
To determine the DCW, 1.5 mL aliquots were collected in pre-weighed centrifuge tubes and centrifuged, then the supernatants were carefully aspirated out while the cell pellets were dried at 70 °C till constant weight. After that, each tube was weighed to determine the dry weight of the cells by subtracting the initial weight from the final weight of each centrifuge tube44,45. The DCW was calculated and expressed as g/L.
Studying the effect of nutritional and environmental factors
In the conventional scaling-up approach, various nutritional and physical parameters were optimized by maintaining all factors at a constant level in the basal medium, except the one under study. Each subsequent factor was examined after taking into account the previously optimized factor(s)46. The impact of several parameters on the decolorizing activity of the selected isolate was investigated. The factors and their ranges were tested as follows:
Inoculum size (% v/v): 1, 1.5, 2, and 2.5.
Incubation time (day): 1, 2, 3, 4, 5, and 6.
Agitation speed (RPM): 120, 160 and 200.
Temperature (ºC): 25, 30, and 35.
Carbon source (0.4% w/v): glucose, sucrose, starch, and the effect of no addition of extra carbon source was also assessed.
Lignin concentration (mg/L): 200, 400, 600, 800, and 1000.
Nitrogen source (0.4% w/v): tryptone, urea, peptone, ammonium nitrate, and yeast extract.
Initial pH of the fermentation medium: 6, 8, 10, and 12.
Optimization of dye decolorizing activity using design expert software
Response surface methodology (RSM), central composite design (CCD) was applied to optimize the environmental factors for the dye decolorization assay. Because the OFAT approach to optimization is time-consuming and tedious, RSM with CCD was selected for appropriate variation of key components. The model for decolorization of AB was implemented with the identification of likely interactions and optimal operating parameters. The effects of four independent factors, including lignin concentration (A), tryptone concentration (B), pH (C), and incubation time (D), each with three levels (low, medium, and high), on one response % decolorization of the dye as a dependent variable were examined in a total of 30 experiments. To determine the percentage of dye decolorization after four hours of dye addition, the combined impacts of four factors were examined in the designated ranges including lignin concentration (400–600 mg/L), tryptone concentration (1–7 g/L), pH (8–10), and incubation period (24–72 h).
Table 1 also displays the experimental design matrix produced from the CCD model using DESIGN EXPERT (DESIGN EXPERT Software, v. 13.0.5.0, Stat-Ease Inc., Statistics Made Easy, Minneapolis, MN, USA) which contributed to the final model equation with observed and predicted responses.
The dye biodegradation media (MSM) of the RSM model-based experiments were prepared and distributed as 25 mL aliquots in 250 mL Erlenmeyer flasks. They were inoculated at 1.5% v/v mL of the seed culture and then incubated at the previously found optimum agitation speed and temperature. After incubation, samples were collected following four hours of dye addition to be tested for dye decolorization activity of the test isolate.
Statistical and graphical investigations
To determine how well the model fits the data, analysis of variance (ANOVA) was examined which included estimation of p-values, lack of fit F-values, adjusted and predicted R2. The model reduction was also implied to improve model statistics. Only factors with p-values lower than 0.05 were considered significant. To ensure model fitness, lack of fit p-value must be greater than 0.1. To illustrate the distinct and combined effects of parameters on the response, 3D response surface and contour graphs were displayed.
Validation of the applied model data
The final model’s equation and an examination of RSM plots were used to figure out and predict the maximum response values (highest decolorization percentages). Two optimized confirmation experiments were run, and the findings were compared with the values predicted by the equation to assess the model’s accuracy. The results obtained under optimum conditions and those obtained under non-optimized conditions were also compared.
Detection of genes responsible for ligninolytic activity
Genomic DNA extraction
DNA extraction was carried out using Quick-DNA™ Bacterial Miniprep kit according to the manufacturer’s instructions.
Quality control pre-tests
The genomic extract of the test isolate was then sent to Beijing Genomics Institute (BGI) Hongkong Tech Solution NGS Lab (Tai Po, New Territories, Hong Kong) by DHL to perform the whole genome sequencing process (WGS). Upon arrival, some quality control pretests were done such as determination of sample concentration, purity, and integrity. Concentration was detected by a fluorometer (e.g., Qubit Fluorometer, Invitrogen). Sample integrity and purity were detected by agarose gel electrophoresis (concentration of agarose gel: 1% voltage: 150 V, electrophoresis time: 40 min).
Genome sequencing and assembly
The genomic DNA extract of the selected test isolate was sequenced using a DNBSEQ G400 PE150 (MGI Tech Co., Ltd). The raw reads of low quality from paired-end sequencing (those with consecutive bases covered by fewer than five reads) were discarded. The sequenced reads were then assembled using Spades v 3.15.2 software.
Bioinformatic analysis for detection of genes responsible for ligninolytic activity
The bioinformatics analysis was proceeded for annotation and detection of genes responsible for ligninolytic activity. Genome annotation is the process of identifying functional elements along the sequence of a genome. The FASTA file of clean reads after assembly was uploaded to BACTERIAL AND VIRAL BIOINFORMATICS RESOURCE CENTER BV-BRC (https://www.bv-brc.org/) for annotation of genes. The annotation service available in BV-BRC uses a modular, updated version of rapid annotation using subsystem technology (RAST)47 that is called the RAST toolkit (RASTtk)48. Genes related to lignin degradation and dye decolorization activity were identified and manually annotated by performing a BLASTP search against the ‘nr’ database.
Submission of the next generation sequencing (NGS) reads to the national center for biotechnology information (NCBI)
The FASTQ files of Next Generation Sequencing (NGS) readings were submitted to the National Center for Biotechnology Information (NCBI) to SRA database (https://submit.ncbi.nlm.nih.gov/about/sra/).
Results
Isolation of bacterial isolates with potential ligninolytic activity
A total of 177 possible ligninolytic bacteria were retrieved from 55 soil samples that were collected from different locations in Egypt. Coded numbers were assigned to soil samples and bacterial isolates (Table 2).
Assessment of the recovered isolates for their ligninolytic activities using dye decolorization assays
Solid phase dye decolorization assay
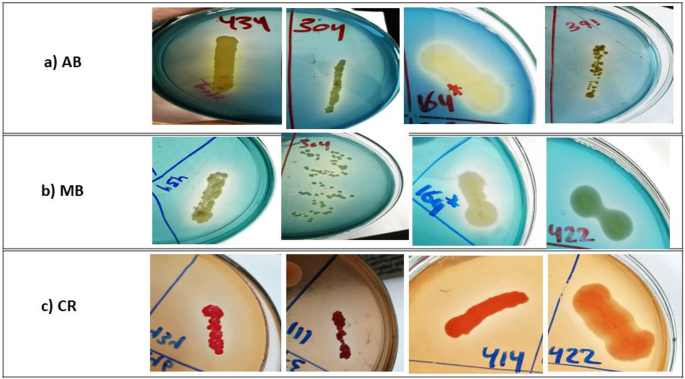
Dye decolorization assay in solid agar plates revealed that 46 (25.9%), 60 (33.8%), and 51 (28.8%) of bacterial isolates showed decolorization zones with AB, MB, and CR, respectively (Fig. 1 and Table 2). As demonstrated in Table 2, some isolates on AB and MB plates showed dye adsorption, while others did not. However, all the tested isolates in the CR assay had adsorbed the CR dye on the growth surface.

Decolorization zones of some representative isolates with potential ligninolytic activity on LB-dye agar plates. (a) AB (Azure B) for isolates 434, 304, 164, and 391; (b) MB (methylene blue) for isolates 434,304, 164, and 422; (c) CR (Congo red) for isolates 434, 111, 414, and 422.
Liquid phase dye decolorization assay
Liquid phase assays were carried out for a total of 16 bacterial isolates which gave positive results on the three different types of dyes in solid phase assay (Table 3). The liquid assay of AB showed that isolates 434 and 304 have the highest decolorization capacity with percentages of 98.5 and 72.5%, respectively, and of 86.3 and 81%, respectively in MB assay after 2 days of incubation with the dye. However, in the CR assay, isolate 391 showed the highest decolorization activity by 29.2% (Fig. 2). Dye adsorption on the pellets’ surface after centrifugation was also monitored and the results are shown in (Fig. 2). Similar to the results obtained in solid phase dye decolorization assay, all the tested isolates in CR assay showed adsorbed dye on the pellets’ surface.

Decolorization percentages produced by the tested isolates in LB broth containing Azure B (AB), methylene blue (MB), or Congo red (CR) as determined in liquid phase assay. (a) Isolates showed adsorption of dye on pellets’ surface after centrifugation.
Molecular identification of bacterial isolates
The 16S rRNA gene was sequenced to identify two selected isolates (434 and 304) that had the highest decolorization activity for the tested dyes. Isolate 434 could be identified as Streptomyces intermedius since it showed 99.86% homology of the 16S rRNA nucleotide sequences with Streptomyces intermedius strain DSM 40372 (NR_119347.1). While isolate 304 could be identified as Streptomyces griseorubens with 99.93% homology of the 16S rRNA nucleotide sequence of Streptomyces griseorubens strain NBRC 12780 (NR_041066.1). The partial nucleotide sequences of 16S rRNA genes of the two isolates (434 and 304) were submitted and deposited in the NCBI GenBank database under the accession numbers OQ928560 and OQ928561, respectively. According to the phylogenetic tree (Fig. 3), S. intermedius and S. griseorubens are the closest strains in similarity to the tested isolates, 434 and 304, respectively.

The phylogenetic tree of isolates 434, and 304 based on 16S rRNA genes sequence.
Extracellular and intracellular ligninolytic activity for the test isolate
Extracellular decolorizing activity of the test isolate
None of the culture supernatants from either LB or lignin containing medium grown culture exhibited significant dye-decolorizing ability (not more than 10% decolorization could be detected) for either static or shaking assay conditions after 2 days of incubation with the dye.
Intracellular decolorizing activity of the test isolate using the crude cell lysate
The results for the crude cell lysate obtained from LB culture showed no decolorizing activities after 2 days of incubation with the dye, while the corresponding results of lignin containing medium’s culture showed slightly decolorizing activities after 2 days of incubation with the dye (less than 5% decolorization).
Decolorizing activity of the test isolate under growth conditions
After 72 h growth age of S. intermedius isolate, the addition of AB dye resulted in decolorization capacity in LB medium of 0.002, 0.06, and 30% and in MSM-KL of 6.8, 21, and 41% following dye addition and re-incubation under the same growth conditions for 0, 4, and 24 h, respectively.
Effect of different factors on growth and decolorizing activity of the test isolate using one-factor-at-a-time approach (OFAT)
Effect of different inoculum sizes
The result of the decolorization assay at 4 and 24 h following the addition of dye to 72 h age culture and re-incubation under the same growth conditions showed that the highest decolorization capacity was 22.69 and 39.18% upon using 1.5% v/v inoculum size, respectively (Fig. 4A).

Effect of different inoculum sizes (A) and different incubation times (B) on decolorization percentage of AB after 0, 4, and 24 h of dye addition to the grown culture of S. intermedius test isolate. Lines above columns indicate standard deviation values.
Effect of incubation time
Two days of incubation growth period produced the maximum decolorization activity, which was 24.7 and 44.92% after 4 and 24 h following the addition of dye and re-incubation under the same growth conditions, respectively (Fig. 4B).
Effects of agitation speed and incubation temperature
As demonstrated in Fig. 5A,B, the maximum decolorization activity was obtained at 160 rpm and 30 ºC which was 33.58 and 47.18% at 4 and 24 h following dye addition to 2 days age culture and re-incubation under the same conditions, respectively.

Effect of agitation speed (A) and different temperatures (B) on decolorization percentage of AB after 0, 4, and 24 h of dye addition to the grown culture of S. intermedius test isolate. Lines above columns indicate standard deviation values.
Effect of carbon source
The maximum activity for dye decolorization was observed by no addition of extra carbon source to MSM-KL. These maximum activities were 25.65 and 50.84% at 4 and 24 h following the addition of dye to 2 days age culture and re-incubation under the same growth conditions, respectively (Fig. 6A).

Effect of carbon source (A) and lignin concentrations (B) on decolorization percentage of AB after 0, 4, and 24 h of dye addition to the grown culture of S. intermedius test isolate. Lines above columns indicate standard deviation values.
Effect of lignin concentration
The ability to decolorize AB by 19.07 and 54.92% at 4 and 24 h following dye addition, respectively, was demonstrated by a lignin concentration of 600 mg/L (Fig. 6B).
Effect of nitrogen source
The highest activity was obtained when using tryptone as a nitrogen source by 38.69, and 55.17% followed by peptone and yeast extract as shown in Fig. 7A at predetermined assay time measurements.

Effect of some nitrogen sources (P: Peptone, T: Tryptone, YE: Yeast extract, AN: Ammonium nitrate, and U: Urea) (A) and different pH (B) on decolorization percentage of AB after 0, 4, and 24 h of dye addition to the grown culture of S. intermedius test isolate. Lines above columns indicate standard deviation values.
Effect of initial pH
After 4 and 24 h of dye addition, pH 8 and 10 showed close decolorization activity by 34.27, 56.59, and by 34.92, 56.76%, respectively (Fig. 7B).
Statistical optimization of process parameters, analysis, and validation
RSM (CCD) was used to perform comprehensive research on dye decolorization by the selected S. intermedius test isolate. The design included 16 factorial points, 8 axial points, and 6 repetitions at the center point for the estimation of the pure error sum of squares. Analysis of variance (ANOVA) for percentage decolorization given in Table 4 revealed that the quadratic model is highly statistically significant, as indicated by the model F value of 24.64 which means that there is only a 0.01% chance that the value is significant by chance. the p- value is a measure of a test’s statistical significance; a value below 0.05 denotes a test parameter’s significance at the 5% level, and multiple model variables were significant (p < 0.05). The model’s effectiveness and capacity to predict the percentage of dye decolorization were demonstrated by the similarity of adjusted R2 (0.8874) and predicted R2 (0.7641) and the difference between them didn’t exceed 0.2. The adequate precision of the model, which assesses signal to noise ratio, was 14.63, with a ratio greater than 4 is desirable and after excluding the non-significant factors (Table 5), the best mathematical model explaining the behavior of the process can be reduced to that presented in Eq. (2).
$$begin{gathered} {text{Decolorization of dye }}left( % right) = – {73}.{6967} + 0.0{289958}*{text{lignin conc}} + {4}.00{44}*{text{tryptone conc}} + hfill \ {6}.{5}0{475}*{text{pH}} + {3}.{49779}*{text{incubation time}} + 0.00{237854}*{text{lignin conc}}*{text{tryptone conc}} + hfill \ 0.0{6965}0{2}*{text{tryptone conc}}*{text{incubation time}} + left( { – 0.{121549}*{text{pH}}*{text{incubation time}}} right) + ( – 0.{738434} hfill \ *{text{tryptone conc}}^{2} ) + left( { – 0.0{343354}*{text{incubation time}}^{2} } right) hfill \ end{gathered}$$
Consequently, by resolving this equation, the optimum levels of lignin conc, tryptone conc, pH, and incubation time were 583 mg/L, 5.4 g/L, 9.8, and 40.6 h, respectively, with a predicted decolorization percentage of 66.85%.
The design-expert program was used to display a contour and response surface graph to investigate the relationship between the independent factors and the response. Figure 8a–c depicts these graphs, which demonstrate the quadratic effect of tryptone concentration and incubation time on the response. This effect can be explained by the statistical significance of the quadratic coefficients of these variables in the model with a convex shape in both axes (Fig. 8b). However, plots of pH and lignin content show a linear effect as seen in (Fig. 8a,c).

Three-dimensional (3D) surface plots for the effects of initial pH, lignin conc, tryptone conc and incubation time (growth age before dye addition) on AB decolorization percentage as obtained from design expert software.
A statistically improved medium was used in two trials to confirm the validity of the experimental model. The response was 59.75 ± 6.94%, which indicated a strong resemblance between actual (59.75 ± 6.94%) and predicted values (66.85%) and confirming the accuracy and reliability of the model.
Dye decolorization in the basal and optimized conditions
Decolorization of AB dye (starting concentration of 6 mg/L) was accomplished using dye decolorization flasks from basal and optimized medium. Only 22.69% of the AB was decolorized from the basal medium after 4 h and 39.18% after 24 h contact periods after dye addition and re-incubation under the same conditions. While, by utilizing an optimized medium, it was observed that the corresponding decolorization percentage of the AB dye was increased to 59.75 and 80.36% after the same contact periods mentioned before (Fig. 9).

Decolorization percentage of AB in the basal and optimized conditions after 4 and 24 h of dye addition to the grown culture of S. intermedius test isolate. Lines above columns indicate standard deviation values.
Detection of the genes responsible for ligninolytic activity
Genomic DNA extraction of S. intermedius test isolate and quality control pretests
The DNA extraction from S. intermedius test isolate was performed, and the quality control pretests yielded the following outcomes, as presented in (Table 6 and Fig. S1). These outcomes indicate that the DNA extraction was successful, with a sample concentration of 37.2 ng/µL. Additionally, the sample integrity test indicated slight degradation. The overall assessment categorizes the sample as level A, signifying that it meets the necessary criteria for library construction and sequencing. The success rate of the sequencing process was around 93.59%.
Genome sequencing and assembly
In sequencing data, there is a certain number of low-quality reads in the raw data obtained. In order to obtain more accurate and reliable results in subsequent bioinformatics analysis, the raw data was treated by removing reads with a certain proportion of low quality, removing adapter contamination, and removing duplication contamination. The Q20%, Q30%, and GC% values were assessed for both pre-processed and post-processed data. Q20% increased slightly from 97.1 to 97.26%, while Q30% improved from 92.59 to 93.49%. The GC content remained relatively stable, changing from 72.44 to 72.1%. The above processes were applied to read 1 (forward read along the forward DNA strand), and read 2 (reverse read along the reverse DNA strand) of the whole genome sequence data obtained for S. intermedius test isolate and the statistical results are shown in (Table 7).
Bioinformatic analysis of whole genome sequence data of S. intermedius test isolate
The whole genome sequence obtained for S. intermedius test isolate after assembly was 7,084,373 bp in size, with 73.07% GC content, 6672 coding sequences (CDS) (Table 8), 4219 proteins with functional assignments, and 2453 hypothetical proteins (Table 9). According to morphological characteristics, 16S rRNA sequence, and complete genome sequence, this isolate was classified as a gram-positive bacterium, Streptomyces sp.
Genome sequence analysis of S. intermedius test isolate identified potential genes that may be accountable for degrading or depolymerizing lignin (Supplementary Materials, Table S1). After functional annotation of the genome sequence, the analysis showed the presence of bacterial peroxidases, especially dye decolorizing peroxidase (DyP) type. The genomic sequence of S. intermedius test isolate also contained a number of oxidoreductases, oxidases, reductases, and multicopper oxidases (laccases). Considering this, DyP and the laccase, which may be responsible for the enzymatic activity seen in this study may be brought on by the existence of these genes. Besides, it has been observed the presence of several genes encoding proteins that may be associated with the protection of cells against reactive species generated during aromatic compound metabolism. This includes superoxide dismutase, glutathione peroxidase, and thioredoxin genes. Additionally, it was found in the genome of this test organism genes related to the class of heme-thiolate proteins known as the cytochrome P450 genes.
Moreover, genome sequence analysis of S. intermedius test isolate identified the presence of multiple genes that are responsible for the catabolism of lignin derived compounds (Table S2). This includes the presence of multiple genes encoding enzymes responsible for aromatic funneling pathways such as hydroxybenzoate hydroxylase enzyme, vanillate demethylase, and dehydrogenases.
Additionally, the genes encoding enzymes involved in the metabolism of key aromatic intermediates through the ring cleavage pathway were discovered. These enzymes include catechol 1,2-dioxygenase, homogentisate 1,2-dioxygenase, 4-hydroxyphenylpyruvate dioxygenase, extradiol dioxygenase and multiple monooxygenases encoding genes (1,2-phenylacetyl-CoA epoxidase). Several transferase and hydratase encoding genes, including acetyl-CoA acetyltransferase and enoyl-CoA hydratase, which were previously reported to play a role in the breakdown of the aromatic molecule benzoate were also found. Additionally, the circular genome map that showed the distribution of some of the genes encoding ligninolytic enzymes was constructed using Proksee49 (Fig. S2).
As shown in Tables S1 & S2, the amino acid sequences of the S. intermedius test isolate’ genes that encode the lignin-degrading products were compared to other amino acid sequences that had been deposited in the NCBI database by using BLASTP algorithm to detect the similarity between them. By using this approach, we were able to locate the expected genes responsible for all of the pathways leading to lignin breakdown.
Submission of sequencing reads to NCBI
The FASTQ files of the WGS project of S. intermedius test isolate have been deposited in the NCBI/SRA database under the accession code SRR25321249.
Discussion
It is approved that ligninolytic enzymes have a wide range of industrial uses. Besides, the production of numerous fine compounds from lignin and detoxification of pulp produced from paper industry processes50, they also have applications in versatile pollutant degradation such as insecticides, and polycyclic aromatic hydrocarbons (PAHs). They are helpful in cleaning up the environment since they may transform toxic substances into less dangerous forms11. Moreover, the ligninolytic enzymes produced by different microorganisms can also decolorize different types of dyes25,51. Those findings hold significant relevance in applying these enzymes in decolorizing textile dye effluent and bioremediation processes21. In this study, we could retrieve 177 potential ligninolytic bacteria from 55 soil samples using minimal salt medium containing KL. The use of lignin as the only source of carbon in minimal salt medium could encourage bacterial growth and raise the possibility that isolates may depolymerize lignin52. As mentioned in the literature, textile dye decolorization is one of the most significant uses of ligninolytic enzymes. Laccases and peroxidases, two types of ligninolytic enzymes, have been used to remove synthetic colors from textile effluent. By using them in textile processing, conventional treatments may be replaced with more environmentally friendly ones11,53,54. Therefore, assessing the isolates’ ligninolytic potential could be carried out through testing their capability to decolorize various dyes. Three different types of dyes were used in this study which included AB, MB, and CR. This method revealed that the bacterial isolate which demonstrated a unique capacity to decolorize distinct types of dyes with or without adsorption of the dye on its growth surface could have potential ligninolytic activity. Out of the total isolates tested, 16 of them displayed decolorization zones with the three different types of dyes used, indicating that they produce oxidative enzymes such as peroxidases and laccases which have the potential to act synergistically to depolymerize lignin. These isolates were subsequently selected for liquid phase testing to further explore isolates with the highest activity for potential application in lignin degradation and also for textile dye effluent bioremediation21. All tested isolates with decolorization activity on either tested dye displayed dye adsorption on cell pellets in the CR assay. Consequently, it is not clear to what extent isolates can degrade the dye rather than being adsorbed. Isolates having the code numbers 434 and 304 exhibited the highest activity by 98.5 and 72.5% in the AB assay and by 86.3% and 81% in the MB assay, respectively without adsorption of dye on cell pellets indicating that, among all the tested isolates, they have the strongest decolorizing activity. Based on the 16S rRNA sequence, isolates 434 and 304 are closely related to different Streptomyces groups, showing the highest similarity to S. intermedius strain DSM 40372 (NR_119347.1) and S. griseorubens strain NBRC 12780 (NR 041066.1), respectively. From the phylogenetic tree (Fig. 3), S. rutgersensis (NR_112495)55, S. griseoflavus (NR_042291), and S. griseorubens (NR_041066)51, S. althioticus (NR_115392)56 as well as S. variabilis (NR_043840)57 strains which are closely related to the two isolates used in this study, were also previously reported to demonstrate lignin-degrading ability. Dye decolorizing capacity in MSM-KL was greater than in LB medium indicating that lignin containing medium can induce the microorganism to produce ligninolytic enzymes within a shorter period of time. Several experiments have been carried out to determine factors affecting dye decolorizing activity by S. intermedius test isolate. The findings in this study showed that the activity was less affected by inoculum sizes (Fig. 4A). However, the maximum activity was observed within 48 h, and these results supported past research showing that actinomycetes exclusively generate ligninolytic enzymes as a growth-associated primary metabolic activity58,59. In this study, the decolorizing activity by S. intermedius isolate was maximum at 160 rpm. The same rpm was used for ligninolytic enzymes production by Streptomyces sp.S631 compared to lower agitation (120 rpm)60,61 and higher agitation (200 rpm)62 used for such types of enzymes’ production. However, in this study, the increase in bacterial growth by agitation rates of 200 rpm, did not boost decolorization activity and consequently related degrading enzymes’ production. This could be interpreted on the basis that a high agitation rate increases the metabolic status of the microbial culture that might be balanced by a reflecting action demonstrated by enzymes synthesis lowering effect as a protective mechanism against cell lysis. Moreover, at lower agitation rates, enzyme output is reduced considerably which is probably attributed to a decrease in biosynthetic capability of the cells as a result of inadequate oxygenation and mixing of the culture media. The rate of agitation has a significant impact on how much dissolved oxygen is present in the fermentation medium62.
Additionally, it was observed that the optimum temperature was 30 ºC, a similar pattern of findings was reported in the biodegradation of KL by Bacillus velezensis strain36. Falade and his colleagues, also found that maximum LiP activity was obtained at a temperature of 30 °C and incubation time of 48 h63, which is likewise in good agreement with the optimum temperature (30 °C) and incubation time (48 h) reported in the present work. Most research articles on the biodegradation of lignin consistently report that the optimal temperature is 30 ºC64.
Some articles reported that ligninolytic bacteria can use KL as their only source of carbon without addition of extra carbon sources known as co-substrates52,65, while others reported that bacteria on fermentation media with glucose or the addition of another carbon source to the medium as a co-substrate can increase the production of ligninolytic enzymes36,66 or even is a must for stimulation of bacterial growth and co-metabolism of KL67. So, in this study, the effect of the addition extra carbon sources to MSM-KL was tested. The maximum activity was shown by no addition of extra carbon source to MSM-KL and thus validated that the isolate could utilize lignin as the only carbon source as a result of the lignin-inducing effect that drives the microorganism to manufacture lignin-degrading enzymes. The S. intermedius isolate was anticipated to depolymerize lignin and absorb the lower molecular weight KL-derived molecules to thrive31.
Based on the findings from previous studies, it has been observed that certain microorganisms exhibit limited tolerance to high concentrations of KL due to the tendency of KL to inhibit the growth of bacteria at higher concentrations67,68. To assess the tolerance effect of the isolate, varying concentrations of lignin were employed for testing this effect, the results revealed that S. intermedius isolate can tolerate a high concentration of lignin (1000 mg/L) with an optimum activity by using KL at a concentration of 600 mg/L which is higher than that found in pulp paper mill effluent (500 mg/L)67.
Enzyme production is influenced by the availability of nitrogen and its concentration in the media. The type of nitrogen supply may also affect the production and decolorization activity of ligninolytic enzymes69. The dye decolorization flasks were supplemented with several inorganic and organic nitrogen sources separately in an effort to identify an optimal nitrogen source for the decolorization activity. Regarding microbial growth, it was observed that some nitrogen sources had a significant effect on bacterial growth. The highest levels of decolorizing ability were observed when tryptone was used as a nitrogen source at a concentration of 0.4% w/v followed by peptone and yeast extract when used at the same concentration. On the contrary, urea and ammonium nitrate reduce microorganisms’ capacity to proliferate. DCW was significantly decreased when using urea and ammonium nitrate as a nitrogen source in comparison to other nitrogen sources used in this study, which may be the most likely reason for the decreased activity. This result ties well with previous studies wherein demonstrated that tryptone stimulates the highest laccase synthesis in comparison to yeast extract in Bacillus sp.70. Others found yeast extract as the optimum nitrogen source for the laccase production for the strain S. psammoticus59,62, while, Olajuyigbe and his colleagues69 reported that ammonium nitrate is the optimal nitrogen source for Stenotrophomonas sp for the production of ligninolytic enzymes.
The effect of initial pH on decolorization activity by S. intermedius test isolate was assessed by preparing fermentation media at different initial pH (6–12). The ranges of pH were selected according to previous studies of isolated ligninolytic bacteria36. Results obtained from this study showed that the optimum AB decolorization percentage from S. intermedius test isolate was achieved at pH 8.0 and 10. However, the decreased microbial growth and thereby enzymatic production at pH 12. These results revealed that pH is an important factor in decolorization activity. The results obtained in this study in agreement with previous studies reported that maximum ligninolytic enzyme production from Bacillus sp. SHC1 and Leucobacter sp.SHC3 occurred at pH 868. It was also reported that Cupriavidus basilensis B-8, a bacterium with ligninolytic activity can grow at an alkaline pH range with optimum activity of KL degradation at pH 764. However, most fungal isolate reports on the generation of ligninolytic enzymes indicated that production was best at an acidic pH69,71.
The level of the chosen variables was then optimized using RSM (CCD). During the evaluation of 3D response surface plots, it has been observed that tryptone and incubation time have an interactive effect on the percentage of dye decolorization with a convex shape in both axes (Fig. 8b) which means that the activity first increases gradually as tryptone and incubation time are increased to their peak levels, then decreases as tryptone levels are raised and incubation times are lengthened. The corresponding response surface plot’s convex form (Fig. 8b) suggested that the response’s optimal value falls within the range of the factors studied, a similar conclusion was reached by Aghaie-Khouzani72, who found that the maximum laccase activity falls within the optimum level of peptone concentration (2.2 g/L) upon using RSM. An alkaline pH range between 8 and 10 may boost the activity of ligninolytic enzyme production. These results go beyond previous reports, showing the same findings36,63. Consequently, when relevant components were used at their optimum levels, a 2.6-fold increase in the dye’s decolorization percentage was attained after 4 h of incubation with bacterium growth of S. intermedius isolate using the model’s optimal settings. These findings showed that the time of decolorization decreased, and the dye disappearance percentage increased, compared to unoptimized conditions.
The lignin’s intricate structure necessitates the cooperative action of several lignin-degrading enzymes that work in harmony. By applying WGS and annotation of genes, we were able to find and locate the anticipated genes in charge of all the processes leading to lignin degradation (Supplementary Materials, Tables S1, S2/Fig. S2). Two mechanisms are primarily responsible for lignin breakdown in nature: lignin is firstly depolymerized to produce low molecular weight lignin fragments, which are then either modified or biologically converted into other compounds. The depolymerization process of native lignin is induced by oxidative enzymes, such as peroxidases and laccases which finally leads to an oxidative degradation of bonds in lignin (Fig. S3A)1. The genomic sequence of S. intermedius isolate revealed the existence of multiple peroxidases and laccase genes. Peroxidase enzymes have been found to be the primary enzymes involved in lignin depolymerization73. DyP-type is a newly discovered family of peroxidases that has been reported to have broad substrate specificity including several types of dyes and lignin compounds74,75. Multicopper oxidase or laccase acts by reducing oxygen to water resulting in oxidation of phenolic and nonphenolic substances to their corresponding radical species which can then be subjected to further reactions51,76. It has been reported in several studies that the laccase enzyme produced by several Streptomyces sp can decolorize or degrade aromatic dyes77. So, the dye decolorizing activity observed in this study may be due to the presence of DyP and laccase encoding genes. As supporting enzymes, the oxidases produce hydrogen peroxides that the peroxidases use to break down lignin and aromatic compounds78,79. The oxidoreductase’s function is to produce unspecific free radicals and reactive intermediates in order to break down lignin and aromatic compounds80. These intermediates may be toxic and affect cell viability. Therefore, they must be eliminated or changed into safer and more stable molecules to allow microbial cell survival. Numerous ubiquinone oxidoreductase-encoding genes were found in S. intermedius isolate, which were tied with previous work by Jindal Li and his colleagues81 who reported the lignin degrading ability of this protein in Aspergillus fumigatus. Therefore, these genes may also be a sign of S. intermedius isolate’s capacity to degrade lignin. Several proteins, such as catalase, superoxide dismutase, glutathione peroxidase, and thioredoxin, associated with the oxidative stress response for defense against reactive species and detoxification pathways during aromatic metabolism, were also discovered in the genomic sequence of S. intermedius isolate. Genes encoded with cytochrome P450 proteins had also been found and were previously reported to assist lignin breakdown. Cytochrome P450 catalyzes many enzymatic reactions for the conversion of aromatic/xenobiotic compounds into more polar and/or less toxic derivatives82. When lignin is successfully depolymerized, a variety of substituted phenols and propyl phenols, as well as oligomers of these, are produced (Fig. S3A)50. After some modifications, those tiny fragments may be interesting chemicals on their own and represent one method of lignin valorization. The presence of several dehydrogenase encoding genes in the genomic sequence of S. intermedius isolate indicated that the isolate also had a role in the aromatic funneling pathway (Fig. S3B). Dehydrogenases act by transforming the various aldehydes produced into the equivalent carboxylic acid, which is typically less toxic to the host cell1. Different pathways were associated with the aromatic funneling process to produce central intermediate metabolites. The most common intermediate metabolite is protocatechuate. It has also been observed that the presence of two genes encoding enzymes of hydroxybenzoate hydroxylase and vanillate demethylase which are previously reported to have significant roles in the aromatic funneling process. Hydroxybenzoate hydroxylase acts by converting hydroxybenzoate into protocatechuate, vanillate demethylase acts by converting vanillic acid into protocatechuate83. This may ensure the potential role of this strain in the aromatic funneling pathways. The aromatic funneling pathways also produce some common central intermediates including catechol, protocatechuate, gentisate, and homogentisate, which will then go through central-ring cleavage catalyzed by ring-cleaving dioxygenase (Fig. S3B)2. Thus, the genomic analysis of S. intermedius isolate revealed the presence of another group of genes encoding enzymes that are responsible for romantic ring cleavages such as catechol dioxygenase, homogentisate dioxygenase, hydroxyphenylpyruvate dioxygenase, and monooxygenases, which contribute to the breakdown of central intermediate pathways. Some transferase and hydratase encoding genes were also detected in the genomic sequence of S. intermedius isolate. Those enzymes were also found to play a part in the breakdown of the aromatic molecule benzoate84. Finally, the analysis and identification of different metabolic pathways in its genomic sequence related to lignin degradation and dye decolorization activities indicated that we could consider this strain as a potential candidate for industrial application in lignin biodegradation and textile dye effluent bioremediation.
Conclusion
The soil serves and can be considered as a reservoir for undiscovered or uncharacterized bacterial isolates that may be more beneficial in environmentally friendly processes. Upon isolation and characterization of bacterial isolates from various soil samples, S. intermedius test isolate 434 is beneficial as it used KL as a sole carbon source for growth and showed considerable decolorization efficiency of AB and other types of dyes. OFAT approach And RSM were used to maximize ligninolytic enzyme production and dye decolorizing capacity over a shorter period (4 h). This resulted in a 2.6-fold increase in the decolorizing capacity of AB within 4 h incubation with bacterium growth of the isolate. This study presents a novel approach to accelerating dye decolorization by optimizing ligninolytic enzyme production, offering a more efficient biotechnological solution.
Unlike previous studies, our work integrates optimization of dye decolorization with WGS, providing both functional and genetic insights into ligninolytic enzymes efficiency for industrial applications.
Data availability
All data generated or analyzed during this study are included in the manuscript and supplementary information files.
Abbreviations
- KL:
-
Kraft lignin
- MSM-KL:
-
Mineral salt medium containing kraft lignin
- LB:
-
Luria–Bertani
- AB:
-
Azure B
- MB:
-
Methylene blue
- CR:
-
Congo red
- DCW:
-
Dry cell weight
- WGS:
-
Whole genome sequencing
- LiP:
-
Lignin peroxidase
- MnP:
-
Manganese peroxidase
- VP:
-
Versatile peroxidase
- DyP:
-
Dye decolorizing peroxidase
- OFAT:
-
One-factor-At-a-time approach
- RSM:
-
Response surface methodology
- CCD:
-
Central composite design
References
-
Abdelaziz, O. Y. et al. Biological valorization of low molecular weight lignin. Biotechnol. Adv. 34, 1318–1346 (2016).
-
Becker, J. & Wittmann, C. A field of dreams: Lignin valorization into chemicals, materials, fuels, and health-care products. Biotechnol. Adv. 37, 107360 (2019).
-
Mathew, A. K., Abraham, A., Mallapureddy, K. K. & Sukumaran, R. K. Chapter 9—Lignocellulosic biorefinery wastes, or resources? In Waste Biorefinery (eds Bhaskar, T. et al.) (Elsevier, 2018).
-
Kang, S., Li, X., Fan, J. & Chang, J. Hydrothermal conversion of lignin: A review. Renew. Sustain. Energy Rev. 27, 546–558 (2013).
-
Cheah, W. Y. et al. Pretreatment methods for lignocellulosic biofuels production: Current advances, challenges and future prospects. Biofuel Res. J. 7, 1115–1127 (2020).
-
Dharmaraja, J. et al. Lignocellulosic biomass conversion via greener pretreatment methods towards biorefinery applications. Bioresour. Technol. 369, 128328 (2023).
-
Chauhan, P. S. Role of various bacterial enzymes in complete depolymerization of lignin: A review. Biocatal. Agric. Biotechnol. 23, 101498 (2020).
-
Shi, X. et al. An acid-stable bacterial laccase identified from the endophyte Pantoea ananatis Sd-1 genome exhibiting lignin degradation and dye decolorization abilities. Biotechnol. Lett. 37, 2279–2288 (2015).
-
Takahashi, K. et al. Characterization of the catabolic pathway for a phenylcoumaran-type lignin-derived biaryl in Sphingobium sp. strain SYK-6. Biodegradation 25, 735–745 (2014).
-
Datta, R. et al. Enzymatic degradation of lignin in soil: A review. Sustainability 9, 1163 (2017).
-
Kumar, A. & Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 6, e03170 (2020).
-
de Gonzalo, G., Colpa, D. I., Habib, M. H. M. & Fraaije, M. W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 236, 110–119 (2016).
-
Sondhi, S. et al. An extracellular thermo-alkali-stable laccase from Bacillus tequilensis SN4 with a potential to biobleach softwood pulp. 3 Biotech 5, 175–185 (2015).
-
Haq, I., Kumar, S., Kumari, V., Kumar, S. & Raj, A. Evaluation of bioremediation potentiality of ligninolytic Serratia liquefaciens for detoxification of pulp and paper mill effluent. J. Hazard. Mater. 305, 190–199 (2016).
-
Gu, J. et al. Bacterial transformation of lignin: key enzymes and high-value products. Biotechnol. Biofuels Bioprod. 17, 1–34 (2024).
-
Rahmanpour, R. & Bugg, T. D. H. Characterisation of dyp-type peroxidases from Pseudomonas fluorescens Pf-5: Oxidation of Mn(II) and polymeric lignin by Dyp1B. Arch. Biochem. Biophys. 574, 93–98 (2015).
-
Yadav, M. & Yadav, H. S. Applications of ligninolytic enzymes to pollutants, wastewater, dyes, soil, coal, paper and polymers. Environ. Chem. Lett. 13, 309–318 (2015).
-
Malherbe, S. & Cloete, T. E. Lignocellulose biodegradation: Fundamentals and applications. Rev. Environ. Sci. Biotechnol. 1, 105–114 (2002).
-
Saratale, R. G., Saratale, G. D., Chang, J. S. & Govindwar, S. P. Bacterial decolorization and degradation of azo dyes: A review. J. Taiwan Inst. Chem. Eng. 42, 138–157 (2011).
-
Aragaw, T. A. A review of dye biodegradation in textile wastewater, challenges due to wastewater characteristics, and the potential of alkaliphiles. J. Hazard. Mater. Adv. 16, 100493 (2024).
-
Rahayu, F. et al. Newly isolated ligninolytic bacteria and its applications for multiple dye degradation. Water. Air. Soil Pollut. 234, 1–11 (2023).
-
Pearce, C. I., Lloyd, J. R. & Guthrie, J. T. The removal of colour from textile wastewater using whole bacterial cells: a review. Dye. Pigment. 58, 179–196 (2003).
-
Singh, R. L., Singh, P. K. & Singh, R. P. Enzymatic decolorization and degradation of azo dyes—A review. Int. Biodeterior. Biodegradat. 104, 21–31 (2015).
-
Harish, B. S., Thayumanavan, T., Nambukrishnan, V. & Sakthishobana, K. Heterogeneous biocatalytic system for effective decolorization of textile dye effluent. 3 Biotech 13, 165 (2023).
-
Wu, K. et al. Decolourization and biodegradation of methylene blue dye by a ligninolytic enzyme-producing Bacillus thuringiensis: Degradation products and pathway. Enzyme Microb. Technol. 156, 109999 (2022).
-
El Awady, M. E. et al. Exploring the decolorization efficiency and biodegradation mechanisms of different functional textile azo dyes by Streptomyces albidoflavus 3MGH. BMC Microbiol. 24, 1–22 (2024).
-
Haq, I. & Raj, M. A. Biodegradation of Azure-B dye by Serratia liquefaciens and its validation by phytotoxicity genotoxicity and cytotoxicity studies. Chemosphere 196, 58–68 (2018).
-
Li, H., Zhang, R., Tang, L., Zhang, J. & Mao, Z. Evaluation of Bacillus sp. MZS10 for decolorizing Azure B dye and its decolorization mechanism. J. Environ. Sci. 26, 1125–1134 (2014).
-
Zammuto, V. et al. Enhancement of biodegradation and detoxification of methylene blue by preformed biofilm of thermophilic bacilli on polypropylene perforated balls. J. Mar. Sci. Eng. 12, 1–16 (2024).
-
Eid, H. A., Mohamed, W. S., Zaki, A. G., Amer, S. K. & El-Shatoury, E. H. Statistical optimization of Congo red biodegradation by a bacterial strain of Alcaligenes faecalis. Bioresour. Technol. Rep. 23, 101573 (2023).
-
Riyadi, F. A. et al. Enzymatic and genetic characterization of lignin depolymerization by Streptomyces sp. S6 isolated from a tropical environment. Sci. Rep. https://doi.org/10.1038/s41598-020-64817-4 (2020).
-
Zhong, H. et al. Whole-genome sequencing reveals lignin-degrading capacity of a ligninolytic bacterium (Bacillus cereus) from Buffalo (Bubalus bubalis) Rumen. Genes (Basel) 13, 842 (2022).
-
Nawaz, M. Z. et al. Genomic insights into the metabolic potential of a novel lignin-degrading and polyhydroxyalkanoates producing bacterium Pseudomonas sp. Hu109A. Chemosphere 310, 136754 (2023).
-
Melo-Nascimento do, A. O. S. et al. Functional characterization of ligninolytic Klebsiella spp. strains associated with soil and freshwater. Arch. Microbiol. 200, 1267–1278 (2018).
-
Bera, S. P., Shah, M. P. & Godhaniya, M. Microbial remediation of textile dye acid orange by a novel bacterial consortium SPB92. Front. Environ. Sci. 10, 1–8 (2022).
-
Verma, M., Ekka, A., Mohapatra, T. & Ghosh, P. Journal of environmental chemical engineering optimization of kraft lignin decolorization and degradation by bacterial strain Bacillus velezensis using response surface methodology. J. Environ. Chem. Eng. 8, 104270 (2020).
-
Falade, A. O., Eyisi, O. A. L., Mabinya, L. V., Nwodo, U. U. & Okoh, A. I. Peroxidase production and ligninolytic potentials of fresh water bacteria Raoultella ornithinolytica and Ensifer adhaerens. Biotechnol. Reports 16, 12–17 (2017).
-
Sanders, E. R. Aseptic laboratory techniques: Plating methods. J. Vis. Exp. https://doi.org/10.3791/3064 (2012).
-
Hooda, R., Bhardwaj, N. K. & Singh, P. Screening and identification of ligninolytic bacteria for the treatment of pulp and paper mill effluent. Water Air Soil Pollut. https://doi.org/10.1007/s11270-015-2535-y (2015).
-
Bandounas, L., Wierckx, N. J. P., de Winde, J. H. & Ruijssenaars, H. J. Isolation and characterization of novel bacterial strains exhibiting ligninolytic potential. BMC Biotechnol. 11, 94 (2011).
-
Tamura, K., Stecher, G. & Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027 (2021).
-
Bholay, A. D. et al. Bacterial lignin peroxidase: A tool for biobleaching and biodegradation of industrial effluents. Univers. J. Environ. Res. Technol. 2, 58–64 (2012).
-
Arora, D. S. & Gill, P. K. Comparison of two assay procedures for lignin peroxidase. Enzyme Microb. Technol. 28, 602–605 (2001).
-
Stone, K. M., Roche, F. W., Thornhiu, N. F., Place, T. & Wcie, L. Dry weight measurement of microbial biomass and measurement variability analysis. Advanced centre for biochemical engineering, University College London. Biotechnol. Tech. 6, 207–212 (1992).
-
Glazyrina, J. et al. High cell density cultivation and recombinant protein production with Escherichia coli in a rocking-motion-type bioreactor. Microb. Cell Fact. 9, 1–11 (2010).
-
Singh, S. K., Singh, S. K., Tripathi, V. R., Khare, S. K. & Garg, S. K. Comparative one-factor-at-a-time, response surface (statistical) and bench-scale bioreactor level optimization of thermoalkaline protease production from a psychrotrophic Pseudomonas putida SKG-1 isolate. Microb. Cell Fact. 10, 114 (2011).
-
Aziz, R. K. et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genomics 9, 1–15 (2008).
-
Brettin, T. et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. https://doi.org/10.1038/srep08365 (2015).
-
Grant, J. R. et al. Proksee: In-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 51, W484–W492 (2023).
-
Milad, S., Saleh, S., Aboulwafa, M. & Hassouna, N. Lignin from a disposable by-product to a repository of value-added compounds. Arch. Pharm. Sci. Ain Shams Univ. https://doi.org/10.21608/aps.2023.229635.1132 (2023).
-
Kaur, R., Salwan, R. & Sharma, V. Structural properties, genomic distribution of laccases from Streptomyces and their potential applications. Process Biochem. 118, 133–144 (2022).
-
Chen, Y. H. et al. Biodegradation of kraft lignin by a bacterial strain Comamonas sp. B-9 isolated from eroded bamboo slips. J. Appl. Microbiol. 112, 900–906 (2012).
-
Kameche, K., Amrani, S., Mouzaoui, S. & Aït-Amar, H. Biodegradation of diazo dye Evans blue by four strains of Streptomyces isolated from soils of Algeria. Biocatal. Agric. Biotechnol. 46, 102529 (2022).
-
Buntić, A. V., Pavlović, M. D., Antonović, D. G., Šiler-Marinković, S. S. & Dimitrijević-Branković, S. I. A treatment of wastewater containing basic dyes by the use of new strain Streptomyces microflavus CKS6. J. Clean. Prod. 148, 347–354 (2017).
-
Hamedi, J., Vaez Fakhri, A. & Mahdavi, S. Biobleaching of mechanical paper pulp using Streptomyces rutgersensis UTMC 2445 isolated from a lignocellulose-rich soil. J. Appl. Microbiol. 128, 161–170 (2020).
-
Mamphogoro, T. P. Laccases from actinomycetes for lignocellulose degradation. M.Sc. thesis, University of the Western Cape (2012).
-
Tripathi, B. M., Kaushik, R., Kumari, P., Saxena, A. K. & Arora, D. K. Genetic and metabolic diversity of streptomycetes in pulp and paper mill effluent treated crop fields. World J. Microbiol. Biotechnol. 27, 1603–1613 (2011).
-
Depart-, M. & Building, L. S. Lignocellulose-degrading actinomycetes. FEMS Microbiol. Rev. 46, 145–163 (1987).
-
Niladevi, K. N., Sukumaran, R. K., Jacob, N., Anisha, G. S. & Prema, P. Optimization of laccase production from a novel strain-Streptomyces psammoticus using response surface methodology. Microbiol. Res. 164, 105–113 (2009).
-
Ashgan, A. G. A., Hassan, A. E. E., Samia, A. A. A. A., Amr, E. H. A. & Abdallah, M. E. The use of biotechnology for biodegradation of paper manufacturing wastes (Kraft Lignin) via egyptian bacterial isolates. Alexandria Sci. Exch. J. 32, 392–399 (2011).
-
Rath, S., Paul, M., Behera, H. K. & Thatoi, H. Response surface methodology mediated optimization of Lignin peroxidase from Bacillus mycoides isolated from Simlipal biosphere. J. Genet. Eng. Biotechnol. 20, 2 (2022).
-
Murugesan, R. & Vembu, B. Production of extracellular laccase from the newly isolated Bacillus sp. PK4. African J. Biotechnol. 15, 1813–1826 (2016).
-
Falade, A., Jaouani, A., Mabinya, L., Okoh, A. & Nwodo, U. Exoproduction and molecular characterization of peroxidase from Ensifer adhaerens. Appl. Sci. 9, 3121 (2019).
-
Shi, Y. et al. Characterization and genomic analysis of kraft lignin biodegradation by the beta-proteobacterium Cupriavidus basilensis B-8. Biotechnol. Biofuels 6, 1–14 (2013).
-
El-Hanafy, A. A., Abd-Elsalam, H. E. & Hafez, E. E. Fingerprinting for the lignin degrading bacteria from the Soil. J. Appl. Sci. Res. 3, 470–475 (2007).
-
Rafael, V. Bacterial degradation of lignin. Enzyme Microb. Technol. 10, 646–655 (1988).
-
Chandra, R. et al. Isolation and characterization of bacterial strains Paenibacillus sp. and Bacillus sp. for kraft lignin decolorization from pulp paper mill waste. J. Gen. Appl. Microbiol. 54, 399–407 (2008).
-
Rahman, N. H. A., Rahman, N. A., Aziz, S. A. & Hassan, M. A. Production of ligninolytic enzymes by newly isolated bacteria from palm oil plantation soils. BioResources 8, 6136–6150 (2013).
-
Olajuyigbe, F. M., Fatokun, C. O. & Oyelere, O. M. Biodelignification of some agro-residues by Stenotrophomonas sp. CFB-09 and enhanced production of ligninolytic enzymes. Biocatal. Agric. Biotechnol. 15, 120–130 (2018).
-
Kaushik, G. & Thakur, I. S. Production of laccase and optimization of its production by Bacillus sp. using distillery spent wash as inducer. Bioremediat. J. 18, 28–37 (2014).
-
Wen, X., Jia, Y. & Li, J. Enzymatic degradation of tetracycline and oxytetracycline by crude manganese peroxidase prepared from Phanerochaete chrysosporium. J. Hazard. Mater. 177, 924–928 (2010).
-
Aghaie-Khouzani, M., Forootanfar, H., Moshfegh, M., Khoshayand, M. R. & Faramarzi, M. A. Decolorization of some synthetic dyes using optimized culture broth of laccase producing ascomycete Paraconiothyrium variabile. Biochem. Eng. J. 60, 9–15 (2012).
-
Brown, M. E. & Chang, M. C. Y. Exploring bacterial lignin degradation. Curr. Opin. Chem. Biol. 19, 1–7 (2014).
-
Ahmad, M. et al. Identification of DypB from Rhodococcus jostii RHA1 as a lignin peroxidase. Biochemistry 50, 5096–5107 (2011).
-
Colpa, D. I., Fraaije, M. W. & van Bloois, E. DyP-type peroxidases: a promising and versatile class of enzymes. J. Ind. Microbiol. Biotechnol. 41, 1–7 (2014).
-
Granja-Travez, R. S. et al. Structural and functional characterisation of multi-copper oxidase CueO from lignin-degrading bacterium Ochrobactrum sp. reveal its activity towards lignin model compounds and lignosulfonate. FEBS J. 285, 1684–1700 (2018).
-
Tian, J. H., Pourcher, A. M., Bouchez, T., Gelhaye, E. & Peu, P. Occurrence of lignin degradation genotypes and phenotypes among prokaryotes. Appl. Microbiol. Biotechnol. 98, 9527–9544 (2014).
-
Palazzolo, M. A. & Kurina-Sanz, M. Microbial utilization of lignin: available biotechnologies for its degradation and valorization. World J. Microbiol. Biotechnol. 32, 173 (2016).
-
Kameshwar, A. K. S. & Qin, W. Qualitative and quantitative methods for isolation and characterization of lignin-modifying enzymes secreted by microorganisms. Bioenergy Res. 10, 248–266 (2017).
-
Bissaro, B., Várnai, A., Røhr, Å. K. & Eijsink, V. G. H. Oxidoreductases and reactive oxygen species in conversion of lignocellulosic biomass. Microbiol. Mol. Biol. Rev. https://doi.org/10.1128/MMBR.00029-18 (2018).
-
Li, J., Feng, Q., Du, X. & Yan, H. Quantitative proteomics analysis of natural lignocelluloses degraded by Aspergillus fumigatus g-13 based on itraq. Polish J. Environ. Stud. 30, 189–200 (2020).
-
Ichinose, H. Cytochrome P450 of wood-rotting basidiomycetes and biotechnological applications. Biotechnol. Appl. Biochem. 60, 71–81 (2013).
-
Iram, A., Berenjian, A. & Demirci, A. A review on the utilization of lignin as a fermentation substrate to produce lignin-modifying enzymes and other value-added products. Molecules 26, 2960 (2021).
-
Wischgoll, S. et al. Gene clusters involved in anaerobic benzoate degradation of Geobacter metallireducens. Mol. Microbiol. 58, 1238–1252 (2005).
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Milad, S., Saleh, S.E., Aboulwafa, M.M. et al. Screening and characterization of soil bacteria for lignin and textile dye effluent bioremediation and optimization using response surface methodology. Sci Rep 15, 25788 (2025). https://doi.org/10.1038/s41598-025-04789-5
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-04789-5
