Introduction
Articular cartilage is composed of water and a dense extracellular matrix (ECM), containing collagen type II fibers and proteoglycans, among others. Embedded within this matrix are chondrocytes, highly specialized cells responsible for maintaining and regulating articular cartilage by synthesizing or degrading ECM proteins. Despite their high metabolic activity, chondrocytes exhibit poor proliferative activity1. The lack of vascular and neural support in articular cartilage makes its repair or the prevention of damage progression intrinsically challenging2.
Photobiomodulation (PBM) is a noninvasive treatment widely explored for promoting tissue repair and it has been proposed to restore articular cartilage injury3,4,5. In PBM, human tissues are irradiated with light in the visible and near-infrared spectrum (600–1100 nm), eliciting photophysical and photochemical events at the cellular level. This therapy uses lasers, light-emitting diodes (LEDs), or broadband lights without causing a significant temperature rise during stimulation6. Lasers or LEDs, which provide stimulation at specific and well-defined wavelengths, are preferred over broadband lights. Although lasers are the most commonly used technology, LEDs offer a cost-effective and versatile alternative to lasers in PBM, delivering comparable therapeutic benefits with greater accessibility and reduced thermal risks6.
Recent research demonstrates that PBM enhances the proliferation and viability of human chondrocytes, increases ECM synthesis, and clinically decreases pain and inflammation7. However, despite this growing body of evidence, most studies are based mainly on pre- and clinical research, with only a limited number of in vitro investigations reporting PBM effects on chondrocytes8,9,10,11,12. In vitro studies are crucial for advancing our current knowledge on PBM’s effects, as they allow the assessment of the therapy effects at the chondrocyte level. Furthermore, exhaustive parameter optimization of PBM dosage can be screened without inflicting pain or harm to animals. The lack of studies aimed at identifying the most appropriate PBM dosage has hindered its broader application for articular cartilage treatment.
This work aims to establish the most appropriate PBM protocols for chondrocyte stimulation. Firstly, a comprehensive PBM optimization was carried out, testing a wide range of wavelengths, intensities, durations, periodicities, and operation modes to evaluate their effect on chondrocyte viability and proliferation. The most promising PBM protocols were then further scrutinized to elucidate their influence on cartilage ECM synthesis.
Materials and methods
Cell culture and seeding
Primary human chondrocytes were isolated from a healthy articular cartilage from the knee of a 46-year-old female and purchased from Innoprot (Spain). The chondrocytes were grown in chondrocyte basal medium (Innoprot, Spain) supplemented with 5% (v/v) of fetal bovine serum (FBS, Innoprot, Spain), 1% (v/v) penicillin/streptomycin (Innoprot, Spain) and 1% (v/v) chondrocyte growth factors (Innoprot, Spain) at 37 °C and with 5% CO2. The culture medium was refreshed every two days. Once the cells reached 90% confluency, they were trypsinized using 0.25% (v/v) trypsin/EDTA (PAN-Biotech GmbH, Germany) and subcultured according to the manufacturer´s indications. Human chondrocytes from passages 3 and 4 were used in all experiments.
Photobiomodulation
Optical actuation setup
A customized device was developed by our research group to deliver PBM at wavelengths of 600, 655, 810, 850, and 940 nm to cultured cells in 12-well plates (TPP, Switzerland). The stimulation device was integrated into a print circuit board containing an array of surface mounted device (SMD) LEDs positioned over each well and distributed in a circular area of 4.9 cm2. The number of LEDs in each array was specific for each wavelength and determined based on a prior experimental characterization of the optical power density measured by a commercial power meter (Newport). The defined arrays were as follows: an 8-LEDs array for 600 nm (Lumileds, USA), a 16-LEDs array for 655 nm (Osram GmbH, Austria), 850 nm (Würth Elektronik eiSos GmbH & Co. KG, Germany) and 940 nm (Osram GmbH, Austria), and a 4-LEDs array for 810 nm (Luminus, USA). The device was connected to a computer via USB, and its current, the stimulation time, and the on/off time (for pulsed light) were controlled through a customized software developed by our research group.
The stimulation device was sterilized using UV light for 20 min inside a laminar airflow chamber (Gelaire®, Australia). Before conducting cellular experiments, the optical actuation setup was tested and calibrated to ensure a correct optical intensity at the bottom of the culture wells.
Temperature measurement
Culture medium temperature during PBM was monitored by immersing a sterilized thermometer into the culture media inside the well plate. For all stimulation parameters, temperature was recorded at 1-min intervals for up to 4 min under experimental conditions like those used for cellular experiments. Prior to measurement, the culture medium temperature was stabilized at room temperature.
Stimulation protocol
Human chondrocytes were seeded at 15,000 cells/mL one day prior to stimulation. Various PBM parameters were employed over three days to carry out an initial screening of the most relevant stimulation protocols for enhancing chondrocyte activity. Specifically, wavelengths of 600, 655, 810, 850, and 940 nm at power densities (i.e., intensities) of 4, 7, 10, 14 and 17 mW/cm2 were tested. The PBM was applied either in continuous mode for 1 min or in pulsed mode for 2 min with a 1 Hz pulse frequency (matching the native walking frequency13) and a 50% duty cycle, assuring that both operation modes received the same irradiation energy. From those parameters, the applied energy density and total energy were obtained. The PBM was applied either daily (three sessions) or every other day (two sessions). Following the identification of the most promising stimulation parameters, the stimulation duration was increased up to 4 min (240 s) to assess the effect of longer stimulation on chondrocyte response. Each stimulation parameter was carefully calibrated to ensure precise energy delivery to the cell culture surface. Table 1 summarizes the stimulation parameters optimized for human chondrocytes.
To assess the influence on cartilage matrix synthesis, chondrocytes were seeded at a density of 12,000 cells/mL and stimulated over seven days with the most effective PBM parameters.
Chondrocyte stimulation was conducted in a sterile environment within a laminar flow cabinet at room temperature. Following stimulation, the 12-well plates were maintained inside the incubator at 37 °C and with 5% CO2. Human chondrocytes were collected one day after the last stimulation session. A control group was included, where the customized stimulation device was turned off but positioned over the cultured 12-well plate.
Metabolic activity
The metabolic activity of both stimulated and non-irradiated human chondrocytes was measured by the 3-(4,5-dimethylthiazol -2-yl) -5-(3-carboxymethoxyphenyl) 2-(4-sulfophenyl) -2H-tetrazolium (MTS) assay (MTS Cell Proliferation Colorimetric Assay Kit, Abcam, United Kingdom) following three days of PBM stimulation. One day after the last stimulation, the MTS reagent was added to the culture medium of each well, according to the manufacturer´s protocol, and incubated for 3 h at 37 °C with 5% CO2. A blank control, containing only the culture medium without cultured cells, was also prepared. After 3 h, the absorbance of the MTS-treated culture medium was read at 490 nm using a microplate reader (Biotek Epoch, United States). The absorbance of the blank was subtracted from that of the remaining wells to obtain corrected values.
Cell proliferation
The number of chondrocytes after three days of stimulation was quantified. On the day following the last stimulation session, cultured human chondrocytes were detached from the well bottom using 0.25% (v/v) trypsin/EDTA (PAN-Biotech GmbH, Germany) for 5 min at 37 °C. The cells were then centrifuged at 300 g for 5 min at room temperature. The obtained cell pellet was resuspended in culture medium and stained with 0.4% (v/v) trypan blue (PAN-Biotech GmbH, Germany). Cell counting was performed in a Neubauer chamber using a microscope (Kern & Sohn GmbH, Germany) at 10 × magnification. The MTS assay measurements were normalized to the cell number for each condition to ensure accuracy.
Immunocytochemistry
After seven days, the presence of collagen type II (COL II), aggrecan (ACAN) and collagen type I (COL I) in human chondrocytes cultures was investigated after stimulating with the most promising PBM protocols. Following three PBS washes, irradiated and non-irradiated cells were fixed with 10% (v/v) formalin (Sigma-Aldrich, USA) for 20 min at room temperature, then blocked with 50 mM ammonium chloride (NH4Cl, ITW Reagents, IT) for 10 min at room temperature. Cells were permeabilized with 0.1% (w/v) sodium dodecyl sulfate (SDS, Thermo Fisher Scientific, USA) for 10 min at room temperature, followed by blocking with 3% (v/v) bovine serum albumin (BSA, NZYtech, PT) in PBS for 20 min at room temperature. Afterwards, chondrocytes were incubated overnight at 4⁰C with primary antibodies diluted in 0.1% BSA (w/v) in phosphate buffered saline (PBS) against COL I (Abbkine, USA, 1:50), COL II (Abbkine, USA, 1:50) and ACAN (Assay Biotechnology, USA, 1:50). After three washes with 0.1% BSA (w/v) in PBS, chondrocytes incubated with primary antibody against COL I and COL II were further incubated with anti-mouse secondary antibody conjugated with Andy fluor 488 (ABP Biosciences, USA, 10 µg/mL) in 0.1% BSA (w/v) in PBS, while chondrocytes incubated with primary antibody against ACAN were incubated with anti-rabbit secondary antibody conjugated with Alexa fluor 488 (Abcam, UK, 1:200) in 0.1% BSA (w/v) in PBS for 1 h at room temperature, protected from light. Next, cells were incubated with 1 µg/mL DAPI (Thermo Fisher Scientific, USA) in PBS for 15 min at room temperature, protected from light. Chondrocytes were visualized using a fluorescent microscope (Olympus BX51) at 40 × magnification, using the appropriate filter settings for Alexa fluor 488, Andy fluor 488 and DAPI. The fluorescent intensity was quantified using the ImageJ software (version 1.54g).
Western blot
For the most effective PBM parameters, non-irradiated and irradiated chondrocytes were collected, and COL II, ACAN and COL I expression were assessed by Western Blot. Protein was extracted from chondrocytes with radioimmunoprecipitation assay (RIPA) buffer containing protease inhibitors. The Pierce BCA Protein Assay kit (Thermo Fisher Scientific, USA) was used to determine the protein concentration and 20 μg of total protein were loaded, separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto a methanol activated polyvinylidene difluoride (PVDF) membrane (Thermo Fisher Scientific, USA). The membranes were blocked with 5% (w/v) non-fat milk in tris-buffered saline containing 0.1% (v/v) Tween 20 (TBS-T) for 1 h at room temperature. The membranes were then incubated with primary antibodies diluted in TBS-T with 5% (w/v) BSA overnight at 4 °C: COL II (1:200 dilution, sc-518017, Santa Cruz Biotechnology, USA), ACAN (1:500 dilution, L0101, Assay Biotechnology, USA), COL I (1:1000 dilution, #72,026, Cell Signaling Technology, USA) and β-actin (1:40,000 dilution, Merck, USA). After three TBS-T washes, the membranes were incubated with secondary antibodies anti-rabbit or anti-mouse horseradish peroxidase (HRP, Jackson immunoResearch, USA) for 90 min at room temperature. The bands were visualized using a chemiluminescent substrate (WesternBright ECL HRP Substrate, Advansta, USA) in a ChemiDoc XRS + system (Bio-Rad). Band intensity was quantified by ImageJ software (version 1.54g).
Alcian blue staining
After seven days, sulfated glycosaminoglycans (GAGs) synthetized by chondrocyte cultures (e.g., chondroitin sulfate and keratan sulfate) were quantified through alcian blue staining, following a protocol adapted from literature14,15. Irradiated and non-irradiated chondrocytes were washed with PBS and fixed with 10% (v/v) formalin for 20 min at room temperature. Afterwards, cells were incubated overnight with 1% (w/v) Alcian Blue 8-GX (Sigma Aldrich, USA) in 0.1 M HCl (pH 1.0) (Sigma Aldrich, USA) at room temperature in gentle agitation, protected from light. After washing three times with PBS, cells were observed under a fluorescence microscope (Kern & Sohn GmbH, Germany) at a 10 × magnification.
Stained chondrocyte cultures were incubated with 6 M guanidine HCl (Sigma-Aldrich, USA) at room temperature, protected from light, for 3 h in gentle agitation to remove the bound alcian blue. The absorbance of the extracted dye as measured at 630 nm using a microplate reader (Biotek Epoch, United States). A blank was established with fresh 6 M guanidine HCl, and its absorbance values were subtracted to the experimental ones.
Statistical analysis
Statistical analysis was performed using the 26.0 version of the Statistical Package for the Social Sciences (SPSS, IBM, Chicago, IL) software for Windows. Firstly, the normality of the data was assessed by the Shapiro–Wilk test. For data that did not follow a normal distribution, log transformation was applied. Normally distributed data was analyzed by the independent sample T-test to check for significant differences between each PBM group with control group. For data without parametric distribution, even after log transformation, the non-parametric Mann–Whitney test was applied. The p < 0.05 was considered statistically significant and the results were depicted as mean ± standard deviation. The results were obtained from at least three independent tests.
Results
Optimization of the most promising dosage on chondrocyte activity
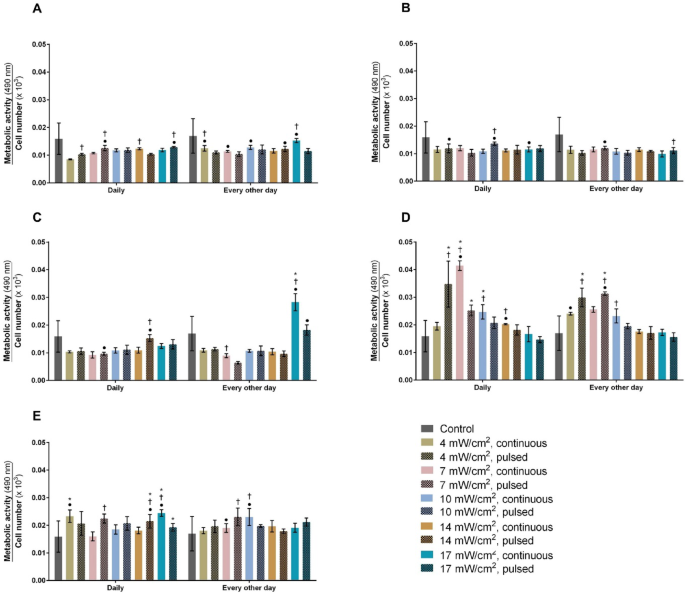
Human chondrocytes were exposed to a wide range of PBM parameters, and their metabolic activity and proliferation were assessed (Fig. 1).

Normalized metabolic activity of non-irradiated (control) and irradiated human chondrocytes with (A) 600 nm, (B) 655 nm, (C) 810 nm, (D) 855 nm and (E) 940 nm LEDs, daily and every other day, in continuous and pulsed modes, at power densities of 4, 7, 10, 14 and 17 mW/cm2, after three days (n = 3). * significant differences compared to the untreated chondrocytes; † significant differences between continuous and pulsed mode; and ● significant differences between daily and every other day application.
Effect of wavelength, operation mode and periodicity
Stimulating human chondrocytes with 600 nm LED did not alter their activity in comparison to untreated cells at any intensity (Fig. 1A). Nonetheless, applying PBM in continuous or pulsed mode, either daily or every other day, significantly increased chondrocyte activity, with variations depending on the combination of parameters. No changes in the chondrocyte activity were detected following 655 nm stimulation compared to non-irradiated cells (Fig. 1B). At this wavelength, continuous and pulsed mode induced similar biological responses, except for intensities of 10 (p < 0.001) and 17 mW/cm2 (p = 0.004), where pulsed mode was statistically superior to continuous mode (Fig. 1B). Daily or every other day application influenced the chondrocytes´ activity under certain PBM parameters, except at 14 mW/cm2, where no effect was observed.
Stimulation with 17 mW/cm2 in continuous mode every other day was the only condition that significantly enhanced the normalized metabolic activity of chondrocytes (p < 0.001) at this wavelength compared to untreated cells (Fig. 1C). The effects of continuous versus pulsed mode and daily versus every other day application differed across PBM parameters. However, no alteration in chondrocyte activity was noticed at intensities of 4 and 10 mW/cm2, regardless of the operation mode (continuous or pulsed) or application frequency (daily or every other day).
The 850 nm stimulation significantly improved the normalized metabolic activity of chondrocytes in comparison to untreated cells under the following conditions: pulsed 4 mW/cm2 applied daily or every other day (p = 0.001); 7 mW/cm2 daily in continuous mode or every other day in both operation modes (p < 0.001); and continuous 10 mW/cm2 (p < 0.001) daily (Fig. 1D). The operation mode or periodicity significantly influenced the chondrocyte activity, depending on the combination of PBM parameters. Nevertheless, increasing the intensity to 10 or 17 mW/cm2 did not produce changes between daily and every other day applications. Also, no changes in metabolic activity were observed when chondrocytes were subjected to 17 mW/cm2, regardless of whether continuous or pulsed mode was used.
Human chondrocytes stimulated at 940 nm displayed significantly enhanced normalized metabolic activity compared to untreated cells under the following conditions: continuous 4 mW/cm2 applied daily (p = 0.006); pulsed 14 mW/cm2 daily (p = 0.0003); and 17 mW/cm2 in continuous (p < 0.001) and pulsed (p = 0.009) modes with daily sessions (Fig. 1E). Both the operation mode and periodicity influenced the chondrocyte activity regardless of the intensity applied, except for 4 mW/cm2, where no effect of the operation mode was observed.
Influence of stimulation time
The wavelengths that induced significant differences in the normalized metabolic activity were 810, 850, and 940 nm, across various power densities, periodicities, and operation modes. Based on the statistical analysis, only a few stimulation conditions were selected for further optimization, focusing on the lowest p value for each stimulation condition, further explained in Supplementary Table S1. For instance, 4 mW/cm2 at 850 nm LED promoted statistically significant differences in normalized metabolic activity in pulsed mode, both daily and every other day. However, the p value for the every-other-day application was lower (p < 0.001) compared to the daily application (p = 0.001), so every-other-day application was selected. This selection was applied accordingly to all conditions that produced significant alterations. The most promising parameters were 17 mW/cm2 (continuous, every other day) at 810 nm stimulation; 4 mW/cm2 (pulsed, every other day), 7 mW/cm2 (continuous, daily) and 10 mW/cm2 (continuous, daily) at 850 nm stimulation; and 4 mW/cm2 (continuous, daily), 14 mW/cm2 (pulsed, daily) and 17 mW/cm2 (continuous, daily) at 940 nm stimulation. For these promising stimulation conditions, the stimulation time was increased up to 240 s, and the normalized metabolic activity was investigated. However, increasing the stimulation time did not result in a proportional enhancement of the normalized metabolic activity (Fig. 2).

Normalized metabolic activity of human chondrocytes stimulated with the most promising PBM parameters, along with increasing stimulation times (60, 120, 180 and 240 s), after three days (n = 3). * significant differences compared to the other stimulation times for the same PBM condition; and † significant differences among all PBM conditions.
Increasing the duration of 810 nm stimulation on chondrocytes did not influence their biological response (Fig. 2). For the 850 nm stimulation, chondrocytes subjected to 4 mW/cm2 (pulsed, every other day) showed significantly enhanced normalized metabolic activity up to 240 s of stimulation, compared to a 60 s stimulation (p = 0.004). However, no differences were observed between 120 s, 180 s, or 240 s stimulation times. Thus, a 120 s stimulation time was preferred over 180 or 240 s. With 7 mW/cm2 (continuous, daily), 240 s of stimulation was required to significantly increase the normalized metabolic activity, as opposed to 60 s (p = 0.006). For 10 mW/cm2 (continuous, daily), stimulating for 120 s resulted in higher normalized metabolic activity compared to 60 s stimulation, although no statistically significant differences were noticed between the two time points. Longer times (180 and 240 s) decreased chondrocyte activity (p < 0.001), making 60 s stimulation sufficient to significantly improve chondrocyte response (Fig. 2).
For the three 940 nm conditions, normalized metabolic activity increased up to 180 s of stimulation, showing a downward tendency at 240 s (Fig. 2). Thus, a 180 s stimulation time was deemed optimal for these conditions (p < 0.004).
Considering the stimulation time of each PBM protocol that significantly promoted chondrocyte activity, the protocol using 14 mW/cm2 (pulsed, daily) with the 940 nm LED for 180 s (p < 0.002) and 17 mW/cm2 (continuous, every other day) with the 810 nm LED (p < 0.008) resulted in significantly lower improvements in normalized metabolic activity of human chondrocytes compared to other conditions (Fig. 2).
Temperature measurement
The highest temperature increase was detected with continuous 600 nm LEDs at 17 mW/cm2, resulting in a rise of 1.6 °C. The majority of PBM protocols, including the most effective ones, induced minimal temperature increases of up to 0.4 °C, with a few conditions causing variations between 0.5 and 0.9 °C (Supplementary Table S2).
Influence of photobiomodulation on cartilage matrix synthesis and chondrocyte dedifferentiation
Following the extensive optimization of PBM protocols, human chondrocytes were further stimulated for seven days using the most effective protocols: 4 mW/cm2, 850 nm, pulsed (1 Hz), 120 s, every other day (LA); 4 mW/cm2, 940 nm, continuous, 180 s, daily (LB); 7 mW/cm2, 850 nm, continuous, 240 s, daily (LC); 10 mW/cm2, 850 nm, continuous, 60 s, daily (LD); 17 mW/cm2, 940 nm, continuous, 180 s, daily (LE). These PBM protocols were applied to investigate their ability to induce the synthesis of cartilage proteins, namely COL II and ACAN, and to decrease the expression of COL I, a dedifferentiation marker in chondrocytes.
Effects on chondrocyte metabolic activity after seven days
The metabolic activity of human chondrocytes was reassessed after irradiation with the most promising PBM protocols for seven days (Fig. 3). Chondrocytes stimulated with 4 mW/cm2 at 850 nm (pulsed, every other day, p < 0.001) or 940 nm (continuous, daily, p < 0.001), and with 7 mW/cm2 at 850 nm (continuous, daily, p < 0.001), exhibited statistically significant enhanced normalized metabolic activity compared to non-irradiated cells, i.e., untreated controls, highlighting the responsiveness of chondrocytes to near-infrared stimulation. In contrast, the PBM protocol 17 mW/cm2 at 940 nm (continuous, daily) resulted in significantly reduced metabolic activity compared to control (p = 0.002).

Normalized metabolic activity of non-irradiated (control) and irradiated human chondrocytes with 4 mW/cm2, 850 nm, pulsed (1 Hz), 120 s, every other day (LA); 4 mW/cm2, 940 nm, continuous, 180 s, daily (LB); 7 mW/cm2, 850 nm, continuous, 240 s, daily (LC); 10 mW/cm2, 850 nm, continuous, 60 s, daily (LD); and 17 mW/cm2, 940 nm, continuous, 180 s, daily (LE) for 7 days (n = 3). Statistical differences compared to non-irradiated chondrocytes denoted as * p < 0.05 and *** p < 0.001.
Effects of cartilage matrix
Immunocytochemistry analyses (Fig. 4) revealed that human chondrocytes were positively stained for COL II (Fig. 4A) and ACAN (Fig. 4B), with a more pronounced staining observed in cells stimulated with specific PBM protocols, in comparison to non-irradiated chondrocytes. In contrast, COL I was slightly stained in chondrocytes, particularly in the stimulated chondrocytes, compared to control (Fig. 4C).

Representative fluorescence microscopic images of non-irradiated (control) and irradiated human chondrocytes with 4 mW/cm2, 850 nm, pulsed (1 Hz), 120 s, every other day (LA); 4 mW/cm2, 940 nm, continuous, 180 s, daily (LB); 7 mW/cm2, 850 nm, continuous, 240 s, daily (LC); 10 mW/cm2, 850 nm, continuous, 60 s, daily (LD); and 17 mW/cm2, 940 nm, continuous, 180 s, daily (LE) for 7 days (n = 3). Cell nucleus was stained in blue with DAPI and (A) COL II, (B) ACAN and (C) COL I expression were stained in green. Scale bar: 20 µm.
Quantification of COL II staining intensity (Fig. 5A) revealed that stimulated chondrocytes significantly increased COL II production in the ECM for most of the PBM protocols applied, except for 10 mW/cm2 at 850 nm (continuous, daily, p < 0.001), which significantly reduced its expression. ACAN expression (Fig. 5B) was also significantly elevated following PBM (p < 0.001). Stimulating chondrocytes with 4 mW/cm2 at 850 nm (pulsed, every other day) did not alter ACAN production. COL I production was also observed in chondrocytes (Fig. 5C), but its expression was significantly downregulated following PBM exposure with 4 mW/cm2 at 850 nm (pulsed, every other day, p < 0.001); or 7 mW/cm2 at 850 nm (continuous, daily, p = 0.025). In contrast, PBM with 10 mW/cm2 at 850 nm (continuous, daily, p < 0.001) significantly increased COL I production in the ECM. No significant effects on COL I expression were observed for the remaining PBM protocols.

Quantification of the fluorescence staining intensity: (A) COL II, (B) ACAN and (C) COL I in non-irradiated (control) and irradiated human chondrocytes with 4 mW/cm2, 850 nm, pulsed (1 Hz), 120 s, every other day (LA); 4 mW/cm2, 940 nm, continuous, 180 s, daily (LB); 7 mW/cm2, 850 nm, continuous, 240 s, daily (LC); 10 mW/cm2, 850 nm, continuous, 60 s, daily (LD); and 17 mW/cm2, 940 nm, continuous, 180 s, daily (LE) for 7 days (n = 3). Staining intensity was measured using ImageJ software (version 1.54 g). Statistical differences compared to non-irradiated chondrocytes denoted as *p < 0.05, ** p < 0.01, *** p < 0.001.
Protein content of collagen type II, aggrecan and collagen type I
Western blot analysis was carried out to assess the protein content of chondrocyte cultures (Fig. 6). There was no significant increase in COL II protein levels in chondrocytes exposed to PBM (Fig. 6A). On the contrary, stimulation with 4 mW/cm2 at 850 nm (pulsed, every other day, p < 0.001) or 4 mW/cm2 at 940 nm (continuous, daily, p = 0.002) significantly reduced COL II expression compared to control. A trend towards increased COL II expression was observed with increasing intensity. ACAN content was significantly upregulated following stimulation with 10 mW/cm2 at 850 nm (continuous, daily, p = 0.049), increasing by 2.0-fold, and 17 mW/cm2 at 940 nm (continuous, daily, p = 0.001), increasing by 1.4-fold, compared to non-irradiated chondrocytes (Fig. 6B). No alteration in ACAN production was detected for the other PBM protocols. COL I expression was significantly reduced after 4 mW/cm2 at 850 nm (pulsed, every other day, p = 0.003), but remained unaffected in chondrocytes exposed with other PBM protocols (Fig. 6C).

Representative images of western blot analysis of protein expression of (A1) COL II, (B1) ACAN and (C1) COL I and western blot analysis of protein expression of (A2) COL II, (B2) ACAN and (C2) COL I in non-irradiated (control) and irradiated human chondrocytes with 4 mW/cm2, 850 nm, pulsed (1 Hz), 120 s, every other day (LA); 4 mW/cm2, 940 nm, continuous, 180 s, daily (LB); 7 mW/cm2, 850 nm, continuous, 240 s, daily (LC); 10 mW/cm2, 850 nm, continuous, 60 s, daily (LD); and 17 mW/cm2, 940 nm, continuous, 180 s, daily (LE) for 7 days (n = 3). Statistical differences compared to non-irradiated chondrocytes denoted as *p < 0.05, ** p < 0.01, *** p < 0.001. Uncropped Western Blot images can be found in Supplementary Figure S1.
Synthesis of sulfated glycosaminoglycans
Sulfated GAGs were quantified by staining chondrocytes with alcian blue. Microscopic images revealed that chondrocytes were positively stained for GAGs (Fig. 7A). After extracting the alcian blue dye, absorbance measurements indicated that chondrocytes stimulated with 17 mW/cm2 at 940 nm (continuous, daily, p < 0.001) presented a higher production of GAGs compared to untreated chondrocytes (Fig. 7B).

(A) Representative microscopic images of non-irradiated (control) and irradiated human chondrocytes stained with alcian blue and (B) quantification of alcian blue staining. Chondrocytes were stimulated with 4 mW/cm2, 850 nm, pulsed (1 Hz), 120 s, every other day (LA); 4 mW/cm2, 940 nm, continuous, 180 s, daily (LB); 7 mW/cm2, 850 nm, continuous, 240 s, daily (LC); 10 mW/cm2, 850 nm, continuous, 60 s, daily (LD); and 17 mW/cm2, 940 nm, continuous, 180 s, daily (LE) for 7 days (n = 3). Scale bar: 20 µm. Statistical differences compared to non-irradiated chondrocytes denoted as *** p < 0.001.
Discussion
This study identified crucial findings regarding the optimal PBM dosage for enhancing chondrocyte biological responses. The most effective PBM parameters comprised wavelengths of 850 nm and 940 nm, applied with intensities ranging from 4 to 17 mW/cm2, for up to 240 s and up to 15 J. These parameters consistently and significantly enhanced metabolic activity and the secretion of cartilage-specific proteins, while also reducing the dedifferentiation process in human chondrocytes.
Chondrocytes are highly specialized cells in articular cartilage, typically in a quiescent state. Cell division occurs in less than 1% of chondrocytes1. Despite their limited cell division, they remain metabolically active by secreting ECM proteins to maintain the normal tissue homeostasis. Therefore, metabolic activity was the primary outcome used to select the most promising PBM protocols for chondrocytes.
Previous in vitro studies applying wavelengths of 632 nm, 808 nm, 830 nm, and 910 nm to chondrocytes reported statistically significant increases in activity following PBM8,9,10,11,12. In comparison, our findings showed that red irradiation at 600 nm or 655 nm did not significantly enhance normalized metabolic activity on chondrocytes. This discrepancy may be due to the higher energy densities used in previous studies, up to 10 J/cm2, whereas this study applied much lower energy densities, up to 1 J/cm2. This suggests that higher power densities or energy densities may be required for these wavelengths to potentiate chondrocyte activity. Notably, wavelengths in the near-infrared spectrum (810 nm, 850 nm, and 940 nm) induced significant variations in chondrocyte activity even at low intensity, suggesting their greater potential for stimulating human chondrocytes compared to red radiation.
The influence of operation mode, periodicity, and dose-dependency of PBM on chondrocytes was investigated. Both operation modes were tested to analyze whether human chondrocytes exhibited distinct responses when stimulated with the same energy but delivered continuously for 1 min or pulsed for 2 min (with 50% duty cycle and 1 Hz pulse frequency). Both operation modes (i.e., continuous vs pulsed) equally and significantly augmented chondrocyte biological responses, with the choice of mode influenced by the combination of parameters. A similar trend was observed for periodicity (i.e., daily vs every-other day), which agrees with a previous study showing similar results in chondrocyte proliferation when subjected to a single or multiple PBM sessions12. To investigate the dose-dependency of PBM, stimulation time was increased from 1 min (60 s) to 4 min (240 s) for the most promising conditions, thereby increasing energy density. However, increasing the stimulus intensity did not result in a higher biological response, suggesting the presence of a threshold for each PBM condition. Beyond this threshold, the biological response of chondrocytes either plateaued or declined, supporting previous findings16,17. Based on the exhaustive optimization of PBM parameters, the most promising conditions for potentiating chondrocyte metabolic activity were further studied.
The remarkable mechanical performance of articular cartilage is attributed to the interplay between COL II fibers and proteoglycans. Upon damage, disruption of COL II fibers and loss of proteoglycans may occur, compromising the tissue’s integrity18,19. Herein, PBM is proposed to stimulate cartilage repair by inducing human chondrocytes to produce more ECM components to fill the damage. In fact, PBM can modulate chondrocyte activity, stimulating the synthesis of cartilage matrix proteins9,12, although the effect is influenced by the applied stimulation protocols. In this study, not all PBM protocols exhibited a stimulatory effect on chondrocytes. Specifically, 7 mW/cm2 at 850 nm, and 4 mW/cm2 or 17 mW/cm2 at 940 nm significantly improved both COL II and ACAN expression, as verified in the immunocytochemistry experiments. In fact, ACAN synthesis was 1.5- to 2.1-fold elevated, while COL II was upregulated by 1.3- to 1.5- fold. When analyzing protein levels, only 10 mW/cm2 at 850 nm and 17 mW/cm2 at 940 nm enhanced ACAN expression by up to 2.0-fold, while not affecting COL II protein expression compared to control. A similar trend was noticed for the experimental condition 17 mW/cm2 at 940 nm, which significantly promoted GAGs synthesis in chondrocytes cultures. This finding is consistent with a previous study utilizing PBM at 940 nm20.
In vitro culture of human chondrocytes is challenging since they undergo dedifferentiation with increasing number of passages, which is characterized by a higher production of COL I relative to COL II21. However, stimulating chondrocytes with PBM in this study reversed this tendency. The stimulation protocols herein implemented either declined or preserved the COL I expression when compared to non-irradiated cells, aside from 10 mW/cm2 at 850 nm which exhibited a stimulatory effect on COL I production. These findings strengthen the existing literature, which reports the ability of PBM to reverse chondrocyte dedifferentiation9,12.
Pre-clinical and clinical research have been exploring the application of PBM on cartilage tissue. Although there is much research supporting the use of PBM, its efficacy in restoring articular cartilage is still controversial. Animal studies have shown that PBM upregulated the ECM proteins and downregulated cartilage degradation-related markers22,23,24, but, when combined with other therapies, PBM was not superior25,26,27. Similarly, patients in randomized clinical studies experienced improved function and pain relief, following PBM28,29, while other studies did not report any effect30,31. These inconsistent therapeutic effects may be a result of the lack of dosage standardization. In vitro models offer an appropriate framework to study PBM dosage as various stimulation protocols can be tested and adapted, according to the bioeffects on chondrocytes. Although these models do not mimic the native environment, the influence of PBM on chondrocytes can be screened out and further translated into animal models. Therefore, this in vitro study aims at providing directional guides towards the most adequate PBM dosage for chondrocytes stimulation. The most promising PBM dosage is the one that can increase cartilage proteins synthesis without affecting the natural dedifferentiation process of chondrocytes. Our findings pinpoint the potential of the 17 mW/cm2 at 940 nm PBM protocol in stimulating cartilage regeneration, as it consistently promoted the secretion of COL II, ACAN and GAGs (Supplementary Table S3). Moreover, this stimulation protocol did not influence the chondrocyte phenotype. Future studies could explore alternative parameters, such as increasing the number of sessions or the intensity, as these may further potentiate the chondrocyte biological response.
The demonstrated effectiveness of LEDs in stimulating chondrocytes, particularly in comparison to laser diodes, presents a cost-effective and customizable alternative for PBM. This could lead to the development of an efficient therapeutic intervention that is accessible to a wider population, particularly those suffering from articular cartilage injuries. The minimal temperature rise (< 1.0 °C) observed during stimulation further supports the feasibility and safety of using LEDs in clinical applications. The most promising PBM parameters align with the World Association for Laser Therapy (WALT) guidelines32,33, suggesting a potential integration of these findings into existing clinical protocols for knee stimulation. However, these guidelines currently apply only to laser diodes. Therefore, the inclusion of LEDs in these guidelines for cartilage stimulation, particularly for near-infrared LED radiation at 850 nm and 940 nm, is warranted, since it significantly enhanced key markers of cartilage repair, such as COL II and ACAN. These findings align with the biological premise that near-infrared radiation penetrates cellular structures more efficiently, stimulating deeper tissue responses.
This study’s in vitro design, while providing a robust framework for optimizing PBM parameters, presents limitations since it does not fully replicate the physiological environment of cartilage. Future research should extend these findings to preclinical and clinical settings to validate their translational applicability.
The insights gained from this study have promising implications for cartilage stimulation and repair. In addition to identifying optimal PBM parameters for enhancing cartilage regeneration, the study emphasizes the importance of considering factors such as operation mode, power density, and periodicity when tailoring PBM protocols. Integrating LEDs into therapeutic guidelines, along with a more nuanced understanding of PBM parameters, holds the potential to significantly impact and restore the articular cartilage degeneration.
Conclusions
The PBM stimulation using LEDs in near-infrared spectrum (i.e., 850 and 940 nm) proved to be the most effective in enhancing chondrocyte activity, even at low intensities (i.e., 4 to 17 mW/cm2) and energy densities (i.e., up to 3 J/cm2), demonstrating significant potential in promoting cartilage repair. Particularly, 940 nm with 17 mW/cm2, applied daily for seven days, consistently promoted the synthesis of cartilage proteins while delaying the normal dedifferentiation of chondrocytes during monolayer culture and successive expansion. By enhancing chondrocyte activity and matrix synthesis while minimizing dedifferentiation, this approach offers a promising avenue for developing accessible and scalable therapeutic interventions. This study considerably advances the current knowledge of PBM effectiveness, unveiling its ability to effectively restore articular cartilage, as well as an important step in the definition of the specifications of future LED-based PBM medical devices.
Data availability
Data is provided within the manuscript or supplementary information files.
References
-
Lyndin, M. et al. Morphofunctional features of articular cartilage structure, Folia Med. Cracov. 59, 81–93. https://doi.org/10.24425/fmc.2019.131138 (2019).
-
Baumann, C. A., Hinckel, B. B., Bozynski, C. C. & Farr, J. Articular cartilage: Structure and restoration. Jt. Preserv. Knee A Clin. Case https://doi.org/10.1007/978-3-030-01491-9_1 (2019).
-
Bozkurt, S. B., Hakki, E. E., Kayis, S. A., Dundar, N. & Hakki, S. S. Biostimulation with diode laser positively regulates cementoblast functions, in vitro. Lasers Med. Sci. 32, 911–919. https://doi.org/10.1007/s10103-017-2192-z (2017).
-
Tschon, M., Incerti-Parenti, S., Cepollaro, S., Checchi, L. & Fini, M. Photobiomodulation with low-level diode laser promotes osteoblast migration in an in vitro micro wound model. J. Biomed. Opt. 20, 078002. https://doi.org/10.1117/1.jbo.20.7.078002 (2015).
-
Salehpour, F. et al. Transcranial near-infrared photobiomodulation attenuates memory impairment and hippocampal oxidative stress in sleep-deprived mice. Brain Res. 1682, 36–43. https://doi.org/10.1016/j.brainres.2017.12.040 (2018).
-
Heiskanen, V. & Hamblin, M. R. Photobiomodulation: Lasers: vs. light emitting diodes?. Photochem. Photobiol. Sci. 17, 1003–1017. https://doi.org/10.1039/c8pp00176f (2018).
-
Oliveira, S. et al. In vitro and in vivo effects of light therapy on cartilage regeneration for knee osteoarthritis : A systematic review. Cartilage. 13, 1–20. https://doi.org/10.1177/19476035211007902 (2021).
-
Sakata, S. et al. High-frequency near-infrared diode laser irradiation attenuates IL-1β-induced expression of inflammatory cytokines and matrix metalloproteinases in human primary chondrocytes. J. Clin. Med. 9, 881. https://doi.org/10.3390/jcm9030881 (2020).
-
Yang, X. et al. Promoted viability and differentiated phenotype of cultured chondrocytes with low level laser irradiation potentiate efficacious cells for therapeutics, front. bioeng. Biotechnol. 8, 1–16. https://doi.org/10.3389/fbioe.2020.00468 (2020).
-
Jia, Y. L. & Guo, Z. Y. Effect of low-power he-ne laser irradiation on rabbit articular chondrocytes in vitro. Lasers Surg. Med. 34, 323–328. https://doi.org/10.1002/lsm.20017 (2004).
-
Tim, C. R. et al. Effects of photobiomodulation therapy in chondrocyte response by in vitro experiments and experimental model of osteoarthritis in the knee of rats. Lasers Med. Sci. 37, 1677–1686. https://doi.org/10.1007/s10103-021-03417-8 (2022).
-
Hang, N. L. T. et al. Photobiomodulation associated with alginate-based engineered tissue on promoting chondrocytes-derived biological responses for cartilage regeneration. Int. J. Biol. Macromol. 280, 135982. https://doi.org/10.1016/j.ijbiomac.2024.135982 (2024).
-
Athanasiou, K. A., Darling, E. M., Duraine, G., Hu, J. C., Reddi, A. H., Structure and Function of Cartilage. In: (Eds Athanasiou, K. A., Darling, E. M., Duraine, G., Hu, J.C., Reddi A. H.) Articulate Cartilage 3–79 (Taylor & Francis Group, 2017).
-
Prosser, A., Scotchford, C., Roberts, G., Grant, D. & Sottile, V. Integrated multi-assay culture model for stem cell chondrogenic differentiation. Int. J. Mol. Sci. https://doi.org/10.3390/ijms20040951 (2019).
-
Khajeh, S. et al. Enhanced chondrogenic differentiation of dental pulp-derived mesenchymal stem cells in 3D pellet culture system: Effect of mimicking hypoxia. Biologia (Bratisl). 73, 715–726. https://doi.org/10.2478/s11756-018-0080-z (2018).
-
Flores Luna, G. L. et al. Biphasic dose/response of photobiomodulation therapy on culture of human fibroblasts. Photobiomodulation Photomed. Laser Surg. 38(7), 413–418 (2020).
-
Dellagrana, R., Rossato, M., Sakugawa, R. L., Baroni, B. M. & Diefenthaeler, F. Photobiomodulation therapy on physiological and performance parameters during running tests: Dose-response effects. J. Strength Cond. Res. 32, 2807–2815 (2018).
-
Henao-Murillo, L., Pastrama, M. I., Ito, K. & van Donkelaar, C. C. The relationship between proteoglycan loss overloading-induced collagen damage, and cyclic loading in articular cartilage. Cartilage 13, 1501S-1512S. https://doi.org/10.1177/1947603519885005 (2021).
-
Inamdar, S. R., Prévost, S., Terrill, N. J., Knight, M. M. & Gupta, H. S. Reversible changes in the 3D collagen fibril architecture during cyclic loading of healthy and degraded cartilage. Acta Biomater. 136, 314–326. https://doi.org/10.1016/j.actbio.2021.09.037 (2021).
-
AbuBakr, N., Salem, Z., Ali, Z. & El Assaly, M. Comparative evaluation of the early effects of the low-level laser therapy versus intra-articular steroids on temporomandibular joint acute osteoarthritis in rats: A histochemical, molecular and imaging evaluation. Dent. Med. Probl. 55, 359–366. https://doi.org/10.17219/dmp/96290 (2018).
-
Darling, E. M. & Athanasiou, K. A. Rapid phenotypic changes in passaged articular chondrocyte subpopulations. J. Orthop. Res. 23, 425–432. https://doi.org/10.1016/j.orthres.2004.08.008 (2005).
-
Trevisan, E. S. et al. Effectiveness of led photobiomodulation therapy on treatment with knee osteoarthritis: A rat study. Am. J. Phys. Med. Rehabil. 99, 725–732. https://doi.org/10.1097/PHM.0000000000001408 (2020).
-
Yamada, E. F. et al. Photobiomodulation therapy in knee osteoarthritis reduces oxidative stress and inflammatory cytokines in rats. J. Biophotonics. 13, 1–14. https://doi.org/10.1002/jbio.201900204 (2020).
-
Stancker, T. G. et al. Can photobiomodulation associated with implantation of mesenchymal adipose-derived stem cells attenuate the expression of MMPs and decrease degradation of type II collagen in an experimental model of osteoarthritis?. Lasers Med. Sci. 33, 1073–1084. https://doi.org/10.1007/s10103-018-2466-0 (2018).
-
Sanches, M. et al. Chondroitin sulfate and glucosamine sulfate associated to photobiomodulation prevents degenerative morphological changes in an experimental model of osteoarthritis in rats. Lasers Med. Sci. 33, 549–557. https://doi.org/10.1007/s10103-017-2401-9 (2018).
-
Tomazoni, S. S. et al. Effects of photobiomodulation therapy, pharmacological therapy, and physical exercise as single and/or combined treatment on the inflammatory response induced by experimental osteoarthritis. Lasers Med. Sci. 32, 101–108. https://doi.org/10.1007/s10103-016-2091-8 (2017).
-
Tomazoni, S. S. et al. Isolated and combined effects of photobiomodulation therapy, topical nonsteroidal anti-inflammatory drugs, and physical activity in the treatment of osteoarthritis induced by papain. J. Biomed. Opt. 21, 108001. https://doi.org/10.1117/1.jbo.21.10.108001 (2016).
-
Angelova, A. & Ilieva, E. M. Effectiveness of high intensity laser therapy for reduction of pain in knee osteoarthritis. Pain Res. Manag. 2016, 1–11. https://doi.org/10.1155/2016/9163618 (2016).
-
Fukuda, T. Y. et al. Pulsed shortwave treatment in women with knee osteoarthritis: A multicenter, randomized, placebo-controlled clinical trial. Phys. Ther. 91, 1009–1017. https://doi.org/10.2522/ptj.20100306 (2011).
-
Hsieh, R. L., Lo, M. T., Liao, W. C. & Lee, W. C. Short-term effects of 890-nanometer radiation on pain, physical activity, and postural stability in patients with knee osteoarthritis: A double-blind, randomized, placebo-controlled study. Arch. Phys. Med. Rehabil. 93, 757–764. https://doi.org/10.1016/j.apmr.2012.01.003 (2012).
-
Vassão, P. G. et al. Level of pain, muscle strength and posture: Effects of PBM on an exercise program in women with knee osteoarthritis–a randomized controlled trial. Lasers Med. Sci. 35, 1967–1974. https://doi.org/10.1007/s10103-020-02989-1 (2020).
-
WALT: World Association for Photobiomodulation Therapy, Recommended treatment doses for Low Level Laser Therapy 780 – 860 nm wavelength, https://waltza.co.za/Wp-Content/Uploads/2012/08/Dose_table_780-860nm_for_Low_Level_Laser_Therapy_WALT-2010.Pdf. (2010) 2010. http://waltza.co.za/wp-content/uploads/2012/08/Dose_table_904nm_for_Low_Level_Laser_Therapy_WALT-2010.pdf%0Ahttps://waltza.co.za/wp-content/uploads/2012/08/Dose_table_780-860nm_for_Low_Level_Laser_Therapy_WALT-2010.pdf (accessed December 10, 2020).
-
WALT: World Association for Photobiomodulation, Recommended treatment doses for Low Level Laser Therapy 904 nm wavelength, https://Waltza.Co.Za/Wp-Content/Uploads/2012/08/Dose_table_904nm_for_Low_Level_Laser_Therapy_WALT-2010.Pdf. (2010) 2010. https://waltza.co.za/wp-content/uploads/2012/08/Dose_table_904nm_for_Low_Level_Laser_Therapy_WALT-2010.pdf (accessed December 10, 2020).
Acknowledgements
This research was supported by the national funds of Portuguese Foundation for Science and Technology (FCT) under the scope of the UID/4436/2025 project; by LABBELS–Associate Laboratory in Biotechnology, Bioengineering and Microelectromechanical Systems, LA/P/0029/2020; by the research projects Stimcart—PTDC/EME-EME/4520/2021 (https://doi.org/10.54499/PTDC/EME-EME/4520/2021), and BrainStimMap-PTDC/EME-EME/1681/2021 (https://doi.org/10.54499/PTDC/EME-EME/1681/2021); by the European Regional Development Fund through the Operational Competitiveness Program and the National Foundation for Science and Technology of Portugal (FCT) under the projects UID/CTM/00264/2020 of Centre for Textile Science and Technology (2C2T) on its components Base (https://doi.org/10.54499/UIDB/00264/2020) and programmatic (https://doi.org/10.54499/UIDP/00264/2020); and by the PhD fellowship grants UI/BD/150951/2021 and UI/BD/09375/2020 provided to Sofia Oliveira and Francisca Monteiro, respectively. Débora Ferreira also acknowledge FCT for the for Scientific Employment Stimulus (https://doi.org/10.54499/2022.01681.CEE CIND/CP1718/CT0013)
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Oliveira, S., Ferreira, D., Padrão, J. et al. Near-infrared photobiomodulation stimulates viability and cartilage matrix synthesis in human chondrocytes. Sci Rep 15, 44885 (2025). https://doi.org/10.1038/s41598-025-28792-y
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-28792-y