
Introduction
Aquaculture plays a vital role in sustainable food production, providing valuable products for global human consumption1. The increasing effects of global warming on water temperatures, marine biodiversity, and aquatic flora underscore the critical need to enhance fish farming practices2. Among farmed species, Litopenaeus vannamei, commonly known as the Pacific white shrimp, stands as the most important cultured species within the Penaeidae family. Its high commercial value, exceptional breeding ability3and adaptability to a wide range of salinities, coupled with the availability of specific pathogen-free (SPF) broodstock4, make it the dominant species in global shrimp aquaculture. Consequently, shrimp farming has emerged as a primary food production method in coastal regions, driving economic growth5and fostering small-scale entrepreneurial activity6.
The sustainability of L. vannamei farming is significantly threatened by various factors, dominant them the outbreaks of bacterial diseases. Intensification, suboptimal water quality, and undernutrition across different production stages exacerbate this issue, leading to global industry impacts7,8.
Vibrio parahaemolyticus, a Gram-negative, halophilic bacterium prevalent in marine, estuarine, and coastal environments, is a primary concern, causing substantial mortalities and significant economic losses worldwide9,10. Intensive aquaculture’s reliance on antibiotics for bacterial infection control presents a significant challenge. While crucial for disease management, widespread antibiotic use promotes drug-resistant bacteria, compromising future treatment efficacy. Moreover, most of administered antibiotics (70–80%) contaminate the environment, adversely affecting ecosystems and aquatic life11,12. Consequently, there is a growing imperative for sustainable, environmentally friendly bioactive materials that can enhance immunity and productivity for disease control, thereby overcoming the limitations of antibiotics13.
Nature provides a range of disease-mitigation strategies for aquaculture, such as microalgae, herbal extracts and oils, probiotics, and fungi. However, honeybee products have recently garnered significant attention as a promising alternative. Building on the suggestion that bee-derived compounds possess the potential to treat bacterial diseases in fish14 researchers have begun to investigate the utility of royal jelly, propolis, and pollen in aquaculture settings15. Bee venom (BV), or apitoxin, is a complex biotoxin produced by honeybees (family Apidae) in a specialized gland connected to their sting16. While primarily a defense mechanism against predators17, BV exhibits a wide array of pharmacological actions, including anti-inflammatory, antimutagenic, antibacterial, radioprotective, anti-nociceptive, immunostimulatory, hepatoprotective, and anti-cancer effects18,19,20. Notably, BV has demonstrated antimicrobial activity against several fish pathogens21,22, and its dietary supplementation significantly enhanced growth performance in Thinlip mullet, Chelon ramada23. Beyond individual benefits, BV also contributes to social antisepsis and collective immunity within the bee colony.
Chitosan, a naturally occurring compound, is widely recognized as a promising, environment friendly feed additive, finding applications across aquaculture, agriculture, medicine, food, and the chemical industry24. Its utility extends to drug delivery systems in various fields25. In aquaculture, chitosan supplementation in fish diets has significantly enhanced growth performance, immune function, and antioxidant status26. However, its nano-conjugation with BV remains a relatively unexplored area. The use of polymers like chitosan as drug carriers offers a novel approach in aquaculture, improving the stability, efficacy, and water solubility of natural compounds27.
While BV has applications in complementary medicine for treating inflammation-related diseases in humans28. and its antibacterial effects against bacteria from diverse sources have been studied29,30, its use in shrimp aquaculture, whether in natural or nanoparticle form, remains unexplored. To our knowledge, this study is the first to evaluate the dietary incorporation of bee venom as a nano-nutrient in shrimp feed, investigating its potential as both a nutritional and therapeutic supplement. Consequently, this study aimed to comprehensively evaluate the impact of dietary bee venom-loaded chitosan nanoparticles (BV-CSNP) as a novel growth promoter on the health of Pacific white shrimp (Litopenaeus vannamei). The investigation encompassed an assessment of growth parameters, whole-body composition, non-specific immune indicators, the transcriptional expression of antioxidants and immune-related genes, organ histology, and disease resistance against a V. parahaemolyticus challenge.
Materials and methods
Sources of Bee-Venom and chitosan
Fresh exuviae shells from Hermetia illucens (AMININS Co., Egypt) were washed with distilled water, then dried in an oven at 70 °C for 36 h. After drying, the shells were crushed using a blender and stored at room temperature for future use. Chitosan production commenced with preconditioning the shells to remove loosely bound proteins. One hundred grams of shells were immersed in a 0.05 M acetic acid solution (Sigma-Aldrich Co., USA) for 24 h, followed by washing with distilled water (D.W) and drying in an oven at 70 °C for 4 h. The next stage, deproteination, involved dissolving proteins and sugars. The preconditioned powder was soaked in a 12.5 M NaOH solution (1 part shell to 15 parts solution by w/v) for 2 h under constant magnetic stirring, then cooled at room temperature for 30 min. Next, demineralization entailed filtering the shells, washing the residues with D.W until neutral pH, and re-drying them. Subsequently, the resulting powder was treated with 1 L of 1% HCl for 24 h. After filtration, the residue was washed with D.W until neutral pH and dried in an oven at 70 °C for 4 h. The final step, deacetylation, partially removed acetyl groups from the chitin structure. This process involved adding a 50% NaOH solution (1 part shell to 10 parts solution w/v) and stirring magnetically at 100 °C for 6 h. The sample was then cooled to room temperature, filtered, washed with D.W until neutral pH, and dried in an oven at 70 °C. The resulting material was chitosan31,32.
Synthesis of Bee-Venom loaded chitosan nanoparticles (BV-CS NPs)
Chitosan nanoparticles (CSNPs) were fabricated using the ionotropic gelation method33. First, 0.3 g of chitosan was dissolved in 100 mL of 1% acetic acid (in D.W) and magnetically stirred at room temperature until a clear solution was obtained. The chitosan solution’s pH was then adjusted to 5 with 1 M NaOH. Concurrently, a sodium tripolyphosphate (TPP) solution (Sigma-Aldrich Co., USA) was prepared by dissolving 0.1 g of TPP in 33.3 mL of D.W. This TPP solution was added dropwise to the chitosan solution under magnetic stirring at 1000 rpm at room temperature, maintaining a 3:1 (v/v) chitosan: TPP ratio. Sonication was performed for 20 min. The resulting chitosan particle suspension was cooled, centrifuged at 12,000 rpm for 60 min, lyophilized, and stored at 4 °C. For bee venom-loaded nanoparticles, 0.1 g of honeybee venom (purchased from the Department of Honeybee Research, Institute of Plant Protection, Giza, Egypt) was added to the TPP solution before its addition to the chitosan solution. Finally, the mixture was centrifuged at 11,000 rpm and 4 °C for 60 min, lyophilized, and stored at 4 °C. All chemicals and solvents used were of analytical grade and high purity.
Characterization of BV-CSNPs
The morphology of CSNPs and BV-CSNP (combination) was identified. For TEM analysis, a drop of each sample was placed on a carbon-coated copper grid and allowed to dry at room temperature. Electron micrographs were acquired using a JEOL JEM-1010 transmission electron microscope operating at 80 kV at the Regional Center for Mycology and Biotechnology (RCMB), Al-Azhar University34.
Shrimp and rearing conditions
After two weeks of acclimation, 240 healthy L. vannamei with an average initial weight of 6.00 ± 0.02 g were randomly selected and uniformly distributed into four triplicate groups in 1m3 hapas (20 shrimp/hapa). This took place at a private earthen pond shrimp farm in Damietta, Egypt. Throughout the experimental period, the shrimp rearing conditions maintained the following water parameters: dissolved oxygen 6.60 ± 0.03 mg/L, pH 8.16 ± 0.01, salinity 37.50 ± 0.04 g/L, ammonia nitrogen content 1.22 ± 0.01 mg/L, toxic ammonia 0.13 ± 0.00, and temperature 27.13 ± 0.07 °C. Shrimp were fed daily at 5% of their body weight, with 15% of the rearing water changed daily.
Diet Preparation and feeding protocol
-
The nutritional ingredients and chemical composition of these experimental diets are detailed in Table 1. We prepared experimental diets by thoroughly mixing a commercial shrimp diet (Skretting Company, Egypt®) with various supplements. Chitosan nanoparticles (CS NPs) were incorporated at a concentration of 1 g per kg of diet, based on established protocols26. Bee venom loaded chitosan nanoparticles (BV-CSNPs) was then added to the commercial diet at three different concentrations: 0.1, 0.2, and 0.3 mg per kg of diet. A control group received only the basal commercial diet without any supplementation. Shrimp were fed these diets three times daily, at 8:00 AM, 12:00 PM, and 4:00 PM, for a period of 63 days. The daily feeding rate was maintained at 5% of their total body weight.
Growth performance
For assessing growth attributes, shrimp (20 shrimp) in each group were weighted before and after 63 days after the feeding. The feed conversion ratio (FCR), final weight (FW), specific growth rate (SGR), and weight gain (WG) were determined using the following formulas.
-
WG (%) = (Final weight (g)-Initial weight (g))/(Initial weight (g)) ×100.
-
SGR= (ln FW – ln IW)/(t)×100.
Where: ln is the natural logarithmic of FBW and IBW; t = time in days.
-
FCR= (Feed intake (g))/(weight gain (g)).
Shrimp body composition analysis
Shrimp body samples (n = 3) were evaluated for their proximate components by a conventional method35. The Kjeldahl method, which traps NH3 with boric acid, was used to calculate crude protein (N × 6.25). Dry matter was assessed by drying samples in an oven at 105 °C. The ash concentration was established by burning 2 g of samples in a muffle furnace at 550 °C for 4 h. The Soxtect System HT extraction unit extracted the total lipids with ether. Additionally, gross energy was established through a Schimadzu CA-4P bomb calorimeter36.
Hemolymph sampling and non-specific immunological parameters analysis
Hemolymph samples (250 µl) were picked up from five randomly selected shrimp per treatment from the ventral-sinus cavity using a 25-gauge needle and a 1 ml syringe that was inserted with a pre-cooled (4 °C) anticoagulant37.
Total hemocyte count
Fifty µl of the anticoagulant-hemolymph mixture was diluted in 150 µl of formaldehyde (4%). Then 20 µl of the diluted mixture was examined on a Neubauer hemocytometer to measure the total hemocyte count, THC. Hemocytes were counted using a light microscope at 100× magnification and calculated as; THC (cells/mL) = count×106 × dilution factor.
Phenoloxidase activity
Phenoloxidase (PO) activity was determined spectrophotometrically by monitoring the production of dopachrome from L-dihydroxyphenylalanine (L-DOPA)37,38. The shrimp’s PO activity was assessed by measuring the absorbance of dopachrome at 490 nm, which was quantified by observing the increase in optical density per minute per milliliter of protein.
The lysozyme activity
Lysozyme activity (U ml−1) was evaluated in shrimp hemolymph using the ELISA micro-well technique with a lysozyme ELISA kit (CAT. No. SL0050FI, Sunlong Biotech Co., China). Following the manufacturer’s instructions, the reaction was carried out at room temperature, and the absorbance at 450 nm was recorded using a microplate ELISA reader at 30 s and 4 min. One unit of lysozyme activity was defined as the amount of sample needed to cause a decrease in absorbance by 0.01 per minute39.
The phagocytotic activity
The phagocytotic assay followed the procedures of a previous study40 with some modifications. Shrimp hemocytes were cleansed with a saline solution (NaCl 28.4 g, MgCl2-6H2O 1.0 g, MgSO4-7H2O 2.0 g, CaCl2-2H2O 2.25 g, KCl 0.7 g, glucose 1.0 g, and HEPES 2.38 g/l) and the viable cell count was adjusted to 1 × 106 cells/ml. The cell suspension (200 µl) was applied to a cover slip. The cell suspension was withdrawn after 20 min and thoroughly washed with shrimp saline 3 times. The Candida albicans preparation (2 mL) was added and incubated for 2 h. Afterwards, the cell suspension was rinsed 5 times with shrimp saline to attain a concentration of 5 × 108 cells/ml, then fixed with 100% methanol. The cover slip was stained with Giemsa stain, mounted and examined under the microscope. Two hundred hemocytes were counted for each sample. The phagocytic activity and index were calculated using the following equation.
-
-Phagocytic rate (PR) (%) = (Number of phagocytic hemocytes)/(Total number of hemocytes)×100,
-
-Phagocytic index (PI)= (The Number of phagocytic C. albicans cells by hemocytes)/(Total number of phagocytes hemocytes) ×100.
Bactericidal activity: Bactericidal activity was determined as described by Adams41. Hemolymph samples were serially diluted in a sterile buffer saline (pH 7.5) at 1:2, 1:4, 1:8, 1:16, and 1:32. V. parahaemolyticus suspensions were appropriately diluted with their respective saline diluents before 100 µl of each bacterial suspension was incubated with 100 µl of hemolymph, which was incubated at room temperature for 3 h. Fifty microliters of each hemolymph-bacterial mixture were then inoculated onto TSA plates and incubated at 28 °C for 24 h, alongside a positive control plate. Colony-forming units (CFU) were counted using the plate counting method, and the results were expressed as the percentage inhibition (PI) of V. parahaemolyticus was calculated as follows:
Inhibition % = [100- (mean CFU in sample/mean CFU positive control)] × 100.
Antioxidant activity assessment
Superoxide dismutase (SOD) activity was measured using a commercial kit (Biodiagnostic Co., Egypt; SD 2521) according to the manufacturer’s instructions. Absorbance was measured calorimetrically at 560 nm42, and SOD activity was expressed as units per milliliter (U/mL). Catalase (CAT) activity was measured at 550 nm using a microplate reader, following the method described by Johansson and Håkan Borg43, and expressed as units per milligram of protein (U/mg protein). Malondialdehyde (MDA) levels were measured according to a previously described method44, and the lipid peroxide level was expressed in µmol l −1.
Digestive enzyme activity
Lipase and amylase activities were measured colorimetrically for each treatment. Intestinal samples (n = 3) were homogenized in sterile, ice-cold saline solution and centrifuged at 3500 × g for 10 min at 4 °C. Supernatants were collected for analysis. Lipase activity was determined using a commercial kit (Spectrum, Egypt; REF: 281 001) and expressed as units per milligram of protein (U/mg protein)45. Amylase activity was measured using a commercial kit (Biodiagnostic Co., Egypt; CAT. NO. AY 10 50) according to the manufacturer’s instructions and expressed as U/mg protein. Protease activity was measured using a fluorometric protease activity assay kit (ab112152).
Transcriptional expression of antioxidant and immune-related genes
Total RNA was isolated from the hepatopancreas tissues of each experimental group using the RNA purification kit (Thermo Fisher Scientific, USA) according to the manufacturer’s instructions. A nanodrop lite spectrophotometer (Thermo Scientific, US) was used to measure the O.D. 260 nm/O.D. 280 nm ratio to assess the quality of the extracted RNA. Complementary DNAs (cDNAs) were synthesized from 1 µg of RNA using the SuperScript TM III First-Strand Synthesis System (Invitrogen, USA) and Oligo-dT primers following the manufacturer’s instructions. After that, the cDNA samples were stored at −20 °C until they were needed again. Consequently, the mRNA expressions of immune (Lipopolysaccharide and β−1,3-glucan-binding protein (LGBP), Peroxinectin (PX), Prophenoloxidase activating (ppA) enzyme and antioxidant (cytosolic manganese superoxide dismutase (cytMnSOD), mitochondrial manganese superoxide dismutase (mtMnSOD) related genes were valued using qPCR (SensiFast SYBR Lo-Rox kit, Bioline, London, UK) to quantify the gene transcription folds (Table 2). The thermal cycle conditions for the reaction were adjusted as follows: 10 min at 95 °C, 40 cycles at 95 °C for 15 s, 30 min at 60 °C, and finally 5 min at 85 °C. The transcription levels were standardized to the β-actin gene as an internal control gene according to the 2−ΔΔCT method46.
Histopathological analysis
The hepatopancreas, intestine, and muscle tissue from shrimp of each treatment group (three shrimp/hapa) were collected after 63 days and fixed in Davidson solution for 24 h. The tissues were then dehydrated in a series of ethanol concentrations (70%, 80%, 90%, and 100%), cleared in xylene, embedded in paraffin, and sectioned at a thickness of 4 mm. The tissue slides were stained with H&E dyes and examined under a light microscope (Leica, UK) for histological evaluation, with images taken for documentation47. The villus height (VH), villus width (VW), and intestinal muscle thickness (MT) were measured using the software ImageJ imaging software (1.54d, NIH, USA) and evaluated.
Challenge assay
Following the feeding trial, a challenge test was conducted to evaluate shrimp resistance to a pathogenic V. parahaemolyticus strain48. The bacterial strain was cultured in Brain Heart Infusion broth supplemented with 2% NaCl at 28 °C for 24 h. After incubation, the bacterial suspension was centrifuged, and the resulting pellet was washed several times with sterile saline solution. The pellet was then resuspended to a final concentration of 10⁷ CFU/mL. Thirty shrimp from each dietary treatment (Control, T1, T2, and T3) were randomly distributed into twelve tanks (60 × 35 × 30 cm³) with three replicates per treatment (10 shrimp/tank). All groups were challenged by immersion in water containing the V. parahaemolyticus suspension at a concentration of 10⁷ CFU/mL for 24 h at 28 °C49. Shrimp were monitored for six days post-challenge, or until mortalities ceased. Dead shrimp were removed and recorded daily. V. parahaemolyticus was re-isolated from the hepatopancreas of freshly dead shrimp using Thiosulfate-Citrate-Bile Salts-Sucrose Agar (TCBS). Mortality rates were calculated as: Mortality (%) = (Number of dead shrimp/Total number of shrimp) × 100.
Statistical analysis
To analyze the data and identify significant differences among treatments, we employed one-way ANOVA followed by Duncan’s multiple range test. Statistical analyses were conducted using SPSS version 26.0 (IBM Corp., Armonk, NY, USA). Results are presented as means ± standard error of the mean (SEM), with statistical significance set at P < 0.05.
Results
Characterization of CSNPs and BV-CSNPs
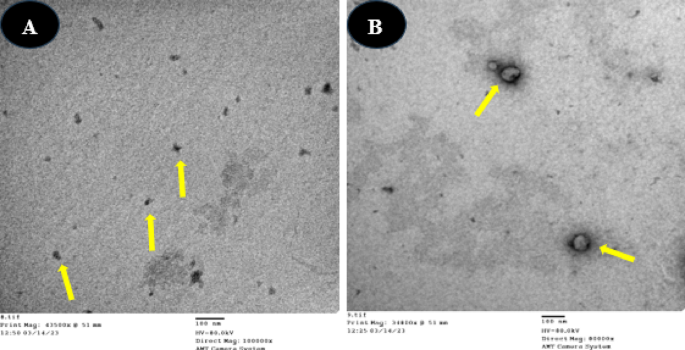
TEM screening was employed to examine the particle size distribution and morphology of both chitosan nanoparticles (CSNPs) and bee venom-loaded chitosan nanoparticles (BVCSNPs). TEM images were captured at a magnification of 48.000×. The analysis showed that CS NPs were spherical with smooth surfaces and had an average size of 39.5 ± 8.21 nm. In contrast, BV CS NPs exhibited an average size of 98.25 ± 7.22 nm (Fig. 1).

TEM images of the biosynthesized nanoparticles: (A) CSNPs, and (B) BV-CSNPs. The nanoparticles are clearly visible as distinct, well-dispersed spherical structures. TEM images were captured at a magnification of 48.000×.
Growth and survival
After a 63-day feeding trial, shrimp growth performance was evaluated, and the results are summarized in Table 3. Shrimp fed diets containing varying concentrations of BV-CSNPs exhibited significantly enhanced FW, WG, SGR, and biomass compared to the free diet of BV-CSNPs group (P < 0.05). Furthermore, the FCR was notably lower in all BV-CSNP-supplemented groups compared to the control (P < 0.05). Interestingly, there were no significant differences in FCR observed among the various BV-CSNP concentrations. It’s also worth noting that survival rates, and FI did not differ substantially among any of the treatment groups (P < 0.05). The most favorable growth, as indicated by the highest FW, WG, and average daily gain (ADG), was observed in shrimp fed the diet containing 0.2 mg/kg BV-CSNPs. Shrimp fed the diet containing 0.2 mg/kg of BV-CSNPs showed significantly greater biomass compared to the control group (P < 0.05). However, no significant differences in biomass were observed among the various BV-CS NP-treated groups (P > 0.05).
Whole body composition
Table 4 presents the whole-body composition of shrimp after a 63-day feeding assessment with diets containing BV-CSNPs and a control diet. Overall, no significant alterations (P > 0.05) were observed in moisture, dry matter, protein, or lipid content between the BV-CSNPs-fed groups and the control. The shrimp fed 0.1 mg BV-CSNPs exhibited meaningfully higher moisture content and lower dry matter and protein content. In contrast, ash content was drastically greater (P < 0.05) in all BV-CSNP-fed groups when compared to the basal diet. Interestingly, gross energy tended to decrease (P > 0.05) with raising amounts of BV-CSNPs in the shrimp diets.
Non-Specific immunological parameters
Table 5 exhibits the immunological parameters of shrimp after 63 days of feeding with experimental and control diets. A significant increase (P < 0.05) in total hemocyte count was noted in the hemolymph of shrimp fed diets supplemented with BV-CSNPs, with higher dietary inclusion levels leading to greater counts. Furthermore, phenoloxidase activity was significantly elevated (P < 0.05) in shrimp fed 0.3 mg of BV-CSNPs compared to all other groups. However, there were no significant differences were observed among the other BV-CSNPs groups. Lysozyme activity was significantly enhanced (P < 0.05) in the group fed the highest level of BV-CSNPs (0.3 mg/kg) compared to the control and other BV-CSNPs groups. Shrimp fed the diet containing 0.3 mg/kg of BV-CS NPs exhibited significantly higher phagocytic activity compared to both the control group and all other treatment groups. The phagocytic index was significantly higher (P < 0.05) in shrimp fed 0.2 mg/BV-CSNPs linked to the control and other BV-CSNPs groups. Furthermore, hemolymph from all shrimp fed BV-CSNP diets exhibited significantly greater bactericidal activity (P < 0.05) compared to the control group. This indicates an improved capacity to combat bacterial infections. Within the BV-CSNP groups, the shrimp in 0.3 group demonstrated the highest bactericidal activity, suggesting a dose-dependent effect on this immune response.
Antioxidants status and lipid peroxidation
As shown in Fig. 2, supplementation with 0.2 mg/kg (T2) and 0.3 mg/kg BV-CSNPs (T3) led to a significant increase (P < 0.05) in the activities of both superoxide dismutase (SOD) (Fig. 2A) and catalase (CAT) (Fig. 2B) compared to the control and T1 groups. The CAT level was highest in shrimp fed diets containing 0.3 mg/kg of bee BV-CSNPs, significantly surpassing (P < 0.05) all other experimental groups. Furthermore, both the 0.2 mg/kg (T2) and 0.3 mg/kg BV-CSNPs groups exhibited a significant reduction (P < 0.05) in malondialdehyde (MDA) levels (Fig. 2C) compared to the control.

Antioxidant parameters of Pacific white shrimp, L. vannamei fed the control and the various levels of BV-CS NPs supplemented diets after 63-day feeding trial (means ± SE). A: SOD activity, B: CAT activity, and C: MDA activity. Superscripts represent significant (P < 0.05) differences among treatments.
Digestive enzymes
Lipase activity (Fig. 4 A) was significantly higher in shrimp fed diets containing 0.2 mg/kg (T2) and 0.3 mg/kg (T3) of BV-CSNPs compared to other groups (P < 0.05). The highest levels of lipase activity were observed in the 0.3 mg/kg BV-CSNP (T3) group. The addition of gradual levels of BV-CS NPs to the diet led to a dose-dependent improvement in protease activity (P < 0.05), with the highest levels of protease activity were observed in the 0.3 mg/kg BV-CSNP (T3) group (Fig. 4 C). Both the 0.2 mg/kg (T2) and 0.3 mg/kg (T3) concentrations of BV-CSNPs resulted in significantly higher amylase levels (P < 0.05) compared to the other experimental groups (Fig. 4B).

Digestive enzyme activity of Pacific white shrimp, L. vannamei fed the control and the various levels of BV-CS NPs supplemented diets after 63-day feeding trial (means ± SE).A: lipase activity, B: amylase activity, and C: Protease activity. Superscripts represent significant (P < 0.05) differences among treatments.
Gene expression
Gene expression analysis (Fig. 4) revealed a significant up-regulation (P < 0.05) of LGBP (Fig. 4A), PX (Fig. 4B), and PPT (Fig. 4C) genes in all groups of shrimp fed diets containing BV-CS NPs compared to the control group, showing a dose-dependent response. Similarly, cytMnSOD (Fig. 4D) and mtMnSOD (Fig. 4E) genes were significantly up-regulated (P < 0.05) in all BV-CS NPs-supplemented groups compared to the control group.

The relative mRNA expressions of LGBP, PX, PPT, mtMnSOD, and cytMnSOD genes among Pacific white shrimp, L. vannamei fed the control and the various levels of BV-CS NPs supplemented diets after 63-day feeding trial (means ± SE). Superscripts represent significant (P < 0.05) differences among treatments.
Histopathological analysis
Histological examination of shrimp hepatopancreas tissue (Fig. 5) provided insights into the cellular structure and overall health of this vital organ across the different dietary groups. In both the control (Fig. 5A) and T1 (Fig. 5B) groups, the hepatopancreas exhibited a normal cellular structure, with clear identification of R-cells (resorptive cells), B-cells (blister-like cells), and F-cells (fibrillar cells). However, these groups also showed mild edema between the tubules and some hemocyte infiltration, suggesting minor stress or inflammatory responses. In contrast, the T2 (Fig. 5C) and T3 (Fig. 5D) groups displayed a normal hepatopancreatic architecture, characterized by well-defined tubules connected by basophilic connective tissue strands. Notably, these groups, especially the T3 group, showed a noticeable increase in the number of R-cells and B-cells.
As shown in Fig. 6 (A-D), shrimp fed BV-CSNP (0.2 or 0.3 mg/kg) diets exhibited an improved intestinal structure compared to the control group (Fig. 6A) and T1 (Fig. 6B). Specifically, the T2 (Fig. 6C) and T3 (Fig. 6D) groups displayed normal epithelium and microvilli morphology. Furthermore, these groups showed significantly greater (P < 0.05) enterocyte height (EH) and intestinal wall muscle thickness (WT) when compared to the control group (Fig. 6E & F). These findings suggest enhanced nutrient absorption and overall intestinal health with BV-CS NP supplementation. Regarding muscle tissue, mild intramuscular edema was observed in the control and lower BV-CS NP groups (Fig. 7A-C). In contrast, the T3 group presented normal muscle tissue structure (Fig. 7D), indicating that higher concentrations of BV-CS NPs may help mitigate muscle stress or inflammation.

Photomicrograph of hepatopancreas tissues of L. vannamei fed with the control and the three BV-CS NPs diets at 63 days. (A&B) Control & T1 groups showing normal histological structure with hemocytes infiltration (Short arrow); besides mild edema between hepatopancreatic tubules; (C) T2 group showing normal histological structure with hemocytes infiltration (short arrow); (D)T3 group showing obvious increase in B-cells (thin arrow), and R-cells (arrowhead). F-cells (double arrows), L: lumen. Stain H&E. Scale Bars = 50 μm.

Photomicrograph of intestinal tissues of L. vannamei fed the control and the three BV-CS NPs diets at 63 days. (A) Control group, T0, showing short epithelium with a thin circular muscle layer besides sloughing of some enterocytes; (B) T1 group showing intact epithelium with thin circular muscle layer; (C& D) T2 & T3 groups showing increased microvilli folding with improved enterocyte height with thick intestinal wall. (E) Enterocytes height, (F) Intestinal wall thickness. 1: Mucosa brush border, 2: Intestine epithelium, 3: Epithelium height, 4: Wall thickness, and 5: Intestine lumen. Stain H&E. Scale Bars = 50 μm.

Photomicrograph of muscular tissues of L. vannamei fed the control and the three BV-CS NPs diets at 63 days. (A) The control and the T1 groups showing intramuscular oedema (black arrow), (C) The T2 group showing mild oedema between muscle bundles, and (D) The T3 group exhibits normal tissue appearance. mf: muscle fibres, n: muscle nucleus. Stain H&E. Scale Bars = 50 μm.
Challenge against V. parahaemolyticus
Following a challenge with virulent Vibrio parahaemolyticus, cumulative mortality rates were significantly lower in shrimp fed diets containing BV-CS NPs compared to the control group (Fig. 8). Mortality decreased in a dose-dependent manner, with the lowest mortality rate observed in the T3 group (20%), followed by the T2 group (25%), and then the T1 group (30%). All BV-CS NPs-supplemented groups exhibited substantially lower mortality than the control group (65%).

The mortality rate in Pacific white shrimp, L. vannamei fed the control and the three BV-CSNPs diets over 63 days and at 5 days post-challenge with V. parahaemolyticus.
Discussion
The pervasive use of antibiotics in aquaculture has led to significant environmental concerns and poses potential risks to both human and aquatic animal health. This pressing issue underscores the urgent need for natural, sustainable alternatives to antibiotics to safeguard the environment while simultaneously boosting productivity and health in farmed fish. Over the past two decades, the exploration of natural molecules has emerged as a promising strategy in aquaculture. However, their efficacy has been dramatically amplified by the integration with cutting-edge advancements in nanotechnology. This synergy allows for enhanced delivery, bioavailability, and targeted action of these natural compounds, paving the way for more environmentally responsible and effective aquaculture practices.
Recent research highlights the significant benefits of various nano-products in aquaculture, offering promising solutions for improving aquatic animal health and productivity. For example, Eissa, et al.26 investigated the use of pumpkin seed oil-loaded chitosan nanoparticles in whiteleg shrimp (Litopenaeus vannamei). Their findings showed that this nano-product significantly enhanced growth, boosted digestive enzyme activity, improved immune response, and increased survival rates against V. parahaemolyticus. Another recent study investigated the effect of BV-CSNPs as a potential therapeutic strategy for enhancing tilapia health and resistance to Aeromonas hydrophila50. Recent studies have shown that using chitosan-loaded particles in aquaculture can have numerous positive impacts. Likewise, Mabrouk, et al.51 found that supplementing the diet of Nile tilapia broodstock with Arthrospira platensis nanoparticles led to improvements in growth performance, a more balanced steroid hormone profile, and enhanced reproductive productivity. Similarly, Said, et al.52 concluded that dietary selenium nanoparticles derived from Spirulina platensis positively impacted growth performance and physio-biochemical components in Pacific white shrimp (L. vannamei), while also offering an alleviating effect against cadmium toxicity. Furthermore Eissa, et al.53 demonstrated that the dietary administration of nano curcumin to red tilapia (Oreochromis sp.) resulted in increased growth performance, improved body composition, healthier blood parameters, beneficial histopathological changes, and enhanced resistance against Aspergillus flavus. This experiment is the first to investigate the favorable consequences BV-CSNPs on growth performance, immune response, hemolymph health, organ histology, and survival following challenge with V. parahaemolyticus. Dietary supplementation with BV-CSNPs at levels of 0.2 or 0.3 mg/kg improved growth variables and overall health. This enhancement was demonstrated by enhanced antioxidant status (increased SOD and CAT activity), reduced MDA levels, and up-regulated expression of LGBP, PX, and PPT genes.
These improvements can be attributed to the unique characteristics of bee venom, which exhibits promising biological activities. Numerous studies have demonstrated that BV has a unique composition containing a variety of bioactive compounds54,55,56. These compounds contribute to BV’s diverse therapeutic properties, including anti-inflammatory57immunostimulatory activity58anticancer properties59and antimicrobial effects60as well as the potential to enhance reproductive61 and growth performances62.
In the present study, dietary supplementation with BV-CSNPs for 63 days enhanced growth performance in L. vannamei, as evidenced by increased final weight and a reduced FCR. Similar improvements in weight gain and survivability have been reported in pigs and broiler chicks supplemented with BV63,
This growth enhancement may be attributed to the bioactive compounds in BV, which could improve nutrient absorption and utilization by promoting healthy digestive tract function. This, in turn, would lead to a lower FCR and more efficient growth64,65. The observed normal morphology of the intestinal epithelium and microvilli in shrimp fed diets containing higher levels of BV-CSNPs supports this hypothesis. Furthermore, the significant increase in epithelial height and intestinal wall muscle thickness indicates improved intestinal health, which is crucial for nutrient absorption in shrimp66. Therefore, dietary BV-CSNPs likely enhance digestive capacity and promote nutrient absorption. These results align with previous research67 highlighting the importance of the intestinal mucosa as a primary barrier against pathogens and suggesting that intestinal health significantly influences overall health and immunity in shrimp.
Dietary BV-CSNPs supplementation significantly increased moisture and ash content while decreasing whole-body protein, lipid, and gross energy levels. In a recent study we conducted using red tilapia, we observed similar trends: moisture, protein, and ash content increased, while lipid content decreased. These variations may be attributed to species-specific differences and environmental conditions68. Further supporting the positive impact of bee venom, a study in broiler chickens demonstrated that bee venom in drinking water led to increased carcass weight and reduced abdominal fat69. Similarly, Kim, et al.70 reported that BV supplementation enhanced relative breast meat yield in broiler chickens, although it did not significantly affect abdominal fat weight. These variations could be attributed to the source of bee venom (BV), the study duration, animal type and breed, and other experimental conditions.
This study highlights the immunostimulatory activity of BV in shrimp, demonstrating its positive impact on both cellular and humoral components of the innate immune system. Shrimp fed diets containing BV-CS NPs (0.2 or 0.3 mg/kg diet) showed a significant increase in hemocyte numbers, a key indicator of cellular immune response. This increase was directly correlated with the inclusion level of BV-CSNPs in their diet.
Phagocytic activity of hemocytes was also enhanced in a dose-dependent manner. This rise in phagocytic cells, which are crucial for engulfing and destroying pathogens, aligns with previous research on BV’s effects, such as those observed in male rabbits61. The improved immune response likely contributes to greater disease resistance in shrimp55. The anti-inflammatory activities of BV are attributed to its bioactive peptides, such as melittin and MCD-peptide, as well as key enzymes, particularly phospholipase A2 (PLA2)71. Notably, PLA2 may also play a role in the observed increase in immune cell populations, further bolstering the shrimp’s defense mechanisms72. The secondary arm of the shrimp’s innate defense is the humoral response, which involves the release of various immune molecules. This includes the prophenoloxidase (Pro-PO) system73and antimicrobial peptides such as penaeidins, and lysozymes, which serve as a first line of defense against invaders, in addition to antioxidant defense enzymes like SOD and CAT play a crucial role in this response74.
This study reveals that adding BV to shrimp diets, specifically in the form of BV-CSNPs, can significantly improve their immune system. A key finding is the potential enhancement of phenoloxidase (PO) levels in all shrimp fed BV-CS NPs. PO is a critical enzyme in the proPO-activating system, essential for converting inactive proPO into active PO75. This process is vital for melanin biosynthesis, a crucial part of the innate immune response that helps shrimp recognize foreign invaders and activate cellular defenses like hemocyte recruitment, phagocytosis, and melanization. Therefore, higher PO levels directly indicate a stronger immune response. It’s worth noting that a reduction in the proPO system’s activity can impair phagocytosis, underscoring PO’s importance73. In addition to PO, the study observed a significant increase in lysozyme activity in shrimp fed the highest concentration of BV-CSNPs. Lysozyme is known to modulate both cellular and humoral defense mechanisms, protecting against diseases. A study by Mai and Wang76 confirmed that lysozyme modulated the cellular and humoral defense mechanisms to protect shrimp against diseases. The enhanced immune response in shrimp is likely due to BV’s potent anti-inflammatory and mast cell-degranulating peptides. These peptides stimulate the secretory mechanisms of inflammatory cells, thereby strengthening the overall immune defense77.
In this context, BV proves to be a powerful ally in enhancing shrimp health. Its bioactive components, like melittin and adolapin, do more than just scavenge free radicals and reduce oxidative stress. This study shows that in shrimp given high levels of BV-CSNPs, these components significantly ramp up the activity of key antioxidant enzymes such as SOD and CAT. At the same time, they bring down MDA levels, which are indicators of oxidative damage. Essentially, these components work together to neutralize harmful free radicals, lessen oxidative stress, and bolster immune function56,58. This protects shrimp cells from damage and keeps them healthy, aligning with findings from various animal studies61,62.
This study emphasizes the antioxidant efficacy of BV, evidenced by significantly reduced levels of MDA in shrimp78,79. MDA is a widely recognized biomarker for oxidative stress in biological systems80. The decrease in MDA indicates that BV effectively protects polyunsaturated fatty acids in cell membranes from lipid peroxidation and subsequent oxidative damage. Therefore, monitoring MDA levels proved to be a crucial indicator for evaluating the effectiveness of dietary antioxidants in mitigating oxidative stress and improving shrimp health.
These findings are further supported by the preservation of normal histological architecture observed in the muscle tissue of shrimp fed high levels of BV-CSNPs. In stark contrast, the control group and those receiving lower doses of BV-CSNPs exhibited clear signs of cellular and tissue degeneration. This provides strong visual evidence of BV’s protective effects against oxidative damage. These findings are corroborated by the preservation of normal histological architecture in the muscle tissue of shrimp fed high levels of BV-CS NPs. In contrast, the control group and those receiving lower doses displayed cellular and tissue degeneration.
This study revealed that shrimp fed diets supplemented with 0.2 and 0.3 mg/kg of BV-CSNPs exhibited significantly higher activity of digestive enzymes, including lipase, amylase, and protease, compared to other groups. These findings support the hypothesis that BV’s immunostimulatory effects contribute to overall shrimp health, leading to improved feed intake and, consequently, enhanced digestive enzyme secretion81. Beyond its immune benefits, the anti-inflammatory properties of BV play a critical role in maintaining the integrity of the digestive tract epithelium. This preservation of gut health fosters optimal digestive function, enabling more efficient nutrient digestion and absorption, which ultimately boosts growth performance and overall shrimp health. Further supporting these results is the observation of normal histopathological structure in the hepatopancreatic tissues of shrimp fed BV-CSNPs diets. There was also a notable increase in the number and activity of B-cells, responsible for secreting digestive enzymes, and R-cells, which absorb and store lipids and glycogen.
This study found that shrimp fed diets containing BV-CSNPs showed a significant increase in the expression of several important immune-related genes: LGBP (lipopolysaccharide and β−1,3-glucan binding protein), PX (peroxinectin), and PPA (prophenoloxidase)82. LGBP, PX83and ppA74 are crucial immune molecules connected to the prophenoloxidase (proPO) system, a vital part of the shrimp’s innate immunity. The upregulation of the LGBP gene in the hepatopancreas of L. vannamei shrimp, as observed in this study, aligns with previous research highlighting LGBP’s critical role in shrimp defense. These findings are comparable with prior studies82,84 that found the upregulation of the LGBP gene in the hepatopancreas of L. vannamei and proposed that the LGBP has a crucial role in shrimp defense.
Similarly, the PX gene of shrimp-fed diets containing BV-CS NPs significantly increased, suggesting that an increase in PX can increase the biological activities of cell adhesion85opsonin86degranulation87peroxidase88and encapsulation89 of shrimp. Moreover, the transcription level of mtMnSOD in hepatopancreas was enhanced, indicating that the immune response depending on the ROS system played an important role in shrimp responses against fungal, bacterial, and viral infection, similar to other studies in shrimp90,91.
This study found that shrimp fed BV-CSNPs exhibited significantly lower mortality rates after exposure to Vibrio parahaemolyticus. This enhanced protection is likely due to dietary supplements improving the shrimp’s gut health and microbial community92,93which in turn positively influences the water microbiota.
The inclusion of BV-CS NPs in the diet not only improves survival during a V. parahaemolyticus challenge by altering the gut bacterial population but also enhances key immune molecules such as pattern recognition proteins, antimicrobial peptides, and prophenoloxidase. These immune enhancements collectively aid in bacterial clearance and improve infection tolerance, providing a robust defense against pathogens. Adding BV-CSNPs to the diet does more than just help shrimp survive Vibrio parahaemolyticus infections. It also improves their ability to fight off bacteria by changing the gut’s bacterial makeup. On top of that, it boosts important immune molecules like pattern recognition proteins, antimicrobial peptides, and prophenoloxidase, all of which help clear out bacteria and increase the shrimp’s tolerance to infections83. Further studies are needed to evaluate the economic feasibility of bee venom application in commercial aquaculture and to assess its potential impact on shrimp meat quality.
Conclusion
This study clearly shows that adding BV-CSNPs to shrimp diets at concentrations of 0.2 or 0.3 mg/kg significantly enhances several key aspects of their health. We observed notable improvements in growth performance, hemocyte count, and the activity of crucial immune enzymes like phenoloxidase and lysozyme. Digestive enzyme activity also increased, alongside positive changes in intestinal histomorphology, indicating better nutrient utilization and gut health. Furthermore, BV-CSNPs led to the upregulation of mRNA expression for important immune genes, including LGBP, PX, ppA, cytMnSOD, and mtMnSOD. These genetic changes suggest a stronger, more robust immune response. The findings indicate that BV-CSNPs may help mitigate immune-mediated oxidative stress, protecting shrimp cells from damage. Higher levels of BV-CSNP inclusion in the diet appear to strengthen overall immunity, potentially making shrimp more resilient to environmental stressors and pathogen infections. While the use of bee venom in aquaculture is a relatively new area, continued research is highly promising. BV-CSNPs could become a valuable nutritional and therapeutic supplement, especially as a nano-nutrition approach, to improve shrimp health and sustainability in aquaculture.
Data availability
All data to support the conclusions have been provided in the manuscript.
Abbreviations
- BV-CS NPs:
-
Bee venom loaded with nano-chitosan
- SOD:
-
Superoxide dismutase
- CAT:
-
Catalase
- HP:
-
Hepatopancreases
- SPF:
-
Pathogen-free
- IW:
-
Initial shrimp weight
- FW:
-
Final shrimp
- WG:
-
Weight gain
- FI:
-
Feed intake
- FCR:
-
Feed conversion ratio
- SGR:
-
Specific growth rate
- ADG:
-
Average daily gain
- SR:
-
Survival rate
- THC:
-
Total hemocyte count
- PO:
-
Phenoloxidase activity
- LYZ:
-
Lysozyme activity
- PA:
-
Phagocytic activity
- PI:
-
Phagocytic index
- BA:
-
Bactericidal activity
- ANOVA:
-
Analysis of variance
References
-
Eissa, M. E. H. et al. Dietary inclusion of Pediococcus acidilactici probiotic promoted the growth indices, hemato-biochemical indices, enzymatic profile, intestinal and liver histomorphology, and resistance of nile Tilapia against Aspergillus flavus. Anim. Feed Sci. Technol. 306, 115814. https://doi.org/10.1016/j.anifeedsci.2023.115814 (2023).
-
Han, S. M., Lee, K. G., Park, K. K. & Pak, S. C. Antimicrobial activity of honey bee venom against select infectious fish pathogens. North. Am. J. Aquaculture. 75, 445–448. https://doi.org/10.1080/15222055.2013.802264 (2013).
-
Rajeev, R., Adithya, K. K., Kiran, G. S. & Selvin, J. Healthy microbiome: a key to successful and sustainable shrimp aquaculture. Reviews Aquaculture. 13, 238–258. https://doi.org/10.1111/raq.12471 (2021).
-
Kumaran, M. et al. Is Pacific white shrimp (Penaeus vannamei) farming in India sustainable? A multidimensional indicators-based assessment. Environ. Dev. Sustain. 23, 6466–6480. https://doi.org/10.1007/s10668-020-00881-0 (2021).
-
Ahmed, N. Linking Prawn and shrimp farming towards a green economy in bangladesh: confronting climate change. Ocean. Coastal. Manage. 75, 33–42. https://doi.org/10.1016/j.ocecoaman.2013.01.002 (2013).
-
Belton, B., Padiyar, A., Ravibabu, G. & Gopal Rao, K. Boom and bust in Andhra pradesh: development and transformation in india’s domestic aquaculture value chain. Aquaculture 470, 196–206. https://doi.org/10.1016/j.aquaculture.2016.12.019 (2017).
-
Zou, Y. et al. Determination of the infectious agent of translucent Post-Larva disease (TPD) in Penaeus vannamei. 9, 741, (2020). https://doi.org/10.3390/pathogens9090741
-
Luo, K. et al. Evaluation of paraprobiotic applicability of Clostridium butyricum CBG01 in improving the growth performance, immune responses and disease resistance in Pacific white shrimp, Penaeus vannamei. Aquaculture 544, 737041. https://doi.org/10.1016/j.aquaculture.2021.737041 (2021).
-
Letchumanan, V., Chan, K. G. & Lee, L. H. J. F. i. m. Vibrio parahaemolyticus: a review on the pathogenesis, prevalence, and advance molecular identification techniques. 5, 705, (2014). https://doi.org/10.3389/fmicb.2014.00705
-
Aly, S. M., ElBanna, N. I., Elatta, M. A., Hegazy, M. & Fathi, M. Prevalence, molecular typing, antibiogram and histopathological changes of V. harveyi and V. parahaemolyticus isolated from Gilthead sea bream (Sparus aurata). Aquaculture and Fisheries, (2023). https://doi.org/10.1016/j.aaf.2023.11.002
-
Yang, H. et al. Efficacy of Hydroxy-L-proline (HYP) analogs in the treatment of primary hyperoxaluria in Drosophila Melanogaster. BMC Nephrol. 19, 167. https://doi.org/10.1186/s12882-018-0980-8 (2018).
-
Zhong, Y. et al. Investigation of the interaction between the fate of antibiotics in aquafarms and their level in the environment. J. Environ. Manage. 207, 219–229. https://doi.org/10.1016/j.jenvman.2017.11.030 (2018).
-
Bansemir, A., Blume, M., Schröder, S. & Lindequist, U. Screening of cultivated seaweeds for antibacterial activity against fish pathogenic bacteria. Aquaculture 252, 79–84. https://doi.org/10.1016/j.aquaculture.2005.11.051 (2006).
-
Tukmechi, A., Ownagh, A. & Mohebbat, A. In vitro antibacterial activities of ethanol extract of Iranian propolis (EEIP) against fish pathogenic bacteria (Aeromonas hydrophila, Yersinia ruckeri & Streptococcus iniae). Brazilian J. Microbiol. 41 https://doi.org/10.1590/S1517-83822010000400030 (2010).
-
de la Cruz-Cervantes, J. A., Benavides-González, F., Sánchez-Martínez, J. G., Vázquez-Sauceda, M. L. & Ruiz-Uribe, A. J. Propolis in aquaculture: A review of its potential. Reviews Fisheries Sci. Aquaculture. 26, 337–349. https://doi.org/10.1080/23308249.2018.1424798 (2018).
-
Rady, I., Siddiqui, I. A., Rady, M. & Mukhtar, H. J. C. L. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. 402, 16–31, (2017). https://doi.org/10.1016/j.canlet.2017.05.010
-
Baracchi, D., Francese, S. & Turillazzi, S. Beyond the antipredatory defence: honey bee venom function as a component of social immunity. Toxicon 58, 550–557. https://doi.org/10.1016/j.toxicon.2011.08.017 (2011).
-
Kim, H. et al. Bee venom mitigates cisplatin-induced nephrotoxicity by regulating CD4 + CD25 + Foxp3 + regulatory T cells in mice. (2013). https://doi.org/10.1155/2013/879845
-
Lim, B. S. et al. Effect of bee venom acupuncture on Oxaliplatin-Induced cold allodynia in rats. Evidence-Based Complement. Altern. Med. 369324 https://doi.org/10.1155/2013/369324 (2013).
-
Park, Y. C. et al. Long-Term effectiveness of bee venom acupuncture and physiotherapy in the treatment of adhesive capsulitis: A One-Year Follow-Up analysis of a previous randomized controlled trial. J. Altern. Complement. Med. 20, 919–924. https://doi.org/10.1089/acm.2014.0220 (2014).
-
Kabakci, D., Ürkü, Ç. & Önalan, Ş. Determination of the antibacterial effect of bee venom against rainbow trout pathogens and antibiotic resistance gene expression. Acta Vet. 73, 374–388. https://doi.org/10.2478/acve-2023-0028 (2023).
-
El-Bilawy, E. H., Mamdouh, I., Behiry, S. & Teiba, I. I. Evaluating the antibacterial efficacy of bee venom against multidrug-resistant pathogenic bacteria: Escherichia coli, Salmonella typhimurium, and Enterococcus faecalis. World J. Microbiol. Biotechnol. 41, 40. https://doi.org/10.1007/s11274-024-04248-9 (2025).
-
El Basuini, M. F. et al. Bee venom enhances performance and immune function in thinlip mullet: A promising approach for sustainable aquaculture. Fish Shellfish Immunol. 151, 109713. https://doi.org/10.1016/j.fsi.2024.109713 (2024).
-
Mohan, K. et al. Chitin, Chitosan and chitooligosaccharides as potential growth promoters and immunostimulants in aquaculture: A comprehensive review. Int. J. Biol. Macromol. 251, 126285. https://doi.org/10.1016/j.ijbiomac.2023.126285 (2023).
-
Parhi, R. Drug delivery applications of Chitin and chitosan: a review. Environ. Chem. Lett. 18, 577–594. https://doi.org/10.1007/s10311-020-00963-5 (2020).
-
Eissa, E. S. H. et al. Pumpkin seed oil-loaded Chitosan nanoparticles enhance growth, digestive enzymes, and immune response in whiteleg shrimp (Litopenaeus vannamei): impacts on histopathology and survival against Vibrio parahaemolyticus. Aquaculture Rep. 40, 102599. https://doi.org/10.1016/j.aqrep.2024.102599 (2025).
-
Selvasudha, N., Sweety, J. P., Dhanalekshmi, U. M. & Ruckmani, K. Springer Nature Singapore, in Marine Biomaterials: Drug Delivery and Therapeutic Applications (eds Sougata Jana & Subrata Jana) 97–137, (2022). https://doi.org/10.1007/978-981-16-4787-1_4
-
Im, E. J., Kim, S. J., Hong, S. B., Park, J. K. & Rhee, M. H. Anti-Inflammatory Activity of Bee Venom in BV2 Microglial Cells: Mediation of MyD88-Dependent NF-κB Signaling Pathway. Evidence-Based Complementary and Alternative Medicine 3704764, (2016). https://doi.org/10.1155/2016/3704764 (2016).
-
Leandro, L. F. et al. Antimicrobial activity of apitoxin, Melittin and phospholipase A 2 of honey bee (Apis mellifera) venom against oral pathogens. 87, 147–155, (2015). https://doi.org/10.1590/0001-3765201520130511
-
Al-Safar, M. & Salman, J. J. Antibacterial activity of bee venom against multidrug resistant Acinetobacter baumannii locally isolates. I J. R P S. 9, 1510–1514 (2018).
-
Ploydee, E. & Chaiyanan, S. Production of High Viscosity Chitosan from Biologically Purified Chitin Isolated by Microbial Fermentation and Deproteinization. International Journal of Polymer Science 162173, (2014). https://doi.org/10.1155/2014/162173 (2014).
-
De Queiroz Antonino, R. S. et al. Preparation and characterization of Chitosan obtained from shells of shrimp (Litopenaeus vannamei Boone). Mar. Drugs. 15 https://doi.org/10.3390/md15050141 (2017).
-
Hosni, A. et al. Therapeutic significance of thymoquinone-loaded Chitosan nanoparticles on streptozotocin/nicotinamide-induced diabetic rats: in vitro and in vivo functional analysis. Int. J. Biol. Macromol. 221, 1415–1427. https://doi.org/10.1016/j.ijbiomac.2022.09.048 (2022).
-
Amin, B. H., Ahmed, H. Y., El Gazzar, E. M. & Badawy, M. M. M. Enhancement the mycosynthesis of selenium nanoparticles by using gamma radiation. Dose-Response 19 (15593258211059323). https://doi.org/10.1177/15593258211059323 (2021).
-
Lee, M. J. C. & International, A. O. A. C. Official methods of analysis of AOAC international. Edited Patricia A. 359, 399300 (1995). 16th ed.
-
Du, L. & Niu, C. J. Effects of dietary substitution of Soya bean meal for fish meal on consumption, growth, and metabolism of juvenile giant freshwater Prawn. Macrobrachium rosenbergii. 9, 139–143. https://doi.org/10.1046/j.1365-2095.2003.00239.x (2003).
-
Liu, C. H. & Chen, J. C. Effect of ammonia on the immune response of white ShrimpLitopenaeus vannamei and its susceptibility to Vibrio alginolyticus. Fish Shellfish Immunol. 16, 321–334. https://doi.org/10.1016/S1050-4648(03)00113-X (2004).
-
Hernández-López, J., Gollas-Galván, T. & Vargas-Albores, F. Activation of the prophenoloxidase system of the brown shrimp Penaeus californiensis Holmes). Comp. Biochem. Physiol. Part. C: Pharmacol. Toxicol. Endocrinol. 113, 61–66. https://doi.org/10.1016/0742-8413(95)02033-0 (1996).
-
Lin, Y. C., Tayag, C. M., Huang, C. L., Tsui, W. C. & Chen, J. C. White shrimp Litopenaeus vannamei that had received the hot-water extract of Spirulina platensis showed earlier recovery in immunity and up-regulation of gene expressions after pH stress. Fish Shellfish Immunol. 29, 1092–1098. https://doi.org/10.1016/j.fsi.2010.09.002 (2010).
-
Rengpipat, S., Rukpratanporn, S., Piyatiratitivorakul, S. & Menasaveta, P. Immunity enhancement in black tiger shrimp (Penaeus monodon) by a probiont bacterium (Bacillus S11). Aquaculture 191, 271–288. https://doi.org/10.1016/S0044-8486(00)00440-3 (2000).
-
Adams, A. Response of Penaeid shrimp to exposure to Vibrio species. Fish Shellfish Immunol. 1, 59–70. https://doi.org/10.1016/S1050-4648(06)80020-3 (1991).
-
Bolann, B. J. & Ulvik, R. J. Improvement of a direct spectrophotometric assay for routine determination of superoxide dismutase activity. Clin. Chem. 37, 1993–1999. https://doi.org/10.1093/clinchem/37.11.1993 (1991).
-
Johansson, L. H. Håkan borg, L. A. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem. 174, 331–336. https://doi.org/10.1016/0003-2697(88)90554-4 (1988).
-
Ohkawa, H., Ohishi, N. & Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 95, 351–358. https://doi.org/10.1016/0003-2697(79)90738-3 (1979).
-
Borlongan, I. G. Studies on the digestive lipases of milkfish, Chanos chanos. Aquaculture 89, 315–325. https://doi.org/10.1016/0044-8486(90)90135-A (1990).
-
Livak, K. J. & Schmittgen, T. D. J. M. Analysis of relative gene expression data using real-time quantitative PCR and the 2 – ∆∆CT M.thod. 25, 402–408, (2001). https://doi.org/10.1006/meth.2001.1262
-
Duan, Y. et al. Changes in the intestine microbial, digestion and immunity of Litopenaeus vannamei in response to dietary resistant starch. Sci. Rep. 9, 6464. https://doi.org/10.1038/s41598-019-42939-8 (2019).
-
Eissa, E. S. H. et al. Protective effects of Chlorella vulgaris as a feed additive on growth performance, immunity, histopathology, and disease resistance against Vibrio parahaemolyticus in the Pacific white shrimp. Aquacult. Int. 32, 2821–2840. https://doi.org/10.1007/s10499-023-01298-y (2024).
-
Balcázar, J. L., Rojas-Luna, T. & Cunningham, D. P. Effect of the addition of four potential probiotic strains on the survival of pacific white shrimp (Litopenaeus vannamei) following immersion challenge with Vibrio parahaemolyticus. 96, 147–150 (2007). https://doi.org/10.1016/j.jip.2007.04.008
-
Eissa, M. E. H., Hendam, B. M., ElBanna, N. I. & Aly, S. M. Nano-chitosan encapsulation of bee venom: a potential therapeutic strategy for enhancing tilapia health and resistance to Aeromonas hydrophila. Aquacult. Int. 33, 368. https://doi.org/10.1007/s10499-025-02043-3 (2025).
-
Mabrouk, M. M. et al. Arthrospira platensis nanoparticles dietary supplementation improves growth performance, steroid hormone balance, and reproductive productivity of nile tilapia (Oreochromis niloticus) broodstock. PLOS ONE. 19, e0299480. https://doi.org/10.1371/journal.pone.0299480 (2024).
-
Said, R. M. et al. Impacts of dietary selenium nanoparticles from Spirulina platensis on growth performance, Physio-Biochemical components and alleviating effect against cadmium toxicity in Pacific white shrimp Litopenaeus vannamei. Catalysts 13 https://doi.org/10.3390/catal13111389 (2023).
-
Eissa, E. S. H. et al. Dietary effects of nano Curcumin on growth performances, body composition, blood parameters and histopathological alternation in red Tilapia (Oreochromis sp.) challenged with Aspergillus flavus. Fishes 8 https://doi.org/10.3390/fishes8040208 (2023).
-
Zhang, S. et al. Bee venom therapy: potential mechanisms and therapeutic applications. Toxicon 148, 64–73. https://doi.org/10.1016/j.toxicon.2018.04.012 (2018).
-
Wehbe, R. et al. Bee venom: overview of main compounds and bioactivities for therapeutic interests. Molecules 24 https://doi.org/10.3390/molecules24162997 (2019).
-
Khalil, A., Elesawy, B. H., Ali, T. M. & Ahmed, O. M. Bee venom: from venom to drug. Molecules 26 https://doi.org/10.3390/molecules26164941 (2021).
-
Billingham, M. E. J., Morley, J., Hanson, J. M., Shipolini, R. A. & Vernon, C. A. An Anti-Inflammatory peptide from bee venom. Nature 245, 163–164. https://doi.org/10.1038/245163a0 (1973).
-
Komi, E. A., Shafaghat, D., Zwiener, R. D. & F. & Immunology of bee venom. Clin. Rev. Allergy Immunol. 54, 386–396. https://doi.org/10.1007/s12016-017-8597-4 (2018).
-
Oršolić, N. Bee venom in cancer therapy. Cancer Metastasis Rev. 31, 173–194. https://doi.org/10.1007/s10555-011-9339-3 (2012).
-
Perumal Samy, R. et al. Antibacterial activity of snake, Scorpion and bee venoms: a comparison with purified venom phospholipase A2 enzymes. J. Appl. Microbiol. 102, 650–659. https://doi.org/10.1111/j.1365-2672.2006.03161.x (2007).
-
El-Hanoun, A., El-Komy, A., El-Sabrout, K. & Abdella, M. Effect of bee venom on reproductive performance and immune response of male rabbits. Physiol. Behav. 223, 112987. https://doi.org/10.1016/j.physbeh.2020.112987 (2020).
-
Han, S. M. et al. Effects of honeybee venom supplementation in drinking water on growth performance of broiler chickens. Poult. Sci. 89, 2396–2400. https://doi.org/10.3382/ps.2010-00915 (2010).
-
Han, S. M. et al. Effects of bee venom treatment on growth performance of young pigs. 37, 253–260, (2009). https://doi.org/10.1142/S0192415X09006813
-
Rady, I., Siddiqui, I. A., Rady, M. & Mukhtar, H. Melittin, a major peptide component of bee venom, and its conjugates in cancer therapy. Cancer Lett. 402, 16–31. https://doi.org/10.1016/j.canlet.2017.05.010 (2017).
-
Abd El-Wahed, A. A. et al. In Studies in Natural Products ChemistryVol. 60459–484 (Elsevier, 2019). ed Rahman Atta ur.
-
Jobling, M. & National Research Council (NRC). Nutrient requirements of fish and shrimp. Aquacult. Int. 20, 601–602. https://doi.org/10.1007/s10499-011-9480-6 (2012).
-
Duan, Y., Liu, Q., Wang, Y., Zhang, J. & Xiong, D. Impairment of the intestine barrier function in Litopenaeus vannamei exposed to ammonia and nitrite stress. Fish Shellfish Immunol. 78, 279–288. https://doi.org/10.1016/j.fsi.2018.04.050 (2018).
-
Eissa, E. S. H. et al. Potential symbiotic effects of β-1,3 glucan, and fructooligosaccharides on the growth performance, immune response, redox status, and resistance of Pacific white shrimp, Litopenaeus vannamei to fusarium Solani infection. Fishes 8 https://doi.org/10.3390/fishes8020105 (2023).
-
El-Banna, B., Abouzeid, A., El-Damrawy, S. & El-Rayes, T. Effect of bee venom on production performance and immune response of broilers. Egypt. J. Nutr. Feeds. 26, 91–100. https://doi.org/10.21608/ejnf.2023.298109 (2023).
-
Kim, D. H. et al. Evaluation of bee venom as a novel feed additive in fast-growing broilers. Br. Poult. Sci. 59, 435–442. https://doi.org/10.1080/00071668.2018.1476675 (2018).
-
Moreno, M. & Giralt, E. Three valuable peptides from bee and Wasp venoms for therapeutic and biotechnological use: melittin, Apamin and mastoparan. Toxins 7, 1126–1150. https://doi.org/10.3390/toxins7041126 (2015).
-
Ye, M. et al. Neuroprotective effects of bee venom phospholipase A2 in the 3xTg AD mouse model of alzheimer’s disease. J. Neuroinflamm. 13 https://doi.org/10.1186/s12974-016-0476-z (2016). 10.
-
Amparyup, P., Charoensapsri, W. & Tassanakajon, A. Prophenoloxidase system and its role in shrimp immune responses against major pathogens. Fish Shellfish Immunol. 34, 990–1001. https://doi.org/10.1016/j.fsi.2012.08.019 (2013).
-
Tassanakajon, A., Somboonwiwat, K., Supungul, P. & Tang, S. Discovery of immune molecules and their crucial functions in shrimp immunity. Fish Shellfish Immunol. 34, 954–967. https://doi.org/10.1016/j.fsi.2012.09.021 (2013).
-
Cerenius, L., Lee, B. L. & Söderhäll, K. The proPO-system: pros and cons for its role in invertebrate immunity. Trends Immunol. 29, 263–271. https://doi.org/10.1016/j.it.2008.02.009 (2008).
-
Mai, W. & Wang, W. Protection of blue shrimp (Litopenaeus stylirostris) against the white spot syndrome virus (WSSV) when injected with shrimp lysozyme. Fish Shellfish Immunol. 28, 727–733. https://doi.org/10.1016/j.fsi.2010.01.002 (2010).
-
Banks, B. E., Dempsey, C. E., Vernon, C. A., Warner, J. A. & Yamey, J. Anti-inflammatory activity of bee venom peptide 401 (mast cell degranulating peptide) and compound 48/80 results from mast cell degranulation in vivo. Br. J. Pharmacol. 99, 350–354. https://doi.org/10.1111/j.1476-5381.1990.tb14707.x (1990).
-
Del Rio, D., Stewart, A. J. & Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metabolism Cardiovasc. Dis. 15, 316–328. https://doi.org/10.1016/j.numecd.2005.05.003 (2005).
-
Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: analytical and biological challenges. Anal. Biochem. 524, 13–30. https://doi.org/10.1016/j.ab.2016.10.021 (2017).
-
Bava, R. et al. Therapeutic use of bee venom and potential applications in veterinary medicine. Veterinary Sci. 10 https://doi.org/10.3390/vetsci10020119 (2023).
-
Carrillo-Farnés, O., Forrellat-Barrios, A. & Guerrero-Galván, S. Vega-Villasante, F. A review of digestive enzyme activity in Penaeid shrimps. Crustaceana 80, 257–275 (2007).
-
Cheng, W., Liu, C. H., Tsai, C. H. & Chen, J. C. Molecular cloning and characterisation of a pattern recognition molecule, lipopolysaccharide- and β-1,3-glucan binding protein (LGBP) from the white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 18, 297–310. https://doi.org/10.1016/j.fsi.2004.08.002 (2005).
-
Sritunyalucksana, K., Wongsuebsantati, K., Johansson, M. W. & Söderhäll, K. Peroxinectin, a cell adhesive protein associated with the ProPO system from the black tiger shrimp, Penaeus monodon. Dev. Comp. Immunol. 25, 353–363. https://doi.org/10.1016/S0145-305X(01)00009-X (2001).
-
Zokaeifar, H. et al. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 33, 683–689. https://doi.org/10.1016/j.fsi.2012.05.027 (2012).
-
Chiu, C. H. et al. Immune responses and gene expression in white shrimp, Litopenaeus vannamei, induced by Lactobacillus plantarum. 23, 364–377, (2007). https://doi.org/10.1016/j.fsi.2006.11.010
-
Thörnqvist, P. O., Johansson, M. W. & Söderhäll, K. Opsonic activity of cell adhesion proteins and β-1,3-glucan binding proteins from two crustaceans. Dev. Comp. Immunol. 18, 3–12. https://doi.org/10.1016/0145-305X(94)90247-X (1994).
-
Johansson, M. W. & Söderhäll, K. A cell adhesion factor from crayfish haemocytes has degranulating activity towards crayfish granular cells. Insect Biochem. 19, 183–190. https://doi.org/10.1016/0020-1790(89)90090-5 (1989).
-
Johansson, M. W., Lind, M. I., Holmblad, T., Thornqvist, P. O. & Soderhall, K. Peroxinectin, a novel cell adhesion protein from crayfish blood. Biochem. Biophys. Res. Commun. 216, 1079–1087. https://doi.org/10.1006/bbrc.1995.2731 (1995).
-
Kobayashi, M., Johansson, M. W. & Söderhäll, K. The 76 kD cell-adhesion factor from crayfish haemocytes promotes encapsulation in vitro. Cell Tissue Res. 260, 13–18. https://doi.org/10.1007/BF00297485 (1990).
-
Cheng, W., Tung, Y. H., Liu, C. H. & Chen, J. C. Molecular cloning and characterisation of cytosolic manganese superoxide dismutase (cytMn-SOD) from the giant freshwater Prawn Macrobrachium rosenbergii. Fish Shellfish Immunol. 20, 438–449. https://doi.org/10.1016/j.fsi.2005.05.016 (2006).
-
Gómez-Anduro, G. A. et al. The cytosolic manganese superoxide dismutase from the shrimp Litopenaeus vannamei: molecular cloning and expression. Dev. Comp. Immunol. 30, 893–900. https://doi.org/10.1016/j.dci.2006.01.002 (2006).
-
Gainza, O. & Romero, J. J. S. r. Effect of Mannan oligosaccharides on the microbiota and productivity parameters of Litopenaeus vannamei shrimp under intensive cultivation in Ecuador. 10, 2719, (2020). https://doi.org/10.1038/s41598-020-59587-y
-
Noble, T. et al. Novacq™ improves survival of Penaeus vannamei when challenged with pathogenic Vibrio parahaemolyticus causing acute hepatopancreatic necrosis disease. 545, 737235, (2021). https://doi.org/10.1016/j.aquaculture.2021.737235
Acknowledgements
The authors extend their gratitude to the staff at the shrimp farm for their assistance with rearing and sample collection during the study.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
All experimental procedures were conducted in conformity with institutional guidelines for the care and use of laboratory animals in Suez Canal University, Egypt, and conformed to the College of Veterinary Medicine Commission for Care and Use of Laboratory Animals (Code No. SCU 2023077).
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Eissa, M.E.H., Hendam, B.M., ElBanna, N.I. et al. Bee venom loaded chitosan nanoparticles enhances growth, immunity and resistance to vibrio parahaemolyticus in pacific white shrimp. Sci Rep 15, 26179 (2025). https://doi.org/10.1038/s41598-025-11011-z
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-11011-z