
Introduction
The global spread of Varroa destructor mites is one of the major pest invasions in the last decades1,2,3. The parasitic mite has infested honey bee (Apis mellifera) hives worldwide and caused rapid increases in colony mortality, especially in combination with the viruses it vectors4,5,6. Therefore, a vast majority of beekeepers choose to treat their colonies against V. destructor, often multiple times in a season7,8. According to the latest colony loss survey in New Zealand, over 97% of all bee hives received mite treatments in 2023 with synthetic miticides used in more than 80% of cases9,10.
The mite’s high virulence and fast reproduction require a regular treatment schedule11. Varroa destructor foundresses complete their reproductive cycle inside the honey bee brood cell in less than 12 days5,12. They invade brood cells containing 5th instar larvae just before capping, feed on larval haemolymph to sustain their energy demand and start laying their eggs approximately 3 days after invasion13. Offspring maturation to reproductive adults and mating of the new females occur inside the same brood cell within these 12 days5,14. Once the fully developed honey bee hatches, a new generation of mites will emerge with it. This high turnover enables V. destructor populations to grow quickly and population dynamics models predict an up to 12-fold increase within one brood season and even more in warmer climates where brood is continuously present15. To prevent exponential rise and keep levels beneath economic damage thresholds beekeepers intervene multiple times a year11,16,17.
Currently, a range of different tools are used in mite control, often in combination to maximise outcome and minimise their harmful side effects8,18. The most common form of chemical treatments are synthetic miticides such as pyrethroids, amitraz, or organophosphates, due to their efficacy and ease of application19. Yet, their lipophilic nature causes them to accumulate inside the hive more easily20. This buildup can be hazardous in different ways as it results in increased exposure and potential sublethal effects for bees and a higher risk of miticide resistance in V. destructor12,21. An increasingly popular alternative are organic treatments with essential oils (thymol, menthol) or organic acids (formic acid, oxalic acid)7,8,19. While these alternatives can be effective, treatment success is often less predictable and even organic products can affect brood and bee health negatively22. In addition to chemical approaches, hive management can also improve mite control. Screen bottom boards to trap mites, splitting colonies, planned brood breaks or drone brood removal can be valuable tools to reduce mite loads short-term or slow down their growth6,23. However, an effective, easily applicable and low-risk treatment does not exist yet12.
A novel approach is the use of biopesticides based on RNA interference (RNAi) technology to target honey bee pathogens such as deformed wing virus (DWV), Israeli acute paralysis virus (IAPV) or Varroa destructor mites24,25,26. RNA interference is a naturally occurring mode of gene silencing in eukaryotes that operates on a post-transcriptional level27,28. Short double-stranded RNA (dsRNA) fragments derived from the genome or from exogenous sources like viruses are bound to a large enzyme complex (RNA-induced silencing complex) that induces specific degradation of complementary RNA sequences28. RNAi-based treatments proved to be efficient against V. destructor mites when targeted against specific genes26,29,30. The Varroa destructor-specific dsRNA used in this trial is being developed under the brand name NORROA™ by GreenLight Biosciences, Inc., and the active ingredient’s common name is vadescana (CAS# 2,643,947–26-4). It targets the calmodulin gene in V. destructor and inhibits the mite’s egg production and thus reproduction31.
Most mite control therapeutics are applied repeatedly during the brood season or as long-acting formulas regardless of the beekeeper’s choice of treatment, resulting in an extended exposure of bees to these substances32,33. Therefore, assessing their toxicity to bees and the extent of sublethal effects are crucial steps on the way to a safe treatment. Many commercially available miticides have been evaluated for their topical and systemic toxicity, median lethal dose and impact on health and gene expression22,33,34. Yet, effects of novel mite control approaches such as dsRNA products and their impact on the lifespan and behaviour of honey bees have not been reported.
In this study we used radiofrequency identification (RFID) technology to monitor the movements of individual bees in and out of the hive. RFID chips attached to bees appear not to compromise their natural behaviour and can be automatically detected by electronic readers at the hive entrance35. RFID technology has been used successfully to monitor honey bees after direct administration of insecticides36,37,38.
Our main aim was to investigate the effects of V. destructor treatments on individual bees. Bees from hives treated with vadescana were compared to bees from hives without any mite treatment as well as hives treated with a conventional miticide (amitraz strips). To determine overall health levels and quantify potential sublethal effects we looked at three key health indicators. We recorded lifespan and survival of bees, their foraging behaviour and measured the development of mite populations over time with bottom board counts. For our trial, all treatments were applied to the hives according to manufacturer’s instructions to achieve concentrations comparable to actual field conditions. This experiment also illuminated patterns of drifting and robbing between apiaries and hives.
Material and methods
Apiary sites and hives
The trial was conducted over a total of 17 weeks between January and May 2024 with 9 hives in two different apiary sites 300 m apart on the grounds of Victoria University in Wellington, New Zealand (latitude: −41.290, longitude: 174.767). Hives at the same apiary site were placed about 2 m apart, so they could be connected to the same RFID system. At the start of the trial in late January, bee hives were brought onto campus from a commercial apiary (Dave’s bees, Ashurst, New Zealand). At this time, all hives consisted of 8–10 frames of bees and a laying queen and had not been treated for mites for at least 2 months prior to the start of the experiment. All hive boxes were placed on top of a wooden bottom block, so bottom boards could be inserted underneath each hive to monitor mite infestations.
All weather data for these apiary sites was sourced from New Zealand’s National Climate Database (Cliflo, https://cliflo.niwa.co.nz). Information was obtained from the nearest weather station within 1 km of the apiary sites (Kelburn automated weather station: latitude: −41.284, longitude: 174.767) and measured daily at 9 am local time.
Treatment groups
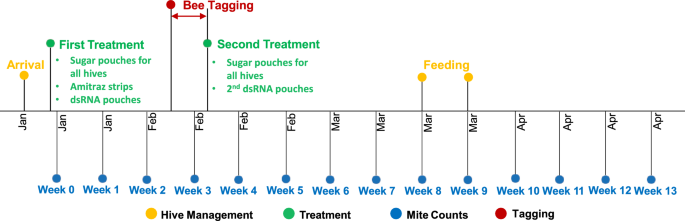
Hives were divided into three different treatment groups and randomly assigned one of the treatments. We would have preferred to have more than 3 replicate hives per treatment, but were logistically limited. One group of 3 hives was did not receive any mite treatments at all and served as the negative control, a second group of 3 hives was treated with miticide strips containing amitraz as a conventional miticide control group and the third group of 3 hives was treated with vadescana at a concentration of 4 g/L. Vadescana is the active ingredient in a product owned, developed and synthesized by GreenLight Biosciences, Inc. and includes a 402 base pair dsRNA sequence complementary to the calmodulin-like gene in Varroa destructor (GenBank accession number XM_022799184.1)39. This gene encodes the Ca2+ binding protein calmodulin, which plays a central role in calcium signaling40. The product’s brand name is NORROA™ and vadescana is formulated and administered within a ~ 60% sucrose-based solution in a sealed pouch with perforated openings (Supp. Mat. 1). This pouch is placed inside the hive on top of the brood box or on the bottom board, so bees take up the vadescana solution and then distribute it throughout the brood chamber to developing larvae. At the start of the trial (week 0), all hives received a 500 mL pouch with ~ 60% sucrose-based solution identical to the formulation of NORROA™ but without the active ingredient vadescana dsRNA. This application ensured that all hives received the same nutritional source, but only 3 hives were given pouches containing vadescana (Fig. 1). Miticide hives were treated with two amitraz strips containing 500 mg amitraz/strip (Apivar®, Veto-pharma S.A.S., France) per brood box according to manufacturer’s instructions in addition to the sucrose-based solution. Three weeks later, all colonies were given a second pouch with either just the ~ 60% sucrose-based solution or the vadescana-treated hives received their second dose as per the manufacturer’s instructions.

Timeline of the hives’ arrival, mite treatments and following mite counts. The experiment was conducted during the austral summer and autumn from January to May. Hives were treated with amitraz strips and dsRNA in week 0 and again with dsRNA in week 3. In the same week newly-hatched bees were tagged with RFID chips. Mite counts were performed on a weekly basis.
RFID tagged bees
To track an individual bee’s activity and movements, bees were tagged with RFID chips (Mini-Transponder mic3®Q1.6, microsensys GmbH, Erfurt, Germany), each holding a unique identification (UID) number (Fig. 2). The dimensions of the transponder were 1.6 × 1.6 × 0.4 mm3 and its weight < 5 mg to ensure they did not interfere with the bee’s normal behaviour41.

RFID tagged bees (left): 150 newly-hatched bees per hive were tagged with RFID chips to track their activities and RFID readers (right): Readers were mounted at the entrance of each hive to register all bee movements.
In week 3, when a cohort of bees that would have developed from eggs while being exposed to the mite control treatments was ready to emerge, one frame of capped brood from each hive was taken into the lab and stored at 34 °C in a temperature-controlled room overnight. The next morning, newly hatched bees were collected directly from the frame. Bees were anaesthesised at −20°C for 5 min and placed on ice for the RFID tagging procedure. All bees had a drop of cyanoacrylate-based glue (Shelleys®, Australia) applied to the dorsal part of their thorax and a RFID transponder was placed on top in between their wings. All tags were initialized, assigned to their individual hive and checked for their function with a handheld RFID-Penreader (iID PENsolid UHFcc, microsensys GmbH, Erfurt, Germany). To organise the bees’ UIDs into groups and assign their date of birth, the iID®BEE suite software (microsensys GmbH, Erfurt, Germany) was used in combination with the penreader. This process was repeated for each hive until approximately 150 bees per hive had been tagged with RFID chips. After the bees had fully regained consciousness, they were placed back into their hives within 4 h of being tagged.
Radiofrequency identification (RFID)-based monitoring
For continuous monitoring of the hives and movements of the individual bees, all hives were equipped with RFID technology (iID®BEEscience professional Bundle v313.2D, microsensys GmbH, Erfurt, Germany). Each of the hives was equipped with a RFID reader (iID® Reader Block – model roof antenna holder) mounted on top of a small platform at the hive entrance (Fig. 2). These reading devices detect and identify all RFID chips that pass the entrance. All logs include the chip’s unique identification number (UID), an exact timestamp and the direction of the movement. All readers at an apiary site were connected to a central RFID controller (iID®controller) with 1 GB RAM and a 16 GB USB stick (intenso®, Germany) for data storage.
Hive management
For the duration of the trial, hives were disturbed as little as possible, and no boxes were added or removed. After the honey flow had ended, all hives were fed twice (week 8 and 9) with 2 L of ~ 50% sucrose solution each time as a diet supplement. Sugar solutions were given in a top feeder, so bees were not disturbed.
To monitor the development of V. destructor populations without disturbing the hives, weekly mite fall counts were performed. Cardboard sheets with a size of 505 mm × 405 mm which equals the base of a NZ Langstroth hive box were sprayed with canola oil spray (Woolworths®, NZ) to make them more adhesive for mites. These sticky bottom boards were placed underneath all hives for 24 h, then all mites present on the bottom board were counted and counts from all hives of a treatment were averaged42. While there are better methods for sampling mites, we only used bottom boards because they are the least invasive technique to obtain mite counts and we wanted to avoid sampling or injuring any RFID-tagged bees during the course of the experiment. Any stress or disturbance from alternative mite sampling techniques could have influenced their lifespan and foraging behaviour and therefore compromised our results23,43.
Data analysis
All statistical analyses were performed using R version 4.3.1 (R Development Core Team, 2023) and RStudio (RStudio/2023.06.1, Posit Software PBC, USA, 2023). Data collected by the RFID controllers was saved for every day of the trial and imported directly into R, but due to power outages datasets remained incomplete on 2 occasions. Detections were saved by the readers in 15 s intervals to account for multiple detections during the same bee passage and avoided a long series of consecutive detections. Metadata containing information on birthday and home hive was added and combined with daily records. No limits for trip duration were set and no RFID records were removed from the collection.
All available data was categorised into logging data without direction information (reader entry: unknown) used for general activity and lifespan analysis and data with direction information (reader entry: departing/arriving). The latter were used to generate a list of trips each bee made over her lifetime.
The lifespan of all individuals was determined as the time between their date of birth and the last time they were detected by a RFID reader. For survival analysis this lifespan data was used to generate Kaplan–Meier survival plots with the survival and survminer packages (survival 3.6–4, survminer 0.4.9)44,45.
Foraging analysis was performed using only data containing direction information. In our algorithm, an individual trip was defined as a corresponding pair of data entries, the first ‘departing’ and the following ‘arriving’ of the same UID. Number of trips per bee and trip duration were calculated from these individual trips. Foraging onset and foraging lifespan were derived from the same dataset using the first trip of a bee as her foraging onset date and the time difference between first and last foraging trip for the foraging lifespan calculations. Foraging lifespan in this broader definition encompasses all out of-hive tasks of a honey bee, including a short pre-foraging stage with orientation or learning flights as well as actual foraging activity46,47.
For all datasets the Shapiro–Wilk test was used to determine whether they form a normal distribution. Then a Kruskal–Wallis test was performed for nonparametric datasets to discover significant differences (p < 0.05) in between groups. If differences were found to be significant, a multiple comparison test after Kruskal–Wallis was done for pairwise comparisons using the pgirmess package in R (pgirmess 2.0.2)48.
Since all bees were assigned to their home hive, trips to other hives could be recorded. If bees moved permanently into another hive and did not return to their home hive, they were counted as drifting bees49,50. If they returned from their visit back into their home, they were classified as robbing bees51. Network plots for data on robbing and drifting were created using the igraph package (igraph 2.0.3.9019)52.
For the analysis of mite loads over time across the different treatment groups, a linear regression model was fitted using the lm function from the lme4 package53. In the model treatment and weeks since trial start were considered as factors. Subsequently, the emmeans package was used to create pairwise comparisons and estimated marginal means which were necessary to identify significant differences in the post-hoc analysis54.
Results
Hives were monitored for a total of 105 days during the austral summer and autumn, between February and May 2024. At the end of the trial, all hives were found to be alive and still active. A total of 1396 bees from 9 hives had been tagged with RFID chips and out of those 1327 bees were detected at least once by the RFID readers (Supp. Mat. 2). A total of 977 bees (69.99%) lived to forage at least once in their lifetime. The second antenna on one of the RFID readers (hive yellow 3) was found to be faulty and no direction information was gathered from these records, so this hive was later excluded from the foraging analysis and only data from bees of untreated hives 1 and 2 (n = 191) was included in the analysis.
Lifespan
Survival times were calculated for all bees (n = 1327) that were detected at least once in their hive. The average lifespan differed significantly between treatment groups (chi-squared = 45.4, df = 2, p < 0.001), although there was also significant variation within the treatment group between individual hives (Fig. 3). Bees (n = 439) in untreated hives had the shortest survival spans with an average of 21.8 days [95% CI = 20.5- 23.2]. Bees from vadescana-treated hives (n = 430) ranged intermediate with 25.4 days [95% CI = 24.0- 26.8] and bees from amitraz treated hives (n = 458) had the longest average lifespans with 28.7 days [95% CI = 27.4- 30.1].

Kaplan–Meier plot displaying the survival rate of bees assigned to different treatments: Lifespans in days were recorded for all RFID-tagged bees in the three treatment groups: Untreated control (n = 439), amitraz treated group (n = 458) and vadescana dsRNA group (n = 430). Shaded areas show values within the 95% confidence interval.
Differences in survival times between the treatment groups were most apparent in the first 30 days of the trial. The lower average lifespan times in the control group were primarily driven by higher initial mortality rates (Fig. 3). There was variability in survival in all treatment groups, with some individual bees that lived for more than twice the average lifespan. The oldest bee in the trial survived for 86 days as an adult.
Activity
Daily movements were recorded from bees in all hives to ensure that the RFID equipment was functioning and active throughout the entire trial. An activity curve was combined with weather data from a nearest weather station (700 m away) to correlate activity with external factors such as low outside temperatures or rainfall. Drops in activity can be seen to correspond with drop in temperature or rainfall (Fig. 4).

Detections per day correlated with daily weather data: The graph on the bottom measured the daily activity of bees in detections of RFID tags per day as seen on the y-axis. Detections were summarised per treatment group. dsRNA treated hives are displayed in green, miticide treated hives in red and untreated control hives in yellow. The top curve shows temperature (red) and rainfall data (blue). Days with low bee activity corresponded to drops in temperature or rainfall.
Activity levels generally reflect the lifecycle of a worker bee and tagged bees were shown to pass through these typical behaviour phases55. Little activity was detected for the first 10 days, likely when the bees mainly performed in-hive tasks as nurse or house bees. Afterwards activity increased as they set off for their first orientation flights outside the hive46. This rise in movement was followed by a very active foraging period of 4 weeks before activity started to decrease indicating that bees were dying off55,56. Untreated bees were seen to pass through those life stages faster, as their activity peaked early on and then started to decline (Supp. Mat. 3). Over 98% (n = 433) of the untreated bees had died after 7 weeks. Activity in vadescana-treated hives was recorded for 11 weeks and the last bee of an amitraz-treated hive died after 14 weeks of recording.
Foraging behaviour
For foraging analysis, only data with direction information was analysed. When filtering the available data, the reader of one of the untreated hives (Y3) was found to be faulty, so this hive was excluded from the foraging analysis. In total 31,257 trips of 977 bees were included in this analysis.
The bees from vadescana-treated hives started foraging the earliest with 8.5 days of age [95% CI = 8.20–8.79], amitraz-treated bees were intermediate with 10.4 days [95% CI = 9.88- 10.9] foraging onset and untreated bees had the latest foraging onset age with 12.6 days [95% CI = 11.6- 13.5] (Fig. 5). The differences between all groups were found to be statistically significant (chi-squared = 52.085, df = 2, p < 0.001). Total duration of the foraging period was similar for groups that received mite treatments, the hives treated with vadescana had a foraging lifespan of 19.3 days [95% CI = 18.1- 20.5] and hives with amitraz strips had a foraging lifespan of 20.9 days [95% CI = 19.8- 22.1]. For untreated hives the recorded foraging lifespan was the shortest with 14.2 days [95% CI = 12.7- 15.6] and their time spent as forager was found to be significantly shorter (chi-squared = 45.195, df = 2, p < 0.001) than for the other two groups. There was no statistically significant difference between the amitraz and the vadescana group (p = 0.2). Both treatment groups included outliers that foraged for over 50 days, more than twice the average foraging period.

Foraging onset age (left) and foraging lifespan (right): Foraging onset is defined as the first time a bee takes a flight outside her hive and foraging lifespan as the timespan between her first and last flight. Both values are displayed in days on the y-axis. Boxes range from first to the third quartile, whiskers extend to 1.5 times the interquartile range and values outside this range are plotted as individual points. Green boxes represent dsRNA treated bees (n = 381), red boxes miticide-treated bees (n = 405) and yellow boxes untreated bees (n = 191). Significance brackets indicate these groups were significantly different (p < 0.05). Asterisks indicate significance levels: p ≤ 0.05: *, p ≤ 0.01: ** and p ≤ 0.001: ***.
In terms of foraging effort bees from vadescana hives did the most trips (41.1 trips per bee [95% CI = 36.9- 45.2]) and had the shortest trip lengths of 59.7 min [95% CI = 48.5- 71.0] on average (Fig. 6). Amitraz-treated hives ranked in the middle with 30.4 trips per bee [95% CI = 27.3- 33.4] and 89.0 min average trip length [95% CI = 71.2- 106.9]. Untreated bees from control hives were the least productive in this study with only 17.3 trips per bee [95% CI = 14.1- 20.5] and had the longest foraging time with an average time of 371.5 min [95% CI = 271.3- 471.6].

Average length of foraging trips per bee (left) and number of foraging trips per bee (right): Trip length is displayed in hours on the y-axis (left). Note the axis has been compressed above 12 h to incorporate longer trips lasting over 12h. Boxes range from first to the third quartile, whiskers extend to 1.5 times the interquartile range and values outside this range are plotted as individual points. Green boxes represent dsRNA treated bees (n = 381), red boxes miticide-treated bees (n = 405) and yellow boxes untreated bees (n = 191). Significance brackets indicate these groups were significantly different (p < 0.05) and asterisks indicate significance levels: p ≤ 0.05: *, p ≤ 0.01: ** and p ≤ 0.001: ***.
The average number of trips per bee was significantly different between all treatment groups (chi-squared = 84.662, df = 2, p < 0.001). Trip duration was found to be significantly longer only for the untreated control group compared to both treatment groups (chi-squared = 80.332, df = 2, p < 0.001). Untreated hives had far more outliers and some of the untreated bees showed unusual foraging behaviour with few and very long trips, especially over 24 h which indicates they did not return to their hives overnight.
Movements between hives
Bees were found to move between all hives and both apiary sites. About 8% (n = 113) of all tagged bees were found to visit other hives than their home hive. If bees abandoned their home hive permanently, they were categorized as drifting bees whereas bees that came back to their home hive after a trip to another hive were classified as robbing bees.
Drifting bees (n = 50) were found to move hives earlier in their life at an average of 18.1 days [95% CI = 14.4—21.7], but robbing behaviour (n = 63) could start at any time, mostly later in their lives at an average age of 26.9 days [95% CI = 23.7—30.1]. This age difference between drifting and robbing bees was found to be significantly different (chi-squared = 11.479, df = 1, p < 0.001) (Supp. Mat.4).
On a hive level (counts of more than 1 bee) drifting occurred only between neighbouring hives at the same apiary site and generally on lower levels than robbing (Fig. 7a). This pattern indicates drifting might be common in between hives at the same apiary sites and there is an exchange of bees (about 3.5% here) between hives, but it is more a series of individual bees randomly moving to a neighbouring hive than a coordinated swarm phenomenon.

Movement of bees in between hives: Apiary 1 and 2 display our 2 apiary sites and hive boxes represent individual hives. Arrows connecting hives show bees moving in between those hives. Bees were found to drift only to hives within their apiary site (a), but robbing bees would even travel in between apiaries (b). Robbing activities originated mainly from one hive (Red 3). Illustrations were created with BioRender.com.
Robbing activities, in contrast, originated mainly from one hive (Red 3) and they were detected in 7 out of 8 other hives, even at the other apiary site (Fig. 7b). When comparing data on robbing behaviour with other datasets such as foraging trip data and lifespans, it was found that some of the robbing trips occurred on the same day and at the same hive and thus were part of more coordinated robbing events (Supp. Mat. 5 and 6). On days of these robbing events the number of robbing trips and foraging trips in total was higher than usually, bees were unusually active and an unusual number of bees died.
Mite counts
Weekly mite counts from bottom boards were performed to investigate the effects of different treatments on the mite population over the study period (Fig. 8). While this method of mite monitoring posed no risk to tagged bees and caused little if any disturbance to the hives, it is known to be less accurate than other diagnostic tools23,43. In week 0 at the beginning of the trial, all treatments had moderate mite counts of about 5 mites/100 bees on average. Mite counts remained below these initial counts on average until week 8 when untreated hives had an average of 6.67 mites/100 bees. Mite levels in all hives continued to rise and in week 10 even vadescana treated hives had an average of 5.33 mites/100 bees.

Total mite counts from sticky bottom boards: Weekly mite counts were done over the trial period to monitor population development. Varroa treatments were applied in week 0 (pouches for dsRNA group and strips for miticide group) and week 3 (only dsRNA pouches). Total counts are displayed on the y-axis on a logarithmic scale and treatment weeks are shown on the x-axis. dsRNA treated hives had lower mite numbers than the untreated control group throughout the trial.
A linear regression model including the main effects of “Treatment” (F = 4.12, df = 2, p = 0.019) and “Week” (F = 15.83, df = 13, p < 0.001) on V. destructor counts found both to have significant effects on bottom board mite counts. Further analysis with pairwise comparisons found that the mite counts in untreated hives were significantly higher than in vadescana treated hives (p = 0.014). Amitraz-treated hives had intermediate mite counts that were not significantly different to either the untreated hives or vadescana treated hives (p > 0.3). The effects of “Week” on mite counts were also found to be significant from week 11 onwards (p < 0.05). This development indicates that treatments were effective in controlling mite levels for about 10 weeks and treatment effects were waning afterwards. The exponentially rise in mite numbers is also typical for the season, as bee numbers decrease in autumn, but mite loads increase heavily.
Discussion
In this study, we investigated the effects of different V. destructor treatments on individual bees. Bees from hives that were not treated against Varroa destructor displayed reduced longevity and foraging performance. Both mite control treatments, i.e. vadescana and amitraz strips, were found to improve lifespan and health of bees compared to the untreated control group.
Vadescana-treated bees survived significantly longer than untreated bees and spent more time on tasks outside their hive, at the pre-foraging and foraging stage. They did more trips during their lifetime and had the shortest trips on average whereas untreated bees displayed opposite behaviour, i.e. fewer and longer trips. Longer time periods away from the hive, especially overnight, could be interpreted as a sign for loss of orientation and potential cognitive impairment of those bees57. As V. destructor infested bees have previously been shown to spend more time outside the hive, have longer flight times and lower return rates, the results of the untreated group could be explained by their higher mite levels57. Their shorter lifespan could also be attributed to these lower return rates, the negative effects of higher mite infestation levels and possibly virus infections vectored by mites19,58. Multiple studies have previously demonstrated that V. destructor parasitised bees have shorter lifespans and lower body weights59,60,61.
Foraging performance and lifespan of vadescana-treated bees are also likely connected. Out of-hive tasks such as orientation or foraging flights are known to be associated with various risks and a higher mortality rate than in-hive duties56,62. An earlier transition to foraging and a higher investment in foraging activities, might come at the cost of a shorter lifespan46,56. This negative correlation could explain why bees treated with dsRNA completed more flights over their lifetime than amitraz-treated bees, even though they had shorter average lifespans. Bees from both treatment groups showed normal foraging behaviour with trip numbers and lengths similar to results from other studies with a comparable RFID setup46,47.
Vadescana-treated hives had constantly lower mite loads indicating that treatment was efficient. Due to its mode of action, vadescana appears to not be lethal to mites and therefore is not expected to increase mite fall, instead it reduces reproduction of the mites and as a result stops population growth and therefore reduces the number of mites per colony31. Amitraz treatment did not change mite fall significantly compared to the untreated hives. Although bottom board counts are less accurate than other mite count methods, weekly counts over an extended period of time can provide insight into the development of mite numbers and are helpful to compare colonies within an experiment23,42. Weekly mite counts showed a similar trend for all treatment groups which is most likely due to seasonal changes in bee and mite populations in autumn63,64. Small colony sizes, rising mite numbers and re-invasion from surrounding hives can lead to a quick buildup in mite numbers11,65,66,67. At the end of the trial in May, all our hives had mite numbers considered to be at or above treatment threshold and economically damaging17,18,42.
Movement of bees between hives can contribute to the spread of mites and growing infestation rates63,66. Here, about 8% of tagged bees moved into neighbouring hives. We found two distinctly different patterns. Drifting bees moved into a non-natal hive permanently and generally did so within the same apiary site and at younger ages, which perhaps suggests they got lost during orientation flights49,50. In contrast, robbing bees visited hives in the other apiary as well as within their own apiary, but they returned to their home hives afterwards and their trips showed no clear age-related pattern. Robbing activity is initiated on a hive level by robbing bees recruiting other foragers after they have found a suitable target hive and robbing hives are characterised by shifts in foraging and guarding behaviour68,69. During robbing events foraging activity, aggression and thus mortality risk increase rapidly69. These changes were also evident here in activity and lifespan data as observed robbing events corresponded with heightened activity and mortality in the robbing hive (Supp. Mat. 5 and 6). Beyond the direct risks of fights with victim hive bees and potentially fatal injuries, robbing events can have long-term consequences for hive health and survival. Mites and other pathogens such as AFB or DWV can be transmitted horizontally to other hives via visiting bees63,70,71,72. Re-invasion of previously treated hives by mites from surrounding colonies can render earlier treatment efforts inefficient63,66. Therefore, movements of bees between hives can have a considerable impact on colony health and mite levels.
Amitraz and vadescana-treated bees had average lifespans of 28 and 25 days, respectively, which lies within the usually cited normal lifespan for a honey bee of 25–35 days73,74. Literature specifically on lifespans of RFID tagged bees tends to report slightly shorter lifespans of around 20 days, even for control groups, most likely because handling and tagging is an additional stressor that cannot be avoided entirely46,47,75. Consequently, even 21 days for the untreated hives seem to be within a natural range.
We did not find any negative impact of amitraz on bees, in contrast to other studies reporting effects of miticides such as flumethrin or amitraz on bee health and behaviour75,76,77. To observe these effects, pesticides or miticides have often been experimentally administered to individual bees directly either orally or through cuticle contact, but they might not influence bee performance in the field as colony-level data suggests76,77,78.
The vadescana-treated honey bees exhibited significantly earlier and increased foraging activity compared to amitraz-treated honey bees. As discussed previously, out of-hive foraging and even orientation flights are known to be associated with a higher mortality rate than in-hive duties62[,56. An earlier transition to foraging might come at the cost of a shorter lifespan56[,46. This foraging behaviour may have contributed to the slightly shorter lifespan of vadescana-treated honey bees compared to amitraz-treated honey bees.
RNAi-based treatments represent a new group of biopesticides that utilize a different mode of action compared to traditional miticides and therefore, can be used to manage populations that have developed resistance to other miticides and can be an important tool to prevent or delay resistance development in populations which do not have resistance to other miticides79. Integrated resistance management suggests the alternating use of multiple modes of action can delay or prevent the development of resistance and rotating between synthetic miticides and RNAi-based miticides could benefit beekeepers that struggle with resistant mite populations in their colonies80. dsRNA treatments have been used successfully to control V. destructor populations on a small scale26,31. Yet, data on their effects in the field and over longer time periods is still limited81,82,83. Here, we applied treatments to full-strength honey bee colonies (8-10 frames of bees) according to manufacturer’s instructions and thus bees and mites were exposed to concentrations comparable to field conditions. Therefore, these findings resemble what beekeepers can expect in terms of efficacy and impact on foraging performance in the field.
Furthermore, the impact of RNAi-technology on the mite’s host species and other insects present in the mite’s environment needs to be considered. Exposure of monarch butterflies (Danaus plexippus) and waxmoths (Galleria mellonella) to vadescana found no adverse effects on these species84,85. Previous studies also reported no effects of vadescana on honey bee pupae31. In line with these previous results, we did not observe negative impacts on adults in their foraging or lifespan, and on the contrary, vadescana-treated bees lived longer than bees from untreated hives and had earlier and increased foraging behavior compared to untreated or amitraz-treated hives.
Data availability
All data supporting the findings of this study are available within the paper and its Supplementary Information.
References
-
Chapman, N. C. et al. The final frontier: Ecological and evolutionary dynamics of a global parasite invasion. Biol. Let. 19, 20220589. https://doi.org/10.1098/rsbl.2022.0589 (2023).
-
Martin, S. J. et al. Global honey bee viral landscape altered by a parasitic mite. Science 336, 1304–1306. https://doi.org/10.1126/science.1220941 (2012).
-
Boncristiani, H. et al. World honey bee health: The global distribution of western honey bee ( Apis mellifera l.) pests and pathogens. Bee World 98, 1–5. https://doi.org/10.1080/0005772X.2020.1800330 (2020).
-
Soroker, V., Kovačić, M. & Hatjina, F. The coloss varroa task force: Combating the mite in the 21st century. Bee World 99, 14–16. https://doi.org/10.1080/0005772X.2021.1981678 (2022).
-
Traynor, K. S. et al. Varroa destructor: A complex parasite, crippling honey bees worldwide. Trends Parasitol. 36, 592–606. https://doi.org/10.1016/j.pt.2020.04.004 (2020).
-
Boecking, O. & Genersch, E. Varroosis – the ongoing crisis in bee keeping. J. Verbr. Lebensm. 3, 221–228. https://doi.org/10.1007/s00003-008-0331-y (2008).
-
Bąk, B., Wilde, J. & Siuda, M. The condition of honey bee colonies (Apis mellifera) treated for Varroa destructor by different methods. J. Apic. Res. 57, 674–681. https://doi.org/10.1080/00218839.2018.1495440 (2018).
-
Roth, M. A., Wilson, J. M., Tignor, K. R. & Gross, A. D. Biology and management of Varroa destructor (Mesostigmata: Varroidae) in Apis mellifera (Hymenoptera: Apidae) colonies. J. Integr. Pest Management https://doi.org/10.1093/jipm/pmz036 (2020).
-
Landcare Research New Zealand, M. W. Nz colony loss survey summary. (Manaaki Whenua, Landcare Research New Zealand, 2023). 2023
-
McGruddy, R. A. et al. Are increasing honey bee colony losses attributed to Varroa destructor in new zealand driven by miticide resistance?. J. Apicultural Res. https://doi.org/10.1080/00218839.2024.2364146 (2024).
-
Jack, C., Oliveira, I., Kimmel, C. & Ellis, J. Seasonal differences in Varroa destructor population growth in western honey bee (Apis mellifera) colonies. Front. Ecol. Evol. 11, 1102457. https://doi.org/10.3389/fevo.2023.1102457 (2023).
-
Rosenkranz, P., Aumeier, P. & Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 103(Suppl 1), S96-119. https://doi.org/10.1016/j.jip.2009.07.016 (2010).
-
Han, B. et al. Life-history stage determines the diet of ectoparasitic mites on their honey bee hosts. Nat. Commun. 15, 725. https://doi.org/10.1038/s41467-024-44915-x (2024).
-
Elias-Neto, M., Soares, M. P. M. & Bitondi, M. M. G. Changes in integument structure during the imaginal molt of the honey bee. Apidologie 40, 29–39. https://doi.org/10.1051/apido:2008064 (2008).
-
Martin, S. A population model for the ectoparasitic mite Varroa jacobsoni in honey bee (Apis mellifera) colonies. Ecol. Model. 109, 267–281. https://doi.org/10.1016/S0304-3800(98)00059-3 (1998).
-
Jack, C. J., Boncristiani, H., Prouty, C., Schmehl, D. R. & Ellis, J. D. J. Insect Sci. https://doi.org/10.1093/jisesa/ieae011 (2024).
-
Strange, J. P. & Sheppard, W. S. Optimum timing of miticide applications for control of Varroa destructor (Acari: Varroidae) in Apis mellifera (Hymenoptera: Apidae) in washington state, USA. J. Econ. Entomol. 94, 1324–1331. https://doi.org/10.1603/0022-0493-94.6.1324 (2001).
-
Jack, C. J. & Ellis, J. D. Integrated pest management control of Varroa destructor (Acari: Varroidae) the most damaging pest of (Apis mellifera l. (Hymenoptera: Apidae)) colonies. J. Insect Sci. https://doi.org/10.1093/jisesa/ieab058 (2021).
-
Noël, A., Le Conte, Y. & Mondet, F. Varroa destructor: How does it harm Apis mellifera honey bees and what can be done about it?. Emerging Topics in Life Sciences 4, 45–57. https://doi.org/10.1042/ETLS20190125 (2020).
-
Benito-Murcia, M. et al. Residual tau-fluvalinate in honey bee colonies is coupled with evidence for selection for Varroa destructor resistance to pyrethroids. Insects https://doi.org/10.3390/insects12080731 (2021).
-
Hernández-Rodríguez, C. S. et al. Large-scale monitoring of resistance to coumaphos, amitraz, and pyrethroids in Varroa destructor. Insects https://doi.org/10.3390/insects12010027 (2021).
-
Tihelka, E. Effects of synthetic and organic acaricides on honey bee health: A review. Slovenian Veterinary Res. https://doi.org/10.26873/SVR-422-2017 (2018).
-
Liu, F., Xu, X., Zhang, Y., Zhao, H. & Huang, Z. Y. A meta-analysis shows that screen bottom boards can significantly reduce Varroa destructor population. Insects https://doi.org/10.3390/insects11090624 (2020).
-
Smeele, Z. E., Baty, J. W. & Lester, P. J. Effects of deformed wing virus-targeting dsRNA on viral loads in bees parasitised and non-parasitised by Varroa destructor. Viruses 15, 2259. https://doi.org/10.3390/v15112259 (2023).
-
Maori, E. et al. IAPV, a bee-affecting virus associated with colony collapse disorder can be silenced by dsRNA ingestion. Insect Mol. Biol. 18, 55–60. https://doi.org/10.1111/j.1365-2583.2009.00847.x (2009).
-
Garbian, Y., Maori, E., Kalev, H., Shafir, S. & Sela, I. Bidirectional transfer of RNAi between honey bee and Varroa destructor: Varroa gene silencing reduces varroa population. PLoS Pathog. 8, e1003035. https://doi.org/10.1371/journal.ppat.1003035 (2012).
-
Brutscher, L. M. & Flenniken, M. L. RNAi and antiviral defense in the honey bee. J. Immunol. Res. 2015, 941897. https://doi.org/10.1155/2015/941897 (2015).
-
Wilson, R. C. & Doudna, J. A. Molecular mechanisms of RNA interference. Annu. Rev. Biophys. 42, 217–239. https://doi.org/10.1146/annurev-biophys-083012-130404 (2013).
-
Leonard, S. P. et al. Genetic engineering of bee gut microbiome bacteria with a toolkit for modular assembly of broad-host-range plasmids. ACS Synth. Biol. 7, 1279–1290. https://doi.org/10.1021/acssynbio.7b00399 (2018).
-
Cedeño, A. G., Nakaya, P., Niz, J., Sciocco-Cap, A. & Salvador, R. Double-stranded RNA synthesized in bacteria can be transferred to bee and varroa tissues. J. Apic. Res. 54, 99–100. https://doi.org/10.1080/00218839.2015.1103541 (2016).
-
McGruddy, R. A. et al. RNA interference as a next-generation control method for suppressing Varroa destructor reproduction in honey bee (Apis mellifera) hives. Pest Manag. Sci. 80, 4770–4778. https://doi.org/10.1002/ps.8193 (2024).
-
Greatti, M., Milani, N. & Nazzi, F. Reinfestation of an acaricide-treated apiary by Varroa jacobsoni oud. Exp. Appl. Acarol. 16, 279–286. https://doi.org/10.1007/BF01218569 (1992).
-
Gashout, H. A., Goodwin, P. H. & Guzman-Novoa, E. Lethality of synthetic and natural acaricides to worker honey bees (Apis mellifera) and their impact on the expression of health and detoxification-related genes. Environ. Sci. Pollut. Res. 25, 34730–34739. https://doi.org/10.1007/s11356-018-3205-6 (2018).
-
Dahlgren, L., Johnson, R. M., Siegfried, B. D. & Ellis, M. D. Comparative toxicity of acaricides to honey bee (Hymenoptera: Apidae) workers and queens. J. Econ. Entomol. 105, 1895–1902. https://doi.org/10.1603/ec12175 (2012).
-
Ai, H. & Takahashi, S. The lifelog monitoring system for honeybees: RFID and camera recordings in an observation hive. Journal of Robotics and Mechatronics 33, 457–465 (2021).
-
Schneider, C. W., Tautz, J., Grünewald, B. & Fuchs, S. RFID tracking of sublethal effects of two neonicotinoid insecticides on the foraging behavior of Apis mellifera. PLoS ONE 7, e30023. https://doi.org/10.1371/journal.pone.0030023 (2012).
-
Shi, J. et al. Sublethal acetamiprid doses negatively affect the lifespans and foraging behaviors of honey bee (Apis mellifera l.) workers. Sci. Total Environ. 738, 139924 (2020).
-
Colin, T., Meikle, W. G., Wu, X. & Barron, A. B. Traces of a neonicotinoid induce precocious foraging and reduce foraging performance in honey bees. Environ. Sci. Technol. 53, 8252–8261. https://doi.org/10.1021/acs.est.9b02452 (2019).
-
Inberg, A., Mahak, K. & Evans, J. Compositions and methods for controlling arthropod parasite and pest infestations. United States patent (2017).
-
Clapham, D. E. Calcium signaling. Cell 131, 1047–1058. https://doi.org/10.1016/j.cell.2007.11.028 (2007).
-
Streit, S., Bock, F., Pirk, C. W. W. & Tautz, J. Automatic life-long monitoring of individual insect behaviour now possible. Zoology 106, 169–171. https://doi.org/10.1078/0944-2006-00113 (2003).
-
Dietemann, V. et al. Standard methods for varroa research. J. Apic. Res. 52, 1–54. https://doi.org/10.3896/IBRA.1.52.1.09 (2013).
-
Posada-Flórez, F. J., Abban, S. K., Smith, I. B. Jr. & Cook, S. C. Development and evaluation of a new effective tool and method for assessing Varroa destructor (Acari: Varroidae) mite populations in honey bee colonies. Insects https://doi.org/10.3390/insects13050457 (2022).
-
A package for survival analysis in s (2023).
-
Survminer: Drawing survival curves using ‘ggplot2’ (2021).
-
Prado, A. et al. Honeybee lifespan: The critical role of pre-foraging stage. Royal Soc. Open Sci. 7, 200998. https://doi.org/10.1098/rsos.200998 (2020).
-
Colin, T., Warren, R. J., Quarrell, S. R., Allen, G. R. & Barron, A. B. Evaluating the foraging performance of individual honey bees in different environments with automated field RFID systems. Ecosphere 13, e4088. https://doi.org/10.1002/ecs2.4088 (2022).
-
Pgirmess: Spatial analysis and data mining for field ecologists (2024).
-
Pfeiffer, K. J. & Crailsheim, K. Drifting of honeybees. Insectes Soc. 45, 151–167. https://doi.org/10.1007/s000400050076 (1998).
-
Free, J. B. The drifting of honey-bees. J. Agric. Sci. 51, 294–306. https://doi.org/10.1017/S0021859600035103 (1958).
-
Willingham, R., Klopchin, J. & Ellis, J. D. Robbing behavior in honey bees. Edis 2015, 3–3. https://doi.org/10.32473/edis-in1064-2015 (2015).
-
Csárdi, G. et al. The igraph software package for complex network research. Inter. J. https://doi.org/10.32614/CRAN.package.igraph (2006).
-
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. https://doi.org/10.18637/jss.v067.i01 (2015).
-
Emmeans: Estimated marginal means (2018).
-
Robinson, G. E. Regulation of division of labor in insect societies. Annu. Rev. Entomol. 37, 637–665. https://doi.org/10.1146/annurev.en.37.010192.003225 (1992).
-
Rueppell, O., Bachelier, C., Fondrk, M. K. & Page, R. E. Jr. Regulation of life history determines lifespan of worker honey bees Apis mellifera l. Exp. Gerontol. 42, 1020–1032. https://doi.org/10.1016/j.exger.2007.06.002 (2007).
-
Kralj, J. & Fuchs, S. Parasitic Varroa destructor mites influence flight duration and homing ability of infested Apis mellifera foragers. Apidologie 37, 577–587. https://doi.org/10.1051/apido:2006040 (2006).
-
Yang, X. & Cox-Foster, D. L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. 102, 7470–7475. https://doi.org/10.1073/pnas.0501860102 (2005).
-
Annoscia, D., Del Piccolo, F. & Nazzi, F. How does the mite Varroa destructor kill the honeybee Apis mellifera? Alteration of cuticular hydrcarbons and water loss in infested honeybees. J. Insect Physiol. 58, 1548–1555. https://doi.org/10.1016/j.jinsphys.2012.09.008 (2012).
-
Morfin, N., Goodwin, P. H. & Guzman-Novoa, E. Varroa destructor and its impacts on honey bee biology. Front. Bee Sci https://doi.org/10.3389/frbee.2023.1272937 (2023).
-
Yang, X. & Cox-Foster, D. Effects of parasitization by Varroa destructor on survivorship and physiological traits of Apis mellifera in correlation with viral incidence and microbial challenge. Parasitology 134, 405–412. https://doi.org/10.1017/S0031182006000710 (2007).
-
Visscher, P. K. & Dukas, R. Survivorship of foraging honey bees. Insectes Soc. 44, 1–5. https://doi.org/10.1007/s000400050017 (1997).
-
Frey, E. & Rosenkranz, P. Autumn invasion rates of Varroa destructor (Mesostigmata: Varroidae) into honey bee (Hymenoptera: Apidae) colonies and the resulting increase in mite populations. J. Econ. Entomol. 107, 508–515. https://doi.org/10.1603/ec13381 (2014).
-
Leza, M. M., Miranda-Chueca, M. A. & Purse, B. V. Patterns in Varroa destructor depend on bee host abundance, availability of natural resources, and climate in mediterranean apiaries. Ecol. Entomol. 41, 542–553. https://doi.org/10.1111/een.12327 (2016).
-
Genersch, E. et al. The german bee monitoring project: A long term study to understand periodically high winter losses of honey bee colonies. Apidologie 41, 332–352. https://doi.org/10.1051/apido/2010014 (2010).
-
Kulhanek, K., Garavito, A. & vanEngelsdorp, D. Accelerated Varroa destructor population growth in honey bee (Apis mellifera) colonies is associated with visitation from non-natal bees. Sci. Rep. 11, 7092. https://doi.org/10.1038/s41598-021-86558-8 (2021).
-
Giacobino, A. et al. Short communication: Varroa destructor re-invasion dynamics during autumn and winter in Apis mellifera colonies from a temperate climate. J. Invertebr. Pathol. 197, 107890. https://doi.org/10.1016/j.jip.2023.107890 (2023).
-
Napier, T. C., Westwick, R. R., Kane, C. W. & Rittschof, C. C. Evaluating the cues that coordinate a shift towards the robbing foraging tactic in the honey bee (Apis mellifera). Behav. Ecol. Sociobiol. 77, 46. https://doi.org/10.1007/s00265-023-03321-x (2023).
-
Grume, G. J., Biedenbender, S. P. & Rittschof, C. C. Honey robbing causes coordinated changes in foraging and nest defence in the honey bee Apis mellifera. Ani. Behav. 173, 53–65. https://doi.org/10.1016/j.anbehav.2020.12.019 (2021).
-
Peck, D. T. & Seeley, T. D. Mite bombs or robber lures? The roles of drifting and robbing in Varroa destructor transmission from collapsing honey bee colonies to their neighbors. PLoS ONE 14, e0218392. https://doi.org/10.1371/journal.pone.0218392 (2019).
-
Schittny, D., Yañez, O. & Neumann, P. Honey bee virus transmission via hive products. Veterinary Sci. 7, 96. https://doi.org/10.3390/vetsci7030096 (2020).
-
Milbrath, M. Honey bee bacterial diseases Honey bee med. veterinary practitioner 277–293 (2021).
-
Amdam, G. V. & Omholt, S. W. The regulatory anatomy of honeybee lifespan. J. Theor. Biol. 216, 209–228. https://doi.org/10.1006/jtbi.2002.2545 (2002).
-
Knoll, S., Pinna, W., Varcasia, A., Scala, A. & Cappai, M. G. The honey (Apis mellifera l., 1758) and the seasonal adaptation of productions. Highlights on summer to winter transition and back to summer metabolic activity. Rev Livest. Sci. 235, 104011. https://doi.org/10.1016/j.livsci.2020.104011 (2020).
-
Wu, X. et al. The adverse impact on lifespan, immunity, and forage behavior of worker bees (Apis mellifera linnaeus 1758) after exposure to flumethrin. Sci. Total Environ. 858, 160146. https://doi.org/10.1016/j.scitotenv.2022.160146 (2023).
-
Begna, T. & Jung, C. Effects of sequential exposures of sub-lethal doses of amitraz and thiacloprid on learning and memory of honey bee foragers, Apis mellifera. J. Asia-Pacific Entomol. 24, 77–83. https://doi.org/10.1016/j.aspen.2021.03.012 (2021).
-
O’Neal, S. T., Brewster, C. C., Bloomquist, J. R. & Anderson, T. D. Amitraz and its metabolite modulate honey bee cardiac function and tolerance to viral infection. J. Invertebr. Pathol. 149, 119–126. https://doi.org/10.1016/j.jip.2017.08.005 (2017).
-
Colin, T. et al. Effects of late miticide treatments on foraging and colony productivity of european honey bees (Apis mellifera). Apidologie 52, 474–492. https://doi.org/10.1007/s13592-020-00837-3 (2021).
-
Lester, P. J. Integrated resistance management for acaricide use on Varroa destructor. Front. Bee Sc. https://doi.org/10.3389/frbee.2023.1297326 (2023).
-
Helps, J. C., Paveley, N. D., White, S. & van den Bosch, F. Determinants of optimal insecticide resistance management strategies. J. Theor. Biol. 503, 110383. https://doi.org/10.1016/j.jtbi.2020.110383 (2020).
-
Bortolin, F. et al. First evidence of the effectiveness of a field application of RNAi technology in reducing infestation of the mite Varroa destructor in the western honey bee (Apis mellifera). Parasit. Vectors 18, 28. https://doi.org/10.1186/s13071-025-06673-7 (2025).
-
Masucci, J. D. Developing double-stranded RNA as a new varroa control product. Am. Bee J. 160, 1–6 (2020).
-
McGruddy, R. A. et al. RNAi can be effective for the control of Varroa destructor in honey bee colonies. J. Apic. Res. In press. https://doi.org/10.1080/00218839.2026.2622814 (2026).
-
Krishnan, N., Hall, M. J., Hellmich, R. L., Coats, J. R. & Bradbury, S. P. Evaluating toxicity of varroa mite (Varroa destructor)-active dsRNA to monarch butterfly (Danaus plexippus) larvae. PLoS ONE 16, e0251884. https://doi.org/10.1371/journal.pone.0251884 (2021).
-
Bulgarella, M. et al. In silico analysis of potential off-target effects of a next-generation dsRNA acaricide for varroa mites (Varroa destructor) and lack of effect on a bee-associated arthropod. Insects 16, 317. https://doi.org/10.3390/insects16030317 (2025).
Acknowledgements
We thank Dave Wiessing from Dave’s bees, Ashurst, for the hives and his professional expertise.
Funding
Funding for this study was provided by the Faculty of Science Postgraduate Research Support at Victoria University of Wellington (grant number: 400095) and GreenLight Biosciences Inc.
Ethics declarations
Competing interests
Brian Manley is employed by GreenLight Biosciences Inc. All other authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Merk, J., Anastasi, M., McGruddy, R. et al. Longevity and foraging performance of honey bees treated with an RNAi-based Varroa destructor biopesticide. Sci Rep 16, 8208 (2026). https://doi.org/10.1038/s41598-026-38557-w
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-026-38557-w