
Introduction
The succession and variation of the human microbiome profoundly affect human health and disease from cradle to grave. Correlation between mounting diseases and gut microbiome has been revealed nowadays, and probiotic interventions have consequently gained recognition as potential therapeutic modalities of cancers, autoimmune and metabolic diseases, etc. Despite centuries of clinical use1,2,3, probiotic therapy is extreme individualized variation in practical probiotic colonization, and beneficial effects are often limited to the administration period4,5. Evolutionary biology ascribes this vulnerable probiotic colonization to the poor adaptability of bacteria individuals. Specifically, bacterium colonization is a dynamic process including reversible attachment, irreversible adhesion, microcolony formation, and biofilm maturation6. However, practice indicates that stable biofilm formation is extremely difficult under the constant environmental assaults7,8. Recent studies revealed that the formation of biofilm is highly modulated in distinct ontogenetic phases in all aspects of gene, protein, and phenotype, analog to the Phylogeny mode of multicellular eukaryotic embryos9. Additionally, specific multicellular behaviors such as intracellular signaling, cell differentiation, and labor division among bacteria in the biofilm have long been discovered10,11. Evidence have indicated the critical role of multicellularity in biofilm formation, yet the practical application of this advanced perspective has never been explored. Therefore, we intend to redefine the underlying principle of probiotic colonization by integrating this multicellular perspective in developing innovative strategies for probiotic therapy.
By drawing an analogy between the growth dynamics of bacterial colonies and the embryonic development of multicellular eukaryotes, we have proposed an innovative concept of utilizing multicellular probiotic microcolonies as delivery units. Compared to probiotic individuals, probiotic microcolonies demonstrate superior colonization competitiveness due to two key advantages (1) Bacterial microcolonies rely on quorum sensing (QS) system to coordinate and display physiological and behavioral characteristics such as antagonizing foreign bacteria, resisting harsh microenvironment, and stimulating luminescence, whereas individuals cannot12,13; (2) Extracellular matrix, constituting three-dimensional (3D) structure of biofilm, not only promotes spatial aggregation but also enhances bacteria adaptability against antimicrobials and competing microorganisms as natural barrier14. While the multicellular microcolony strategy conceptually conforms to biofilm formation modality, but microcolony from traditional gas/solid interface cultivation is uncontrollable, uncollectable, and untransportable. Thus, strategy integrating tunable cultivation and delivery of microcolony is urgently needed.
This multicellular microcolony strategy requires both finely regulated nutrient supply and extracellular matrix accumulation to support microcolony cultivation, and 3D external structure to maintain colony integrity15,16,17. However, the protective armors designed for bacterial protection or functionalization in probiotic delivery studies18,19,20 cannot accommodate the volumetric expansion of the microcolony, flexibly manipulate microcolony growth, or maintain the structural integrity of microcolony. Some studies have demonstrated that hydrogel with the 3D network can encapsulate organisms, allow nutrient penetration, and even influence metabolism of the bacteria21,22,23, but the cross-linked network may also suppress bacteria growth and proliferation via spatial limitation and mechanical interaction, impeding the development of microcolonies. The internal stress of stress-relaxing hydrogel could gradually decrease over time under sustained external forces, allowing hydrogel to deform and provide space for cell growth24,25,26. Here, the viscoelastic hydrogels with combined physical and covalent crosslinks could adapt to microcolony growth via stress-relaxation property, and recent studies also indicated that tunable viscoelasticity of 3D hydrogels selectively affects the growth, metabolism, and biological behavior of the encapsulated organisms23,27,28. Therefore, we hypothesize that stress relaxation and mechanical stress of viscoelastic hydrogels help manipulate the fate of the probiotic colony in the 3D culture systems and construct a compact, dynamic, and metabolically active probiotic microcolony with precisely tunable spatial constraints.
In this work, inspired by the rapidity, efficiency, safety, and reliability of Express Mail Service (EMS), we co-opte the acronym EMS and designed an Express Microcolony Service (EMS) system with tunable matrix stiffness and viscoelasticity to support microcolony based on a multicellular self-organized strategy (Fig. 1a), which integrates the free-radical polymerization covalent crosslinking network with ion-crosslinking network of classical calcium alginate, exploring the biomechanical properties that affect key factors of colonization including microcolony self-organization, quorum sensing, stress response and adhesion matrix formation in 3D hydrogel network. Optimized culture time and colony density that provide maximized colonization benefits are studied on EMS micro-cargo, and the change of colonization-related gene expression and stress resistance of self-organized microcolony are analyzed via prokaryotic transcriptome sequencing. Moreover, the hybrid hydrogel recipe is optimized multidimensionally to achieve highest colonization efficiency and activity of microcolony in probiotic therapy, finally achieving sustainable health benefits.

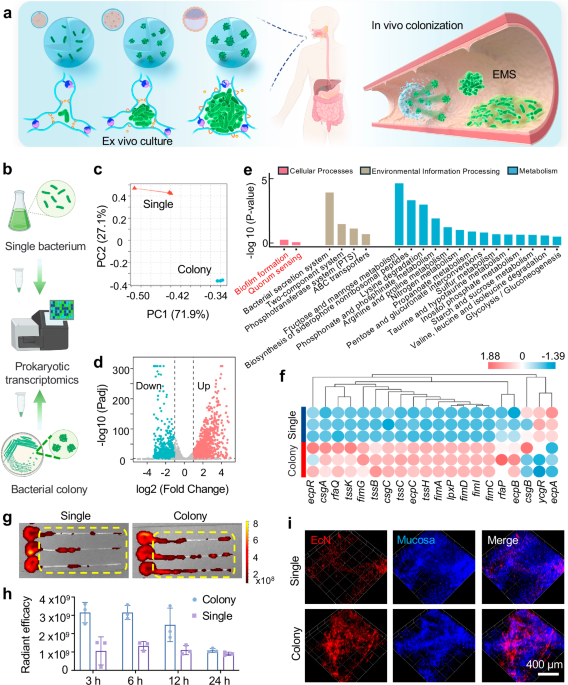
a Schematic illustration of the Express Microcolony Service (EMS) micro-cargo system which supports oral delivery of microcolony for biofilm formation of probiotics. Created in BioRender. Liu, H. (2025) https://BioRender.com/7lpn56z. b Schematic illustration of prokaryotic transcription sequencing. Created in BioRender. Liu, H. (2025) https://BioRender.com/h654r91. c The principal coordinate analysis between the colony group and the single bacterium group. d Volcano plot of the differential expressed genes (DEGs), analyzed by a two-tailed nbinomTest in DESeq. e Kyoto Encyclopedia of Gene and Genomes (KEGG) enrichment analysis, the colony group verse the single bacterium group. f Heatmap of biofilm formation-related gene expression. g Representative fluorescence image of colons (6 h after gavage). h IVIS radiant efficiency analysis displayed bacterial adhesion in the colon 3 h, 6 h, 12 h, 24 h after oral gavage. i Representative confocal image of intestinal mucosa with bacterial adhesion, scale bar 400 µm. Data in h were presented as mean ± SD (n = 3 biologically independent samples). Source data are provided as a Source Data file.
Results and discussion
Prokaryotic transcriptome sequencing of single bacterium and multicellular microcolony
The transition from a single bacterium to surface-attached bacterial colony involves coordinated regulatory cascades triggered by environmental, interbacterial or intracellular signals, and diverse alterations in genetic expression are essential for the formation of the intricate structure and distinct physiological characteristics of the biofilm29. In the study, we applied Escherichia coli Nissle 1917 (EcN), a traditional probiotic provides anti-inflammatory effect, protects the mucosal barrier, and inhibits intestinal pathogen invasion30,31,32. Firstly, prokaryotic transcriptomics was used to study the differential expressed genes (DEGs) between the single bacterium and multicellular colony from agar plate. As shown in Fig. 1b, we collected both bacteria suspension from LB broth media and bacterial colonies on Luria-Bertani (LB) agar medium for prokaryotic transcriptomic sequencing analysis. Principal component analysis (PCA) suggested that the transcriptome structure of bacteria differs between two culture conditions (Fig. 1c). The volcano plot showed the distribution of DEGs by different gene difference multiples and P-values (Fig. 1d). We found that nearly half of the gene expression changed between the single bacterium and multicellular colony from agar plate, there were 991 down-regulated genes and 1129 up-regulated genes in the multicellular colony compared with the single bacterium. Next, the Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis was performed (Fig. 1e), and we found that two important pathways including biofilm formation (tssC, tssB, tssK, et al.) and quorum sensing (ydcU, livH, secG, et al.) in terms of cell biological processes were upregulated in the multicellular colony rather than the single bacterium. Additionally, some amino acid metabolism and carbohydrate metabolism were upregulated in multicellular colony (Fig. 1e). This change may be attributed to the upregulation of quorum sensing, a mechanism of intercellular communication that enables microorganisms to modulate gene expression in response to cell density33,34. The enhancement of quorum sensing appears to stimulate metabolic pathways associated with biofilm formation, potentially including increased production of extracellular polysaccharides (EPS) for colony protection35. This could consequently alter the bacteria’s energy and carbon source metabolism.
mRNA expression of genes associated with biofilm formation is depicted in Fig. 1f. csg operons encoding adhesive organelles like curli fimbriae and fim-related genes (fimA, fimI, fimC, fimD, fimG) encoding type I flagella and contributing to the formation of antibiotic-resistant capsules, all of which are critical to the formation of biofilm were significantly upregulated in the multicellular colony compared with the single bacterium36. ecpC and ecpB encoding common fimbriae of Escherichia coli and ecpR encoding fimbriae synthesis transcription regulators37 also exhibited significant up-regulation. These findings demonstrated that EcN exerted superior ability in adhesion and colonization in the colony group compared to the single bacterium group. Additionally, bacterial stress resistance mechanisms critically facilitated its colonization and biofilm formation by enabling resistance against external challenges like heat, osmotic pressure, and acidic environments. The analysis presented in Supplementary Fig. 1 reveals differential expression patterns for stress-related genes all significantly up-regulated in the multicellular colony, including BhsA, pspA, pspC rpoE rseA rpoS, along with the heat shock proteins (htpG hspQ htpX ibpAB) and cold shock protein (cspA)37. Notably, higher mRNA expression of acid resistance-related genes38 involving glutamate decarboxylase (gadA, gadB, gadC), arginine decarboxylase (adiA), and acid-resistant chaperone proteins (hdeBA, hdeD) were all enhanced in multicellular colony group compared to the single bacterium group. The anti-acid property is essential for the effective treatment of probiotic therapy, because oral probiotics must withstand the strong bactericidal effect of gastric fluid before reaching the intestine. In summary, these findings suggest that bacteria in the colony-growing mode exhibit improved colonization potential than individuals in bacteria suspension.
For further verification, bacteria planktonic suspension and colony suspension were collected for oral gavage. The bacteria entered the colonic lumen about 2–3 h post-administration, so the bacterial adhesion was measured by fluorescence quantification at 3 h, 6 h, 12 h, and 24 h after oral gavage (Supplementary Fig. 2a). Images acquired from In Vivo Imaging System (IVIS) indicated that within 12 h, the adhesion rate of the colony group was much higher than that of the single bacterium group, whereas this difference became insignificant by 24 h (Fig. 1g, h), suggesting the failed bacterial colonization. This change may be attributed to (1) oral administration leading to the direct exposure of bacteria to the gastric acid, resulting in damaged bacterial viability; (2) it is difficult to control the morphology and size of colonies in 2D agar plates, and collecting the colonies from these agar plates could damage the colony structure, which hinders colonization. Also, bacteria suspension and colonies were injected separately into colon to validate the direct adhesion ability (Supplementary Fig. 2b). IVIS images showed that the adherence rate of the colony group was significantly higher than single bacterium group after 1 h of incubation (Supplementary Fig. 2c, d). Finally, confocal microscopy was applied for the 3D scanning on the surface of mucosa. Figure 1j showed that the adhesion pattern of single bacterium group to the intestinal mucosa was scattered, while the adhesion of multicellular colony group from the LB agar plate was relatively concentrated and had a relatively higher adhesion rate. In summary, we expect to utilize the 3D hydrogel to (1) protect bacterial viability against the gut environment, (2) integrate and simplify the process of cultivation-delivery of microcolonies, (3) flexibly manipulate size and morphology of microcolonies in the 3D hydrogel network.
Optimization of stress-relaxed hydrogel and preparation of EMS micro-cargo system
3D network hydrogel can encapsulate microbial organisms while allowing nutrient diffusion, and hydrogel with stress-relaxation property allows hydrogel deformation under the forces generated by bacterial proliferation, thereby providing space for growth of microcolonies24,25,26. Moreover, current studies are exploring rules by which the biophysical parameters of stress-relaxing hydrogels modulate fundamental biological processes including cellular morphological changes, proliferation, and stem cell differentiation39,40. Nevertheless, it is still unclear how the transcriptome and biological behavior of bacteria change within 3D hydrogel matrices. Here, we introduced the covalent cross-linking network (methylacrylylated sodium alginate hydrogel, AlgMA) into the classical dynamic cross-linking network of calcium alginate hydrogel to obtain a 3D-hydrogel culture system (Fig. 2a) with adjustable matrix stiffness and stress-relaxation property. A series of hydrogels with different Alginate to AlgMA ratios were designed and divided into five groups according to percentage of AlgMA: 0%, 25%, 50%, 75%, and 100%. The rheological study of these hydrogels revealed that the initial modulus was varied under different crosslinking conditions (Fig. 2b). The average storage moduli of group 0%, 25%, 50%, 75%, and 100% under only optical crosslinking showed an increasing trend which were 0.8, 4.8, 86.5, 201.2, and 469.1 Pa, respectively. The average storage moduli under only Ca2+ crosslinking showed a decreasing trend which were 5063.9, 3974.9, 2974.8, 1912.2, 1025.8 Pa, respectively. The average storage moduli with double cross-linking of each group were 4726.5, 4736.7, 3826.6, 2956.4, and 2040.3 Pa, respectively. This data implied that the dynamic crosslinking by calcium ions provides better mechanical properties for the hydrogel. In addition, stress relaxation, the key characteristic of this series of dual-network hydrogels mentioned above also showed gradient changes with the adjustment of components, which is the higher the proportion of covalence network, and the worse the stress relaxation characteristics (Fig. 2c, d). Moreover, half decay time (τ 1/2), the parameter that represents the time that reach half of max relaxation stress is 8.6 s, 39.1 s, 108.0 s, 290.9 s, and 388.1 s, respectively. These rheological parameters suggested that the lower the proportion of AlgMA in the dual-network hydrogel, the faster the stress relaxation rate of the hydrogel.

a Schematic illustration of the design of dual-network hydrogel. Created in BioRender. Liu, H. (2025) https://BioRender.com/7lpn56z. b Evaluation of initial storage modulus of hydrogels under different crosslinking conditions (ion crosslink, covalent crosslink and double crosslink). c Stress-relaxation profiles of hydrogels with different Alginate to AlgMA ratios. d Relaxation half-time (τ1/2) of the AlgMA hydrogels. e Relative change in storage modulus (G’) compared to the initial storage modulus (G’₀) of the hydrogel during swelling in LB medium, expressed as G’/G’₀. f Weight change of swelling hydrogel in LB medium. g Weight change of swelling hydrogel in AGF and ASF. h Weight change of swelling hydrogel in ACF. i Schematic illustration of the preparation of EMS. Created in BioRender. Liu, H. (2025) https://BioRender.com/7lpn56z. j Optical image of the hydrogel micro-cargo. Scale bar 800 µm. k The size distribution of the micro-cargo. l Representative images of microcolonies in micro-cargo after being cultured for 2 h (I), 4 h (II), 6 h (III), and 8 h (IV), scale bar 200 µm. m Representative SEM image of the micro-cargo system (MS), primary micro-cargo system (PMS, encapsulated dispersed single bacteria) and the EMS micro-cargo system (encapsulated bacterial microcolonies). Representative images for 2j, l, m are shown, each experiment was repeated at least three times independently with similar results. Data in (d, g, h) were presented as mean ± SD (n = 5 independent samples). Source data are provided as a Source Data file.
In addition, the synthesized hydrogel designed for probiotic culture and oral delivery has to maintain its stability throughout LB culture and gastrointestinal challenge until reaching the colon. Therefore, we conduct a series of swelling-degradation tests in LB medium and artificial digestive fluids. First, we measured the change of storage modulus of hydrogels (0%, 25%, 50%, 75%, and 100% AlgMA group) in LB medium (Fig. 2e), and found that 100%AlgMA group maintained the highest storage modulus after 12-h of swelling. Meanwhile, weight changes of hydrogels after 3 h of swelling reached 155.56%, 123.64%, 114.78%, 112.07%, and 107.66% of the initial mass, which showed a decreasing trend in turn (Fig. 2f). Among them, the weight of 0%AlgMA hydrogels decreased significantly after reaching the peak value at 3 h, indicating a degraded state afterward, while the degradation was not obvious in the other groups due to the existence of the covalent crosslinking network. Although the storage modulus all decreased significantly, all groups of hydrogels maintained a relatively stable state during swelling-degradation except for the 0%AlgMA group.
Then, we systematically evaluated the swelling-degradation of hydrogel micro-cargo from each group in different artificial digestive fluids in turn (Fig. 2g). Alginate is relatively stable in the acidic environment, so all five groups did not degrade in the artificial gastric fluid (AGF). However, 0%AlgMA hydrogel micro-cargo degraded rapidly in the artificial intestine fluid (ASF), indicating that 0%AlgMA was not suitable for this oral system. The other groups swelled to different degrees in the ASF due to their stable covalent network. The degradation in the artificial colon fluid (ACF) is shown in Fig. 2h, we found that the 50%AlgMA group degraded more rapidly than other groups. It is speculated that most of the cross-linking sites in 0% and 25% AlgMA groups are occupied by calcium ions, and the loss of calcium ions in ASF is less, so its degradation in ACF is slower than that of the 50%AlgMA group, while the degradation of the 75% and 100%AlgMA groups was slow due to their covalency network. Although ionically crosslinked networks provide beneficial stress relaxation for microcolony formation, stable covalent networks are also indispensable for effective oral delivery. Therefore, 50%AlgMA hydrogel was chosen for the following investigation, because it maintained stability in AGF and ASF but degraded the fastest in ACF according to the swelling-degradation evaluation.
In this study, micro-cargo for oral administration was prepared via gas-shearing microfluidic strategy which is efficient in production of homogeneous microspheres on a large scale41, and the probiotics were dispersed in the micro-cargo and cultured in the LB medium in appropriate condition (Fig. 2i). As shown in Fig. 2j, k, the micro-cargo was round and uniform in size, the average particle size was 434 ± 12.75 µm. It is critical to study the changes in biological behavior of bacteria in the stress-relaxed hydrogel. Here, micro-cargo produced from hydrogel containing 2 × 105 CFU/mL bacteria were cultured in the liquid medium at 37 °C. The micro-cargo as well as the growing microcolonies during culturing were observed under the microscope (Supplementary Fig. 3). The formation of self-organized microcolonies at the time points of 2, 4, 6, and 8 h was observed (Fig. 2l), and fluorescence demonstrated a temporal increase in the volume of green fluorescent-labeled microcolonies. The microcolonies grew into uniform microspheres rather than undergoing morphological instability and roughening. This phenomenon is primarily attributed to the hydrogel’s elastic stress that physically constrains rapid microcolony expansion, thereby enforcing spherical morphological confinement. Then stress-relaxation property of hydrogel facilitates gradual microcolony growth by diminishing elastic stress over time. In addition, the surface morphology of empty micro-cargo (MS, without bacteria), primary micro-cargo system (PMS, containing dispersed single bacteria without undergoing culturing), and EMS (containing bacterial microcolonies after culturing) were observed using a scanning electron microscope (Fig. 2m), we can observe the structure of matured microcolonies on the surface of the cultured micro-cargo.
Gene expression pattern of self-organized microcolonies in micro-cargo
Subsequently, the expression of colonization-related genes in microcolonies cultured within distinct dual-network hydrogels was investigated. It was found that the mRNA expression changes of quorum sensing gene (sdiA), adhesion gene (csgA), extracellular polysaccharide gene (adrA) and stress response gene (iraM) in microcolonies were not linearly correlated with the AlgMA percentage (hydrogels with 0%, 25%, 50%, 75% and 100% AlgMA) under the same initial bacteria concentration and same culture conditions (Fig. 3a–d). Although the changes between groups were not uniform, we found that sdiA, csgA, adrA, and iraM in the 50% AlgMA group exhibited higher expression compared with other groups. In addition, we measured the total number of bacteria in different hydrogels and the bacterial growth curve after the bacteria released from the hydrogel (Fig. 3e, f). OD 600 absorbance of bacteria from the five groups after 6-h culture were 0.3452, 0.3942, 0.4722, 0.3872, and 0.2772, respectively. The total bacteria in the high covalent hydrogel network (100%AlgMA group) were less than other groups, it was speculated that the poor viscoelasticity of the high covalent network limited the growth of microcolonies. Also, the colony growth curve indicated that the 50%AlgMA group had a faster proliferation rate (steeper proliferative slope) and a higher OD600 value (0.4722). Previous literature has reported that flexible design of matrix may control the function or composition of living elements inside, and growth restriction provided by hydrogel network in a certain range may promote the formation of compact, vigorous, and metabolically active bacterial microcolonies27, which is similar to our findings. Combined with the finding in colonization-related gene expression and the swelling-degradation tests, the 50%AlgMA hydrogel was not only more suitable for the culture-oral delivery system but also exhibiting greater proliferative capacity and enhanced survival rates compared to other formulations. Therefore, the 50% AlgMA hydrogel was chosen for the following investigation. Further study analyzing the difference of the colonization-related genes between the single bacterium and 50% EMS (contain multicellular microcolony in 50% AlgMA micro-cargo) was carried out (Fig. 3g). The mRNA expressions of multicellular microcolony (quorum sensing gene sdiA, global regulatory genes csrA and csgD, adhesion gene csgA, extracellular polysaccharide synthesis gene adrA, and stress response gene iraM) were all significantly higher than the single bacterium group. In summary, the multicellular microcolony from 50%AlgMA hydrogel micro-cargo has significantly improved colonization potential.

a mRNA level of quorum sensing-related gene sdiA of microcolony from different hydrogels. b mRNA level of adhesion-related gene csgA of microcolony from different hydrogels. c mRNA level of exopolysaccharide-related gene adrA of microcolony from different hydrogels. d mRNA level of stress response-related gene iraM of microcolony from different hydrogels.e Bacterial quantification in different hydrogel micro-cargoes. f Growth curve of bacteria after releasing from the hydrogel micro-cargoes. g mRNA level of colonization-related genes of microcolony from different hydrogels. h The principal coordinate analysis of bacterial transcriptome from 50% EMS group and the single bacterium group. i Volcano plot of DEGs of the two groups, analyzed by a two-tailed nbinomTest in DESeq. j Heat map of transcriptional differential genes of the two groups (important up-regulated genes of 50%EMS group were listed). k Up-regulated pathway of the 50% EMS group according to KEGG enrichment analysis. l Down-regulated pathway of the 50% EMS group according to KEGG enrichment analysis. For in k and l, the horizontal axis represents gene ratio, the vertical axis represents gene term, and the bubble size represents the enriched gene counts. The P-value was calculated by hypergeometric distribution method. Data in (a–d, g) were presented as mean ± SD (n = 3 independent samples). Source data are provided as a Source Data file.
To explore how the culturing-delivery system influences the colonization potential of probiotics from the biological perspective, we studied the transcriptional characteristics of 50% EMS and the single bacterium through prokaryotic cell transcription. The principal component analysis suggested that the transcriptional profile of probiotics under the two culture conditions differs a lot (Fig. 3h). Besides, Fig. 3i, j showed 226 up-regulated and 158 down-regulated genes in 50% EMS group compared with the single bacterium according to the volcano plot and heat map of DEGs, important upregulated genes (including genes related to biofilm formation, two-component systems, phosphotransferase system (PTS), and amino acid metabolism pathways) of the 50% EMS group marked on the right side of the heatmap. According to the KEGG enrichment analysis (Fig. 3k, l), biofilm formation pathways were enriched in 50% EMS group in the up-regulated pathway enrichment diagram. The two-component system that can sense a variety of environmental signals (such as osmotic pressure, temperature, pH value, small molecules, etc.) to manipulate bacterial activity42 was also upregulated in 50% EMS. Additionally, metabolism-related pathways such as phenylalanine metabolism, amino sugar and nucleotide sugar metabolism, D-amino acid metabolism, nitrogen metabolism, and sulfur metabolism were all up-regulated, which contributes to the promotion of bacteria viability. Moreover, the PTS pathway43, which facilitates the transport of sugars and their derivatives via a phosphoric acid cascade reaction, was also up-regulated in 50% EMS group. This pathway is integral to carbon and nitrogen metabolism, the regulation of intracellular metal ion homeostasis, and influences stress responses and other biological processes. Finally, the mRNA expression of quorum sensing related genes (trpE, fadD), biofilm formation related genes (csrA, tssC) were significantly up-regulated in 50% EMS group compared with the single bacterium group (Supplementary Fig. 4). In conclusion, the transcriptional differences between 50% EMS group and single bacterium group suggest that probiotics from multicellular microcolony perform better in colonization, further experiments will be conducted to clarify.
Additionally, we analyzed the transcriptomic difference between the 50%EMS and colony group (From Fig. 1) to distinguish whether hydrogel solution has an impact on colony growth. PCA analysis indicated a significant difference between the colony group and the 50%EMS group (Supplementary Fig. 5a), and 1044 up-regulated and 1057 down-regulated genes were found in 50% EMS group compared with the colony group (Supplementary Fig. 5b). Further KEGG analysis (Supplementary Fig. 5c, d) revealed that cell cycle was enriched in the 50%EMS group, suggesting the active bacterial proliferation. Additionally, many metabolic pathways concerning amino acid and glycolysis were upregulated in 50%EMS group compared with colony group, especially the energy metabolism pathway—oxidative phosphorylation was significantly upregulated. These data suggest that 50%EMS can promote bacterial energy metabolism and proliferation. Reasons for those change can be: (1) The stress relaxation characteristics of 50%EMS, which provide appropriate spatial constraint forces that promote the formation of highly proliferative and metabolically active microcolonies; (2) sodium alginate is a natural polysaccharide, which may be utilized by bacteria as a carbon source, leading to change in glycolysis metabolism. However, we did not find significant enrichment of quorum sensing and biofilm formation pathways in KEGG analysis, suggesting that the enhanced colonization effect result from the colony structure, and the hydrogel may promote bacterial metabolism and proliferation to some extent.
Self-organization and release of microcolony
The biofilm formation experiences five successive processes including reversible adhesion, irreversible adhesion, microcolony formation, maturation, and detached diffusion. Although the water channels in hydrogel can transport water and nutrients, the nutrient transport will be insufficient for all bacteria during microcolony self-organization, which affects the colonization potential of microcolonies. Here, we intend to explore the characteristics of microcolony self-organization in dual-network hydrogels. We studied the effects of initial bacterial concentration and culture time on the density and size of the microcolony in 50%EMS. Given that high bacterial density in hydrogel micro-cargo intensifies competition for nutrients and space, whereas low density results in excessive microcolony growth, and the concentration of 5 × 10⁶ CFU/mL already resulted in narrow bacterial spacing and slowed microcolony growth, we chose three bacterial concentrations with a 5-fold serial dilution (2 × 105, 106, and 5 × 106 colony-forming unit (CFU) /mL) to prepare micro-cargo. As shown in Fig. 4a and Supplementary Fig. 6, the maximum average diameter of microcolonies in the three groups of hydrogel micro-cargo (initial bacteria concentration of the 2 × 105, 106, and 5 × 106 CFU/mL) was 23.94 μm, 14.96 μm and 9.86 μm. It is indicated that the higher initial bacterial densities correlated with slower colony self-organized kinetics (observing the slope of the proliferation curve), and the smaller the microcolonies after reaching the plateau (Fig. 4b, c). Additionally, it was found that the bacterial proliferation reached the plateau period at different period in three groups with an initial bacterial concentration of 2 × 105, 106, and 5 × 106 CFU/mL (Fig. 4d), suggesting that the higher initial bacterial concentration in the micro-cargo, the faster bacterial proliferation, so that the bacteria quantity reach the plateau rapidly. However, total bacterial content in equal amounts of hydrogel micro-cargo was similar regardless of the initial bacterial concentration after culturing over 12 h.

a Representative image showing change of microcolony under different initial bacterial concentration (2 × 105, 106, or 5 × 106 CFU/mL) and culture time (0 h, 2 h, 4 h, 6 h, 8 h, 12 h and 24 h). b Size analysis of microcolony according to the diameter. c The maximum diameter of the microcolony in hydrogel with different initial bacterial concentration. d Change of total bacteria in the micro-cargoes. e Live/Dead staining of microcolonies in hydrogel micro-cargoes. f Representative images of swelling and degradation process of hydrogel micro-cargo in ACF and the release of microcolony. The significance between the two groups was calculated utilizing a two-tailed Student’s t-test. Data are presented as mean ± SD (n = 10 independent samples for b, c; n = 3 independent samples for d). Source data are provided as a Source Data file.
Meanwhile, the bacteria viability inside the micro-cargo during culture was studied via fluorescence cell viability assay (Fig. 4e and Supplementary Fig. 7). The dead-to-live ratio of bacteria at 0 h, 2 h, 6 h, and 12 h was 0.0%, 7.1%, 2.8%, and 27.3%, respectively, which suggested that dead bacteria increased over time after the proliferation in microcolonies reached saturation. Among them, the mortality ratio of 6 h is lower than that of 2 h may be caused by the active proliferation at 6 h that significantly increased the proportion of live bacteria. Finally, the release of microcolonies from the hydrogel micro-cargo was studied. As shown in Fig. 4f, alginate hydrogel gradually degraded in ACF, and the internal microcolonies were dispersed in the ACF in 30 min. In summary, this section explored influence of initial bacterial concentration and culture time on the formation and survival of microcolonies and elucidated the release characteristics of microcolonies.
Environmental resistance and colonization potential of self-organized microcolony
Key to the successful oral delivery of the probiotic is the stress resistance against the harsh gastrointestinal environment. Traditional probiotic delivery strategies provided protective armor for each bacterium via chemical/genetical engineering, which the protective layer of liposomes, metal-phenols, polydopamine, or surface group modifications shelter the bacteria from the gastric acid, bile salt, and antibiotics in the digestive tract. Here, we analyzed the stress-resistant property of single bacterium and multicellular microcolony in the micro-cargo. We also compared the microcolonies with an equivalent quantity of uncultured probiotics encapsulated in hydrogel micro-cargo to determine the protective effect was attributed to the hydrogel matrix or the microcolony structure (Fig. 5a). We treated the Single bacterium, EMS micro-cargo system, and primary micro-cargo system (PMS) with AGF, ASF (containing bile salt), and antibiotics. The spot test44 revealed that the hydrogel protects the bacteria from gastric acid, but not antibiotics or bile salts. However, the multicellular microcolony structure significantly enhanced resistance to bile salts and antibiotics compared to the other two groups (Fig. 5b–e). The growth curves of the single bacterium group and EMS group were comparable, indicating that the structure of the microcolony did not affect the proliferation of bacteria (Supplementary Fig. 8). Finally, we investigated the bacterial viability in PMS and EMS after antibiotic treatment (Fig. 5f, g). The bacterial mortality in EMS was significantly lower than that of PMS, which may be ascribed to the dense structure of the microcolony and the cumulative extracellular polysaccharides. In summary, we proved that microcolony structure conferred higher resistance against environmental assault including acidic fluid, bile salt, and antibiotics.

a Schematic illustration of microcolony resistance to environmental assaults. Created in BioRender. Liu, H. (2025) https://BioRender.com/7u2jmla. b Spot assay showing the bacterial resistance against AGF, bile acid, and antibiotic. c Quantitative analysis of surviving bacteria after AGF treatment. d Quantitative analysis of surviving bacteria after bile acid treatment. e Quantitative analysis of surviving bacteria after antibiotic treatment. f Live/Dead staining of PMS after antibiotic treatment. g Live/Dead staining of EMS after antibiotic treatment. h Quantitative analysis of bacteria adhered to intestinal epithelial cell. i Quantitative analysis of EPS. j Representative image of crystal violet staining of biofilm, analyzing the biofilm-forming potential of PMS group, EMS group, PMS-D (dispersed) group, EMS-D group (dispersed means cells were dispersed and extracellular matrix was washed off). k Quantitative analysis of biofilm formation by measuring the crystal violet absorbance at 590 nm. l Timeline of mouse colonization model (n = 5). Created in BioRender. Liu, H. (2025) https://BioRender.com/jxkryqz. m Quantitative analysis of fecal EcN abundance within 3 days after gavage. n Quantitative analysis of fecal EcN abundance in small intestine (SI), cecum, and colon 3 days after gavage. The significance between the two groups was calculated utilizing the two-tailed Student’s t-test or Mann–Whitney U-test. Data are presented as mean ± SD (n = 3 independent samples for c–e, h, n = 5 independent samples for I; n = 6 independent samples for k; n = 5 independent animal samples for m, n). Source data are provided as a Source Data file.
Next, we analyzed the actual colonization capacity of microcolony. The self-aggregation ability (Supplementary Fig. 9) and cell adhesion ability (Fig. 5h, Supplementary Fig. 10) of the single bacterium in PMS and microcolony in EMS were evaluated. The results indicated that the aggregation ability of bacteria in microcolony was slightly higher than that of the single bacterium, and its adhesion to intestinal epithelial cell Caco-2 was also significantly higher than that of the single bacterium. Bacterial-secreted extracellular polysaccharides constitute fundamental extracellular structural of the biofilm, critically mediating probiotic adhesion, mechanical stabilization, and biofilm protection45. Therefore, we utilized the phenol-sulfuric acid method to detect exopolysaccharides, and the results suggested its production in the multicellular microcolony group was twice more than that of the single bacterium group (Fig. 5i). Additionally, the biofilm thickness (Supplementary Fig. 11) and the quantity (Fig. 5j, k) were detected to analyze the biofilm formation ability. It was observed that microcolonies from EMS produce more biofilm matrix than EMS-dispersed group (matrix of microcolony was washed off) under the same culture conditions, implying the key role of microcolony structure during colonization. Previous studies used biofilm matrix to encapsulate probiotics for oral delivery and proved that it improved environmental tolerance and adhesion of probiotics in the digestive tract46. In our study, the microcolony from EMS intrinsically generates robust environmental resistance and adhesion properties, without requiring additional processing steps like matrix extraction or coating.
Then the in vivo colonization capacity of oral EMS or PMS was evaluated (Fig. 5l). To promote the colonization of probiotics, all mice were treated with kanamycin (50 μg/mL) dissolved in drinking water prior to probiotic administration. Then each mouse was given 20 mg EMS through oral gavage, while the control group was given 20 mg PMS containing matched bacterial counts as EMS. During the next 3 days, using spot assay to quantify the fecal EcN abundance. We found that fecal EcN abundance decreased gradually over time in the PMS group, while EMS group only increased slightly on the second day and had a similar abundance on the third day compared with the first day (Fig. 5m). Given fecal probiotic content cannot represent bacterial colonization in the gut, we dissected the small intestine, cecum, and colon from these mice for tissue bacterial quantitative analysis (Fig. 5n, Supplementary Fig. 12a). The results demonstrated that cecum EcN content was the highest, followed by colon, and almost no EcN colonized in the small intestine. The average EcN abundance of the EMS group was significantly higher than the PMS group, specifically 89-fold higher in cecum, 52-fold higher in colon, two-fold higher in small intestine. The colonization of EcN (expressing red fluorescent protein) can be visualized in the frozen tissue sections (Supplementary Fig. 12b).
Therapeutic efficacy of oral EMS
EcN has been widely used in the treatment of inflammatory bowel disease (IBD), and it is the only probiotic approved by European Crohn’s and Colitis Organization Guidelines for remission maintenance in IBD patients47. It was reported that EcN possesses comparable therapeutic effect to mesalamine30, relying on its anti-inflammatory and intestinal flora regulation properties48. Both literature and the clinical application suggest that EcN plays a significant role in inducing and maintaining remission of IBD, but usually limited to the administration period, suggesting that probiotics did not colonize. Here, mouse model with colitis induced by dextran sulfate sodium (DSS) (Fig. 6a) was constructed, we examined the therapeutic efficacy of EcN microcolony through oral administration of EMS. We prepared hydrogel micro-cargo (without culturing) directly loaded with EcN (the PMS group) as the control group to distinguish the effect from microcolony or individual probiotic. First-line drug 5-ASA was also used as the control of effective treatment. The EMS group significantly maintained the body weight and colon length and reduced the disease activity with only one dose of probiotic therapy (Fig. 6b–f, Supplementary Fig. 13). Compared with 5-ASA, the EMS group did not differ significantly in weight change and colon length but had a significantly lower disease activity score.

a Experimental design of DSS-induced colitis, all mice were randomly divided into 7 groups (n = 5 per group): healthy control, DSS group, DSS + 5-ASA group (positive control), DSS + EMS group (oral gavage of micro-cargo containing EcN microcolonies), DSS + PMS (oral gavage of micro-cargo containing EcN), DSS + EcN (oral gavage of EcN suspension), DSS + MS (oral gavage of blank micro-cargo). Created in BioRender. Liu, H. (2025) https://BioRender.com/jxkryqz. b Change of mice body weight at the end of modeling (day 9). c Daily changes of body weight of mice. d Analysis of disease activity index (DAI) score of each group. e Analysis of colon length of each group. f Macroscopic colon appearance. g Representative images of Hematoxylin and eosin (H&E) staining of colon tissue section. h Quantification of intestinal EcN abundance using qPCR method. i mRNA analysis of ZO-1 and Occludin of colon tissue. j mRNA analysis of IFN-γ, IL-1β, IL-6, TNF-alpha and MCP-1 of colon tissue. k Histogram of LDA value showing the difference of gut microbiota between the DSS and the EMS group. l Functional annotation clustering heat map showing the difference of gut microbiota between the DSS and the EMS group. m Schematic illustration of the colonization process of EcN from EMS and its beneficial effect. Created in BioRender. Liu, H. (2025) https://BioRender.com/7lpn56z. The significance between the two groups was calculated utilizing the two-tailed Student’s t-test. Data are presented as mean ± SD (n = 5 independent animal samples for b–e, h; n = 3 independent animal samples for i, j). Source data are provided as a Source Data file.
Hematoxylin and Eosin (H&E) staining of colon tissue (Fig. 6g) revealed that the integrity of colon epithelium was well maintained in the EMS group, and the immune cell infiltration was also significantly reduced. H&E staining of tissue section from major organs of the mice (liver, lung, heart, and kidney) also demonstrated that this micro-cargo system possesses good biocompatibility (Supplementary Fig. 14). Previous literature found that EcN could regulate epithelial tight junction protein that protects the intestinal epithelial barrier32,49. In addition, quantitative analysis of EcN in colon tissue was performed by quantitative polymerase chain reaction (qPCR) (Fig. 6h), and it was found that the tissue EcN abundance in EMS group was remarkably higher than the EcN group (oral gavage of EcN suspension), suggesting that the colonization effect of EMS microcolony in the intestine was higher than that of scattered bacteria. Finally, mRNA expression of cell junction protein and inflammatory factors in colon tissues was analyzed (Fig. 6i, j). It was found that mRNA expression of ZO-1 and Occludin in the EMS group was a little bit lower than that in healthy control group but is significantly higher than that in the DSS group. Meanwhile, the mRNA expression of cytokines (IFN-γ, IL-1β, IL-6 and MCP-1) in the colon of the EMS group was also considerably lower than those in the DSS group, which is consistent with the trend in disease activity score. In summary, a single dose of oral EMS can effectively achieve predicted colonization and therapeutic efficacy in the acute colitis model without the need for additional maintenance.
16S rRNA sequencing was conducted to analyze the fecal microbiota alteration in DSS group and EMS group. Initially, fecal samples collected one day before the administration of DSS were analyzed to make sure each mouse started with the same microbiota composition. Alpha (Supplementary Fig. 15a) and beta diversity (Supplementary Fig. 15b) between the DSS group and the EMS group showed no significant difference, indicating that the bacterial abundance and microbial diversity were similar. Further analysis suggested that the microbial composition at the phylum level (Supplementary Fig. 15c) and functional prediction analysis (Supplementary Fig. 15d) based on PICRUSt2 system displayed no significant difference. Then, fecal samples collected after DSS and EMS administration were analyzed, we found that 91 ASVs were common between the two groups (Supplementary Fig. 16a). Shannon and Simpson indexes (Supplementary Fig. 16b) showed no notable difference in alpha diversity between DSS group and EMS group, which might be ascribed to the short-term of acute colitis model. In addition, the 3D principal coordinates analysis (PCoA) plot indicated the microbiome between the DSS group and the EMS group significantly differed (Supplementary Fig. 16c). Further investigation was conducted for the detailed microbiota alteration, and the gut microbiota distribution at the phylum and genus levels is shown in Supplementary Fig. 17. Here, we identified key bacteria relevant to IBD showing different abundance between the two groups. According to the literature, Enterococcaceae50 and Gammaproteobacteria51 were found to increase in IBD patients, whereas showed significantly lower abundance in the EMS-treated group compared to DSS controls. Conversely, Bacteroides52,53 and Lachnospiraceae50 which were found to significantly decrease in Crohn’s disease patients, were markedly increased in EMS-administrated mice. Lactobacillales was found to increase in the feces of the DSS group, align with reports of increased Lactobacillus abundance in feces of new-onset and treatment-naive Crohn’s disease52. Importantly, the EMS group exhibited significant upregulation of Akkermansia muciniphila (Supplementary Fig. 17b) a mucin-decomposing bacteria that can sustain intestinal homeostasis by alleviating the inflammatory response and reducing intestinal permeability of intestinal pro-inflammatory lipopolysaccharide54,55. Moreover, Blautia was also found notably more abundant in the EMS group. This beneficial strain plays a critical role in modulating the flora growth and intestinal inflammation56, mainly due to its production of bacteriocin that inhibit the colonization of intestinal pathogens such as Clostridium perfringens and vancomycin-resistant enterococci. Additionally, linear discriminant analysis (LDA) effect size showed the statistically significant and biologically relevant species between the two groups (Fig. 6k, Supplementary Fig. 18), The cladogram showed the changes under different taxa from phylum to genus. Finally, the Tax4Fun2 system that can predict the function profile of gut microbiota (Fig. 6l) was applied for functional annotation clustering analysis. We found that the DSS group was functionally enriched in Human Diseases and Environmental Information Processing. However, enrichment in Metabolism, Genetic Information Processing, and Organismal Systems was found in the EMS group, which was consistent with the remission phenotype of the colitis model.
To overcome the limitations of traditional probiotic delivery, most current research focuses on designing protective armor for probiotics to enhance their survival rate or modifying bacterial surface to promote adhesion to the human tissue46,57,58. Nevertheless, these physical and chemical strategies are passive and short-term, which cannot address the colonization issue fundamentally. Here, the oral Express Microcolony Service (EMS) strategy inspired by parallels between bacterial colony development and eukaryotic embryogenesis, possess advantage of economic feasibility, biosafety, delivery efficiency, and treatment reliability. Notably, this advanced system enhances the colonization of EcN which regulates homeostasis of the entire gut microenvironment and promotes formation of a healthier gut microbiota composition, which benefits the alleviation of colitis (Fig. 6m).
Methods
All research conducted for this study complies with the relevant ethical regulations. The Animal Care Committees of the Shanghai Jiao Tong University School of Medicine have approved all the experimental protocols for this work (approval number: B-2021-009).
Bacterial culture
Bacteria strain Escherichia coli Nissle 1917 (EcN, serotype O6:K5:H1, Cat. T0023, Biosci, China) belonged to the species Escherichia coli was cultured in an LB medium. EcN strain expressing red fluorescent protein (REcN) (Shanghai Microbiological Culture Collection, China) was cultured in liquid LB medium (with 50 μg/mL kanamycin). Bacterial resuscitate: Inoculating the bacteria to 3 mL liquid LB medium, then the bacteria were cultured in an incubator shaker (220 rpm at 37 °C) overnight. Then, 500 μL bacteria suspension were transferred into 5 mL LB medium and cultured until OD600 reached 0.50. The bacteria suspension was centrifuged at 3500 g for 5 min to collect for subsequent experiments. To induce expression of red fluorescent protein in REcN, 1 mL bacteria suspension was inoculated into 100 mL liquid medium and cultured until OD600 reached 0.50. Then, adding 1 mL 100 mM isopropyl β-D-thiogalactoside solution and continuously culturing for 4 h to induce red fluorescent protein, the bacteria could be collected by centrifugation for follow-up experiments.
Prokaryotic transcriptomic analysis
Sample preparation: The bacteria suspension was inoculated in 5 mL liquid medium until OD600 reached 0.50, then centrifuged at 3500 g to collect the bacteria precipitation. To collect bacterial colonies from the LB agar medium, the prepared bacterial suspension was extended evenly on the plate and placed in the incubator at 37 °C overnight. The bacterial colonies were scraped off the surface of the agar LB medium with a sterile scraper.
Prokaryotic transcriptome sequencing procedure: (1) Extracting total RNA from bacteria by Trizol reagent; (2) Conducting total bacterial RNA quality detection and removing ribosomal RNA; (3) Breaking the RNA into 200–300 bp fragments; (4) Synthesizing cDNA strand 1 by random primers and reverse transcriptase, the chain specific library was established after the second strand was synthesized. (5) The purified double-stranded cDNA then underwent end-repair, A-tailing, and adapter ligation. AMPure XP beads were used for size selection (300–400 bp) of the fragments. (6) PCR amplification was performed using a thermostable DNA polymerase to cyclically denature, anneal, and extend DNA strands, thereby exponentially increasing the number of target DNA fragments to acquire the cDNA library. (7) The insert size of the cDNA library was detected using the Agilent 2100, and the effective concentration of the library was accurately quantified by amplifying specific sequences using fluorescent detection, ensuring the concentration exceeds 2 nM for subsequent sequencing. (8) Sequencing on Illumina platform. (9) Data analysis: First filtering original data, then comparing with reference EcN genome, calculating the gene expression, and finally conducting expression difference analysis, enrichment analysis, and cluster analysis.
Colon adhesion evaluation
In vitro: Fresh mouse colon was taken, and both ends of colons were tied off with surgical sutures. Bacterial suspension and colony suspension containing same number of bacteria were injected into colonic lumen using a sterile syringe. After incubation for 1 h, unattached bacteria were gently washed away with PBS, and adherent bacteria were examined via in vivo imaging system (IVIS) analysis.
In vivo: 8 weeks C57BL/6 male mice (n = 3 per group) were fasted for 24 h, and the abdominal skin was fully depilated using depilatory cream, then 200 μL of bacteria suspension or colonies suspension were administered by oral gavage. The bacterial adhesion in the colon were measured by fluorescence quantification at 3 h, 6 h, 12 h, and 24 h after oral gavage.
Preparation of alginate methacryloyl (AlgMA)
Dissolving 2 g sodium alginate in 200 mL of 10 mM 2-morpholinoethanesulphonic acid buffer (MES, Aladdin, China); dissolving 380 mg 2-aminoethyl methacrylate hydrochloride (Aladdin, China) in 6 mL MES solution to obtain solution A; dissolving 1.268 g 4-(4,6-dimethoxy-1,3,5, -triazine-2-yl) -4-methylmorpholine hydrochloride hydrate (Aladdin, China) in 6 mL MES solution to obtain solution B. Solution A and B were divided into three times each, and added into sodium alginate solution (Sigma, USA) every 30 min, stirring all solution overnight waiting for reaction. Adding 2 g sodium chloride (Aladdin, China) into the solution the next day and stirring until completely dissolved. Then, put all solution into a dialysis bag (molecular weight cut-off 8000–14000, Solarbio, China) for dialysis (3 days). Finally, all liquid was transferred and freeze-dried in a freeze-drying machine.
Hydrogel preparation for rheometric analysis
Preparing aqueous solutions of alginate (1%) and AlgMA (1%) respectively. Then mixing the two solutions at different proportions including: 0% AlgMA group (containing 100% Alginate), 25% AlgMA group (containing 75% Alginate), 50% AlgMA group (containing 50% Alginate), 75% AlgMA group (containing 25% Alginate), 100% AlgMA group (containing 0% Alginate). A 20 mm diameter round mold was used to prepare the hydrogel. 600 μL hydrogel solution was added to the mold, and 300 μL calcium chloride solution (100 mM) was added to the hydrogel for ionic crosslinking. Irradiation using a 365 nm light source provided the covalent crosslinking. Three groups of hydrogels were obtained by ionic crosslinking, covalent crosslinking, or double crosslinking, respectively, for rheological testing.
Rheometric analysis
The rheological properties of the hydrogels were measured using an AR-G2 Rheometer (TA Instrument) with stainless-steel parallel plate geometry (P20 TiL, 20 mm diameter). Samples of different proportions were prepared for storage modulus, loss modulus, and the stress-relaxation test. 15% constant shear strain was applied to the hydrogel, then measuring the change of stress relaxation within 10,000 s. The timescale at which the stress relaxed to half of the initial level was also analyzed to quantify the stress relaxation rate. Besides, the dynamic measurement includes: (1) Analysis of the storage modulus change under different cross-linking conditions (covalent cross-linking, ionic cross-linking, and double cross-linking); (2) The change of stress relaxation under different AlgMA proportion, and calculating the half decay time (T1/2); (3) Gels were incubated in LB medium in a shaker, and the time-modulus curve was detected at continuous time points.
Swelling-degradation analysis
Swelling-degradation monitoring in LB medium. 0%, 25%, 50%, 75%, and 100% AlgMA hydrogels were all double cross-linked and the initial weight of each gel was obtained before being immersed in the LB medium. We transferred the gel from the liquid using the filter mesh, then quickly blotted the liquid from both sides of the gel using absorbent paper. Subsequently, weighed the gel using a balance and recorded the measurement. The weight of hydrogels was measured at 30 min, 1 h, 3 h, 6 h, and 12 h. Swelling-degradation monitoring in artificial digestive fluid. 0%, 25%, 50%, 75%, and 100% AlgMA hydrogels were immersed into AGF (2 h), ASF (4 h), and ACF (continuous monitoring), respectively, and periodically weighed the hydrogels. The swelling and degradation of hydrogel were analyzed according to the weight change.
Preparation of the micro-cargo of probiotics
Fresh bacteria in the exponential growth period (OD600 = 0.5) were washed and resuspended. The bacteria were added to the prepared hydrogel solution (the final bacterial concentration was 2 × 105, 106, and 5 × 106 CFU/mL) and mixed by vortex. The gas-shearing microfluidic method was employed for preparing the micro-cargo of probiotics. Specifically, the prepared hydrogel solution with bacteria was injected (2 mL/min) into the inner axis of the biaxial needle (inner diameter 0.31 mm, outer diameter 0.82 mm) at a uniform speed using the micropump. The nitrogen (0.4 L/min) from the outer axis cut the solution into uniform droplets by shearing force. The droplets were collected using a 10-cm dish containing 0.1 mol/L CaCl2 aqueous solution and were then crosslinked under a 365 nm light source for about 1 min to acquire double cross-linking hydrogel. Finally, hydrogel micro-cargo was collected and quickly washed with deionized water twice for subsequent experiments.
Morphological characterization of micro-cargo
The prepared hydrogel micro-cargo-containing bacteria were evenly spread in the LB medium and cultured in a 37 °C incubator. The bacteria in the micro-cargo were stained (live bacteria were stained with green fluorescence), and microcolony size progression was monitored using confocal microscopy (Carl Zeiss, Germany). Additionally, after freeze-drying and the gold spray (10 mA, 45 s), the micro-cargo was photographed by scanning electron microscope (Thermo Fisher Scientific, USA).
mRNA expression analysis of bacteria
Fresh bacteria suspension in the exponential growth period (microcolonies in micro-cargo were obtained by degradation of hydrogel) was centrifuged at 3500 g, the bacterial precipitates were washed and centrifuged again. Total RNA was extracted using the Trizol reagent, the other qPCR procedure was as previous. The primer sequences used in the experiment were as follows:
16S forward: AGGCCTTCGGGTTGTAAAGT, reverse: GTTAGCCGGTGCTTCTTCTG;
adrA forward: CTGGCGGAACATAAACGCAG, reverse: ATGCCGCCGACAGTTATCAA;
csgD forward: AACGTGTTGTCAATGGGTTGC, reverse: AGGGCTGATTCCGTGCTGT;
csgA forward: GCGGTAACTCTGCACTTGCTC, reverse: ATCTGAGCCCTGACCAACATC;
flhC forward: GCCCACAAGCAGAAGAAGG, reverse: GGGATGGCGGTTGACATAA;
csrA forward: GATTGGGGATGAGGTCACCG, reverse: CAGAAACTTCCTTCGGGGCA;
luxS forward: CAATCACCGTGTTCGATCTGC, reverse: CGCATAAAACCAGCAAACAGG;
iraM forward: TTTCTCCCTCCTGGCAGTAT, reverse: CCTTGTTGAATGGTGCGATGT;
rpoS forward: CGAAAAAGCGTTGCTGGACA, reverse: GCTCGAACAGCCATTTGACG;
sdiA forward: GATGACTTATTCAGCGAAGCACA, reverse: GCAACGGGAAAAGGACAAAA.
Bacterial proliferation activity
Fresh bacteria suspension in exponential growth period (microcolonies in micro-cargo was obtained by degradation of hydrogel) was centrifuged at 3500 g, the bacterial precipitates was resuspended and diluted to OD600 = 0.2. Then 100 μL diluted bacteria suspension was divided into 96-well plates, and the wells were covered by the respiratory sealing membrane. The OD600 absorbance was detected every 30 min in the microplate reader for microbial growth monitoring.
Cultural optimization of microcolony
The hydrogel solutions with an initial bacterial concentration of 2 × 105, 106, and 5 × 106 CFU/mL were used to prepare the micro-cargo and then cultured in the LB medium. At 0 h, 2 h, 4 h, 6 h, 8 h, 12 h and 24 h, the bacteria were stained, and the changes in microcolony size were observed. The bacteria in the micro-cargo were quantified by OD600 absorbance after hydrogel degradation. The LIVE/DEAD™ BacLight™ bacterial activity detection kit (Thermo Fisher Scientific) was used to stain the bacteria. Micro-cargoes were aspirated from the culture medium, rapidly washed three times with PBS to remove residual liquid and then mixed evenly with the LIVE/DEAD working solution. After incubating in the dark for 15 min, bacterial fluorescence was observed under a confocal microscope (Carl Zeiss, Germany). The live bacteria displayed green fluorescence, and the dead bacteria displayed red fluorescence. The ratio of dead cells to live cells was calculated by fluorescence intensity.
Environmental resistance evaluation
Pre-cultured micro-cargo was treated with artificial gastric fluid (1 h), bile salt solution (30 mg/mL, 4 h), and antibiotic (1% penicillin-streptomycin solution, 4 h), respectively. The micro-cargo was washed three times to remove residual liquid, degraded using 100 mM sodium citrate aqueous solution, and finally collected by centrifugation. The bacterial suspension was diluted in a 10-fold gradient with the bacterial medium, and 10 μL of each group was successively dropped on the LB agar medium. The bacteria’s survival under environmental assaults was counted after 24 h of incubation. The bacteria suspension containing the same number of bacteria and the hydrogel micro-cargo (encapsulated uncultured and dispersed bacteria) containing the same number of bacteria were used as controls.
Bacterial adhesive evaluation
The human colorectal adenocarcinoma cell line Caco-2 (Cat. CL-0050) for this experiment was purchased from Wuhan Procell Life Science & Technology Co., Ltd., Caco-2 cells were cultured in standard 24-well plates at 37 °C in a 5% CO2 incubator. After removing the unattached cells, approximately 107 CFU/mL EcN in the logarithmic growth phase were added to each well and cultured for 3 h. Then washing twice with sterile PBS to remove loosely attached bacteria, and Caco-2 cells were lysed using 0.1% Triton X-100. The lysis buffer in each well was subjected to a series of gradient dilutions and plated on the LB agar medium. The adhesive property of single bacterium and microcolony was evaluated by colony counting, respectively.
Bacterial self-aggregation evaluation
The bacteria in suspension or microcolony from micro-cargo were collected and resuspended. The OD600 absorbance was aligned between groups. 4 mL of each group was put into a tube placed at 37 °C for 18 h. At 3 h, 6 h, 9 h, 12 h, and 18 h, the static culture medium and the resuspended medium in the middle of the test tube were taken out to evaluate the OD600 absorbance.
Bacterial biofilm formation
100 μL bacteria suspension (107 CFU/mL) were inoculated into each well. After 24 h culture, the unattached bacteria were removed by gently washing 2 to 3 times. After drying and fixing at 60 °C for 30 min, 0.1% crystal violet was added to stain. Then, each well was gently washed before being observed under the microscope. For quantification of biofilm, the plate was dried and decolorized with 33% glacial acetic acid. The OD590 absorbance was measured by a microplate reader to quantify the biofilm content. Biofilm thickness was evaluated through the confocal microscope. FilmTracer™ SYPRO™ Ruby biofilm matrix dye (Sigma) was used for biofilm visualization. The staining solution was gently added to avoid damage to the biofilm. The samples were incubated in dark at room temperature before observation.
EPS content evaluation
Sterile slides were placed in the 12-well plate, the bacteria were inoculated (107 CFU/mL) and incubated for 24 h in an incubator. The slide was taken out and covered by 500 μL phenol solution (5%) for sufficient oscillatory elution. Then concentrated sulfuric acid was added and mixed by vibration. The OD490 absorbance was measured to quantify EPS content in the biofilm.
In vivo colonization
6–8 weeks-old C57BL/6 male mice were treated with kanamycin (50 μg/mL) dissolved in drinking water for 1 week. Two days later, the mice were given oral gavage of micro-cargo loaded with EcN microcolonies once, and the control group was given oral gavage of EcN bacteria suspension. The feces of each mouse were collected every day for colonization counting. On the fourth day, all mice were sacrificed, all intestinal tissues (including colon, small intestine, and cecum) were separated for evaluation of bacterial colonization.
Spot assay for bacterial quantification
30 mg feces of each mouse were ground and homogenized in 500 μL LB medium, the supernatant was acquired and diluted in a 10-fold gradient. The gradient-diluted liquid was successively added to the LB agar medium (containing kanamycin), and the colony count was performed after 24 h of incubation. For bacterial colonization of tissue, 50 mg of intestinal tissue (small intestine, cecum, or colon) was prepared and ground in 400 μL LB medium in a tissuelyser (60 Hz, 120 s). The supernatant was acquired and diluted by LB medium and was successively added to the LB agar medium. Colony counts were performed after 24 h of incubation.
Acute colitis model
6–8 weeks C57BL/6 male mice were housed in a specific pathogen-free facility at the Animal Center of Shanghai Jiao Tong University School of Medicine. The facility maintained controlled temperature and humidity with a 12 h light/dark cycle. All mice were randomly divided into seven groups: healthy control, DSS group, DSS + 5-ASA group (positive control), DSS + EMS group (oral gavage of micro-cargo containing EcN microcolonies), DSS + PMS (oral gavage of micro-cargo containing EcN), DSS + EcN (oral gavage of EcN suspension), DSS + MS (oral gavage of blank micro-cargo). On the third day of DSS treatment, the mice were given an oral gavage of 20 mg micro-cargo. The mice were sacrificed 2 to 4 days after DSS drinking stopped. The change of body weight and the disease activity index were continuously measured as previously reported59,60, the feces were separately collected for colonization evaluation, the colon was dissected for length measurement and then prepared for mRNA detection and H&E staining.
16S rRNA sequencing
All fecal samples were subjected to DNA extraction, PCR amplification, product purification, library preparation and inspection, and finally NovaSeq sequencing. The raw data were processed to generate ASV, and each ASV sequence was annotated for the corresponding species information and species-based abundance distribution. Based on this, the subsequent sample diversity analysis, the difference in community structure between groups, and the significance test of difference were conducted. In addition, functional prediction of the microbiome based on functional databases was also carried out.
Statistical analysis
Statistics were mainly analyzed via the GraphPad Prism 8.4 (USA). Quantitative data are displayed as mean ± standard deviation (SD), the categorical data are displayed as numbers (percentages). Statistical analysis between two groups was performed by the two-tailed Student’s t-test or Mann–Whitney U-test. Finally, p < 0.05 was considered statistically significant.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The 16 S rRNA sequencing data and RNA sequencing data generated in this study are deposited in the National Genomics Data Centers’ Genome Sequence Archive (GSA) database (https://ngdc.cncb.ac.cn/), under accession code CRA027249 and CRA021170. Source data are provided with this paper.
References
-
O’Toole, P. W., Marchesi, J. R. & Hill, C. Next-generation probiotics: the spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2, 17057 (2017).
-
Miao, G. et al. RemodeliNg Intestinal Microbiota Alleviates Severe Combined Hyperlipidemia-induced Nonalcoholic Steatohepatitis And Atherosclerosis in LDLR-/- Hamsters. Research 7, 0363 (2024).
-
Li, N. et al. The role of gut microbiota associated metabolites in digestive disorders. Eng. Regener. 5, 228–246 (2024).
-
Veiga, P., Suez, J., Derrien, M. & Elinav, E. Moving from probiotics to precision probiotics. Nat. Microbiol. 5, 878–880 (2020).
-
Guyonnet, D., Schlumberger, A., Mhamdi, L., Jakob, S. & Chassany, O. Fermented milk containing Bifidobacterium lactis DN-173 010 improves gastrointestinal well-being and digestive symptoms in women reporting minor digestive symptoms: a randomised, double-blind, parallel, controlled study. Br. J. Nutr. 102, 1654–1662 (2009).
-
Yin, W., Wang, Y., Liu, L. & He, J. BiofilMs: The Microbial “Protective Clothing” In Extreme Environments. Int. J. Mol. Sci. 20, 3423 (2019).
-
Caballero-Flores, G., Pickard, J. M. & Núñez, G. Microbiota-mediated colonization resistance: mechanisms and regulation. Nat. Rev. Microbiol. 21, 347–360 (2023).
-
Sharahi, J. Y. et al. Advanced strategies for combating bacterial biofilms. J. Cell Physiol. 234, 14689–14708 (2019).
-
Futo, M. et al. Embryo-Like Features In Developing Bacillus Subtilis Biofilms. Mol. Biol. Evol. 38, 31–47 (2021).
-
van Gestel, J., Vlamakis, H. & Kolter, R. Division of labor in biofilms: the ecology of cell differentiation. Microbiol. Spectr. 3, Mb-0002–Mb-2014 (2015).
-
Vlamakis, H., Chai, Y., Beauregard, P., Losick, R. & Kolter, R. Sticking together: building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 11, 157–168 (2013).
-
Whiteley, M., Diggle, S. P. & Greenberg, E. P. Progress in and promise of bacterial quorum sensing research. Nature 551, 313–320 (2017).
-
Kumar, S. et al. Delineating virulence of Vibrio campbellii: a predominant luminescent bacterial pathogen in Indian shrimp hatcheries. Sci. Rep. 11, 15831 (2021).
-
Yan, J. & Bassler, B. L. Surviving as a community: antibiotic tolerance and persistence in bacterial biofilms. Cell Host Microbe 26, 15–21 (2019).
-
Liu, H. et al. Advance in oral delivery of living material. Biomed. Technol. 3, 26–39 (2023).
-
Yu, Y., Wang, Q., Wang, C. & Shang, L. Living Materials for Regenerative Medicine. Eng. Regener. 2, 96–104 (2021).
-
Yang, N. et al. Emerging microfluidics for the modeling and treatment of arthritis. Eng. Regener. 5, 153–169 (2024).
-
Pan, C. et al. Polymerization-mediated multifunctionalization of living cells for enhanced cell-based therapy. Adv. Mater. 33, e2007379 (2021).
-
Luo, H. et al. Chemical reaction-mediated covalent localization of bacteria. Nat. Commun. 13, 7808 (2022).
-
Cao, Z. & Liu, J. Surface nanocoating of bacteria as a versatile platform to develop living therapeutics. Nat. Protoc. 19, 3162–3190 (2024).
-
He, J. et al. Honeycomb-like hydrogel microspheres for 3D bulk construction of tumor models. Research 2022, 9809763 (2022).
-
Martínez-Calvo, A. et al. Morphological instability and roughening of growing 3D bacterial colonies. Proc. Natl. Acad. Sci. USA 119, e2208019119 (2022).
-
Butelmann, T. et al. Metabolism control in 3D-printed living materials improves fermentation. ACS Appl. Bio Mater. 4, 7195–7203 (2021).
-
Chrisnandy, A., Blondel, D., Rezakhani, S., Broguiere, N. & Lutolf, M. P. Synthetic dynamic hydrogels promote degradation-independent in vitro organogenesis. Nat. Mater. 21, 479–487 (2022).
-
Chaudhuri, O. et al. Hydrogels with tunable stress relaxation regulate stem cell fate and activity. Nat. Mater. 15, 326–334 (2016).
-
Chaudhuri, O., Cooper-White, J., Janmey, P. A., Mooney, D. J. & Shenoy, V. B. Effects of extracellular matrix viscoelasticity on cellular behaviour. Nature 584, 535–546 (2020).
-
Bhusari, S., Sankaran, S. & Del Campo, A. Regulating bacterial behavior within hydrogels of tunable viscoelasticity. Adv. Sci. 9, e2106026 (2022).
-
Bhusari, S. et al. Rheological behavior of Pluronic/Pluronic diacrylate hydrogels used for bacteria encapsulation in engineered living materials. Soft Matter 20, 1320–1332 (2024).
-
Kuchma, S. L. & O’Toole, G. A. Surface-induced and biofilm-induced changes in gene expression. Curr. Opin. Biotech. 11, 429–433 (2000).
-
Rembacken, B. J., Snelling, A. M., Hawkey, P. M., Chalmers, D. M. & Axon, A. T. Non-pathogenic Escherichia coli versus mesalazine for the treatment of ulcerative colitis: a randomised trial. Lancet 354, 635–639 (1999).
-
Pujo, J. et al. Bacteria-derived long chain fatty acid exhibits anti-inflammatory properties in colitis. Gut 70, 1088–1097 (2021).
-
Teng, G. et al. Probiotic Escherichia coli Nissle 1917 expressing elafin protects against inflammation and restores the gut microbiota. Front Microbiol. 13, 819336 (2022).
-
Liu, L. et al. AHL-mediated quorum sensing to regulate bacterial substance and energy metabolism: a review. Microbiol. Res. 262, 127102 (2022).
-
Liu, X. et al. Roles and opportunities of quorum sensing in natural and engineered anaerobic digestion systems. Water Res. 275, 123190 (2025).
-
Vandana & Das, S. Genetic regulation, biosynthesis and applications of extracellular polysaccharides of the biofilm matrix of bacteria. Carbohydr. Polym. 291, 119536 (2022).
-
Wood, T. K. Insights on Escherichia coli biofilm formation and inhibition from whole-transcriptome profiling. Environ. Microbiol. 11, 1–15 (2009).
-
Yim, J. et al. Transcriptional profiling of the probiotic Escherichia coli Nissle 1917 strain under simulated microgravity. Int. J. Mol. Sci. 21, 2666 (2020).
-
Liou, G. G., Chao Kaberdina, A., Wang, W. S., Kaberdin, V. R. & Lin-Chao, S. Combined transcriptomic and proteomic profiling of E. coli under microaerobic versus aerobic conditions: the multifaceted roles of noncoding small RNAs and oxygen-dependent sensing in global gene expression control. Int. J. Mol. Sci. 23, 2570 (2022).
-
Roth, J. G. et al. Tunable hydrogel viscoelasticity modulates human neural maturation. Sci. Adv. 9, eadh8313 (2023).
-
Darnell, M. et al. Material microenvironmental properties couple to induce distinct transcriptional programs in mammalian stem cells. Proc. Natl. Acad. Sci. USA 115, E8368–e8377 (2018).
-
Yang, S. et al. Fabricated technology of biomedical micro-nano hydrogel. Biomed. Technol. 2, 31–48 (2023).
-
Peruzzi, J. A., Galvez, N. R. & Kamat, N. P. Engineering transmembrane signal transduction in synthetic membranes using two-component systems. Proc. Natl. Acad. Sci. USA 120, e2218610120 (2023).
-
Xu, T. et al. Functional and structural diversification of incomplete phosphotransferase system in cellulose-degrading clostridia. ISME J. 17, 823–835 (2023).
-
Shukla, P. S., Agarwal, P., Gupta, K. & Agarwal, P. K. Molecular characterization of an MYB transcription factor from a succulent halophyte involved in stress tolerance. AoB Plants 7, plv054 (2015).
-
Alotaibi, G. F. Factors influencing bacterial biofilm formation and development. Am. J. Biomed. Sci. Res. 12, 617–626 (2021).
-
Wang, X. et al. Bioinspired oral delivery of gut microbiota by self-coating with biofilms. Sci. Adv. 6, eabb1952 (2020).
-
Scaldaferri, F. et al. Role and mechanisms of action of Escherichia coli Nissle 1917 in the maintenance of remission in ulcerative colitis patients: an update. World J. Gastroenterol. 22, 5505–5511 (2016).
-
Chen, H. et al. Advances in Escherichia coli Nissle 1917 as a customizable drug delivery system for disease treatment and diagnosis strategies. Mater. Today Bio 18, 100543 (2023).
-
Luo, X. et al. Encapsulation of Escherichia coli strain Nissle 1917 in a chitosan-alginate matrix by combining layer-by-layer assembly with CaCl(2) cross-linking for an effective treatment of inflammatory bowel diseases. Colloids Surf. B Biointerfaces 189, 110818 (2020).
-
Schirmer, M., Garner, A., Vlamakis, H. & Xavier, R. J. Microbial genes and pathways in inflammatory bowel disease. Nat. Rev. Microbiol 17, 497–511 (2019).
-
Morgan, X. C. et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 13, R79 (2012).
-
Gevers, D. et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe 15, 382–392 (2014).
-
Guo, X. et al. Gut microbiota is a potential biomarker in inflammatory bowel disease. Front Nutr. 8, 818902 (2021).
-
Zhai, Q., Feng, S., Arjan, N. & Chen, W. A next generation probiotic, Akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 59, 3227–3236 (2019).
-
He, K. Y. et al. Akkermansia muciniphila protects the intestine from irradiation-induced injury by secretion of propionic acid. Gut Microbes 15, 2293312 (2023).
-
Liu, X. et al. Blautia-a new functional genus with potential probiotic properties?. Gut Microbes 13, 1–21 (2021).
-
Cao, Z., Wang, X., Pang, Y., Cheng, S. & Liu, J. Biointerfacial self-assembly generates lipid membrane coated bacteria for enhanced oral delivery and treatment. Nat. Commun. 10, 5783 (2019).
-
Liu, Y. et al. Improved gastric acid resistance and adhesive colonization of probiotics by mucoadhesive and intestinal targeted konjac glucomannan microspheres. Adv. Funct. Mater. 30, 2001157 (2020).
-
Liu, H. et al. Platelet membrane fragment self-assembled oral hydrogel microspheres for restoring intestinal microvascular injury. Adv. Funct. Mater. 33, 2302007 (2023).
-
Liu, H. et al. Colon-targeted adhesive hydrogel microsphere for regulation of gut immunity and flora. Adv. Sci. 8, 2101619 (2021).
Acknowledgements
This work was supported by the Noncommunicable Chronic Diseases-National Science and Technology Major Project (2023ZD0500700 to W.C.), National Natural Science Foundation of China (32101104 to Z.C. and 32470951 to Z.W.), Shanghai Municipal Health Planning Commission (202140127 to Z.C. and 202340092 to Z.W.), Shanghai university teachers’ production, study and practice plan (RC20230003 to H.L.), Shanghai Committee of Science and Technology (22ZR1439600 to Z.W.), The Seed Program for Research and Translation of New Medical Technologies, Shanghai Municipal Health Commission (2024ZZ2028 to Z.C.).
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Liu, H., Chen, Z., Lin, Q. et al. A multicellular self-organized probiotic platform for oral delivery enhances intestinal colonization. Nat Commun 16, 7060 (2025). https://doi.org/10.1038/s41467-025-62349-x
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41467-025-62349-x