Introduction
The growing recognition of the health and environmental burdens of fossil-derived energy carriers has intensified the search for bio-based replacements within a biorefinery framework. Among these, biodiesel and biolubricants are prominent: life-cycle assessments report 40–86% lower GHG emissions for biodiesel than petroleum diesel (feedstock- and pathway-dependent), and biolubricants mitigate the ecological risks associated with mineral-oil releases1,2,3,4. Biolubricants are becoming more popular than petroleum-based lubricants because the latter can pollute up to one million liters of fresh water for every kilogram released5,6. Recent studies increasingly couple microbial conversion of carbon-rich industrial wastewaters with the co-production of biodiesel and related bio-products, placing process efficiency and feedstock selection at the center of current research7,8.
Arthrospira (spirulina) platensis is a filamentous cyanobacterium (formerly known as blue-green algae, where microalgae are a broad group of microbes having photosynthetic activity classified as cyanobacteria and true algae) containing proteins (50.0–71.3% DCW), carbohydrates (16.0–20.0% DCW), and fats (0.9–14.2% DCW)9. Arthrospira demonstrates significant COD removal capacity while simultaneously synthesizing valuable carotenoids. Across real wastewaters, Spirulina removed 93.1% of COD in piggery effluent at 25% strength (v/v), with TN, TP, Cu, and Zn removal efficiencies of 76%, 72%, 82.5%, and 93.5%, respectively, and a biomass yield of 0.56 g L⁻¹ in 8 d10. It achieved 62.59 ± 1.88% and 46.68 ± 1.40% COD removal in two different textile effluents within 5 d11, and in synthetic mariculture reduced 86.5% of TN, and 98.1% of TP with a biomass yield of 3.5 g L⁻¹12. Under 35 °C and 100 µmol photons m⁻² s⁻¹, A. platensis C1 accumulated carotenoids up to 3.64 mg L⁻¹ (4.66 mg g⁻¹ DW) at 96 h13. Although microalgae and cyanobacteria share several morphological and physiological characteristics, microalgae are eukaryotic organisms with membrane-bound organelles, such as a true nucleus and chloroplasts; in contrast, cyanobacteria are prokaryotic photoautotrophs without a nucleus and chloroplasts14.
The red oleaginous Rhodosporidium babjevae, which accumulates lipids up to 64.83% DCW on wheat straw hydrolysate after 96 h15 and carotenoids up to 43.1 mg g-1 at 96 h in N-limited, glycerol-fed medium (glycerol as carbon source)16, is one of the most promising species for biodiesel, biolubricant, and carotenoids production17,18.
Global paper and board production exceeds 400 Mt yr⁻¹, producing 4.3 to 40 kg of dry pulp-and-paper mill sludge (PPMS) per tonne of product; PPMS typically contains 40–50 wt% organic matter19. Conventional disposal methods, landfilling, incineration, and pyrolysis, pose serious environmental problems, especially for high-moisture organic waste. With its high cellulose content (up to 68% DW) and low lignin fraction (7.6–26% DW), primary PPMS (collected from the primary clarifier, 70% of the total dry PPMS produced) represents an ideal substrate for hydrolysis into fermentable sugars (e.g., glucose, arabinose, xylose), enabling microbial conversion into biofuels and biolubricants while offering an economic advantage of $25–80 per wet ton compared with disposal20,21,22,23,24.
Pretreatment is essential before hydrolysis to disrupt fiber structure, solubilize hemicellulose, and expose cellulose for saccharification; enzymatic routes then give high selectivity and fermentability, while dilute-acid routes are faster but create inhibitors (furfural, 5-HMF, acetic acid, and soluble phenolics) that must be managed. However, dilute-acid hydrolysis for lignocellulosic biomass generally shows low sugar yield efficiency. Therefore, combining dilute-acid hydrolysis with subsequent enzymatic hydrolysis significantly enhances the release of reducing sugars25. For PPMS or closely similar PPMS systems, central composite design (CCD) has been commonly used for dilute-acid hydrolysis optimization26,27, while Box–Behnken design (BBD) has been frequently applied to enzymatic saccharification and to downstream SSF or cultivation parameters for producing bioethanol or other bioproducts28,29,30.
Under continuous illumination and aeration, A. platensis operates predominantly photoautotrophically, fixing inorganic carbon (CO₂/HCO₃⁻). The soluble organics in PPMS hydrolysate are therefore consumed primarily by R. babjevae. In this co-culture, O₂–CO₂ exchange underpins a division of labor: A. platensis supplies photosynthetic O₂, elevates and stabilizes alkaline pH via bicarbonate uptake, and assimilates inorganic N and P, whereas R. babjevae converts dissolved organics into storage lipids, thereby lowering the soluble organic load (and potentially mitigating inhibition by organics)31,32,33.
This study evaluates a co-culture of A. platensis and R. babjevae grown on PPMS hydrolysate for lipid biosynthesis, enabling downstream biodiesel and biolubricant production. Addressing the inherently high COD of PPMS, the approach couples value creation with COD removal, thereby improving the environmental profile of this waste stream. Process performance was optimized and statistically validated via RSM (CCD and BBD method), and carotenoids were quantified as a valuable bio-based co-product. To our knowledge, this is the first report to couple a photoautotrophic cyanobacterium with a red oleaginous yeast for PPMS management in an environmental science and engineering context.
Materials and methods
PPMS hydrolysate preparation
The PPMS used in this study was taken from the wood and paper industries in Mazandaran, Iran, which are mainly used for printing and newspaper manufacturing. The raw sludge had a total solid content of about 10% (v/v) and was not biologically pretreated. The main PPMS sample analyzed contained 67.7% cellulose, 16.3% hemicellulose, and 4.5% lignin, according to the standard method described by Sluiter et al., and expressed on oven-dry weight (OD) basis. In addition, the ash content was 6.4% by the ISO 2144 standard method34,35.
Two microbial strains were used: Arthrospira platensis Gomont 1892 (SM; IBRC-M 50177) and Rhodosporidium babjevae Golubev 1993 (A-349; IBRC-M 30088), both obtained from the national center for genetic and biological resources of Iran36.
A commercial type of cellulase enzyme (CAS 9012-54-8, Sigma-Aldrich, Merck KGaA, Germany), derived from Trichoderma reesei, was utilized for the enzymatic hydrolysis of acid-treated PPMS (see below for the details).
The PPMS sample and PPMS hydrolysate were analyzed using the Fourier transform infrared spectroscopy (FTIR) method (Fig. S6.1 as supplementary information). FTIR analysis was conducted using a spectrometer (TENSOR 27, Bruker Optics, Germany) equipped with a DTGS detector. The measurements were performed in transmission mode in the mid-infrared (mid-IR) region, in the wavenumber range of 4000–400 cm⁻¹.
Therefore, a two-step hydrolysis method, acid followed by enzyme, was used. The parameters of dilute-acid hydrolysis were optimized using a CCD based on RSM with factor levels selected from the literature37,38,39,40. Further details on the method description, the design matrix, and the statistical analysis can be found in sections S1 and S2 of the supplementary information (SI). According to Tables S2.6, S2.7, and Fig. S2.2, the optimum conditions were 0.746% (w/w) H₂SO₄, 0.283% (w/w) HNO₃, 134.8 °C for 23.6 min, and pH adjusted to 5 using sodium citrate buffer before enzymatic hydrolysis.
The enzymatic hydrolysis was conducted using 2% (w/w) cellulase at 50 °C for 72 h in a shaking incubator (150 rpm; SHKE4000-6CE, Thermo Fisher Scientific, USA), with polyethylene glycol (1% w/w) added to all samples to improve enzymatic accessibility. Preliminary experiments examined enzyme concentrations (1–3% w/w) and hydrolysis duration (1–5 days), with the design guided by earlier studies41,42,43,44,45.
The concentration of reducing sugars was determined by the 3,5-Dinitrosalicylic acid (DNS) method46, measuring absorbance at 540 nm by UV-Vis spectrophotometer (UV-1800, Shimadzu, Japan). After hydrolysis, the suspension was filtered through a laboratory test sieve (200 mm diameter, 2.0 mm mesh aperture, stainless steel; Endecotts, Netherlands), to give the hydrolysates of PPMS (HPPMS), which were used as the growth medium in co-culture and monoculture experiments.
Procedures and protocols for the growth of microbes
R. babjevae and A. platensis were grown and gradually adapted to HPPMS as a growth medium. A. platensis was cultured in Zarrouk medium, and R. babjevae was cultured in YM medium. Later, both were adapted to HPPMS by successive subculturing with increasing amounts of HPPMS. Details of the media composition, light and dark cycles, aeration systems, centrifugation conditions, storage conditions, and the acclimatization procedure are given in section S3 of SI.
Co-cultivation procedure
Powdered inocula of R. babjevae were introduced into HPPMS medium together with A. platensis, and the two organisms were co-cultured in a single system to test whether co-cultivation improved lipid production and COD removal compared to the respective monocultures. Because co-cultivation is a natural multivariate process, we pre-determined critical process variables for optimization under continuous light and aeration. A systematic experimental design was then employed for quantitative assessment of the impact of the factors and identification of the conditions under which maximum lipid yield and COD removal efficiency are obtained.
The influential variables on the co-cultivation process were identified, and their ranges were established for the experimental design with BBD in the framework of the RSM. The characteristics of variables, experimental matrix, and statistical analysis can be found in SI (section S5.2). According to Table S5.2, each variable of the BBD matrix has three levels chosen after reviewing some past studies31,47,48,49,50. The co-cultivation tests were performed in 1-L graduated cylinders that were foil-covered and thoroughly cleaned to minimize the risk of contamination. For each trial, R. babjevae and A. platensis were added to the growth medium at the ratios specified by the design of experiments (DoE). The HPPMS medium was well aerated (pumping and air stones), and the microbes were well mixed. The air flow rate influencing the mixing effectiveness was carefully controlled by a Digital Flowmeter (M-5SLPM-D, Alicat Scientific, USA) and varied by adjusting the pump speed. A light intensity control system with a fluorescent lamp (TL-D 36 W/965, Philips, Netherlands) was used. The intensity was varied by changing the distance between cylinders and the light source, and the exact light intensity for each condition was measured by a luxmeter (LM 76, Multimetrix-Chauvin Arnoux Group, France). The co-cultivation time was selected according to DoE to vary the length of co-cultivation in each experiment. After each experimental run, biomass yield, lipid yield, and COD removal efficiency were measured to evaluate the effectiveness of the conditions.
Lipid and carotenoid extraction
Carotenoid extraction was performed following a modified DMSO-acetone dry extraction method according to Cheng and Yang51 and Michelon et al.52, and lipid extraction was performed following the Bligh and Dyer method53, using wet biomass and a Soxhlet apparatus. Additional details of the procedures, such as reagent volumes employed, centrifugation conditions, and equipment settings, can be found in the SI (section S4).
Dry cell weight measurement
The dry cell weight (DCW) of microbial cultures was determined by first collecting a specified volume of the culture broth, which was then centrifuged (Sorvall Legend X1R, Thermo Fisher Scientific, USA) at 10,000 rpm for 15 min to separate the biomass. The pellet was washed twice with distilled water to remove residual medium, then transferred to a pre-weighed, anti-static, high-impact polystyrene weigh boat. The sample was placed in an oven (UN55, Memmert, Germany) set to 55 °C and left there for 24 h or until it had a constant weight. After cooling in a desiccator, the dried biomass was weighed using an analytical balance (AUW220D, Shimadzu Corporation, Japan).
Process description for biodiesel and biolubricant synthesis
Based on previously optimized conditions54,55,56, microbial oil was first subjected to esterification with methanol at an oil-to-methanol molar ratio of 1:5, using KOH (1 wt% of oil) as a homogeneous base catalyst. The reaction was conducted in a sealed glass reactor at 60 °C for 60 min under constant magnetic stirring (500 rpm; C-MAG HS 7, IKA-Werke, Germany) to enhance phase contact. Once the mixture was complete, a decanter was used, and the whole blend was allowed to sit and separate into phases for 24 h. The glycerol-rich lower phase was removed, and the upper FAME layer (biodiesel) was collected and stored under controlled conditions.
To improve the physicochemical characteristics and functionalize the FAME for biolubricant applications, a second transesterification step was conducted using neopentyl glycol (NPG) at a NPG-to-FAME molar ratio of 1:2, and sodium methoxide (1.5 wt%) served as the base catalyst. The reaction proceeded at 120 °C under atmospheric pressure for 190 min with continuous magnetic stirring (500 rpm; C-MAG HS 7, IKA-Werke, Germany). The reaction was shifted toward diester formation by continuously removing methanol from the mixture using a rotary evaporator (Rotavapor R-210, Buchi, Switzerland). The resulting biolubricant was purified through simple distillation at 65 °C for 30 min to remove residual methanol. The evaporation of the solvent was performed in a 500 mL round-bottom flask, filled with about 250 mL of the reaction mixture (half of the flask volume) to provide good evaporation under reduced pressure (300 mbar), followed by drying at 85 °C in a laboratory oven (UN55, Memmert, Germany). Anhydrous sodium sulfate was added to eliminate residual moisture57.
To improve performance, additives were incorporated to enhance oxidative stability, viscosity, and thermal resistance. In particular, poly(methyl methacrylate) (PMMA) (10 wt%) was used to control the viscosity, BHT (0.1 wt%) was added as an antioxidant, and TiO₂ nanoparticles (0.5 wt%) were chosen as an antioxidant and a thermal stabilizer. PMMA and BHT were dissolved in ethanol–water mixtures containing approximately 80% ethanol (w/w) and ethanol (10:1 solvent-to-additive ratio), respectively, and at 60 °C under magnetic stirring. TiO₂ nanoparticles were ultrasonically dispersed in tetrahydrofuran (THF) using an ultrasonic processor (Vibra-Cell VCX 750, Sonics & Materials, USA) to ensure uniform dispersion. To prevent agglomeration, TiO₂ nanoparticles were dispersed in THF and sonicated in pulsed mode (120 W; 10 s on/10 s off) for a total on-time of 25 min. These additive solutions were added into preheated NPG diesters (70 °C) and mixed for 60 min under magnetic stirring. Ethanol and THF were evaporated in a rotary evaporator with a water bath temperature of 50 ± 2 °C and a vacuum pressure of about 150 mbar, corresponding to the boiling range of these solvents under vacuum. The 250 mL batch was evaporated under reduced pressure for 35 min until the solvent was removed entirely. To prevent oxidation, the product was stored in amber glass bottles under cool, dry conditions. A schematic representation of the experimental procedures is illustrated in Fig. S4.1, summarizing the key steps involved in the laboratory workflow.
Analytical methods for biolubricant and biodiesel properties
The fatty acid composition was evaluated via gas chromatography (GC; 7890 A, Agilent Technologies, USA) equipped with a flame ionization detector (FID) and a split-splitless injector. Separation was achieved on Agilent J&W DB-23 capillary column (30 m long, 0.32 mm inner diameter, 0.25 mm film thickness) having a stationary phase of 50% cyanopropyl-methylpolysiloxane by using high-purity helium as the carrier gas at a flow rate of 1.8 mL min-1. The oven temperature was programmed from 75 °C (4 min hold) to 250 °C at 20 °C min-1, followed by a final hold at 250 °C for 4 min58. FAMEs were identified and quantified using a certified FAME Mix C8–C22 (CRM18920, Sigma-Aldrich, Merck KGaA, Germany).
Kinematic viscosity at 40 °C and 100 °C was obtained from dynamic viscosity and density measured by a Stabinger viscometer (SVM 3001, Anton Paar, Austria) in accordance with ASTM D7042, and reported as ν (mm² s⁻¹)59. Density at 15 °C was determined by oscillating U-tube densimetry per ASTM D4052 using an automatic density meter (K86201, Koehler, USA)60. Flash and fire points were measured by the Cleveland open-cup method following ASTM D92 with a COC tester (K72000 series, Koehler, USA)61. Low-temperature operability was characterized by pour point (ASTM D5949, automatic pressure-pulsing) using an automatic cloud-and-pour point analyzer (K77001, Koehler, USA)62. Oxidation stability of FAME was assessed by Rancimat according to EN 14112 (neat FAME), using a biodiesel Rancimat (893 Professional, Metrohm, Switzerland)63. Sulfated ash and acid number were determined by ASTM D874-23 and ASTM D974-22, respectively (muffle furnace and indicator titration)64,65. Iodine value (degree of unsaturation) was measured by the Wijs method66. Cold filter plugging point (CFPP) was measured according to EN 116 using a CFPP analyzer (FPP 5Gs, PAC-ISL, France)67. Environmental impact was assessed through COD analysis using the colorimetric method (5220D)68.
Data analysis
Lipid yield, biomass yield, and COD removal efficiency were statistically analyzed using Design Expert v12 software (Stat-Ease, Inc., USA). Graphical representations of data were generated using OriginPro 2024 (OriginLab Corporation, USA) and Design Expert v12.
Results and discussion
Optimization of co-cultivation conditions
A mathematical model was established to predict lipid yield, biomass yield, and COD removal efficiency in R. babjevae–A. platensis co-cultivation, with experimental validation confirming its accuracy; optimization prioritized maximizing lipid yield first, followed by COD removal efficiency, with predictive models and BBD results detailed in SI (section S5.2).
Model development for lipid yield prediction
Using Eq. S2.2, we fitted a reduced quadratic model to predict lipid yield. The final model, expressed in coded variables, treats lipid yield (Y) as the response.
Equation (1): lipid yield prediction model.
$$begin{aligned} {text{Y}}left( {{text{g}}/{text{L}}} right) & = {text{6}}.{text{31}} + 0.0{text{65}}0{text{X}}_{{text{1}}} + {text{1}}.{text{24X}}_{{text{2}}} + 0.{text{1817X}}_{{text{3}}} + 0.{text{195}}0{text{X}}_{{text{4}}} + 0.{text{3575}}left( {{text{X}}_{{text{1}}} times {text{X}}_{{text{3}}} } right) + 0.{text{46}}left( {{text{X}}_{{text{1}}} times {text{X}}_{{text{4}}} } right) \ & quad {-}0.{text{57}}left( {{text{X}}_{{text{3}}} times {text{X}}_{{text{4}}} } right){-}0.{text{4226 X}}_{{text{1}}} ^{{text{2}}} {-}{text{1}}.{text{32 X}}_{{text{2}}} ^{{text{2}}} {-}0.{text{1276 X}}_{{text{3}}} ^{{text{2}}} {-}0.{text{49}}0{text{1 X}}_{{text{4}}} ^{{text{2}}} \ end{aligned}$$
(1)
X₁, X₂, X₃, and X₄ denote the coded variables for the A. platensis to R. babjevae ratio, time, airflow rate, and light intensity (per Eq. S2.1), with the model incorporating main effects, interactions, and quadratic terms to capture linear and nonlinear impacts on lipid biosynthesis, and its validity confirmed by ANOVA (SI, section S5.2).
Model-predicted optimal conditions for lipid yield
The optimized co-cultivation parameters and responses, as presented in Table 1, were determined based on the predictive model described by Eq. (1) and the numerical outputs derived from the software (Fig. S5.1).
Experimental validation of the lipid-yield model at the optimum
To validate the optimized conditions, three independent co-cultivation experiments were performed. As shown in Table 2, observed lipid yields closely matched the model predictions, confirming the robustness of the optimization. Under these conditions, carotenoid content reached 41.22 ± 1.57 mg L⁻¹ as a valuable co-product (data represent mean ± standard deviation, n = 3).
Predictive accuracy of the lipid-yield model
The model showed high predictive accuracy, with average errors of only 0.83% for COD removal, 1.57% for biomass yield, and 3.30% for lipid yield, demonstrating its ability to reliably capture the interactions in the R. babjevae – A. platensis co-culture.
Effects of key factors on lipid yield
Based on Table S5.7, co-cultivation time had the most potent effect on lipid yield (F = 1217.69, p < 0.0001), followed by light intensity (F = 30.03, p < 0.0001) and airflow rate (F = 26.07, p < 0.0001), with significant interactions between light intensity and airflow rate (F = 85.54, p < 0.0001), and between A. platensis to R. babjevae ratio with airflow rate (F = 33.65, p < 0.0001) and light intensity (F = 55.71, p < 0.0001), highlighting strong parameter interdependencies.
In oleaginous yeasts such as R. babjevae and in photosynthetic cyanobacteria such as A. platensis, lipid biosynthesis begins with the ATP-dependent carboxylation of acetyl-CoA to malonyl-CoA by acetyl-CoA carboxylase (ACC), which channels carbon toward fatty acid synthesis in a nutrient- and redox-dependent manner. In R. babjevae, acyl chains are extended by the type I fatty acid synthase complex (FAS). By contrast, in A. platensis, fatty acid synthesis relies on the modular prokaryotic type II system, composed of AccA, AccB, AccC, and AccD together with FabD, FabH, FabG, FabZ, and FabI, which provides environmental responsiveness. In R. babjevae, triacylglycerol accumulates through the Kennedy pathway via glycerol-3-phosphate acyltransferase (GPAT), one-acylglycerol-3-phosphate acyltransferase (LPAAT), and diacylglycerol acyltransferase (DGAT), with an alternative phospholipid diacylglycerol acyltransferase (PDAT) route that remodels membrane phospholipids into storage lipids and can incorporate polyunsaturated acyls such as linoleate (C18:2); depending on species and conditions, additional unusual acyls including ricinoleate may be observed. Under oxidative stress or nutrient limitation, Δ12 and Δ15 desaturases introduce double bonds, further diversifying the lipid pool in R. babjevae69,70.
In A. platensis, neutral-lipid storage as triacylglycerol is minimal in native strains; carbon is preferentially stored as glycogen. De novo membrane glycerolipids arise via the prokaryotic PlsX–PlsY–PlsC route (PlsX generating acyl-phosphate from acyl-ACP, PlsY forming lysophosphatidic acid, and PlsC producing phosphatidic acid). Phosphatidic acid is dephosphorylated by a PAP-like activity to diacylglycerol that is predominantly directed to glycolipid synthesis. Substantial TAG accumulation is non-native and typically requires heterologous WS-DGAT with an acyl-CoA supply71,72,73; several earlier TAG reports were re-assigned to acyl-plastoquinol74.
The biosynthetic precursors acetyl-CoA, the reductant NADPH, and ATP that support lipid anabolism are furnished by central carbon metabolism. In R. babjevae, the oxidative pentose phosphate pathway is the primary source of NADPH; glucose-6-phosphate dehydrogenase (G6PDH) and 6-phosphogluconate dehydrogenase (6PGDH) generate NADPH to support lipogenesis and redox buffering. In A. platensis, light-driven electrons reduce NADP⁺ via ferredoxin–NADP⁺ reductase, supplying NADPH for the photosynthetic light reactions that power the Calvin–Benson–Bassham (CBB) cycle and fatty-acid synthesis. Under nitrogen deprivation, a canonical lipogenic cue, yeasts undergo carbon–nutrient decoupling marked by AMP depletion, activation of Snf1 as the AMPK homolog, and inhibition of mitochondrial isocitrate dehydrogenase, which together promote cytosolic citrate accumulation. ATP-citrate lyase cleaves citrate into acetyl-CoA and oxaloacetate, thereby providing building blocks for de novo fatty acid synthesis and promoting TAG storage69,75,76.
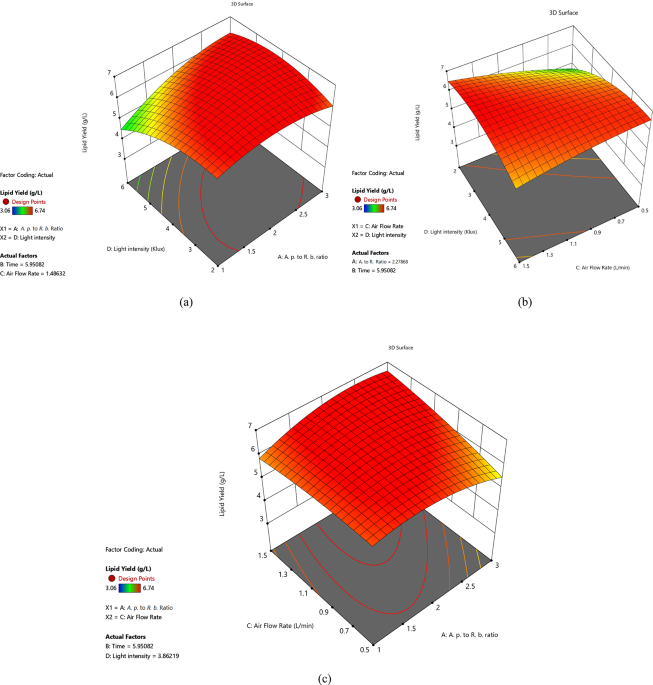
Based on the lipid-yield responses in Fig. 1b and c, aeration rates below around 1.3 L min⁻¹ were associated with reduced lipid accumulation, plausibly reflecting limited O₂ transfer that constrains respiration and ATP generation and promotes fermentative by-products (e.g., lactate and ethanol). In contrast, rates above about 1.48 L min⁻¹ coincided with lower lipid yield, consistent with shear-related damage to A. platensis membranes and photosystems, thereby impairing growth. An intermediate window of around 1.3–1.5 L min⁻¹ produced the highest lipid yields, likely balancing photosynthetic O₂ release for yeast respiration with CO₂ availability for the CBB cycle69,77.
From the lipid-yield responses in Fig. 1a and b, illumination at about 2.8–5.5 klux corresponded to higher lipid accumulation, plausibly reflecting faster CBB turnover, greater availability of triose-phosphate precursors such as G3P, and increased ATP and NADPH supply. At intensities above around 5.5 klux, lipid yield declined, consistent with the onset of photoinhibition, reactive-oxygen-species formation, and pigment loss; the decrease is also evident from reductions in effective quantum yield69,78,79.

3D plot and contour plot of the BBD illustrating the effects of experimental factors on lipid yield: (a) A. platensis to R. babjevae ratio versus light intensity, (b) light intensity versus airflow rate, and (c) A. platensis to R. babjevae ratio versus airflow rate.
From Fig. 1a and c, lipid yield peaks near an A. platensis to R. babjevae biomass ratio of about 2.2:1. This balance likely optimizes syntrophy, A. platensis supplies O₂ (mitigating yeast oxidative stress) and fixes yeast-derived CO₂, whereas off-ratio conditions disrupt exchange and depress lipid accumulation69,80.
Consistent with the trends in Table 3, our optimal conditions, 3.86 klux illumination, 1.48 L min⁻¹ aeration, an A. platensis to R. babjevae ratio of ≈ 2.27, and 5.95 d, fall within a moderate, sub-photoinhibitory light range and a mild aeration regime. Under these conditions, lipid yield reached 6.56 ± 0.22 g L⁻¹ (57.64% DCW), exceeding values reported for comparable co-cultures and monocultures2,81,82,83.
Comparison of co-cultivation and monoculture conditions
According to Fig. 2a and b (the data presented in Fig. 2a and b are provided in Tables S5.11 and S5.12), lipid yields exhibit substantial rises in the co-culture under optimal conditions, with noted augmentations of 31.04% for R. babjevae and for A. platensis after 143 h, relative to the total lipid yield of both strains combined and with a biomass reduction of 9.10% after 143 h, relative to the total biomass yield of both strains combined. Across the 7-day period, peak lipid contents (wt% of DCW) were 48.15% for R. babjevae, 10.41% for A. platensis, and 57.64% for the co-culture.

Comparison of co-cultivation and monoculture under optimal conditions (light intensity: 3.86 klux; A. platensis to R. babjevae ratio: 2.27:1 (for co-cultivation); airflow rate: 1.48 L min−1); (a) lipid yield versus time, (b) biomass yield versus time.
The observed improvements arise from metabolite and gas exchange in co-culture. A. platensis supplies O₂ that boosts R. babjevae respiration, increasing TCA flux, citrate export, and cytosolic acetyl-CoA formation via ATP-citrate lyase; dissolved organics and extracellular carbohydrates are assimilated by the yeast to add acetyl-CoA and reducing power via the pentose phosphate pathway and malic enzyme. This elevates flux through acetyl-CoA carboxylase, fatty acid synthase, and diacylglycerol acyltransferase, favoring TAG formation in the yeast, whereas the mevalonate pathway chiefly supports isoprenoid and carotenoid synthesis. Reciprocally, CO₂ released by R. babjevae drives the CBB cycle in A. platensis, channeling carbon mainly to biomass, glycogen, and membrane lipids in native strains (neutral TAG remains limited). In parallel, nutrient signaling and ROS-driven stress responses promote neutral-lipid accumulation (protective in A. platensis), while competition, shading, and nutrient limitation reduce biomass by diverting carbon from growth to lipogenesis69,70,71,72,73,75,76.
A hypothetical pathway for enzymatic TAG synthesis is shown in Fig. S7.1, based on the co-culture performance of the two microbial strains.
FAME profile and biodiesel property predictions
The fatty-acid composition of co-culture lipids (expressed as FAMEs by GC-FID) is summarized in Table 4. The GC-FID chromatogram is provided in Fig. S7.2.
Under optimal co-culture conditions, the FAME profile totaled ΣSFA 43.16% (C14:0 0.05%, C16:0 0.80%, C18:0 42.08%, C20:0 0.23%), ΣMUFA 47.66% (C16:1 13.05%, C18:1 34.13%, C20:1 0.48%), and ΣPUFA 9.17% (C18:2 6.08%, C18:3 3.09%). Relative to R. babjevae grown on pure glycerol (normalized to 100%), the co-culture exhibits strong stearate enrichment (+ 34.63% points (pp) to 42.08%), marked palmitate depletion (− 24.71 pp to 0.80%), modestly lower oleate (− 6.26 pp to 34.13%), and reduced PUFA (− 13.15 pp). The combination of high C18:0 with substantial MUFA is expected to increase cetane number and oxidative stability; nonetheless, cold-flow and viscosity should be verified via predictive correlations or direct measurements. Mechanistically, the pattern is consistent with a relative limitation of stearoyl-CoA desaturase (Δ9; C18:0→C18:1), altered elongation–desaturation balance, and oxygen and redox constraints, while the high C16:1 with minimal C16:0 suggests active Δ9 desaturation on C16 substrates69,89,90.
Compared with A. platensis monoculture lipids (C16:0 42.85%, C18:0 1.00%, C18:1 3.53%, C18:2 14.43%, γ-C18:3 22.45%, α-C18:3 0.47%; ΣSFA 50.1%, ΣMUFA 8.5%, ΣPUFA 40.6%), the co-culture shifts toward a stearate- and MUFA-weighted signature (C18:0 42.08%, ΣMUFA 47.7%) with minimal palmitate (0.80%) and depressed PUFA (9.17%). Notably, total C18:3 in A. platensis (≈ 22.9%, α and γ) exceeds the EN 14,214 limit of 12% m/m, whereas the co-culture remains compliant at 3.09%91,92,93.
Physicochemical characterization of the produced biodiesel and biolubricant
The physicochemical properties of biodiesel made from the optimized co-culture medium are shown in Table 5. All major requirements for biodiesel, such as viscosity (4.8 mm² s− 1), iodine value (101 g I₂ 100 g− 1), and oxidation stability (8.62 h), meet the specifications set out by EN 14214, showing that this product is suitable as a biodiesel93.
According to Table 6, the formulated biolubricant is comparable to Shell S3 PY 22094. The NPG diester matrix underpins the formulation’s thermal and oxidative robustness and rheological stability; recent studies report viscosity index (VI) = 119.21–190, flash point 235–300 °C, pour point − 15 to + 10 °C (which can be lowered to − 48 °C with a pour point depressant (PPD)), kinematic viscosity 21–191.68 mm² s⁻¹ at 40 °C and 6.6–19.82 mm² s⁻¹ at 100 °C95,96,97.
As a viscosity-index improver (VII), PMMA reduces the temperature dependence of viscosity, mitigating thermal thinning and helping to maintain film-forming viscosity across the operating temperature range; the magnitude of this effect depends on polymer architecture, treat rate, and the formulation’s shear stability98,99. BHT provides chain-breaking radical scavenging, which mechanistically explains the reduced acid number (0.5 mg KOH g⁻¹ vs. 0.6) and delayed peroxide and acid formation, outcomes broadly corroborated in biodiesel and biolubricant oxidation reviews and additization studies100. Beyond their tribochemical role, dispersed TiO₂ nanoparticles exhibit catalytic antioxidant activity that enhances oxidative stability and reduces the formation of peroxides and acids101,102.
Together, these mechanisms justify the Table 6 properties of the formulated biolubricant and support its comparability to Shell S3 PY 220.
Conclusions
Findings of the present study show that joint performances of A. platensis and R. babjevae co-cultures were successful in the context of biodiesel and bio-lubricant production using the PPMS hydrolysate as a type of renewable lignocellulosic feedstock. The synergistic interplay between R. babjevae (organic utilization, COD reduction, CO₂ release) and A. platensis (O₂ and NADPH supply, nutrient recycling) at optimized conditions yielded 6.56 ± 0.22 g L⁻¹ lipids (57.64 ± 1.0% DCW) with 86.36 ± 0.97% COD removal. After 143 h, lipid output in the co-culture increased by 31.04% compared with the combined lipid production of the two monocultures, with maximum lipid accumulation reaching 48.15% and 10.41% DCW for R. babjevae and A. platensis, respectively, versus 57.64% DCW in the co-culture. Fatty acid profiles were dominated by yeast-derived lipids, resulting in biodiesel that met EN 14214 standards (ν₄₀ = 4.8 mm² s⁻¹, iodine value 101 g I₂ 100 g⁻¹, oxidation stability 8.62 h). Further transestrification with NPG and additives (PMMA, BHT, TiO₂) produced a biolubricant matching or exceeding Shell S3 PY 220, with ν₄₀ = 223 mm² s⁻¹, ν₁₀₀ = 20.2 mm² s⁻¹, pour point − 11 °C, flash point 243 °C, and acid number 0.5 mg KOH g⁻¹.
Future prospective
Further research should clarify the molecular pathways expressed during co-cultivation, as the results are promising. Residual biomass, still showing polluting traits after lipid and carotenoid extraction, can be valorized, for instance, in bio-electrochemical systems where microorganisms convert biochemical energy into bioelectricity (e.g., wetland MFC units). Moreover, applying TOPSIS could help rank and evaluate R. babjevae cultures for carotenoid biosynthesis, both with and without A. platensis, while growth kinetics may be statistically quantified to support process optimization.
Data availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
References
-
Xu, H., Ou, L., Li, Y., Hawkins, T. R. & Wang, M. Life cycle greenhouse gas emissions of biodiesel and renewable diesel production in the united States. Environ. Sci. Technol. 56, 7512–7521 (2022).
-
Mohamed, H. et al. Coculturing of Mucor plumbeus and Bacillus subtilis as an efficient fermentation strategy to enhance fungal lipid and γ-linolenic acid (GLA) production. Sci. Rep. 12, 1 (2022).
-
Nguyen, D. T. et al. Microalgae-derived biolubricants: Challenges and opportunities. Sci. Total Environ. 176759 (2024). (2024).
-
Mhatre, N. et al. Microbial-based biolubricants: in-depth analysis. In Multifunctional Materials: Engineering and Biological Applications 133–180Wiley, (2025).
-
Malik, M. A. I., Kalam, M. A., Mujtaba, M. A. & Almomani, F. A review of recent advances in the synthesis of environmentally friendly, sustainable, and nontoxic biolubricants: recommendations for future implementations. Environ. Technol. Innov. 32, 103366 (2023).
-
Barthlott, W. et al. Adsorption and superficial transport of oil on biological and bionic superhydrophobic surfaces: A novel technique for oil–water separation. Philos. Trans. R Soc. A. 378, 2167 (2020).
-
Gopalakrishnan, K., Wager, Y. Z. & Roostaei, J. Co-cultivation of microalgae and bacteria for optimal bioenergy feedstock production in wastewater using response surface methodology. Sci. Rep. 14, 20703 (2024).
-
Jayabal, R. et al. Study of the effects of bio-silica nanoparticle additives on the performance, combustion, and emission characteristics of biodiesel produced from waste fat. Sci. Rep. 13, 18907 (2023).
-
Chwil, M. et al. Comprehensive review of the latest investigations of the health-enhancing effects of selected properties of Arthrospira and Spirulina microalgae on skin. Pharmaceuticals 17, 1321 (2024).
-
Liang, C. et al. Cultivation of Spirulina platensis for nutrient removal from piggery wastewater. Environ. Sci. Pollut Res. 30, 85733–85745 (2023).
-
Selvaraj, D. & Arivazhagan, M. Synergistic effects of Spirulina platensis cultivation in textile wastewater towards nutrient removal and seed germination. Environ. Pollut. 357, 124435 (2024).
-
Fu, X. et al. Nutrient removal from mariculture wastewater by algal–bacterial aggregates developed from Spirulina platensis. Water 15, 396 (2023).
-
Mohammed, I. A., Ruengjitchatchawalya, M. & Paithoonrangsarid, K. Cultivation manipulating zeaxanthin-carotenoid production in Arthrospira (Spirulina) platensis under light and temperature stress. Algal Res. 76, 103315 (2023).
-
Sreeharsha, R. V. & Venkata Mohan, S. Photosynthetic microbes: Evolution, classification, and structural physiology. In Microbial Photosynthesis: from Basic Biology To Artificial Cell Factories and Industrial Applications 3–22 (Springer, Singapore, (2024).
-
Brandenburg, J. et al. Oleaginous yeasts respond differently to carbon sources present in lignocellulose hydrolysate. Biotechnol. Biofuels. 14, 124 (2021).
-
Tahmasebi, Z., Zilouei, H. & Kot, A. M. Investigating the concomitant production of carotenoids and lipids by the yeast Rhodosporidium Babjevae using sugarcane Bagasse hydrolysate or corn steep liquor. Renew. Energy. 228, 120618 (2024).
-
Sánchez-Solís, A., Lobato-Calleros, O., Moreno-Terrazas, R. & Lappe-Oliveras, P. Neri-Torres, E. Biodiesel production processes with yeast: A sustainable approach. Energies 17, 302 (2024).
-
Žganjar, M. et al. High-throughput screening of non-conventional yeasts for conversion of organic waste to microbial oils via carboxylate platform. Sci. Rep. 14, 14233 (2024).
-
Turner, T., Wheeler, R. & Oliver, I. W. Evaluating land application of pulp and paper mill sludge: A review. J. Environ. Manag. 317, 115439 (2022).
-
Shi, S., Kang, L. & Lee, Y. Y. Production of lactic acid from the mixture of softwood pre-hydrolysate and paper mill sludge by simultaneous saccharification and fermentation. Appl. Biochem. Biotechnol. 175, 2741–2754 (2015).
-
Ridout, A. et al. G. É. N. E. Valorisation of paper waste sludge via fermentation and pyrolysis. Cellul. Chem. Technol. 50, 439–442 (2016).
-
Naicker, J. E., Govinden, R., Lekha, P. & Sithole, B. Transformation of pulp and paper mill sludge (PPMS) into a glucose-rich hydrolysate using green chemistry: assessing pretreatment methods for enhanced hydrolysis. J. Environ. Manag. 270, 110914 (2020).
-
Kumar, V. et al. P. A critical review on biofuels generation from pulp-paper mill sludge with emphasis on pretreatment methods: renewable energy for environmental sustainability. BMC Environ. Sci. 2, 2 (2025).
-
Quintana, E., Valls, C. & Roncero, M. B. Valorization of waste paper sludge as a sustainable source for packaging applications. Polym. Bull. 81, 9321–9345 (2024).
-
Oladzad, S. et al. Comparison of acid and hydrothermal pretreatments of date waste for value creation. Sci. Rep. 14, 18056 (2024).
-
Kumar, R., Mondal, P., Sadhukhan, A. K. & Ganguly, A. Optimization of process parameters for production of bio-ethanol from waste potato following staged ultrasonic-assisted acid hydrolysis and saccharification. Biomass Convers. Biorefin 1–13 (2025).
-
Kamzon, M. A., Abderafi, S. & Bounahmidi, T. Multi-objective optimization of the first-stage dilute sulfuric acid hydrolysis of Moroccan beet pulp. Biomass Convers. Biorefin. 13, 4075–4091 (2023).
-
Hu, F., Fang, Y., Khan, Z. & Xing, L. Optimization of triterpenoids biosynthesis in Athelia termitophila as a source of natural products. AMB Express. 15, 61 (2025).
-
Saleh, A. K. et al. Box–Behnken design for the optimization of bioethanol production from rice straw and sugarcane Bagasse by newly isolated Pichia occidentalis strain AS.2. Energy Environ. 33, 1613–1635 (2022).
-
Zambare, V., Jacob, S., Din, M. F. M. & Ponraj, M. Box–Behnken design-based optimization of the saccharification of primary paper-mill sludge as a renewable Raw material for bioethanol production. Sustainability 15, 10740 (2023).
-
Prasetyo, A. et al. Composition and essential nutrients of Cyanobacterium meal produced from spirulina (Arthrospira platensis) fermented by yeast on small-scale. Case Stud. Chem. Environ. Eng. 10, 100940 (2024).
-
Naseema Rasheed, R. et al. Microalgal co-cultivation—recent methods, trends in omic-studies, applications, and future challenges. Front. Bioeng. Biotechnol. 11, 1193424 (2023).
-
Jung, C. G. H. et al. Decarbonization of Arthrospira platensis production by using atmospheric CO₂ as an exclusive carbon source: proof of principle. Int. J. Environ. Sci. Technol. 21, 4635–4644 (2024).
-
Sluiter, A. et al. Determination of structural carbohydrates and lignin in biomass. Lab. Anal. Proced. 1617, 1–16 (2008).
-
International Organization for Standardization. Paper, board, pulps and cellulose nanomaterials—Determination of residue (ash content) on ignition at 900°C (ISO 2144:2019). ISO. Available at: (2019). https://cdn.standards.iteh.ai/samples/73415/edacc5725416400e95aea4d68a0637d4/ISO-2144-2019.pdf
-
National Center for Genetic and Biological Resources of Iran (NCGBRI). Biological Strain Catalog (Agricultural Biotechnology Research Institute of Iran (ABRII), 2024).
-
Karapınar, İ., Kargı, F. & Öztekin, R. Effects of operating parameters on acid hydrolysis of ground wheat starch: maximization of the sugar yield by statistical experiment design. Starch-Stärke 63, 311–318 (2011).
-
Kartini, E., Kasi, H. & Abdullah, N. Fermentable sugar via diluted acid hydrolysis of sugarcane Bagasse. Key Eng. Mater. 908, 435–440 (2022).
-
Moutta, R. et al. Statistical optimization of sugarcane leaves hydrolysis into simple sugars by dilute sulfuric acid catalyzed process. Sugar Tech. 14, 53–60 (2011).
-
Tesfaw, A. & Tizazu, B. Reducing sugar production from teff straw biomass using dilute sulfuric acid hydrolysis: characterization and optimization using response surface methodology. Int. J. Biomater. 1–13 (2021). (2021).
-
Mushtaq, Q., Ishtiaq, U., Joly, N., Martin, P. & Qazi, J. Investigation and characterization of changes in potato peels by thermochemical acidic pre-treatment for extraction of various compounds. Sci. Rep. 14, 63364 (2024).
-
Mao, B., Lu, W. & Huang, G. Ultrasound-assisted enzymatic extraction, process optimization, and antioxidant activity of polysaccharides from sugarcane Peel. Sci. Rep. 15, 5009 (2025).
-
Silvia, S. & Maharani, A. The effect of enzyme concentration and hydrolysis time on the yield of microcrystalline cellulose from rice straw. DHPS 1, 25–32 (2023).
-
Zambare, V. P. & Christopher, L. P. Integrated biorefinery approach to utilization of pulp and paper mill sludge for value-added products. J. Clean. Prod. 274, 122791 (2020).
-
Sheng, Y. et al. Effect of ascorbic acid assisted dilute acid pretreatment on lignin removal and enzyme digestibility of agricultural residues. Renew. Energy. 163, 732–739 (2021).
-
Du, R. et al. In situ assay of the reducing sugars in hydrophilic natural deep eutectic solvents by a modified DNS method. J. Mol. Liq. 385, 122286 (2023).
-
Karitani, Y., Yamada, R., Matsumoto, T. & Ogino, H. UV mutagenesis improves growth potential of green algae in a green algae–yeast co-culture system. Arch. Microbiol. 206, 61 (2024).
-
Tejido-Núñez, Y., Aymerich, E., Sancho, L. & Refardt, D. Co-cultivation of microalgae in aquaculture water: interactions, growth and nutrient removal efficiency at laboratory- and pilot-scale. Algal Res. 49, 101940 (2020).
-
Goldoost, H., Vahabzadeh, F. & Fallah, N. Lipid productivity of Cyanobacterium Anabaena vaginicola in an internally illuminated photobioreactor using LED bar lights. Sci. Rep. 14, 6857 (2024).
-
Thangsiri, S. et al. Bioactive compounds and in vitro biological properties of Arthrospira platensis and Arthrospira maxima: a comparative study. Sci. Rep. 14, 23786 (2024).
-
Cheng, Y. T. & Yang, C. F. Using strain Rhodotorula mucilaginosa to produce carotenoids from food wastes. J. Taiwan. Inst. Chem. Eng. 61, 270–275 (2016).
-
Michelon, M., de Matos de Borba, T., da Silva Rafael, R. & Burkert, C. A. V. De Medeiros Burkert, J. F. Extraction of carotenoids from Phaffia rhodozyma: a comparison between different techniques of cell disruption. Food Sci. Biotechnol. 21, 1–8 (2012).
-
Bligh, E. G. & Dyer, W. J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917 (1959).
-
Erchamo, Y. S., Mamo, T. T., Workneh, G. A. & Mekonnen, Y. S. Improved biodiesel production from waste cooking oil with mixed methanol–ethanol using enhanced eggshell-derived CaO nanocatalyst. Sci. Rep. 11, 6708 (2021).
-
Weldeslase, M. G., Benti, N. E., Desta, M. A. & Mekonnen, Y. S. Maximizing biodiesel production from waste cooking oil with lime-based zinc-doped CaO using response surface methodology. Sci. Rep. 13, 4430 (2023).
-
Li, W. et al. Optimization and comparison of biodiesel production process by electric heating and microwave-assisted heating transesterification for waste cooking oil via one-way experiments and ANOVA. Front. Environ. Sci. 10, 885453 (2022).
-
Acevedo-Serrano, M., Nogales-Delgado, S. & González González, J. F. Catalytic biolubricant production from Canola oil through double transesterification with methanol and neopentyl glycol. Catalysts 14, 748 (2024).
-
Hamano, F. et al. Quantification of fatty acids in mammalian tissues by gas chromatography–hydrogen flame ionization detection. Bio-protoc 10, e3613 (2020).
-
ASTM International. ASTM D7042: Standard Test Method for Dynamic Viscosity and Density of Liquids by Stabinger Viscometer (and Calculation of Kinematic Viscosity) (ASTM International, 2021).
-
ASTM International. ASTM D4052-22—Standard Test Method for Density, Relative Density, and API Gravity of Liquids by Digital Density Meter (ASTM International, 2022).
-
ASTM International. ASTM D92—Standard Test Method for Flash and Fire Points by Cleveland Open Cup Tester (ASTM International, 2024).
-
International, A. S. T. M. ASTM D5949: Standard Test Method for Pour Point of Petroleum Products (Automatic Pressure Pulsing Method) (ASTM International, 2016).
-
CEN (European Committee for Standardization). EN 14112: Fatty Acid Methyl Esters (FAME)—Determination of Oxidation Stability (accelerated Oxidation test) (CEN, 2020).
-
International, A. S. T. M. ASTM D974-22—Standard Test Method for Acid and Base Number by color-indicator Titration (ASTM International, 2023).
-
ASTM International. ASTM D874-23—Standard Test Method for Sulfated Ash from Lubricating Oils and Additives (ASTM International, 2023).
-
Wijs, J. J. A. The Wijs method as the standard for iodine absorption. Analyst 54, 12–14 (1929).
-
CEN (European Committee for Standardization). EN 116: Diesel and Domestic Heating fuels—Determination of Cold Filter Plugging Point (CEN, 2015).
-
Esmaelzadeh, M., Fallah, N. & Vahabzadeh, F. Kinetic study on the degradation of Acid Red 88 azo dye in a constructed wetland–microbial fuel cell inoculated with Shewanella oneidensis MR-1. Environ. Sci. Pollut. Res. 1–23 (2024). (2024).
-
Arora, N. et al. Co-culturing of oleaginous microalgae and yeast: paradigm shift towards enhanced lipid productivity. Environ. Sci. Pollut Res. 26, 16952–16973 (2019).
-
Eungrasamee, K., Zhu, Z., Liu, X., Jantaro, S. & Lindblad, P. Academic Press,. Lipid metabolism in cyanobacteria: biosynthesis and utilization. In Cyanobacteria 85–116 (2024).
-
Tanaka, M. et al. Quantitative and qualitative analyses of triacylglycerol production in the wild-type Cyanobacterium Synechocystis sp. PCC 6803 and the strain expressing AtfA from Acinetobacter baylyi ADP1. Plant. Cell. Physiol. 61, 1537–1547 (2020).
-
Kahn, A., Oliveira, P., Cuau, M. & Leão, P. N. Incorporation, fate, and turnover of free fatty acids in cyanobacteria. FEMS Microbiol. Rev. 47, fuad015 (2023).
-
Rocha, J. et al. Do galactolipid synthases play a key role in the biogenesis of Chloroplast membranes of higher plants? Front. Plant. Sci. 9, 126 (2018).
-
Mori-Moriyama, N., Yoshitomi, T. & Sato, N. Acyl plastoquinol is a major cyanobacterial substance that co-migrates with triacylglycerol in thin-layer chromatography. Biochem. Biophys. Res. Commun. 641, 18–26 (2023).
-
Redding, K. E. et al. Advances and challenges in photosynthetic hydrogen production. Trends Biotechnol. 40, 1313–1325 (2022).
-
Rosas-Paz, M. et al. Nitrogen limitation-induced adaptive response and lipogenesis in the Antarctic yeast Rhodotorula mucilaginosa M94C9. Front. Microbiol. 15, 1416155 (2024).
-
Amanna, B., Bahri, P. A., Zhang, G. & Moheimani, N. R. Enhancing Arthrospira platensis productivity by optimizing mixing rates in a self-cooling flat plate photobioreactor. Algal Res. 88, 104035 (2025).
-
Murata, N., Takahashi, S., Nishiyama, Y. & Allakhverdiev, S. I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta Bioenerg. 1767, 414–421 (2007).
-
Mulo, P., Sakurai, I. & Aro, E. M. Strategies for PsbA gene expression in cyanobacteria, green algae and higher plants: from transcription to PSII repair. Biochim. Biophys. Acta Bioenerg. 1817, 247–257 (2012).
-
Kouzuma, A., Kato, S. & Watanabe, K. Microbial interspecies interactions: recent findings in syntrophic consortia. Front. Microbiol. 6, 477 (2015).
-
Dias, C., Nobre, B., Santos, J. A., Reis, A. & da Silva, T. L. Lipid and carotenoid production by a Rhodosporidium toruloides and tetradesmus obliquus mixed culture using primary brewery wastewater supplemented with sugarcane molasses and Urea. Appl. Biochem. Biotechnol. 194, 5556–5579 (2022).
-
Ibrahim, S., Chin, G. J. W. L. & Misson, M. Enhanced microbial biomass and lipid production through co-cultivation of yeast Rhodotorula toruloides and microalga chaetoceros muelleri. Malays J. Microbiol. 18 (5), 463–472 (2022).
-
Jiru, T. et al. Optimization of cultivation conditions for biotechnological production of lipid by Rhodotorula Kratochvilovae (syn. Rhodosporidium Kratochvilovae) SY89 for biodiesel Preparation. 3 Biotech. 7, 145 (2017).
-
Hamidi, M., Mohammadi, A., Mashhadi, H. & Mahmoudnia, F. Evaluation of effective environmental parameters on lipid, protein and β-carotene production in spirulina platensis microalga. Results Eng. 18, 101102 (2023).
-
Wang, R. et al. Enhanced microalgal biomass and lipid production via co-culture of scenedesmus obliquus and Candida tropicalis in an autotrophic system. J. Chem. Technol. Biotechnol. 91, 1387–1396 (2015).
-
Gómez-Meriño, F. et al. Olivos-Ramírez, G. Potential use of sludge from El ferrol Bay (Chimbote, Peru) for the production of lipids in the culture of Scenedesmus Acutus (Meyen, 1829). Sci. Rep. 14, 6968 (2024).
-
Chunzhuk, E. A. et al. Direct study of CO₂ capture efficiency during microalgae Arthrospira platensis cultivation at high CO₂ concentrations. Energies 16, 822 (2023).
-
Ronda, S. R., Bokka, C. S., Ketineni, C., Rijal, B. & Allu, P. R. Aeration effect on spirulina platensis growth and γ-linolenic acid production. Braz J. Microbiol. 43, 12–20 (2012).
-
Munch, G., Sestric, R., Sparling, R., Levin, D. B. & Cicek, N. Lipid production in the under-characterized oleaginous yeasts, Rhodosporidium Babjevae and Rhodosporidium diobovatum, from biodiesel-derived waste glycerol. Bioresour Technol. 185, 49–55 (2015).
-
Jan, H. A. et al. Biodiesel production from Sisymbrium Irio as a potential novel biomass waste feedstock using homemade Titania catalyst. Sci. Rep. 13, 11282 (2023).
-
Sathish, T. et al. Environmental remediation at vegetable marketplaces through production of Biowaste catalysts for biofuel generation. Sci. Rep. 13, 5067 (2023).
-
Senila, L., Kovacs, E. & Roman, C. Chemical characterization, lipid profile, and volatile compounds in Chlorella sp. and Spirulina platensis: a promising feedstock for various applications. Molecules 30, 1499 (2025).
-
CEN (European Committee for Standardization). EN 14214: Liquid Petroleum products—Fatty Acid Methyl Esters (FAME) for Use in Diesel Engines and Heating applications—Requirements and Test Methods (CEN, 2019).
-
Shell Global. Shell Gas Compressor Oil S3 PY 220—Product technical data sheet. Royal Dutch Shell plc. (2019). Available at: https://www.shell-livedocs.com/data/published/en-GB/b256f22d-c7e0-4de5-bd26-72499c7fca91.pdf
-
Oliveira, A. A. et al. Design of a biolubricant by the enzymatic esterification of the free fatty acids from castor oil with neopentyl glycol. Process. Biochem. 147, 318–331 (2024).
-
Nor, N. M., Salih, N. & Salimon, J. Optimization and lubrication properties of Malaysian crude palm oil fatty acids-based neopentyl glycol diester green biolubricant. Renew. Energy. 200, 942–956 (2022).
-
Raof, N. A., Yunus, R., Rashid, U., Azis, N. & Yaakub, Z. Palm-based neopentyl glycol diester: a potential green insulating oil. Protein Pept. Lett. 25, 171–179 (2018).
-
Mohammed, M. F. Effect of temperature on lubricating oil and poly(methyl methacrylate) additive. Diyala J. Eng. Sci, 205–220 (2012).
-
Liao, W. et al. Synthesis and properties study of a multifunctional polymethacrylate viscosity index improver. ACS Appl. Eng. Mater. 3, 108–117 (2024).
-
Sulistyo, H., Almeida, M. F. & Dias, J. M. Influence of synthetic antioxidants on the oxidation stability of biodiesel produced from acid Raw Jatropha Curcas oil. Fuel Process. Technol. 132, 133–138 (2015).
-
Birleanu, C. et al. Effect of TiO₂ nanoparticles on the tribological properties of lubricating oil: an experimental investigation. Sci. Rep. 12, 5201 (2022).
-
Rahmani, A., Razavi, H. K. & Dehghani-Soufi, M. Green tribology assessment: A comprehensive review of biolubricants and nano enhancers. Energy Convers. Manag. X 24, 100794 (2024).
Acknowledgements
The authors would like to express their sincere gratitude to Mr. Mahdi Esmaelzadeh for his valuable assistance in performing part of the laboratory analyses.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Fazelian, H., Moraveji, M.K., Mozaffarian, M. et al. Production of biodiesel and biolubricant via co-cultivation of Arthrospira platensis and Rhodosporidium babjevae using pulp-paper mill sludge hydrolysate. Sci Rep 15, 44937 (2025). https://doi.org/10.1038/s41598-025-28597-z
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-28597-z