Introduction
In order to perceive and respond adequately to the changing environment, plants have evolved a repertoire of sensory proteins, among which the transmembrane receptor-like protein kinase (RLK) is one of the most important family members, which can trigger the inter- and intra- cellular signal transductions independently trigger signals without further forming a functional complex1. RLKs are ones of a superfamily widespread in plants characterized by highly specific extracellular ligand-binding domain2,3. Among them, leucine-rich repeat RLKs (LRR-RLK) constitute the largest subfamily in Arabidopsis, with approximate 225 members2,4. In general, a typical LRR-RLK protein possesses an extracellular LRR domain, a single transmembrane domain (TM), and a cytoplasmic protein kinase domain. The LRR domain is equipped with the leucine-repeat unit, which is usually 24 amino acids long. The different number and organization of LRR repeats contribute to the diversity of LRR-RLKs, leading to the identification of multiple extracellular signaling ligands. The single-pass TM domain anchors the LRR-RLK to the cell plasma. LRRs perceive and interact with ligands, inducing the conformational change of the cytoplasmic serine/threonine protein kinase domain, resulting in signal transduction via phosphorylation of specific downstream proteins4,5. Although an increasing number of LRR-RLK members have been discovered in plant species, only a few have been functionally characterized, even in the most detailed Arabidopsis genome.
LRR-RLKs are involved in many physiological processes, including plant growth and development, phytohormone signaling, innate immunity, biotic and abiotic stress responses2. CLAVATA 1 (CLV1), one of the typical LRR-RLKs and specifically expressed around the organizing center (OC), which regulates the shoot apical meristem (SAM) homeostasis through recognition of CLV3 peptide ligand7,8,9.
Further, RECEPTOR-LIKE PROTEIN KINASE 2 (RPK2)/TOADSTOOL 2 (TOAD2) together with CLV3 regulates anther and embryo development, as well as SAM homeostasis10. TRACHEARY ELEMENT DIFFERENTIATION FACTOR RECEPTOR (TDR) and PHLOEM INTERCALATED WITH XYLEM (PXY) perceive the TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR (TDIF) peptide ligand and modulates the vascular meristem development11,12. HAESA (HAE) and HAESA-LIKE2 (HSL2) control distinct cell segregation events in lateral root and floral organ with their peptide ligand INFLORESCENCE DEFICIENT IN ABSCISSION (IDA)13. BRASSINOSTEROID INSENSITIVE 1 (BRI1) and its co-receptor BRI1-ASSOCIATED RECEPTOR KINASE 1 (BAK1) mediate the brassinosteroid (BR) signaling pathway and play important roles in multiple aspects of plant development14,15. Some LRR-RLKs participate in multiple signaling pathways. For instance, BAK1 serves as a co-receptor and plays an essential role in plant innate immunity response by interacting with FLAGELLIN SENSITIVE 2 (FLS2) and EF-Tu RECEPTOR (EFR)16. Furthermore, BAK1 and RPK1 are also involved in the ABA-induced stomatal closure in response to drought stress17. Recently, ERECTA (ER) was found to not only regulate stomatal development, pathogen defense, but also participate in salt and osmotic stress responses18,19,20.
Due to their structural complexity and functional diversity, LRR-RLK members have been widely identified in many species, such as Arabidopsis, rice, poplar, tomato, potato, cotton, common beans, Vitis amurensis, Citrus and Rosaceae species3,22,23,24,25,26,27,28,29. Kiwifruit has become one of the most popular cultivated fruit crops in China and even around the world because of its high economic and nutritional value. However, there has been no relevant research on the LRR-RLKs in Actinidia chinensis. In this study, the kiwifruit LRR–RLK family genes were identified and comprehensively studied through gene evolution, gene composition, conserved motifs, functional annotation, and expression profiles. These results may provide genome-scale insights into LRR–RLK genes in kiwifruit growth as well as responses to biotic and abiotic stresses.
Results
Identification and phylogenetic analysis of LRR–RLK genes in Kiwifruit
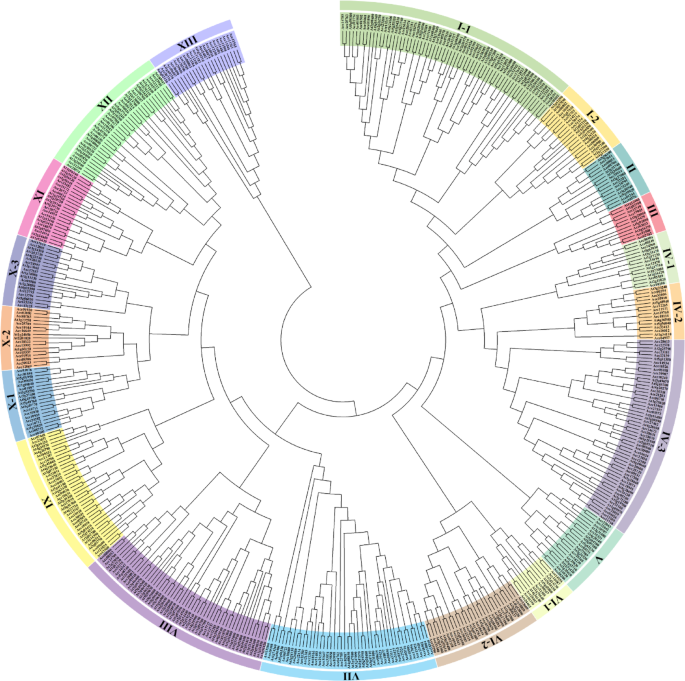
At present, a great number of LRR–RLK genes have been identified in Arabidopsis, rice, poplar, tomato, potato, and other plant genomes3,22,23,24,25,26,27,28,29. To identify the LRR–RLK genes in kiwifruit, genome-wide BLASTP search was employed using the protein sequences of 223 Arabidopsis LRR-RLKs as query4,30 based on the full-length protein sequences. The identified 307 kiwifruit LRR-RLKs were classified into 13 subclades and the corresponding AtLRR-RLK was used as a reference (Fig. 1).

Phylogenetic relationship analysis of LRR-RLK members in kiwifruit. The phylogenetic tree was drawn according to the alignment result of the full-length protein sequences by the neighbor-joining (NJ) method, with the following parameters: Poisson correction, partial deletion and bootstrap (1000 replicates). All AcLRR-RLK members and their Arabidopsis homologs were divided into 13 distinct subclades and labeled with different colors.
The gene size for each LRR-RLK subclade varied significantly. Among these subclades, I and IV had the largest number of AcLRR-RLKs, each contained more than 45 members, and the smallest subclade III with only 6 members. In general, the relative size of AcLRR-RLKs was similar to Arabidopsis within the subclade of I, II, III, IX, X and XII, and showed significantly difference for subclade IV, VII, VIII, XI and XIII (Table 1). For example, the number of LRR-RLKs in kiwifruit and Arabidopsis was 37 and 10 respectively in subclade VII, AcLRR-RLKs accounted for 12.1% and AtLRR-RLKs accounted for only 4.5% of its total LRR-RLKs. In kiwifruit, subclade VIII contained only 11 members comprised 3.6% of the total LRR-RLKs, while 42 AtLRR-RLKs representing 18.9% were included in Arabidopsis (Table 1). It was worth noting that the well-known Arabidopsis LRR-RLKs including CLV1, BAM1, BAM2, PXY, HSL2 and HAE which were involved in peptide perception and regulating plant growth and developmental processes are clustered together within the largest subclade of IV-3.
Chromosome localization, duplication and synteny analysis of LRR-RLK genes in Kiwifruit
The chromosomal distribution of the AcLRR–RLK genes was plotted based on the physical location in the kiwifruit chromosomes31. 307 AcLRR–RLK genes were widely and unevenly distributed on the 29 kiwifruit chromosomes with a maximum of 16 and a minimum of 4 genes located on chromosome 16 and chromosome 15, respectively (Fig. 2A). In addition to whole-genome duplication events, tandem and segmental duplications were the two major mechanisms in the expansion and functional diversification of gene families in plants. A single gene cluster was defined as a chromosomal region containing two or more AcLRR-RLK genes within 200 kb, and tandem duplication events were then calculated in clusters32,33,34. A total of 99 of the 307 AcLRR-RLK genes were grouped into 47 clusters over 25 chromosomes with no gene cluster located on chromosomes 7, 15, 28 and 29. Chromosome 3 contained the maximum of five clusters comprising eight members, and thirteen chromosomes contained only one cluster. The largest cluster comprised six genes on chromosome 19, and the most common cluster contained only two genes (Fig. 2A).
Furthermore, eighteen gene pairs from twelve clusters could be taken as genes derived from tandem duplication events and their full-length protein sequences similarity exceeds 70%32,34, accounting for approximately 8.8% (27/307) of the kiwifruit LRR-RLK genes. Subclades XIII and VII had 8 and 6 tandem duplication events, involving 10 and 9 genes, respectively, and the remaining 4 tandem duplication events were evenly distributed in subclades I-1, IV-3, V and X-3 (Fig. 2A). Among the 307 AcLRR–RLK genes, 63 pairs of 120 genes were derived from segmental duplication events by the syntenic analysis, which representing about 39.1% (120/307) of the kiwifruit LRR–RLK genes (Fig. 2B). The segmental duplication events were unevenly generated among subclades, with a maximum of 10 events in subclades IV-3 and XII, followed by subclades I-1, VI-2 and IX, however none event was found in subclade XIII (Fig. 2B). Overall, the above results suggested that segmental duplication events appeared to play a major role in the expansion of the kiwifruit LRR–RLK gene family. Detailed information of tandem and segmental duplication genes was listed in Table. S2-S4. A total of 47.9% (147/307) of LRR–RLK genes were found to be generated by tandem or segmental duplication events in kiwifruit, implying an important role of gene duplication events in the expansion of this family.

Chromosome localization and duplication analysis of the kiwifruit LRR-RLK genes. (A) 307 AcLRR-RLK genes were mapped to 29 kiwifruit chromosomes. Black lines indicate gene clusters and tandem duplicated genes are indicated in red. (B) Segmental duplication analysis of the kiwifruit LRR-RLK genes. The 63 putative segmental duplicated gene pairs of AcLRR-RLKs are linked by lines respectively.
In addition, the Ka values, Ks values and Ka/Ks values were calculated to evaluate the selection pressure between LRR-RLK paralogous genes in kiwifruit35. The Ka/Ks values in identified segmental duplicated gene pairs were all less than 1, among which the Acc22134/Acc26880 gene pair had the highest Ka/Ks value (0.63) (Table. S4). This result proved that significantly strong selective pressure was applied on the LRR-RLK gene family evolution, indicating that the deleterious mutations in the LRR–RLK gene family were tending to be eliminated, ensuring the preservation of essential functions within the gene sequences36.
Gene structure (exon-intron organization) analysis of LRR-RLK genes in Kiwifruit
To better understand the structural diversity and evolution of the LRR–RLK gene family, the gene structure of the identified 307 AcLRR–RLK genes was analyzed and plotted in Fig. S1, and the representative gene structures of each subclade were selected and displayed in Fig. 3. In terms of the exon-intron composition, 32 genes had no intron, while 142, 30, 11, 5, and 1 gene had 1, 2, 3, 4, and 5 introns, respectively. In addition, 33 AcLRR–RLK genes had 6–10 introns, 28 AcLRR–RLK genes had 11–15 introns, while the remaining 25 genes had more than 15 introns (Fig. S1). The gene structures of AcLRR–RLK were found to have great variability according to their subclades, and showed similar exon-intron organization pattern among closely related LRR–RLK genes within each subclade. All the members of subclade I-2, II, III, IV (1–3), VI (1–2), X-2, XIII and most members of subclade I-1 and VII were fairly simple and included no more than three introns over their full-length genomic sequences. In contrast, the members of subclade VIII, X-3, XI and XII exhibited complex exon-intron organization patterns with more than 10 introns (Fig. 3). Especially for XII, all 22 genes contained 10 introns and had the same exon-intron organization pattern. Subclades of V and IX showed significant variations in the number of introns. In particular, three genes in subclade V were composed of 26 introns, and this complex exon-intron organization pattern was also found in the Arabidopsis ER (At2g26330) gene, which was closely clustered with the three AcLRR–RLK genes (Fig. S1). In addition, comparison of LRR–RLK gene structures within the same subclade of kiwifruit and Arabidopsis, especially those closely clustered together, generally had similar exon-intron organization, indicating that they might have evolved from the same ancestral genes, and already had multiple exon-intron organization patterns. The conservation of LRR–RLK gene structures within subclade strongly suggested their close phylogenetic relationship during gene family evolution (Fig. 1).

Representative kiwifruit LRR-RLK gene structures of each subclade. The exon-intron organization of selected genes represent the majority members within subclade. Exons and introns are displayed by green boxes and black lines. The following parameters was considered in the representative gene selection, and the one with the value closest to the mean value was selected as the representative: length of the gene, length of the CDS, numbers of the exon, length of each exon, length of intron.
Protein structure and conserved motif analysis of LRR-RLKs in Kiwifruit
Physicochemical information of the AcLRR-RLKs were analyzed with ProtParam37, and the lengths of the members ranged from 351 to 1380 aa (Table 2, Table S1). The Acc03716 protein in subclade VII and Acc04973 protein in subclade XIII had the smallest (37.95 kDa) and largest (151.49 kDa) molecular weighted among AcLRR-RLKs, respectively. The theoretical isoelectric point (pI) of AcLRR-RLKs was between 4.8 and 9.73, and only 30.9% were basic proteins (pI > 7.0). Most AcLRR-RLKs consisted of signal peptide, accounting for 92.5% (284/307). Members of subclade I-2, II, III, IV-2, V, VI-1, VIII, X (2 & 3) and XII all contained signal peptide, and subclade VII had the least proportion of 70.3% (Table 2). In order to analyze the protein structure of kiwifruit LRR-RLKs, the full-length protein sequences were submitted to HMMER software for domain analysis, and highly conserved LRR and RLK domains were detected in all 307 kiwifruit LRR-RLKs (Fig. S2). Besides the LRR and RLK domains, all members in subclade VIII and XI possessed a malectin-like domain. Notably, the malectin-like domain in subclade VIII was located at the N-terminus before the LRR repeats and named malectin-like LRR-RLK38. Although malectin-like domains had been also detected in members of subclade XI, however they were located C-terminally before the RLK domain (Fig. S2).
The extracellular LRR domains varied in its number and order of LRR repeats that potentialize LRR-RLKs to perceive and relay various signals and provided the basis of their functional diversity. To further reveal the structural and functional diversification of LRR-RLK members, the extracellular region of AcLRR-RLKs were analyzed for their conserved motifs by the MEME motif search software. The composition and distribution of motifs were found in a same or high similar pattern between AcLRR-RLKs and AtLRR-RLKs within the same subclade, which might imply their close phylogenetic relationship and functional similarity (Fig. S2). In kiwifruit LRR-RLKs, 12 LRR motifs were identified and the distribution patterns of the 24 conserved amino acids were closely matched that plant LRR consensus sequence GxIPxxLxxLxxLxxLxLxxNxLx (Fig. 4). The most conserved amino acid residues Gly (G) at position 1, Pro (P) at position 4, Leu (L) at position 13, 16 and 18, and Asn (N) at position 21 in kiwifruit LRR motifs. Leu (L) at positions 3, 7 and 23 were often substituted by Ile (I) and Phe (F), respectively. In addition, other conserved residues were also observed in some positions in the specific AcLRR motifs. For example, the Glu (E) at position 6 in M10; Leu (L) at position 10 in M01 and M04 (Fig. 4).

Conserved LRR motifs in the LRR domains of kiwifruit LRR-RLK members. The figure was created with MEME motif search software. The conserved motifs of LRR-RLK proteins were identified with the maximum number of motifs and optimum motif width were set as 40 and 26–40 amino residues, respectively. An E-value was defined as the statistical significance of the motif by the software, and E-value of the motifs was 1.6e-906, 2.3e-798, 7.9e-695, 2.0e-584, 5.0e-396, 9.6e-380, 1.5e-347, 6.6e-287, 1.4e-251, 8.9e-239, 2.9e-198, and 2.1e-187 for M01 ~ M12. The occurrence of each motif was analyzed, and the value was 305, 306, 302, 262, 194, 233, 190, 145, 153, 138, 125, and 129 for M01 ~ M12.
Functional annotation and gene ontology analysis of AcLRR-RLKs in Kiwifruit
Functional annotation of 307 kiwifruit LRR-RLK members was conducted via gene ontology (GO) analysis for underlying their biological functions and mechanisms. The biological functions of identified proteins were divided into cellular components (CC), biological processes (BP), and molecular functions (MF) (Table. S5). In the CC category, major terms were associated with following terms: membrane system (plasma membrane and organelle membrane), cell part (symplast, organelle and cell junction) and extracellular region (Table S5). The most 10 enrichment terms were: perinuclear endoplasmic reticulum membrane (GO:1990578), secondary cell wall (GO:0009531), pollen wall (GO:0043667), multivesicular body membrane (GO:0032585), cell outer membrane (GO:0009279), plant-type cell wall (GO:0009505), extracellular matrix (GO:0031012), mitochondrial inner membrane (GO:0005743), organelle inner membrane (GO:0019866) and symplast (GO:0055044) (Fig. 5).
At the BP level, the AcLRR-RLKs were involved in multiple biological processes, and mainly associated with following pathways and terms: meristem maintenance and homeostasis (maintenance of root meristem identity, GO:0010078; embryonic meristem development, GO:0048508), growth and development process (secondary growth, GO:0080117; lateral growth, GO:0080190; floral meristem growth, GO:0010451; phloem development, GO:0010088), response to phytohormones (brassinosteroid homeostasis, GO:0010268; response to growth hormone, GO:0060416), response to biotic stresses (response to herbivore, GO:0080027; defense response to nematode, GO:0002215), response to abiotic stresses (response to mannitol, GO:0010555; response to freezing, GO:0050826) and immune responses (activation of immune response, GO:0002253) (Fig. 5).
The main terms in MF displayed three major functions, including binding: peptide hormone binding (GO:0017046); receptor serine/threonine kinase binding (GO:0033612); steroid binding (GO:0005496), hormone binding (GO:0042562); catalytic activity: transmembrane receptor protein kinase activity (GO:0019199); protein serine/threonine kinase activity (GO:0004674), protein serine/threonine phosphatase activity (GO:0004722) and signal transduction: molecular transducer activity (GO:0060089) (Fig. 5). These results implied that AcLRR-RLKs were mainly involved in membrane and extracellular localization, binding, signal transduction and catalytic functions, and might play important roles in plant growth and development as well as plant responses to phytohormones, biotic and abiotic stress.

Functional annotation of kiwifruit LRR-RLK members. GO enrichment plot shows the functional categorization of biological process, cellular component, and molecular function of kiwifruit LRR-RLK members.
Tissue-specific expression patterns of LRR-RLK genes in Kiwifruit
Gene expression profiles provide preliminary clues to figure out gene function. To explore the potential functions of AcLRR–RLK genes, expression patterns of the identified genes were analyzed using publicly available RNA-seq data retrieved from kiwifruit genome database (Tables S6-S17). The tissue-specific expression profiles showed that most AcLRR–RLK genes were considerable various expressed in different tissues (Table S6). For example, expression of Acc32006 (subclade IV-2), Acc22042 (subclade V), Acc00722 (subclade VII) and Acc11543 (subclade XIII) were not expressed in the detected tissues (Fig. 6A). In general, most of the AcLRR–RLK genes demonstrated a higher expression level in leaf, followed by root and stem, and few genes showed a relative high expression level in all tissues like Acc05159 (subclade IV-1), Acc20724 (subclade XI) and Acc06514 (subclade XI). Meanwhile, there are some AcLRR–RLK genes showed a significantly higher expression level in root and stem, such as Acc18038 (subclade IX) and Acc27829 (subclade IX) which implying a potential role in the vascular tissue development. While, there also were some AcLRR–RLK genes with very low expression level that merely detected which were mainly clustered into subclade I-1 (Fig. 6A). During the fruit development, most of the AcLRR–RLK genes demonstrated a higher expression level in the immature fruits (DAP20_immature) and decreased in mature green (DAP120_mature Green) and ripe fruits (DAP127_ripe). Meanwhile, some members of AcLRR–RLK showed a high and significantly increased expression level (> 2 folds) during fruit development, which indicating a potential positive role of these genes in the fruit ripening process, such as Acc21168 (subclade IV-3), Acc04826 (subclade IV-3) and Acc10594 (subclade VI-2) (Fig. 6A).

Expression profiles of kiwifruit LRR-RLK genes. (A) Tissue-specific expression of kiwifruit LRR-RLK genes. Hierarchically clustered heatmap of all AcLRR-RLK genes expressed across different tissues and organs. Leaf, root, stem, immature fruits (DAP20_immature), mature green (DAP120_mature Green) and ripe fruits (DAP127_ripe). Scale was set independently according to each subclade specific heatmap. (B) Expression profiles of kiwifruit LRR-RLK genes in the process of Psa infection. Hierarchically clustered heatmap of (A) down-regulated LRR-RLK genes in Ac and (B) up-regulated LRR-RLK genes in Ae in the process of Psa infection at 0DPI, 2DPI and 14DPI. Scales were set independently based on specific heatmaps for each kiwifruit variety. The expression data visualized with Heatmap illustrator in TBtools software using the self-normalized log converted TPM values. Significant expression differences were defined as more than 2-fold up-regulation or less than half down-regulation.
Expression profiles of LRR-RLK genes in response to biotic and abiotic stresses in Kiwifruit
In addition, the AcLRR–RLK genes were also participated in the response to biotic and abiotic stresses, and the transcriptome data obtained from kiwifruit under different stresses were compared. Biotic stress treatments included inoculation with Pseudomonas syringae, while abiotic stress treatments included salt, heat, and waterlogging. To screen Psa resistance candidate genes, the differentially expressed AcLRR–RLK genes in two varieties of A. chinensis and A. eriantha were analyzed (Table S7). There were 256 LRR–RLK genes differentially expressed in Ac, of which 242 genes were down-regulated (Fig. S3A). In contrast, only 62 differentially expressed LRR–RLK genes were identified in Ae, with 49 genes were up-regulated (Fig. S3B). Obviously, the total number of differentially expressed LRR–RLK genes in Ac was larger than that in Ae, but there were much more up-regulated genes in Ae. Then 23 LRR–RLK genes whose expression was down-regulated in Ac and up-regulated in Ae were selected and for further candidate gene screening (Fig. 6B). It was worth noting that the expression levels of the two genes Acc23789 and Acc24017 were high at 0DPI in Ac, but their expression levels decreased significantly at 2DPI and 14DPI (Fig. 6B). While in Ae, the expression of Acc23789 and Acc24017 rapidly increased and maintained at high levels during Psa infection (Fig. 6B), which suggested that Acc23789 and Acc24017 might play important roles in kiwifruit Psa infection.
Exposed to different abiotic stresses, 230 AcLRR–RLK genes were induced by at least one stress, and showed distinct expression patterns (Table S8-S17). Among them, 121 AcLRR–RLK genes were induced under salt stress (Fig. S4A), 96 under heat stress (Fig. S4B) and 179 under waterlogging stress (Fig. S4C). Specifically, there were 89 and 32 genes, 6 and 90 genes, 25 and 154 genes up-regulated and down-regulated by salt, heat and waterlogging stress, respectively (Fig. S5). In addition, many AcLRR–RLK genes displayed responses to multiple stress treatments. 34 AcLRR–RLK genes were induced by salt, heat and waterlogging stress, 7 genes were induced by salt and heat stress, 53 genes were induced by salt and waterlogging stress, 37 genes were induced by heat and waterlogging stress (Table S8). Meanwhile, some AcLRR–RLK genes were induced only by specific abiotic stress treatment. For example, Acc15749 and Acc23858 were only up-regulated under salt stress, Acc30909 and Acc24017 were only up-regulated under heat stress (Fig. S5). Overall, these results suggested that AcLRR–RLK genes might be involved in responses to biotic and abiotic stresses, emphasizing their pivotal roles in enhancing resistance to the adverse circumstances, especially of genes which were responsive to multiple stress treatments. However, more work is needed to examine whether these AcLRR–RLK genes could be concerned as candidate genes in future studies.
Discussion
The LRR-RLK family members are considered important regulators of plant growth and development as well as biotic and abiotic stress responses in various plant species. Despite their importance, a detailed understanding of kiwifruit LRR-RLK members has remained elusive. In this study, 307 LRR–RLK genes in kiwifruit were identified, which approximately accounted for 0.91% of the total coding genes in the genome39. The number of LRR–RLK genes in kiwifruit was moderate, more than that of apple, peach, tomato, common beans, sesame, and pine tree23,26,29,40,41, but less than the one of poplar, cotton, tea, peanut, rapeseed, and wheat22,27,42,43,44,45. During the expansion of kiwifruit LRR–RLK gene family, there were 18 pairs of tandem duplication and 63 pairs of segmental duplication, accounting for a total of 47.9% of the LRR-RLK family (Table S2-S4). It has been reported that kiwifruit has undergone two likely duplication events, which may explain the high proportion of AcLRR–RLK duplicated genes46.
Kiwifruit is an important economic fruit crop and faces constantly changing environmental conditions during its growth and development. As a receptor-like protein, LRR-RLK plays important roles in plant growth and development, phytohormone signaling, innate immunity, biotic and abiotic stress responses through intercellular and intracellular signaling2. For example, CLV1, BAM1 and BAM2 play critical roles in the maintenance and differentiation of the shoot apical meristem7,8,9, while TDR and PXY regulate the vascular development11,12. BRI1 and BAK1 are involved in the brassinosteroid signaling14,15. Several LRR-RLK members have been implicated in plant response to biotic and abiotic stresses, including BAK1, RPK1 and ER16,17,18,19,20. Currently, little is known about the function of kiwifruit LRR-RLK protein, and its response to biotic and abiotic stress also requires more research.
Kiwifruit bacterial canker incited by Psa is a major threat to kiwifruit yield and quality (production and development), leading to serious economic losses47,48. Currently, screening for resistance genes and cultivating resistant varieties are regarded as the most effective and safe measures to control the disease in kiwifruit. This study analyzed the expression patterns of LRR–RLK genes in two kiwifruit varieties, the highly susceptible “Hongyang” and the highly resistant A. eriantha49. A higher number of differentially expressed LRR–RLK genes were observed in Ac, with many LRR–RLK genes showed significant expression changes in Ac but not in Ae after exposure to Psa infection (Table S7). Interestingly, during Psa infection, Acc23789 and Acc24017 genes in Ac and Ae varieties showed significantly different expression trends (Fig. 6B). The expression levels of Acc23789 and Acc24017 genes were significantly reduced in Ac, whereas in Ae, Psa infection induced a significant increase in their expression levels. These results indicated that Acc23789 and Acc24017 genes might play an important role in kiwifruit resistance to Psa infection and could be used as candidate genes for Psa-resistant variety breeding. However, whether these candidate genes played a certain role in the process of kiwifruit Psa infection still requires further study in the future.
Moreover, crop growth and yield are also affected by a variety of abiotic stresses. Abiotic stresses such as salt, heat, and waterlogging stresses, also seriously affect plant growth and development. This indicates that LRR–RLK genes play an essential role in improving plant resistance to abiotic stresses in kiwifruit. Most (230/307) AcLRR–RLK genes were differentially expressed under salt, heat, and waterlogging stress (Fig. S4). When exposed to abiotic stress, the AcLRR–RLK genes showed distinct expression patterns. Some genes responded to all three stress treatments, some genes responded to two of the stress treatments, and some genes responded only to specific stress treatment (Fig. S5). For example, Acc23789 and Acc24017 genes were induced by salt, heat, and waterlogging stress treatments, which might be concerned as candidate genes for new variety breeding.
Conclusion
The present study performed a genome-wide analysis of LRR-RLK family genes in kiwifruit. A total number of 307 AcLRR-RLK members were identified and analyzed through evolutionary relationships, conserved motifs, functional annotation, tissue-specific expression, and expression patterns in response to biotic and abiotic stresses, which may supply different insights into the evolutionary conservation of this gene family and emphasize potential candidate genes for future breeding efforts.
Methods
Identification and phylogenetic analysis of Kiwifruit LRR-RLK genes
The LRR-RLK protein sequences of Arabidopsis were used as queries to perform a BLASTP search against the kiwifruit genome database (https://kiwifruitgenome.org/)39 with an E-value threshold of < 1e-10, other parameters kept default. All hit LRR-RLK proteins were analyzed for the presence of LRR, RLK and transmembrane domains using HMMER software (https://www.ebi.ac.uk/Tools/hmmer/)50 and TMHMM 2.0 software (https://services.healthtech.dtu.dk/services/TMHMM-2.0/)51. Then, the candidate AcLRR–RLK genes were obtained after confirmation the presence of at least one LRR-repeat, one single transmembrane and one kinase domain. To retrieve more sequences, the candidate AcLRR–RLK genes were subjected to a second round of screening by a BLASTP search against the kiwifruit genome database. The genomic and protein sequences of all AcLRR–RLK genes were retrieved from the kiwifruit genome database and used for further characterization. The multiple sequence alignment of AcLRR-RLKs and AtLRR-RLKs was carried out by ClustalX52 implemented in MEGA 1153, with the full-length amino acid sequences. The phylogenetic tree was generated with Neighbor-Joining (NJ) method30 with the following parameters: Poisson correction, partial deletion and bootstrap (1000 replicates).
Chromosomal localization, duplication and synteny analysis
The chromosomal localization of AcLRR-RLK genes were analyzed and illustrated by TBtools software (https://github.com/CJ-Chen/TBtools/releases)35. A single gene cluster was defined as two or more genes that localized within 200 kb distance or the neighboring genes interrupted by only one gene33,34. Tandem duplication events among AcLRR–RLK genes were defined as paralogous gene pairs that share similarities of protein sequences not less than 70% identity in a gene cluster32,34, and the identity among gene candidates was achieved by using Clustal Omega software (https://www.ebi.ac.uk/jdispatcher/msa/clustalo)54. A segmental duplication was analyzed by MCScanX software31. The visualization of duplication analysis was accomplished by using circle plotter module in TBtools35. The synonymous (Ks), non-synonymous (Ka) substitutions and Ka/Ks value was calculated with Ka/Ks Calculator module in TBtools35.
Gene structure, physiochemical property and conserved motif analysis
The exon-intron organizations of LRR–RLK genes were visualized with the online Gene Structure Display Server 2.0 (http://gsds.cbi.pku.edu.cn/)55. The physiochemical properties of AcLRR-RLK proteins, including amino acid number, molecular weight, and theoretical isoelectric point (pI), were calculated using ProtParam tools (https://web.expasy.org/protparam)37. The conserved motifs of LRR-RLK proteins were identified by MEME (Multiple Em for Motif Elicitation) online program (http://meme-suite.org/tools/meme)56, with the maximum number of motifs and optimum motif width were set as 40 and 26–40 amino residues, respectively.
Gene ontology (GO) annotation and gene expression analysis
The functional annotation of AcLRR-RLKs were analyzed with goatools and visualized by ggplot module in TBtools35. The kiwifruit transcriptomic data were retrieved from NCBI Sequence Read Archive (SRA) database (https://www.ncbi.nlm.nih.gov/sra) using SRA Toolkit (https://www.ncbi.nlm.nih.gov/home/tools/). Kallisto was introduced to achieved the analysis of the raw SRA transcriptomic data and expression levels in TPM (Transcripts Per Million) form were got57. The transcriptomic data of AcLRR–RLK genes for tissue-specific expression and response to biotic and abiotic stresses were achieved with data under accession number PRJNA328414 (leaf, root and stem; Pseudomonas syringae inoculation: Ac-0DPI, Ac-2DPI and Ac-14DPI; Ae-0DPI, Ae-2DPI and Ae-14DPI), PRJNA187369 (immature fruit: DAP20, mature green: DAP120 and ripe fruit: DAP127), PRJNA726156 (salt: 0.4% medium NaCl-0 h, NaCl-12 h, NaCl-24 h and NaCl-72 h), PRJNA796069 (heat: 50℃ Heat-0 h, Heat-2 h and Heat-4 h) and PRJNA765913 (waterlogging: Water-0d, Water-1d and Water-2d). Hierarchically clustered heatmaps were created with Heatmap illustrator in TBtools software35 using the self-normalized log converted TPM values. A significant expression difference was defined as over 2-fold change in up-regulation or half cutoff in down-regulation. All relevant data are included in the manuscript and supporting information.
Data availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
References
-
Cao, Y. et al. Genome-Wide identification and expression analysis of Kiwifruit Leucine-Rich repeat Receptor-Like proteins reveal their roles in biotic and abiotic stress responses. Int. J. Mol. Sci. 25, 4497 (2024).
-
Shiu, S. H. & Bleecker, A. B. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. U S A. 98, 10763–10768. https://doi.org/10.1073/pnas.181141598 (2001).
-
Shiu, S. H. et al. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant. Cell. 16, 1220–1234. https://doi.org/10.1105/tpc.020834 (2004).
-
Gou, X. et al. Genome-wide cloning and sequence analysis of leucine-rich repeat receptor-like protein kinase genes in Arabidopsis thaliana. BMC Genom. 11, 19. https://doi.org/10.1186/1471-2164-11-19 (2010).
-
Gish, L. A. & Clark, S. E. The rlk/pelle family of kinases. Plant. J. 66, 117–127. https://doi.org/10.1111/j.1365-313X.2011.04518.x (2011).
-
Ogawa, M., Shinohara, H., Sakagami, Y. & Matsubayashi, Y.
-
DeYoung, B. J. et al. The CLAVATA1-related BAM1, BAM2 and BAM3 receptor kinase-like proteins are required for meristem function in Arabidopsis. Plant. J. 45, 1–16. https://doi.org/10.1111/j.1365-313X.2005.02592.x (2006).
-
Hu, C. et al. A group of receptor kinases are essential for CLAVATA signalling to maintain stem cell homeostasis. Nat. Plants. 4, 205–211. https://doi.org/10.1038/s41477-018-0123-z (2018).
-
Wang, C. et al. Truncated BAM receptors interfere the apical meristematic activity in a dominant negative manner when ectopically expressed in Arabidopsis. Plant. Sci. 269, 20–31. https://doi.org/10.1016/j.plantsci.2018.01.003 (2018).
-
Kinoshita, A. et al. RPK2 is an essential receptor-like kinase that transmits the CLV3 signal in Arabidopsis. Development 137, 3911–3920. https://doi.org/10.1242/dev.048199 (2010).
-
Kondo, Y. & Fukuda, H. The TDIF signaling network. Curr. Opin. Plant. Biol. 28, 106–110. https://doi.org/10.1016/j.pbi.2015.10.002 (2015).
-
Zhang, H., Lin, X., Han, Z., Qu, L. J. & Chai, J. Crystal structure of PXY-TDIF complex reveals a conserved recognition mechanism among CLE peptide-receptor pairs. Cell. Res. 26, 543–555. https://doi.org/10.1038/cr.2016.45 (2016).
-
Kumpf, R. P. et al. Floral organ abscission peptide IDA and its HAE/HSL2 receptors control cell separation during lateral root emergence. Proc. Natl. Acad. Sci. U S A. 110, 5235–5240. https://doi.org/10.1073/pnas.1210835110 (2013).
-
Li, J. et al. BAK1, an Arabidopsis LRR Receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 110, 213–222. https://doi.org/10.1016/S0092-8674(02)00812-7 (2002).
-
Nam, K. H. & Li, J. BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell 110, 203–212. https://doi.org/10.1016/S0092-8674(02)00814-0 (2002).
-
Sun, Y. et al. Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex. Science 342, 624–628. https://doi.org/10.1126/science.1243825 (2013).
-
Osakabe, Y. et al. Leucine-rich repeat receptor-like kinase1 is a key membrane-bound regulator of abscisic acid early signaling in Arabidopsis. Plant. Cell. 17, 1105–1119. https://doi.org/10.1105/tpc.104.027474 (2005).
-
Lee, J. S. et al. Competitive binding of antagonistic peptides fine-tunes stomatal patterning. Nature 522, 439–443. https://doi.org/10.1038/nature14561 (2015).
-
Nanda, A. K., Habti, E., Hocart, A., Masle, J. & C. H. & ERECTA receptor-kinases play a key role in the appropriate timing of seed germination under changing salinity. J. Exp. Bot. 70, 6417–6435. https://doi.org/10.1093/jxb/erz385 (2019).
-
Godiard, L. et al. ERECTA, an LRR receptor-like kinase protein controlling development pleiotropically affects resistance to bacterial wilt. Plant. J. 36, 353–365. https://doi.org/10.1046/j.1365-313x.2003.01877.x (2003).
-
Wu, Y. et al. Genome-Wide expression pattern analyses of the < em > Arabidopsis Leucine-Rich repeat Receptor-Like kinases. Mol. Plant. 9, 289–300. https://doi.org/10.1016/j.molp.2015.12.011 (2016).
-
Zan, Y. et al. Genome-wide identification, characterization and expression analysis of populus leucine-rich repeat receptor-like protein kinase genes. BMC Genom. 14, 318. https://doi.org/10.1186/1471-2164-14-318 (2013).
-
Wei, Z., Wang, J., Yang, S. & Song, Y. Identification and expression analysis of the LRR-RLK gene family in tomato (Solanum lycopersicum) heinz 1706. Genome 58, 121–134. https://doi.org/10.1139/gen-2015-0035 (2015).
-
Liu, S. et al. Characterization, and functional investigation of VaHAESA from Vitis amurensis inoculated with Plasmopara viticola. Int. J. Mol. Sci. 19 https://doi.org/10.3390/ijms19041204 (2018).
-
Magalhães, D. M. et al. LRR-RLK family from two Citrus species: genome-wide identification and evolutionary aspects. BMC Genom. 17, 623. https://doi.org/10.1186/s12864-016-2930-9 (2016).
-
Sun, J., Li, L., Wang, P., Zhang, S. & Wu, J. Genome-wide characterization, evolution, and expression analysis of the leucine-rich repeat receptor-like protein kinase (LRR-RLK) gene family in Rosaceae genomes. BMC Genom. 18, 763. https://doi.org/10.1186/s12864-017-4155-y (2017).
-
Yuan, N. et al. Genome-wide identification and characterization of LRR-RLKs reveal functional conservation of the SIF subfamily in cotton (Gossypium hirsutum). BMC Plant Biol. 18, 185. https://doi.org/10.1186/s12870-018-1395-1 (2018).
-
Li, X. et al. Identification and characterization of LRR-RLK family genes in potato reveal their involvement in peptide signaling of cell fate decisions and biotic/abiotic stress responses. Cells 7, 120 (2018).
-
da Silva Dambroz, C. M., Aono, A. H., de Andrade Silva, E. M. & Pereira, W. A. Genome-wide analysis and characterization of the LRR-RLK gene family provides insights into anthracnose resistance in common bean. Sci. Rep. 13, 13455. https://doi.org/10.1038/s41598-023-40054-3 (2023).
-
Saitou, N. & Nei, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. https://doi.org/10.1093/oxfordjournals.molbev.a040454 (1987).
-
Wang, Y. et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 40, e49. https://doi.org/10.1093/nar/gkr1293 (2012).
-
Shiu, S. H., Li, W. H. & Origins, L. S. Expansions, and multiple losses of tyrosine kinases in eukaryotes. Mol. Biol. Evol. 21, 828–840. https://doi.org/10.1093/molbev/msh077 (2004).
-
Holub, E. B. The arms race is ancient history in arabidopsis, the wildflower. Nat. Rev. Genet. 2, 516–527. https://doi.org/10.1038/35080508 (2001).
-
Baloch, A. A. et al. Comparative genomics and evolutionary analysis of plant CNGCs. Biology Methods Protocols. 7 https://doi.org/10.1093/biomethods/bpac018 (2022).
-
Chen, C. et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 13, 1194–1202. https://doi.org/10.1016/j.molp.2020.06.009 (2020).
-
Huang, Z. et al. Molecular evolution, and expression divergence of the auxin/indole acetic acid and auxin response factor gene families in Brassica Rapa shed light on their evolution patterns in plants. Genome Biol. Evol. 8, 302–316. https://doi.org/10.1093/gbe/evv259 (2015). Retention.
-
Wilkins, M. R. et al. in In 2-D Proteome Analysis Protocols. 531–552 (eds Link, A. J.) (Humana, 1999).
-
Hok, S. et al. An Arabidopsis (malectin-like) leucine-rich repeat receptor-like kinase contributes to downy mildew disease. Plant. Cell. Environ. 34, 1944–1957. https://doi.org/10.1111/j.1365-3040.2011.02390.x (2011).
-
Yue, J. et al. Kiwifruit genome database (KGD): a comprehensive resource for Kiwifruit genomics. Hortic. Res. 7, 117. https://doi.org/10.1038/s41438-020-0338-9 (2020).
-
Nie, Z. et al. Comprehensive analysis of LRR-RLKs and key gene identification in Pinus massoniana resistant to pine wood nematode. Front. Plant Sci. 13, 1043261. https://doi.org/10.3389/fpls.2022.1043261 (2022).
-
Yan, W. et al. Comprehensive analysis of Sesame LRR-RLKs: structure, evolution and dynamic expression profiles under macrophomina Phaseolina stress. Front. Plant Sci. 15, 1334189. https://doi.org/10.3389/fpls.2024.1334189 (2024).
-
Zhang, Z., Ye, F., Hu, K., Luo, T. & Miao, Z. New insights into evolution and functional diversification of Camellia sinensis LRR-RLKs. Physiol. Mol. Biology Plants. 30, 851–866. https://doi.org/10.1007/s12298-024-01458-1 (2024).
-
Wang, Q. et al. Genome-wide identification of the LRR-RLK gene family in peanut and functional characterization of AhLRR-RLK265 in salt and drought stresses. Int. J. Biol. Macromol. 254, 127829. https://doi.org/10.1016/j.ijbiomac.2023.127829 (2024).
-
Liu, S. et al. Genome-wide identification and analysis of wheat LRR-RLK family genes following Chinese wheat mosaic virus infection. Front. Plant Sci. 13, 1109845. https://doi.org/10.3389/fpls.2022.1109845 (2023).
-
Song, M. et al. Genome-Wide survey of Leucine-Rich repeat Receptor-Like protein kinase genes and CRISPR/Cas9-Targeted mutagenesis BnBRI1 in Brassica napus. Front. Plant. Sci. 13, 865132. https://doi.org/10.3389/fpls.2022.865132 (2022).
-
Wang, J. P. et al. Two likely Auto-Tetraploidization events shaped Kiwifruit genome and contributed to establishment of the Actinidiaceae family. iScience 7, 230–240. https://doi.org/10.1016/j.isci.2018.08.003 (2018).
-
SCORTICHINI, M., MARCELLETTI, S., FERRANTE, P., PETRICCIONE, M. & FIRRAO, G. Pseudomonas syringae pv. actinidiae: a re-emerging, multi-faceted, pandemic pathogen. Mol. Plant Pathol. 13, 631–640. https://doi.org/10.1111/j.1364-3703.2012.00788.x (2012).
-
Vanneste, J. L. The scientific, economic, and social impacts of the new Zealand outbreak of bacterial canker of Kiwifruit (Pseudomonas syringae pv. actinidiae). Annu. Rev. Phytopathol. 55, 377–399. https://doi.org/10.1146/annurev-phyto-080516-035530 (2017).
-
Song, Y. et al. Comparative transcriptome analysis of resistant and susceptible Kiwifruits in response to Pseudomonas syringae pv. Actinidiae during early infection. PLoS One. 14, e0211913. https://doi.org/10.1371/journal.pone.0211913 (2019).
-
Finn, R. D., Clements, J. & Eddy, S. R. HMMER web server: interactive sequence similarity searching. Nucleic Acids Res. 39, W29–37. https://doi.org/10.1093/nar/gkr367 (2011).
-
Krogh, A., Larsson, B., von Heijne, G. & Sonnhammer, E. L. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J. Mol. Biol. 305, 567–580. https://doi.org/10.1006/jmbi.2000.4315 (2001).
-
Larkin, M. A. et al. Clustal W and clustal X version 2.0. Bioinformatics 23, 2947–2948. https://doi.org/10.1093/bioinformatics/btm404 (2007).
-
Tamura, K., Stecher, G. & Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. https://doi.org/10.1093/molbev/msab120 (2021).
-
Madeira, F. et al. The EMBL-EBI job dispatcher sequence analysis tools framework in 2024. Nucleic Acids Res. 52, W521–W525. https://doi.org/10.1093/nar/gkae241 (2024).
-
Hu, B. et al. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics 31, 1296–1297. https://doi.org/10.1093/bioinformatics/btu817 (2014).
-
Bailey, T. L. et al. MEME suite: tools for motif discovery and searching. Nucleic Acids Res. 37, W202–W208. https://doi.org/10.1093/nar/gkp335 (2009).
-
Bray, N. L., Pimentel, H., Melsted, P. & Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 34, 525–527. https://doi.org/10.1038/nbt.3519 (2016).
Funding
This work was financially supported by grants from Shandong Province Natural Science Foundation for Youths of China (No. ZR2022QC230, No. ZR2022QC226), Scientific Research Foundation of Dezhou University (2021xjrc105, 2021xjrc205) and Foundation for Creative Research Groups of Dezhou University (DZUQC202301).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, C., Tang, X., Liu, W. et al. Identification and expression analysis of LRR–RLK genes reveal their roles in plant development and stress responses in Kiwifruit (Actinidia chinensis). Sci Rep 15, 26346 (2025). https://doi.org/10.1038/s41598-025-12363-2
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12363-2