
Introduction
The years 2030 and 2050 are set to be critical milestones for the future of the energy sector and the energy transition. The global necessity for a transition towards sustainable and renewable energy sources has significantly advanced research and development in the field of renewable energy technologies. Among these technologies, the field of gas fermentation utilizing archaea has rapidly emerged as a promising candidate to make significant contributions to achieving global energy and sustainability goals1,2,3,4. The European Union set the goal to produce 35 billion cubic meters of biomethane per year by 2030 in its REPowerEU Plan. Biomethane should be produced sustainably from organic waste and residual streams, avoiding conflicts related to land use and food security5. These residual biological carbon streams are processed in anaerobic digestion plants to produce biogas, a mixture of approximately 60 vol.-% methane (CH4) and 40 vol.-% carbon dioxide (CO2), which is further upgraded to separate the two gases5. In subsequent biomethanation of CO2, originating from biomass, and renewably produced molecular hydrogen (H2) are converted into CH4 by hydrogenotrophic methanogenic archaea (methanogens) according to Eq. (1)6,7. Methanogens, as strict anaerobes, only grow in anoxic environments, including the gastrointestinal tract8 or potentially in extraterrestrial environments9. They utilize a variety of carbonaceous substrates for biomass and energy production, including methyl compounds, acetate, and C1 compounds, such as CO210. Autotrophic, hydrogenotrophic methanogens obtain their energy from the conversion of H2 and CO2 to CH411,12,13.
Additionally, they are capable of producing proteinogenic amino acids under specific conditions14,15. While biomethanation has not yet been scaled beyond technology demonstration, it is showing promising efficiencies2. A system network of biogas production from organic residues and subsequent biomethanation can therefore act as a CO2 sink. The key benefit of CH4 as a renewable energy carrier over H2 is its ability to utilize existing infrastructure for distribution and storage globally16,17.
$${text{CO}}_{2(g)} + 4{text{H}}_{2(g)} to {text{CH}}_{4(g)} + 2{text{H}}_{2} {text{O}}_{(l)} quad Delta G^{0} = – {135};{text{kJ}};{text{mol}}^{ – 1}$$
(1)
The anaerobic cultivation of methanogens at ambient pressure conditions has been successfully demonstrated, and recent studies have investigated various methanogens to gain scientific knowledge about their growth behavior and CH4 productivity as CH4-producing cell factories11. The cultivation of methanogens under elevated pressure conditions presents several theoretical and practical benefits, which are still underrepresented in existing research18. From a bioprocess engineering perspective, operating under pressurized conditions can enhance CH4 production by increasing gas solubility and facilitating mass transfer, thereby ensuring a more efficient substrate delivery. Furthermore, renewable H2 is often available under pressure from advanced electrolyzers19,20,21,22, negating the need for energy-intensive compression. Biogas upgrading to biomethane under pressure allows for direct integration into the natural gas grid, aligning with industrial infrastructure standards and potentially reducing energy consumption and equipment complexity. These considerations underscore the importance of investigating methanogens under pressurized conditions.
Previous studies have demonstrated the viability of pressurized cultivation of metabolically active methanogens9,11,23, supporting their potential for industrial-scale applications. However, to develop economically viable biomethanation processes that fulfill industry requirements, it is essential to systematically examine methanogens across a broad spectrum of operational conditions to define their physiological limits24. Characterizing these boundaries is critical for optimizing strains with respect to key bioprocess parameters such as high-efficiency gas transfer, nutrient availability, and stable growth media conditions (i.e., temperature, pH, redox potential, and/or by-products of metabolism)17. The traditional cultivation of methanogens is initially conducted on a small scale in closed batch experiments. These experiments facilitate the generation of knowledge regarding the various physiological parameters of the desired methanogenic organism, including CH4 productivity rates, growth behavior, and the optimization of existing growth media11.
Nevertheless, the utilization of these methodologies results in a constrained availability of gaseous substrates to the organism, necessitating the optimization of alternative strategies for scale-up25. It is therefore essential to develop a robust system for the rapid screening and testing of microbes if the ultimate goal is to create a system that can be used on an industrial scale. The growth media play an essential role in maintaining a highly productive organism26,27,28. In this context, we developed a reliable laboratory test stand for gas utilizing microbes and an advanced growth medium. This setup is designed to optimize biological factors like methane productivity and growth behavior while also taking engineering considerations into account. By addressing these parameters, we enhance the system’s robustness and scalability, highlighting the innovation and industrial significance of this work. The bioreactor system comprises four parallel, pressure-resistant reactors. It is designed to operate at a maximum overpressure of 17 bar and a maximum temperature of 135 °C. The subsequent experiments were conducted at pressures of 3 and 10 bar and a temperature of 65 °C to evaluate the organisms’ performance under conditions relevant to industrial applications.
Results and discussion
High-pressure simultaneous bioreactor system
Here, we introduce the development of an advanced second-generation bioreactor system (SBRS-II), engineered to overcome the principal limitations of the original SBRS introduced by Pappenreiter et al.23. Although the original system facilitated high-throughput cultivation under elevated pressure, it was limited by a small working volume of 80 mL, unidirectional shaking motion, and difficulties in ensuring reliable data collection and reproducibility. The SBRS-II addresses these issues by providing a substantially larger reactor volume of 500 mL and a pressure capacity up to 17 bar, coupled with optimized mechanical stirring to enhance mass transfer efficiency. Additionally, the new design features PTFE-lined reactors to ensure long-term corrosion resistance, particularly important for cultivating metal-corrosive organisms such as M. marburgensis. Complemented by improved operational stability and a robust data acquisition interface, these modifications establish the SBRS-II as a versatile, reliable platform suitable for advanced high-pressure gas fermentation research. The microbial system employed in this study has been previously reported to oxidize elemental iron (Fe0)29. Under standard conditions in anaerobic environments, the spontaneous oxidation of Fe0 by potential oxidants such as CO2, nitrate (NO3−), or sulfate (SO42−) is thermodynamically unfavorable. Instead, microbial catalysis is required, whereby microorganisms facilitate the oxidation of Fe0 to Fe2+ for the generation of H2. In the current growth media, sulfide is supplied as Na2S. Based on the dissociation equilibrium (Supplementary Fig. 1) at pH 7, most S2− ions exist as HS− and H2S. This enhances the corrosion potential of the microbial system. The microorganisms subsequently consume the generated H2 in this process, promoting electron transfer and sustaining the overall redox reaction29. In certain cases, direct electron transfer between the reactor surface and microorganisms has also been observed30. Electron transfer via intermediate H2 is a well-documented mechanism in sulfate-reducing microorganisms31,32. Moreover, lower pH conditions enhance the availability of protons (H⁺), further promoting iron corrosion. In anaerobic environments, hydrogenotrophic microorganisms compete for dissolved H2, leading to rapid uptake and maintaining low H2 concentrations, which thermodynamically favors continued iron oxidation. Thus, microbial activity both facilitates and accelerates iron corrosion under strictly anaerobic conditions30. In the SBRS-II design, we addressed this issue by lining the reactor’s inner wall with a PTFE sleeve to prevent corrosion, thereby increasing durability and extending the reactor’s lifespan. In addition to this mechanical upgrade, the PTFE liners also ensure the reactor remains inert under the reaction conditions, preventing the uncontrolled introduction of unwanted components during the experiment.
The control unit of the new SBRS-II, was custom-engineered to meet the specific requirements of high-pressure microbial cultivation. The nominal working volume of each bioreactor was increased to 500 mL. Given the sensitivity of gas fermentation processes to molecular oxygen (O2) contamination, particular attention was devoted to preventing O2 ingress during filling, sampling, and broth harvesting operations18,33. All manipulations were performed under strictly anaerobic conditions, following established protocols, to ensure system closure at all stages.
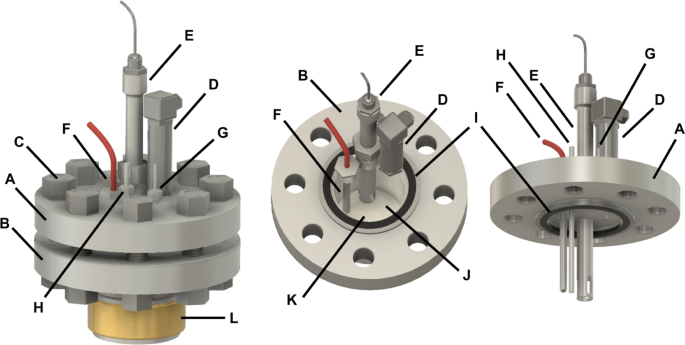
The new system employs mechanical stirring, implemented via magnetic stirrers, aiming to enhance the volumetric mass transfer coefficient (kLα)34. Once the culture broth is introduced, the operating temperature is reached, and the experimental protocol is uploaded to the control unit, the cultivations proceed autonomously. The new system eliminates the need for manual headspace exchange or re-pressurization, resulting in a significantly streamlined workflow and improved throughput for repetitive closed-batch experiments. Figure 1 provides a schematic 3D illustration of one of the SBRS-II reactors, including the arrangement of integrated sensors within the vessel and lid.

Schematic 3D illustration of the SBRS-II reactor system (own representation). Components include: (A) stainless steel reactor lid, (B) stainless steel reactor vessel with an inner diameter of 78 mm and inner height of 100 mm, (C) M24 bolts and nuts, (D) pressure sensor, (E) removable pH probe inside a protection tube, (F) PT100 temperature sensor, (G) gas inlet/outlet, (H) liquid inlet/outlet, (I) graphite gasket with a steel rim, (J) PTFE inlay, (K) PTFE stir bar, and (L) electrical heating band.
In the Supplementary Material to this article, an animation clip (Supplementary Video 1) of the reactor, a detailed P&ID diagram (Supplementary Fig. 2), a photograph (Supplementary Fig. 3), operational constraints, and laboratory safety measures of the developed SBRS-II system are provided.
Comparative assessment of chloride and sulfate-based growth media
In this study, M. marburgensis was cultivated under two distinct initial pressure conditions—3 bar and 10 bar—using the 4:1 H2/CO2 gas mixture in a repetitive closed batch cultivation mode. M. marburgensis is known from previous research as a baro-tolerant organism and has demonstrated the ability to grow and produce CH4 across a broad pressure range11,23,35,36,37. For the cultivation process, both the conventional minimal growth medium reported by Taubner et al.9 (further referred to as MM-CF medium), as well as a novel growth medium, in which sulfate salts are replacing chlorides as counterions for the active components (further referred to as MM-CF-S medium), were used. This new formulation was designed to minimize corrosion risks associated with steel-based reactors, which are typically highly promoted by chloride ions, as discussed earlier.
The conventional MM-CF medium utilizes chloride-based salts, including NiCl2 · 6 H2O (CAS 7791–20-0) in the TE 100× solution, which provides nickel ions as an essential trace element for the organism. However, under the European Registration, Evaluation, Authorisation and Restriction of CHemicals (REACH) regulation, set forth by the European Chemicals Agency (ECHA), NiCl2 · 6 H2O is classified as a toxic and endocrine-disrupting component38. To enhance safety during the preparation and handling of the medium and particularly concerning its large-scale usability without significant limitations, all chloride salts, except for NaCl, were substituted with sulfates in the new formulation.
The final composition of the MM-CF-S medium maintained active component concentrations similar to the MM-CF standard, resulting in 2.6 g L−1 (NH4)2SO4, 6.8 g L−1 KH2PO4, 1.0 g L−1 NaCl, and 1 vol.-% of a 100-fold concentrated sulfate-based trace element (TE-S 100×) solution.
A comparative assessment of M. marburgensis cultivated in the conventional chloride-based MM-CF medium and the novel sulfate-based MM-CF-S formulation using the SBRS-II reactor system revealed substantial differences in process performance. These findings highlight the significant impact of medium composition on the efficiency and robustness of bioprocesses.
As illustrated in Fig. 2, M. marburgensis cultivated in the chloride-based MM-CF medium exhibited increasing methane evolution rates (MER) with rising biomass concentrations, achieving its highest value during experiment set 8 at 10 bar. However, a progressive decline in the specific methane production rate ((q_{{{text{CH}}_{{4}} }})) was observed throughout the 10 bar experiments, indicating a shift in metabolic efficiency despite increased biomass. The corresponding values are presented in Table 1. The MM-CF medium reaction sets 2/3 and 5/6 were performed sequentially without intermediate sampling. Consequently, there is no quantitative data available for OD578 data for these reaction sets is available. ×-marked points representing linearly interpolated values between the respective reaction sets are incorporated into the graph (Fig. 2) and serve to illustrate the overall biomass growth trend over time. For sets 7–9, experiments using the MM-CF medium were carried out in two of the four reactors. Although this provided a smaller sample size (n = 2), the data exhibited consistent trends aligned with expected behavior, and the standard deviation was not notably high. We therefore consider that the reduced number of replicates is unlikely to compromise the reliability of the findings. Our experiments show that M. marburgensis grown on our new sulfate-based MM-CF-S medium delivered increasing MER at increasing biomass concentrations, as also shown in Fig. 3. However, (q_{{{text{CH}}_{{4}} }}) decreased as the number of experiments progressed, both in the 3 bar and 10 bar range, with an overall peak at set 5 at 10 bar. The respective values are given in Table 2.

In 2A, the characteristics of CH4 production at varying pressures using the MM-CF chloride medium. Reaction sets 1–6 correspond to cultivations at 3 bar (n = 4), and sets 7–9 correspond to cultivations at 10 bar (n = 2). For each reaction set, the methane evolution rate (MER) and specific methane production rate ((q_{{{text{CH}}_{{4}} }})) are illustrated as the average of the maximum MER values across all reactors. Colored diamonds represent individual reactor values. Error bars indicate the standard deviation of the mean values, showing variability in CH4 production across replicates. In 2B, biomass growth characteristics at different pressures using the MM-CF chloride medium are shown. Reaction sets 1–4 represent biomass cultivation at 3 bar (n = 4), while reaction sets 5–7 are cultivated at 10 bar (n = 2). Optical density (OD578) values are shown as mean values across all reactors. Colored rectangular markers represent OD578 measurements before each reaction cycle set, and triangular markers represent OD578 measurements after each cycle set, capturing biomass growth over time. × -markers between reaction set 2/3 and 5/6 indicate linear interpolated values. No measured data is available, as reaction sets were performed sequentially without sampling. Colored diamonds indicate individual OD578 values for each reactor, contributing to the mean. Hatched rectangles illustrate the net change in OD578, indicating biomass growth across cycles. Error bars represent standard deviations, showing variability across replicates.

In 3A, the characteristics of CH4 production at varying pressures using the MM-CF-S sulfate medium are shown. Reaction sets 1–4 correspond to cultivations at 3 bar, and sets 5–7 correspond to cultivations at 10 bar. For each reaction set, the methane evolution rate (MER) and specific methane production rate ((q_{{{text{CH}}_{{4}} }})) are illustrated as the average of the maximum MER values across all four reactors. Colored circles represent individual reactor values. Error bars indicate the standard deviation of the mean values, showing variability in CH4 production across replicates. In 3B, biomass growth characteristics at different pressures using the MM-CF-S sulfate medium are presented. Reaction sets 1–4 represent biomass cultivation at 3 bar, while reaction sets 5–7 are cultivated at 10 bar. Optical density (OD578) values are shown as mean values across all four reactors. Colored rectangular markers represent OD578 measurements prior to each reaction cycle set, and triangular markers represent OD578 measurements after each cycle set, capturing biomass growth over time. Colored circles indicate individual OD578 values for each reactor, contributing to the mean. Hatched rectangles illustrate the net change in OD578, indicating biomass growth across cycles. Error bars represent standard deviations, showing variability across replicates.
Across all experimental conditions, high gas conversion efficiencies were observed, with CH4 yields exceeding 95 vol.-% CH4 on average, and consistent biomass growth over all reaction sets.
The sulfate-based MM-CF-S medium achieved a significant improvement in CH4 productivity, with a MERmax of 285.86 ± 22.94 mmol L−1 h−1, nearly double that observed in the chloride-based medium (144.01 ± 9.18 mmol L−1 h−1). Similarly, the maximum biomass accumulation reached an optical density at 578 nm (OD578) of 6.79 ± 0.21 compared to 3.49 ± 0.03 in the chloride-based medium. The (q_{{{text{CH}}_{{4}} }})-rates also improved in the sulfate-based medium, reaching 216.85 ± 17.54 mmol g−1 h−1, compared to 180.36 ± 34.79 mmol g−1 h−1 in the chloride-based medium. The increase in (q_{{{text{CH}}_{{4}} }}) was less pronounced on a relative scale than that of MER or OD578. However, it still reflects a notable enhancement in metabolic efficiency. The determined (q_{{{text{CH}}_{{4}} }}) utilizing the chloride-based growth media can be seen as a baseline standard for the herein reported SBRS-II system and is to some extent comparable to previously reported values by Hofmann et al.35 at low-pressure closed batch cultivations using a gas-pressure controller (GPC) or Abdel Azim et al.27 during low-pressure fed-batch cultivation of M. marburgensis in a bioreactor.
The determined (q_{{{text{CH}}_{{4}} }}) utilizing the sulfate-based MM-CF-S medium, therefore, can be considered about 25% above the reported values determined using the GPC setup at low-pressure conditions35.
The determined conversion rates kmin for both employed growth media could be enhanced compared to values determined using the GPC by a factor of 2 MM-CF and a factor of around 4.25 for the MM-CF-S experiments at 10 bar (8.72 ± 0.56 bar h−1 for MM-CF and 16.93 ± 1.36 bar h−1 for MM-CF-S)35.
Despite the overall improvements, both media displayed signs of phase limitations. In the sulfate-based medium at 3 bar experiments, a plateau in MER despite increasing biomass indicated a gas-phase limitation, likely due to insufficient dissolved H2 (which is equal to a much slower dissolution kinetics compared to the metabolization kinetics) at higher cell densities (Supplementary Fig. 4A–D). This limitation was not apparent under 3 bar conditions in the chloride-based system (Supplementary Fig. 5A–D), as the biomass concentration in those experiments was just about half compared to the sulfate system. This implies a lower total gas–liquid mass transfer at the MM-CF experiments influenced by the different media components, and is confirmed by comparing the maximum kmin values for experimental series. At 10 bar, both media exhibited liquid-phase limitations, characterized by an early peak in MER followed by a decline, indicating that nutrient depletion was a limiting factor during extended cultivation cycles. Product inhibition appears rather unlikely in this case, as the phenomenon occurs in successive reaction cycles, between which product gas is removed. Nevertheless, the potential impact of other metabolic by-products cannot be entirely disregarded.
The exchange of chloride for sulphate ions in the media also changes the total ionic strength of the nutrient medium. It is well established that the total ionic strength exerts an influence on (micro)biological systems39,40,41,42. Consequently, it is plausible that this leads to a (slightly) enhanced bioavailability of nutrients, thereby yielding higher MER and (q_{{{text{CH}}_{{4}} }}) outcomes. Although the total ionic strength has not been specifically studied regarding the influence of the lipid composition of M. marburgensis, it is reasonable to suppose that the membrane lipid composition and/or their head groups could be altered as a result of different ionic strength, as it has already been shown for the intact polar lipid composition of Methanothermobacter thermautotrophicus43 and for the core lipid inventory of M. marburgensis14,44 when different nutrient availabilities or different cultivation conditions had been employed, respectively.
Microscopic imagery of samples taken after the reaction cycle sets showed the formation of organism clusters, as shown in Fig. 4. A comparison of microscopic images for each reaction set can be found in Supplementary Fig. 6. It is postulated that the formation of these clusters is a consequence of an increase in biomass concentration within the culture broth, where it is evident that the stirring employed in the SBRS-II is unable to separate these aggregates sufficiently. To achieve this, it would be necessary to utilize higher stirring speeds.

Microscopic image of M. marburgensis. The picture was captured after the MM-CF-S medium experiments reaction cycle set 4, at 40× magnification, and an OD578 of approximately 3.3. Cell clusters are highlighted with arrows, illustrating cellular aggregation.
The use of sulfate salts introduces an additional sulfur species that may pose risks in mixed microbial communities. In naturally occurring consortia of methanogens and sulfate-reducing bacteria, a tightly coupled metabolic interaction is often observed45,46. Sulfate-reducing organisms consume H2 to reduce sulfate to sulfide, simultaneously oxidizing CH4 to bicarbonate (HCO3−)47. Due to this metabolic competition, H2 is removed from the system as a substrate for the methanogens, resulting in inadequate CH4 production efficiency.
Scheller et al.47 demonstrated that under such conditions, the reverse reaction of methanogenesis can occur via methyl-coenzyme M reductase I (MCR-I), even resulting in net CH4 oxidation under certain conditions. In our study, however, the pure culture of M. marburgensis consistently exhibited unidirectional methanogenesis with no detectable reverse activity, which could have been observed as a significantly prolonged reaction time in the MM-CF-S experiments (see Supplementary Data). This confirms that, under controlled pure-culture conditions, the thermodynamic driving force for CH4 production remains sufficiently high so that the reverse reaction cannot be observed. Nevertheless, for the development of industrial-scale biomethanation processes, it is imperative to maintain a high level of culture hygiene to prevent the establishment of sulfate-reducing populations within the bioreactor, as their presence would compromise H2 availability and overall process efficiency.
Additionally, the pH behavior of the raw MM-CF-S system under varying pressure and temperature conditions was investigated, analogously to what was reported earlier for the MM-CF medium35. Compared to the previous data, here also, increasing pressure led to a predictable decrease in pH, driven by enhanced CO2 solubility. However, for the novel medium, the temperature dependency was far less pronounced than for the chloride system, indicating an enhanced buffer capacity. For the chloride-based MM-CF medium, the pH dropped below the lower physiological threshold for M. marburgensis of around pH 5, at pressures exceeding 12 bar35. Notably, this critical pH threshold was not breached in the sulfate-based MM-CF-S medium, indicating improved pH stability under high-pressure conditions, with a minimum at around pH 6 at 15 bar. The corresponding data is shown in Supplementary Fig. 7.
Conclusions and further perspectives
This study on the cultivation of M. marburgensis represents a significant advancement in our efforts to gain a deeper understanding of the strain’s behavior under elevated pressure conditions. The newly designed high-pressure SBRS-II test setup, described in this report, allows for a comprehensive physiological and biotechnological investigation of M. marburgensis. The results of the cultivation experiments with sulfate-based growth media demonstrate high efficiency in converting the H2/CO2 substrate gas into CH4. Our findings highlight a substantial improvement in gas transfer efficiency achieved with the SBRS-II system compared to its predecessor, enabling more industrial-scale related testing of biomethanation in the lab. The improved performance observed with the sulfate-based medium in pure M. marburgensis cultures further highlights the potential for even further medium optimization, especially for scaling to a high-performance, industrially relevant, and competitive bioprocess.
Methanogenic archaea have the potential to play a significant role in the process of “greening the gas grid”. Once established, biomethanation provides a relatively pure biomethane product that can be transported, distributed, and utilized in a manner comparable to fossil-based natural gas. It is therefore crucial to develop a biomethanation process that is well-characterized and thoroughly understood. It is essential to ensure an optimized substrate supply to the organism to maintain high efficiency rates. As we gain more knowledge, this will become more achievable. The industrial implementation of M. marburgensis in high-pressure biomethanation will depend on progress in key research areas. A major challenge is preventing microbial cross-contamination, as culture instability can significantly reduce efficiency. Future research should develop robust operational frameworks supported by advanced monitoring and rapid detection technologies for reliable large-scale performance. Additionally, exploring how pressure affects cellular metabolism is vital. Understanding pressure’s impact on enzyme activity and genetic evolution will help optimize process conditions and may uncover new biological mechanisms with broader biotech applications. It is also important to evaluate technological aspects. Rigorous benchmarking of yet-to-be-developed reactor designs with traditional systems, considering efficiency, energy use, and economic factors, will identify the best options for industry adoption. Long-term stability remains a key concern, requiring extended trials to detect potential process drift, plan maintenance, and ensure continuous methane production. Overcoming these challenges will advance high-pressure biomethanation technology and establish it as a key component in sustainable energy solutions.
Methods
Archaea strain and chemicals
Methanothermobacter marburgensis DSM 2133 was obtained from the Deutsche Sammlung für Mikroorganismen und Zellkulturen (DSMZ) GmbH (Braunschweig, Germany). Pre-cultures were prepared and maintained as previously reported37,48. Actively growing cultures were stored at 4 °C under a H2/CO2 atmosphere (20 vol.-% CO2 in H2) until experimental use.
All bottled gases, including CO2, H2, H2/CO2 mixture (20 vol.-% CO2 in H2), Biogon® C20 (20 vol.-% CO2 in N2), and N2, were of a minimum purity of 4.5 (Linde Gas, Stadl-Paura, Austria, and Air Liquide, Schwechat, Austria). Low-pressure nitrogen (up to 5 bar, solely for purging purposes) was sourced from the in-house supply and generated through the evaporation of liquefied nitrogen (Linde Gas, Stadl-Paura, Austria). All chemicals utilized for the preparation of the growth media were of standard analytical grade and procured from commercial suppliers without further purification. To ensure a sterile environment, all surfaces and equipment were cleaned using a biocide mixture of 38.6 wt.-% 2-Propanol and 18.2 wt.-% 1-Propanol in water.
Growth media
The growth medium used for cultivating pre- and working cultures was derived from the original medium presented by Schönheit et al.49,50, as well as a simplified medium by Taubner et al.9, referred to as the carbonate-free (MM-CF) medium. The final concentrations of the individual components in the culture broth are 2.1 g L−1 NH4Cl, 6.8 g L−1 KH2PO4, 1.0 g L−1 NaCl, 1 vol.-% of a 100-fold concentrated trace element (TE 100×) solution, and subsequently filled to the desired working volume with UV-sterilized 18 MΩ H2O (TKA Wasseraufbereitungssysteme GmbH, Germany) with the pH adjusted to 7.0 with NaOH. The MM-CF medium is subsequently anaerobicized by repetitive gassing with H2/CO2 gas and vacuuming, followed by an autoclaving step at 121 °C for 15 min. Once cooled to room temperature, 0.25 g L−1 sterile Na2S · 9 H2O is added anaerobically, after which the inoculation step is performed. The TE 100× solution was prepared according to the methodology outlined by Taubner et al.48, except for the replacement of ddH2O with UV-sterilized 18 MΩ H2O, and a final volume of 1000 mL.
To investigate the growth dynamics and CH4 productivity of M. marburgensis, this study employed a sulfate-based medium instead of the chloride-based medium used previously. For simplicity, this medium is further referred to as the sulfate (MM-CF-S) medium. The actual concentrations of the active components were maintained at similar levels to those used in previous studies, resulting in 2.6 g L−1 (NH4)2SO4, 6.8 g L−1 KH2PO4, 1.0 g L−1 NaCl, and 1 vol.-% of a 100-fold concentrated sulfate-based trace element (TE-S 100×) solution. MM-CF and MM-CF-S media were prepared as tenfold concentrates. Detailed recipes for the base media as well as the trace element solutions are available in the Supplementary material (Supplementary Tables 1–5) to this article.
High-pressure cultivation of M. marburgensis
For high-pressure experiments, working cultures were prepared by centrifugation (4000 rcf, 30 min, 20 °C) of the required amount of actively growing pre-culture and resuspending the biomass in the respective media to achieve an OD578 of approximately 0.5 at a final cultivation volume of 175 mL (250 mL Schott bottles, DURAN® pressure plus+ GL 45; DWK Life Sciences GmbH, Wertheim, Germany; butyl rubber seal and GL 45 caps, Ochs Laborbedarf, Bovenden, Germany). Anaerobic conditions were established by repeatedly gassing the bottles with a H2/CO2 gas mixture (maximum 1.5 bar), followed by subsequent vacuuming. All subsequent work with the medium or culture was conducted within a glove box under an N2 atmosphere. For each experiment, the cultivation temperature was set to 65 °C and 3 bar or 10 bar, respectively, with an agitation speed of 150 rpm. Each experiment set consisted of five repetitive closed batch reaction cycles. The endpoint for 3 bar experiments was set to 1.5 bar to prevent air input to the system while exchanging the reactor’s atmosphere prior to the follow-up reaction cycle. To obtain a representative sample after a test pattern, it was necessary to maintain a residual overpressure of approximately 500 mbar. The endpoint for the 10 bar experiments was set to 2.5 bar, respectively. Following the conclusion of an experiment set, the medium was replaced with a fresh supply. Therefore, the biomass was separated by centrifugation (4000 rcf, 30 min, 20 °C) and resuspended in fresh medium.
The here newly reported simultaneous bioreactor system, Gen. 2 (SBRS-II), comprises four identical 316L stainless steel vessels (R-001–R-004, Fitok Group, Germany) lined on the inside with a PTFE inlay with an approximate thickness of 1 mm and insulated on the outside with polyurethane foam. A schematic drawing of the reactors is depicted in Fig. 1, and a P&ID diagram and a photograph of the SBRS-II are shown in Supplementary Figs. 2 and 3. The reactors are mounted on an aluminum frame structure. Each vessel has a nominal volume of 500 mL. A stir bar, set in motion by a magnetic stirrer (IKA-Werke GmbH & Co. KG, Germany), is placed inside the apparatus to ensure proper mixing and enhance gas–liquid mass transfer. The magnetic stirrer was modified by replacing the original magnet with two cylindrical neodymium magnets (12 × 60 mm, nickel plated, grade N38). The individual bioreactors are externally heated by electrical heating bands (TIC10–TIC40, Guangzhou Lingke trade Company Ltd., China), which are setpoint controlled by an external control unit (otom® Group GmbH, Germany). PT100 temperature sensors (TI1–TI4, otom® Group GmbH, Germany), encapsulated in a pressure-resistant stainless steel immersion sleeve, are placed in each reactor to allow for constant monitoring of the actual reaction temperature. The pressure is being measured by four individual pressure sensors (PI1–PI4, Anhui Ruiling Meter Manufacturing Co., Ltd., China) installed in the reactor vessel lid. SBRS-II has supply lines for two single gases or gas mixtures, an off-gassing line, and a vacuum line. The supply lines are connected to the CO2/H2 mixture as well as the Biogon® C20 gas. Each reactor can be equipped with a special pressure-resistant pH probe (QI-pH, Endress + Hauser, Germany, maximum operating overpressure 17 bar) to monitor pH levels during high-pressure cultivation.
Each reactor is equipped with a riser tube for filling and emptying the reactor, as well as for collecting liquid samples. Additionally, the gas line is linked to the riser tube, enabling the manual introduction of gas beneath the fluid surface. All tubing, joints, and valves are constructed from stainless steel 316L. The SBRS-II system is equipped with a custom-manufactured control unit (PICR1–PICR4, Creonia e.U., Perg, Austria). The controller is capable of executing a predefined program, thereby controlling two sets of four solenoid valves (indicated as *-CV-*, Ningbo Brando Hardware Co., Ltd., China). The valves are commanded by the control unit to either open or close, depending on the pressure within the reactor or a predefined timeframe. During an ongoing experiment, the pressure of the respective reactor is recorded, stored in a data file on an SD card, and displayed on the controller display. The data recording interval can be set at the user’s discretion for each reactor. Additionally, the actual temperature inside the reactor is displayed by the SBRS-II’s control unit. Each of the four reactors can be operated fully individually.
A detailed Standard Operating Procedure (SOP) can be found in the Supplementary Material to this article, including the respective programs (Supplementary Tables 6, 7).
Figure 5 depicts a schematic representation of a 10 bar experiment as carried out, showing phases of the exchange of the reactor’s atmosphere and phases of methanogenesis.

Schematic illustration of a five-cycle repetitive closed batch test pattern. Phase A represents the initial exchange of the reactor atmosphere. Phase B illustrates the subsequent pressure decrease resulting from the methanation reaction. Phase C shows the atmosphere exchange between reaction cycles, and Phase D marks the end of the test pattern. Dotted lines indicate sections excluded from data processing; only solid lines are considered in further calculations.
Sample preparation and analytical setup
Analytical monitoring of the methanation reaction was conducted by liquid sampling at a maximum of 2.5 bar absolute reactor pressure following the completion of an experimental test pattern. To obtain samples, cannulas were connected to the corresponding sampling ports, and the samples were transferred into sealed glass vials (vial: 10 mL, Schmidling Labor + Service GmbH, Switzerland; seal: 3.0 mm, Butyl/PTFE, La Pha Pack, Langerwehe, Germany, max. 2.5 bar).
To monitor biomass growth, the optical density (OD578) at 578 nm was determined against a diH2O blank (UV-3600i Plus, Shimadzu). Liquid culture samples were taken after inoculation and completion of a reaction set.
Following the completion of MM-CF-S using reaction sets 2–7, microscopic images (Olympus, Tokyo, Japan) were obtained to observe potential morphological alterations in the organism.
Data processing
Raw pressure data from SBRS-II were obtained in a comma-separated *.csv file and further processed using OriginPro 2024b (OriginLab Corp.) software. A sampling rate of 1 s was set at the SBRS-II control unit. Before calculating the physiological characteristics, the recorded data were manually trimmed to isolate the data associated with the pressure decrease resulting from the methanation reaction. Additionally, a smoothing factor was applied.
As a hydrogenotrophic methanogen, M. marburgensis forms CH4 and H2O, according to Eq. (1), the respective pressure decrease can be used to determine the organism’s gas productivity, while disregarding the carbon flux into biomass formation, as this fraction accounts for a negligible percentage (3–5%) of the total amount of carbon51,52. The methane evolution rate (MER), see Eq. (2), is defined by the number of millimoles CH4 (ΔnCH4) produced over time (∆t) in the liquid reaction volume (Vliq). Following the stoichiometry of the methanation reaction, Eq. (2) can be reformulated, considering the ideal gas law, to process the recorded pressure data, as given in Eq. (3), with the absolute pressure difference (Δp) within Δt, the headspace volume (Vg) and the respective reaction temperature (T)48. The resulting MER versus time curves for each reactor were used to determine the maximum MER (MERmax) within the reaction cycle test pattern (as depicted in Supplementary Figs. 4 and 5). For the calculation of (q_{{{text{CH}}_{{4}} }}) according to Eq. (4), a factor (f) of 0.3148 for the conversion of OD578 to biomass concentration was used.
$$MERleft[ {{text{mmol}};{text{L}}^{ – 1} ;{text{h}}^{ – 1} } right] = frac{{Delta n_{{{text{CH}}_{4} }} }}{{Delta t cdot V_{liq} }}$$
(2)
$$MERleft[ {{text{mmol}};{text{L}}^{ – 1} ;{text{h}}^{ – 1} } right] = frac{{Delta p cdot V_{g} }}{{Delta t cdot V_{liq} cdot 4 cdot R cdot T}}$$
(3)
$$q_{{{text{CH}}_{4} }} left[ {{text{mmol}};{text{g}}^{ – 1} ;{text{h}}^{ – 1} } right] = frac{MER}{{OD_{578} cdot f}}$$
(4)
Statistics and reproducibility
All information on statistics and reproducibility is provided in the respective section. For all graphs in the main article, the data is represented as mean values with standard deviations. For all graphs in the Supplementary Material, the data is presented as is after the described data processing. Experiments were performed as quadruplicates (n = 4). If this is not applicable, it is stated in the respective section, table, or figure description.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
-
Teixeira, L. V., Moutinho, L. F. & Romão-Dumaresq, A. S. Gas fermentation of C1 feedstocks: Commercialization status and future prospects. Biofuels Bioprod. Biorefin. 12, 1103–1117 (2018).
-
Pfeifer, K. et al. Archaea biotechnology. Biotechnol. Adv. 47, 107668 (2021).
-
Straub, C. T. et al. Biotechnology of extremely thermophilic archaea. FEMS Microbiol. Rev. 42, 543–578 (2018).
-
Klein, M. et al. Markerless mutagenesis enables isoleucine biosynthesis solely from threonine in Methanothermobacter marburgensis. Microbiol. Spectr. 13, e03068-24 (2025).
-
REPowerEU Plan. (The European Commission, Brussels, 2022).
-
Angelidaki, I. et al. Biogas upgrading and utilization: Current status and perspectives. Biotechnol. Adv. 36, 452–466 (2018).
-
Awe, O. W., Zhao, Y., Nzihou, A., Minh, D. P. & Lyczko, N. A review of biogas utilisation, purification and upgrading technologies. Waste Biomass Valoriz. 8, 267–283 (2017).
-
Borrel, G. et al. Methanomethylophilus alvi gen. nov., sp. nov., a novel hydrogenotrophic methyl-reducing methanogenic archaea of the order Methanomassiliicoccales isolated from the human gut and proposal of the novel family Methanomethylophilaceae fam. Nov.. Microorganisms 11, 2794 (2023).
-
Taubner, R.-S. et al. Biological methane production under putative Enceladus-like conditions. Nat. Commun. 9, 748 (2018).
-
Kurth, J. M., Op den Camp, H. J. M. & Welte, C. U. Several ways one goal—methanogenesis from unconventional substrates. Appl. Microbiol. Biotechnol. 104, 6839–6854 (2020).
-
Mauerhofer, L.-M. et al. Hyperthermophilic methanogenic archaea act as high-pressure CH4 cell factories. Commun. Biol. 4, 289 (2021).
-
Rittmann, S.K.-M.R., Seifert, A. H. & Bernacchi, S. Kinetics, multivariate statistical modelling, and physiology of CO2-based biological methane production. Appl. Energy 216, 751–760 (2018).
-
Rittmann, S.K.-M.R. A critical assessment of microbiological biogas to biomethane upgrading systems. In Biogas Science and Technology Vol. 151 (eds Gübitz, G. et al.) 117–135 (Springer, Cham, 2015).
-
Taubner, R.-S. et al. Lipidomics and comparative metabolite excretion analysis of methanogenic archaea reveal organism-specific adaptations to varying temperatures and substrate concentrations. mSystems 8, e01159-22 (2023).
-
Reischl, B. et al. Quantitative analysis of amino acid excretion by Methanothermobacter marburgensis under N2-fixing conditions. Sci. Rep. 15, 1–13 (2025).
-
Jensen, M. B., Poulsen, S., Jensen, B., Feilberg, A. & Kofoed, M. V. W. Selecting carrier material for efficient biomethanation of industrial biogas-CO2 in a trickle-bed reactor. J. CO2 Util. 51, 101611 (2021).
-
Thapa, A., Jo, H., Han, U. & Cho, S.-K. Ex-situ biomethanation for CO2 valorization: State of the art, recent advances, challenges, and future prospective. Biotechnol. Adv. 68, 108218 (2023).
-
Hanišáková, N., Vítězová, M. & Rittmann, S. K. M. R. The historical development of cultivation techniques for methanogens and other strict anaerobes and their application in modern microbiology. Microorganisms 10, 412 (2022).
-
Parra, D., Valverde, L., Pino, F. J. & Patel, M. K. A review on the role, cost and value of hydrogen energy systems for deep decarbonisation. Renew. Sustain. Energy Rev. 101, 279–294 (2019).
-
Hermesmann, M., Grübel, K., Scherotzki, L. & Müller, T. E. Promising pathways: The geographic and energetic potential of power-to-x technologies based on regeneratively obtained hydrogen. Renew. Sustain. Energy Rev. 138, 110644 (2021).
-
Schiebahn, S. et al. Power to gas: Technological overview, systems analysis and economic assessment for a case study in Germany. Int. J. Hydrogen Energy 40, 4285–4294 (2015).
-
Ghaib, K. & Ben-Fares, F.-Z. Power-to-methane: A state-of-the-art review. Renew. Sustain. Energy Rev. 81, 433–446 (2018).
-
Pappenreiter, P. A., Zwirtmayr, S., Mauerhofer, L., Rittmann, S. K. R. & Paulik, C. Development of a simultaneous bioreactor system for characterization of gas production kinetics of methanogenic archaea at high pressure. Eng. Life Sci. 19, 537–544 (2019).
-
Chatzis, A., Gkotsis, P. & Zouboulis, A. Biological methanation (BM): A state-of-the-art review on recent research advancements and practical implementation in full-scale BM units. Energy Convers. Manag. 314, 118733 (2024).
-
Rittmann, S., Seifert, A. & Herwig, C. Essential prerequisites for successful bioprocess development of biological CH4 production from CO2 and H2. Crit. Rev. Biotechnol. 35, 141–151 (2015).
-
Rusmanis, D., O’Shea, R., Wall, D. M. & Murphy, J. D. Biological hydrogen methanation systems—an overview of design and efficiency. Bioengineered 10, 604–634 (2019).
-
Abdel Azim, A. et al. The physiology of trace elements in biological methane production. Bioresour. Technol. 241, 775–786 (2017).
-
Hendriks, A. T. W. M., van Lier, J. B. & de Kreuk, M. K. Growth media in anaerobic fermentative processes: The underestimated potential of thermophilic fermentation and anaerobic digestion. Biotechnol. Adv. 36, 1–13 (2018).
-
Lorowitz, W. H., Nagle, D. P. & Tanner, R. S. Anaerobic oxidation of elemental metals coupled to methanogenesis by Methanobacterium thermoautotrophicum. Environ. Sci. Technol. 26, 1606–1610 (1992).
-
Xu, D., Gu, T. & Lovley, D. R. Microbially mediated metal corrosion. Nat. Rev. Microbiol. 21, 705–718 (2023).
-
Dinh, H. T. et al. Iron corrosion by novel anaerobic microorganisms. Nature 427, 829–832 (2004).
-
Belay, N. & Daniels, L. Elemental metals as electron sources for biological methane formation from CO2. Antonie van Leeuwenhoek 57, 1–7 (1990).
-
Mauerhofer, L.-M. et al. Methods for quantification of growth and productivity in anaerobic microbiology and biotechnology. Folia Microbiol. (Praha) 64, 321–360 (2019).
-
Takors, R. et al. Using gas mixtures of CO, CO2 and H2 as microbial substrates: the do’s and don’ts of successful technology transfer from laboratory to production scale. Microb. Biotechnol. 11, 606–625 (2018).
-
Hofmann, W. et al. Method for automated high performance closed batch cultivation of gas-utilizing methanogens. AMB Express 15, 1–15 (2025).
-
Seifert, A. H., Rittmann, S. & Herwig, C. Analysis of process related factors to increase volumetric productivity and quality of biomethane with Methanothermobacter marburgensis. Appl. Energy 132, 155–162 (2014).
-
Rittmann, S., Seifert, A. & Herwig, C. Quantitative analysis of media dilution rate effects on Methanothermobacter marburgensis grown in continuous culture on H2 and CO2. Biomass Bioenergy 36, 293–301 (2012).
-
Nickel dichloride. ECHA Dossier publication, REACH registration, article 10—full, lead (joint submission) https://chem.echa.europa.eu/100.028.858/dossier-list/reach/dossiers/active (27/08/2024).
-
Yin, Y. et al. Deeper insight into the effect of salinity on the relationship of enzymatic activity, microbial community and key metabolic pathway during the anaerobic digestion of high strength organic wastewater. Bioresour. Technol. 363, 127978 (2022).
-
Solon, K. et al. Effects of ionic strength and ion pairing on (plant-wide) modelling of anaerobic digestion. Water Res. 70, 235–245 (2015).
-
George, S. et al. Dynamic modelling the effects of ionic strength and ion complexation on trace metal speciation during anaerobic digestion. J. Environ. Manag. 343, 118144 (2023).
-
Rath, K. M., Maheshwari, A., Bengtson, P. & Rousk, J. Comparative toxicities of salts on microbial processes in soil. Appl. Environ. Microbiol. 82, 2012–2020 (2016).
-
Yoshinaga, M. Y. et al. Methanothermobacter thermautotrophicus modulates its membrane lipids in response to hydrogen and nutrient availability. Front. Microbiol. 6, 5 (2015).
-
Baumann, L. M. F. et al. Quantitative analysis of core lipid production in Methanothermobacter marburgensis at different scales. Bioengineering 9, 169 (2022).
-
Jespersen, M. & Wagner, T. Assimilatory sulfate reduction in the marine methanogen Methanothermococcus thermolithotrophicus. Nat. Microbiol. 8, 1227–1239 (2023).
-
Maslać, N., Sidhu, C., Teeling, H. & Wagner, T. Comparative transcriptomics sheds light on remodeling of gene expression during diazotrophy in the thermophilic methanogen Methanothermococcus thermolithotrophicus. mBio 13, e02443-22 (2022).
-
Scheller, S., Goenrich, M., Boecher, R., Thauer, R. K. & Jaun, B. The key nickel enzyme of methanogenesis catalyses the anaerobic oxidation of methane. Nature 465, 606–608 (2010).
-
Taubner, R.-S. & Rittmann, S.K.-M.R. Method for indirect quantification of CH4 production via H2O production using hydrogenotrophic methanogens. Front. Microbiol. 7, 532 (2016).
-
Schönheit, P., Moll, J. & Thauer, R. K. Nickel, cobalt, and molybdenum requirement for growth of Methanobacterium thermoautotrophicum. Arch. Microbiol. 123, 105–107 (1979).
-
Schönheit, P., Moll, J. & Thauer, R. K. Growth parameters (Ks, μmax, Ys) of Methanobacterium thermoautotrophicum. Arch. Microbiol. 127, 59–65 (1980).
-
de Poorter, L. M. I., Geerts, W. J. & Keltjens, J. T. Coupling of Methanothermobacter thermautotrophicus methane formation and growth in fed-batch and continuous cultures under different H2 gassing regimens. Appl. Environ. Microbiol. 73, 740–749 (2007).
-
Schill, N., van Gulik, W. M., Voisard, D. & von Stockar, U. Continuous cultures limited by a gaseous substrate: Development of a simple, unstructured mathematical model and experimental verification with Methanobacterium thermoautotrophicum. Biotechnol. Bioeng. 51, 645–658 (1996).
Acknowledgements
Markus Ungerank (Creonia e.U.) is acknowledged for his contributions during the design phase of SBRS-II and for providing the control unit. Karl Schütz is acknowledged for his support during the setup of the SBRS-II at the JKU high-pressure lab.
Funding
The Österreichische Forschungsförderungsgesellschaft (FFG) is greatly acknowledged for funding the project FlaeXMethan (Grant 888509). Johannes Kepler University Open Access Publishing Fund and the Federal State of Upper Austria support open access publication.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Orthofer, M., Hofmann, W., Rittmann, S.KM.R. et al. A high-pressure bioreactor system for the cultivation of Methanothermobacter marburgensis on advanced growth media for sustainable energy applications. Sci Rep 15, 35519 (2025). https://doi.org/10.1038/s41598-025-19544-z
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-19544-z