
Introduction
In the absence of global change, it is estimated that the world will need up to 56% more food produced by 2050 relative to 20101. This requirement will be substantially elevated by crop losses caused by increasing extreme climate events2. Current rates of crop yield increase are insufficient to meet this future requirement and new innovations are therefore needed. Photosynthesis, given its low efficiency in crops relative to the theoretical maximum, has emerged as a promising target for bioengineering. Transgenic up-regulation of specific photosynthetic enzymes has already been shown to increase photosynthesis and in turn the yields of rice and soybean in the field3,4. Here we demonstrate a different approach that would substantially increase crop photosynthetic productivity.
Higher plant photosynthesis is primarily driven by photons in the visible spectrum (400–700 nm). While photons in the far-red (FR) spectral region (700–800 nm) are generally insufficient to sustain the full operation of the entire photosynthetic electron transport chain, since they are not absorbed by Photosystem II, FR photons contain sufficient energy to drive charge separation in reaction centers and, in turn, CO2 assimilation5. Indeed, some species of cyanobacteria grow autotrophically in FR light6,7,8, and do so using different strategies9.
These cyanobacteria modify pigment type and composition in response to an increasing ratio of FR to visible light in a process called far-red light photoacclimation (FaRLiP). In visible light, they use chlorophyll a (Chl a), as in higher plants, but when grown in environments depleted of visible light but rich in FR light, the longer-wavelength-absorbing chlorophyll d (Chl d) and f (Chl f) are synthesized10. For example, when grown under FR light, the FaRLiP-capable cyanobacterium, Chroococcidiopsis thermalis contains ~90% Chl a, ~10% Chl f, and <1% Chl d5. Chl f is associated with both photosystems, where it harvests light and acts as the primary electron donor in Photosystem I (PSI)11, while Chl d acts as the primary electron donor in Photosystem II (PSII)6. The presence of these FR absorbing Chls allows these species to utilize photons in the 700–800 nm range with high efficiency, albeit slightly lower than for visible light in the Chl a-containing photosystems12 This is due to the fact that the energy harvested by Chl f needs to be transferred to the reaction center and the transfer involves some uphill steps that lower the efficiency, particularly in PSII12,13,14,15. PSI, however, still works with more than 90% quantum efficiency, even when using 800 nm photons5,16.
Most members of the cyanobacterial genus Acaryochloris use a different strategy to grow in FR light: these species contain mainly Chl d (>90%), independent of the growth radiant energy spectrum, and have a far larger capacity to use FR light of up to 750 nm for photosynthesis8,17. The almost exclusive presence of Chl d results in a high energetic connectivity in both photosystems of Acaryochloris, which therefore reach higher photochemical yields than the photosystems of Chroococcidiopsis thermalis15,18.
Incorporation of the FR light adaptations of these cyanobacteria into crop plants has been frequently suggested as a potential strategy to increase photosynthesis and crop productivity, thereby improving crop yields9,19. However, the magnitude of this benefit has not previously been estimated, despite this being key in determining how worthwhile such an effort would be versus the other opportunities for increasing crop yields via improving other steps of the photosynthetic process20.
Adding far-red absorbing capacity would likely have different effects across the canopy, which need to be taken into account. Leaves at the top of crop canopies capture most (80–90%) incoming visible light and their photosynthesis can be light-saturated for much of the day. Therefore, in full sunlight, capturing the additional energy of FR light in these already photon-saturated leaves would only increase the probability of photodamage. However, in modern crop canopies with several layers of leaves, those in the lower canopy are starved of visible photons for most of the day, excepting a few sunflecks, and so contribute little to crop photosynthesis, net carbon gain, and yield21. Leaves transmit >90% of FR photons, so while these lower canopy leaves are starved of visible photons, they are bathed in FR photons22. As such, the ratio of FR/visible photons rises from <1 at the top of a rice canopy, to 5–6 at the fifth layer of leaves23. One potential strategy for increasing the photosynthetic rate would be to add FR-absorbing chlorophylls to the lower leaves in the canopy, allowing them to use these FR photons.
Averaged over the course of a day, the vertical profile of light extinction through a crop canopy appears similar to that of a solution of chlorophylls or suspension of cyanobacteria24,25. However, this analogy does not account for the complexity imposed by the high heterogeneity of radiant energy and its composition with time and space within a crop canopy, compared to suspensions of cyanobacteria. This is because the solar angle changes throughout the day, such that leaves within the canopy transition between sun and shade as they move in and out of the shadows of overlying leaves. Soybean is one of the top four crops globally, in terms of total seed yield. It is the number one world source of vegetable protein and number two source of vegetable oil26. With worldwide rising demand, increasing its productivity per unit land area in use will reduce land pressure and associated environmental risks27. Importantly soybean provides a canopy that is a good proxy for most major crops.
Here, we use soybean as an example to address canopy heterogeneity and assess the impact of introducing Chl d and Chl f on canopy level CO2 assimilation. We use a reconstruction of a field crop of soybean to determine the diurnal 3D course of photon flux in different wavelengths at all points within the canopy. By coupling the temporal and spatial dynamics of photon flux with a model of the response of CO2 assimilation and microclimate to absorbed photon flux, the daily carbon gain of the canopy is estimated. This framework is then used to determine the additional photosynthetic carbon gain that would result from the addition of Chl d and Chl f. Furthermore, we simulate and compare two strategies of production of FR-absorbing chlorophylls through the canopy, based on the two chlorophyll compositions observed in Chroococcidiopsis thermalis and Acaryochloris.
Results
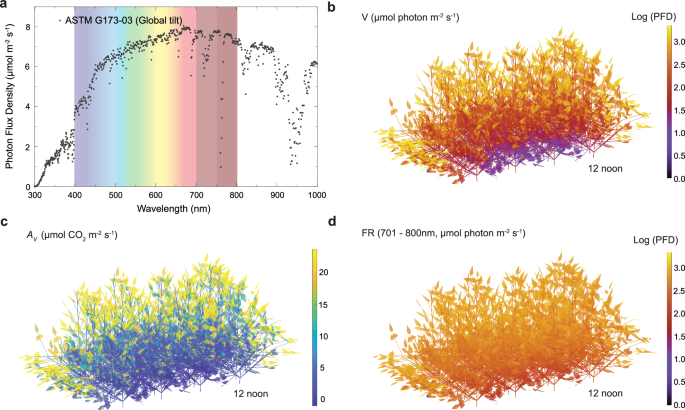
Initially, the visible and far-red light environment in a mature soybean canopy was assessed. This was determined by simulating, in 3D, photon absorption throughout the canopy over the full photoperiod. This model separately tracked blue, green, red, and FR photons, accounting for the different leaf transmittance and reflectance properties to these wavelengths (Fig. 1b, d, Table 1). Figure 1b demonstrates the incident visible photon flux density of leaves in a mature soybean canopy, while Fig. 1d depicts the incident FR photons. In the soybean canopy, the upper layers receive most visible photons (Fig. 1b), which achieve high CO2 assimilation rates (Fig. 1c). However, the lower leaves mainly receive FR photons (Fig. 1d), which barely drive photosynthetic electron transport in soybean28, resulting in near zero CO2 assimilation rate (Fig. 1c) in the lower canopy with the current pigment composition.

a Photon flux density (PFD) of a solar spectrum at top of the canopy, as defined by the standards of an inclined plane at a 37° tilt toward the equator facing the sun (https://www.nrel.gov/grid/solar-resource/spectra-am1.5.html). The PFD at each wavelength was calculated from the solar spectrum’s energy and the energy per photon at that wavelength (Eq. (1)). Shaded areas correspond to the 400–700 nm range (continuous spectrum), 700–750 nm (red) and 750–800 nm (burgundy). b The sum of incident visible (V) photon flux distribution in a soybean canopy (leaf area index of 6.9), including blue (400–500 nm), green (501–600 nm) and red (601–700 nm) photon; c The leaf net CO2 assimilation rate utilizing visible photon (AV) in the canopy. d Incident far-red photon (701–800 nm) in the soybean canopy with leaf FR absorption fraction of 0.1 (AbsFR=0.1). Source data are provided as a Source Data file.
Enhancing FR absorption with Chl d
In contrast to Chl a-containing soybean leaves, the transmittance and reflectance spectra of both extracted Chl d and Acaryochloris marina cells, show effective absorption of visible photons and FR photons between 700 and 750 nm (Supplementary Fig. 1 and Supplementary Table 1). Extracted Chl d in methanol shows 80% absorption at 725 nm, declining rapidly on the red side. However, the fraction of photons absorbed at 740 nm of Chl d-containing Acaryochloris marina cells is 80% at 740 nm (Supplementary Fig. 1c). This difference indicates a red-shift toward longer wavelength absorption of about 15 nm, resulting from the incorporation of the Chl d into chlorophyll binding proteins. Consequently, Chl d-containing photosynthetic proteins potentially absorb most of the FR photons in the 701–750 nm (Supplementary Fig. 1c and Supplementary Table 1). By considering the photon flux within this region of the solar spectrum (Fig. 1a), the addition of Chl d to the soybean canopy would increase the number of utilizable photons by 18.3% (Fig. 1a, Table 1). This suggests a great potential and scope for optimizing crop photosynthesis by utilizing Chl d.
To assess the impact of Chl d on canopy photosynthesis across various soybean growth stages, simulations were conducted every 15 days, from dawn to dusk, starting from the first trifoliolate (a growth stage of V1, the day of year, i.e. DOY, 171) stage to the beginning of pods development (R3, DOY 231) (Fig. 2a). In these simulations, different capacities of FR absorption were tested, while maintaining a constant FR absorption fraction (AbsFR) across all leaves from the top to the bottom of the canopy in each simulation. In this work we have frequently used the absorption fraction (Abs), which is the percentage of photons absorbed in a given spectral range. For example, AbsFR = 0.1 is equal to 10% of FR photons absorbed.

a The soybean canopy of different date of year (DOY). Leaf area index (LAI) was 0.05, 0.49, 2.1 6.1 and 6.9 for the 5 simulated dates, respectively. b, c Present the Ac and the absorbed light of the soybean canopy at different times of the day. d Shows the effect of different AbsFR settings on daily average light absorption, Ac, Ec and WUE. V visible photons, FR far-red photons. AbsFR, leaf FR absorption fraction. Source data are provided as a Source Data file.
Results indicate that the increase in net CO2 assimilation rate correlates positively with soybean growth (Fig. 2b, d). During the vegetative stages (DOY 171, 186, and 201), the increase in photosynthesis due to increased FR photon absorption is minimal and outpaces by the rise in transpiration (Fig. 2d), resulting in a slight decrease in water use efficiency (Fig. 2d). However, as the plants entered the reproductive stages (DOY 216 and DOY 231), the benefits of utilizing Chl d become more apparent. Simulations showed that at the early growth stages the canopy FR absorption and net CO2 assimilation rate increase linearly with increasing AbsFR (Fig. 2c, d), whereas the maximum enhancement in net CO2 assimilation rate reaches a maximum at an AbsFR of 0.4 and 0.3 on DOY 216 and DOY 231, respectively. Further increases in AbsFR then results in progressively smaller improvement in canopy CO2 assimilation (Ac) (Fig. 2d).
Pod filling is driven by the photosynthesis occurring during the reproductive phase of growth29. Therefore, the increase in photosynthesis resulting from addition of FR-absorbing Chl during this period is the best guide to the likely increase in yield. The increased photosynthesis during the vegetative stage could improve crop establishment and may allow more pod initials when the reproductive phase commences. Our estimates of yield gain based on photosynthesis during the reproductive phase might therefore be regarded as a minimum.
Two strategies were compared for extending the absorption using Chl d in the reproductive stage.
Strategy 1: The addition of Chl d to the same amount in all leaves. Based on the above results an optimal AbsFR of 0.3 was chosen (Fig. 3a). Simulations showed that this addition results in a 13.5 % increase in daily average assimilation of CO2 and a 13.8% increase in photon absorption (Fig. 3g, h and Supplementary Table 2). Although transpiration rises 12.9%, the daily average water use efficiency improves by 0.6%. Leaves at the top of the canopy absorb much more FR photons compare to the middle and bottom leaves (Fig. 3c).

Two different strategies for setting leaf absorption fraction of FR (AbsFR) were tested: Strategy 1 (a, c, e) with constant AbsFR, and Strategy 2 (b, d, f) with AbsFR varies linearly with the(,{{Ratio}}_{{FR}/{R}}) (({{Abs}}_{{FR}}=kcdot {{Ratio}}_{{FR}/R})) in the soybean canopy(Aug 18th, 2023, in Champaign, IL, USA). a, b Display the AbsFR values at various canopy heights. Simulated daily average incident far-red (730 ± 5 nm) to red (667 ± 5 nm) photon flux ratio (FR/R ratio) for each leaf pixel (blue dots) using ray tracing algorithm, with the average incident FR/R ratio of each canopy layer (every 10 cm in height, Supplementary Table 5) marked by black dots. Leaf FR absorption fraction was set as 0.3. k is set to 0.056, which was obtained by scaling the AbsFR at bottom of the canopy to the value of 0.86, which is the maximum absorption fraction from the measurement (Supplementary Fig. 1, Supplementary Tables 1, 6). c, d Simulated far-red photons (FR, 701–750 nm) absorption of a soybean canopy with the two strategies respectively. e, f The enhancement of leaf net CO2 assimilation with additional far-red photon (AV+FR-AV). g Depicts the Ac and h presents the absorbed light of the soybean canopy at different times of the day for both strategies. Source data are provided as a Source Data file.
Strategy 2: Increasing Chl d content and, in turn, AbsFR of leaves in response to the FR/red (FR/R) light ratio at different canopy depths (Fig. 3b), so that AbsFR is maximized in the lowest leaves. This strategy offers the advantage of being implementable by regulating Chl d synthesis through the control of phytochromes, which detect the ratio of FR (730 nm) to R (667 nm)30. We simulated change in the phytochrome absorbing wavelengths of FR (730 ± 5 nm) to R (667 ± 5 nm) in the soybean canopy. The average ratio at different heights is calculated and AbsFR is linked to this ratio (Fig. 3b). The maximum AbsFR of 0.86 (Fig. 3b) is obtained from the average transmission spectrum of Chl d between 701 and 750 nm when the optical density (OD) at the absorption maximum was equal to 5. This OD value was chosen to match the typical total chlorophyll OD in leaves (Supplementary Table 1).
This strategy results in a daily average increase in Ac of 13.7%, with a daily average increase in photon absorption of 11.6% (Fig. 3g, h and Supplementary Table 2). Additionally, the daily average water use efficiency improves by 0.7%. The simulated results with a spectral resolution of 50 nm are identical to those obtained with a finer spectral resolution of 10 nm for the FR range of 701–750 nm (Supplementary Fig. 2. Compared to Strategy 1, the canopy absorbs less FR photons in Strategy 2 (Fig. 3g), because AbsFR in the upper leaves is smaller, and less FR photons are absorbed (Fig. 3c, d). Consequently, more FR photons are transmitted to the canopy’s deeper layers. This increase in FR absorption at greater depths enhances the CO2 uptake in the visible photon-starved leaves of the lower canopy (Fig. 3e, f).
Enhancing FR absorption with Chl f
Even more FR photons might be harvested by using Chl f rather than Chl d, which in FaRLiP PSI was shown to absorb and efficiently use wavelengths up to 800 nm16, although most of the absorption occurs in the region up to 760 nm. If Chl f could be incorporated into the soybean canopy, the number of photons utilizable in photosynthesis would be increased by 21.9% if the upper wavelength limit is assumed to be 760 nm, or by 35.7% if further extended to 800 nm. This increase in light absorption also correlated positively with soybean growth (Supplementary Fig. 3). In FaRLiP cyanobacteria, Chl f is 10% of the total chlorophyll content. Analogous to the cyanobacteria, we assume a leaf absorption fraction (AbsFR) of 0.1. Here, both canopy CO2 assimilation and light absorption are enhanced throughout the day in the reproductive stage (DOY 231) (Fig. 4a, b). The daily average increases in Ac are 11.4 % and 18.4% and photon absorption of 9.4% and 15.3%, respectively for upper wavelength limits of 760 nm and 800 nm (Supplementary Fig. 3 and Supplementary Table 3). The daily average water use efficiency increases by 0.6% and 0.9% for these two wavelength ranges, respectively (Supplementary Table 3). The impact of the Chl f utilization on the CO2 assimilation in each leaf in the canopy can be better appreciated in Fig. 4c. The additional absorption of FR photon increases the photosynthetic rate predominantly in the middle layers of the canopy.

Simulated canopy net CO2 assimilation rate (Ac) and absorbed light throughout the day (Aug 18th, 2023, Champaign, IL, US) in a soybean canopy with constant AbsFR of 0.1, and with AbsFR varies with the ({{Ratio}}_{{FR}/R})(({{Abs}}_{{FR}}=kcdot {{Ratio}}_{{FR}/R})), where k is set to 0.056. a, b Show Ac and absorbed light changes with the two strategies. Leave with Chl f used FR photons (701–760 or 800 nm). c, d The enhancement of leaf net CO2 assimilation from additional FR photons (AV+FR– AV) at noon, where FR represents in FR photons of 701–800 nm. Source data are provided as a Source Data file.
If the synthesis of Chl f is controlled by the FR/R photon ratio (as in Strategy 2 above for Chl d utilization), then photosynthetic CO2 uptake is increased further in both the middle and lower layers of the canopy (Fig. 4d). The average daily increase in Ac with this strategy would be 16.3% and 26.1% for upper wavelength limits of 760 nm and 800 nm, respectively (Fig. 4a, and Supplementary Table 3) and daily average increases in photon absorption are 13.9% and 22.7% (Fig. 4b, and Supplementary Table 3). The daily average water use efficiency enhances 0.8% and 1.2% (Supplementary Table 3). Although additional light absorption raises leaf temperature (Supplementary Fig. 4), the implementation of the two strategies we proposed ensures that the increase in leaf temperature, compared to using only visible light, remains within 1 °C at noon (Supplementary Figs. 5 and 6). The boundary layer conductance we used was close to that of leaves measured under the typical average windspeed during the Illinois soybean growing season. Note that the organically rich moist soils typical of the soybean growing region of the US Midwest absorb 85% of incident FR between 700 nm and 800 nm31. Therefore, what FR is not absorbed by the canopy will be absorbed by the soil, so heating of the crop stand should be similar regardless of the presence of chlorophylls absorbing in these wavelengths.
Discussion
Crop leaves typically absorb 90% of visible photons, but transmit or reflect most FR. However, FR photons are still able to drive photosynthetic carbon assimilation, as demonstrated in A. marina and FaRLiP cyanobacteria, where photosystems incorporate Chl d or/and Chl f to utilize FR photons up to 750 nm and 800 nm, respectively6,13. Integration of these Chls, and additional components needed for their function, into crop plants could substantially expand their photosynthetically active spectrum32, promising enhanced growth and productivity. However, the precise extent of these potential benefits has remained unquantified until now. Two cyanobacterial adaptation strategies allowing FR photon utilization offer insights. First, Acaryochloris marina primarily uses Chl d (>90%)33,34,35, effectively utilizing FR from 700 to 750 nm. Second, FaRLiP cyanobacteria like Chroococcidiopsis thermalis, containing mostly Chl a and small amounts of Chl f (ca. 10%), can utilize FR light from 700 to 800 nm5. These cyanobacterial approaches were independently evaluated using a soybean canopy model to focus on photon absorption and photosynthetic efficiency over the course of a day, providing a basis for generalized predictions in other crops. Our simulations indicated that incorporating Chl d and Chl f into plants could increase productivity by as much as 26%, which if achieved would be massive compared to current rates of improvement of around 1–2% per year, and achieved only in the most intensively bred crops36.
By incorporating Chl d into photosynthetic proteins (photosystems and antenna), the FR absorption of leaves could be dramatically increased. In particular, the measurements on Acaryochloris marina cells have shown that Chl d-containing photosynthetic proteins potentially absorb 86% of FR photons (701–750 nm, Supplementary Fig. 1c and Supplementary Table 1). Using this FR absorption fraction as the maximum limit, we explored two FR utilization strategies in crop canopies: constant leaf FR absorption fraction across the canopy (Strategy 1, Fig. 3a) and increasing absorption as the FR/R photon ratio rises with depth into the canopy (Strategy 2, Fig. 3b). Strategy 2 focuses on enhancing FR absorption predominantly in the middle and lower parts of the canopy (Fig. 3d), where leaves typically receive limited visible light. This targeted increase in light absorption (Fig. 3b), effectively improves photon distribution throughout the canopy. The strategy results in an estimated 13.7% increase in the net daily canopy CO2 assimilation (Ac), with an 11.6% increase in light absorption (Supplementary Table 2, Fig. 3g, h). While the increase in Ac is similar for both strategies, Strategy 2 reduces the risk of photoinhibition, photodamage and heating of the uppermost leaves, presenting a significant advantage over Strategy 1.
Incorporating Chl f into crop leaves could enable absorption and utilization of a broader FR range, from 700 up to 800 nm. However, given that Chl f comprises only about 10% of the total Chl content in Chroococcidiopsis thermalis, we mimicked this strategy by limiting the amount of Chl f incorporated into crop leaves. Our simulations suggest that even a low absorption fraction of 0.1 could boost Ac up to 18.4% (Fig. 4a, Supplementary Table 3). If Chl f’s induction is also linked to FR/R increase with depth into the canopy (as with Strategy 2 of Chl d), simulated daily Ac increases to 26.1 % for upper wavelength limits of 800 nm (Supplementary Table 3).
To optimize the use of FR photons in photosynthesis through Chl d and Chl f, we evaluated the impact of these chlorophylls across various growth stages. Inducing Chl d/f at vegetative stage might not be fully beneficial (Fig. 2 and Supplementary Fig. 3) as the canopy isn’t fully developed and light penetration is not yet limited. Additionally, the water uses efficiency declines with increased absorption of FR photon, which could adversely affect soybean growth in regions with inadequate precipitation or limited irrigation capacity. Our proposed Strategy 2 involves enhancing the expression of Chl d or Chl f at the reproductive stage, particularly in the lower parts of the canopy where there are less visible photons but a higher proportion of FR photons. Adjusting the expression levels of Chl d or Chl f in response to the FR/R ratio can significantly improve light use efficiency of the canopy (Supplementary Tables 2 and 3). The improvement in CO2 assimilation can be achieved under well-watered conditions, as transpiration may also rise with increased light absorption. However, the overall water use efficiency of the modified soybean canopy shows a slight improvement (Supplementary Tables 2 and 3), which slightly reduces the cost of water used per unit of CO2 assimilation. Consequently, this strategy not only boosts productivity but also has the potential to enhance yield per unit of water used, i.e. increase sustainability.
This adaptive expression can be mediated by phytochromes, which are light-sensing proteins found in both plants and cyanobacteria37. FaRLiP cyanobacteria use the red/far-red, knotless phytochrome, RfpA, to detect FR light and trigger the signal cascade that allows transcription of the FaRLiP gene cluster which results in the accumulation of Chl f in the photosystems. Similarly, plants phytochromes similarly detect shade through changes in the ratio of FR/R, triggering specific gene expression responses to adapt to light conditions38. Leveraging phytochrome-mediated transcriptional regulation provides a practical method to bioengineer increased Chl d or Chl f content as shade intensifies with depth into the canopy19,39.
We have shown how incorporation of FR absorbing chlorophylls could increase crop carbon assimilation by 26% in the critical pod-filling phase of soybean growth. A perennial question is whether increased photosynthesis will actually result in increased yield. For soybean, as for wheat and rice, countless experiments have shown that when photosynthesis is artificially increased by elevation of CO2 concentration around the crop in the field a concomitant increase in yield is obtained40,41,42. Similarly, where photosynthetic efficiency has been increased by transgenic modifications significant yield increases have been achieved in the field3,4,43. One concern is that more yield will require more resource, in particular nitrogen. When soybean photosynthesis, and in turn yield, was increased by transgenic improvement of photosynthesis or in separate experiments by season-long growth in the field under elevated CO2 the protein and nitrogen content of the seed was unchanged, despite yield increases of 20% or more4,40,44. Since, in common with agronomic practice, no fertilizer was added in these field experiments, this parallel increase in nitrogen could only be explained by a portion of the additional photosynthate fueling additional N-fixation by the associated root-nodule Bradyrhizobia.
Introducing Chl d and Chl f into plant leaves will involve two primary steps: biosynthesis of these chlorophylls and their subsequent integration into plant photosystems. The biosynthesis of Chl d and Chl f is considered feasible based on their structural differences from Chl a: Chl d differs from Chl a with a formyl group replacing the ethenyl group at the C3 site and Chl f has a formyl group in place of the methyl group at the C2 site. The enzyme that converts Chl a to Chl f in cyanobacteria, Chl f synthase (ChlF), has been identified as a paralog of the D1 (PsbA) protein of the photosystem II45. Transgenic expression of the ChlF protein in Synechocystis sp. PCC 6803 and Synechococcus sp. PCC 7002 resulted in the synthesis of Chl f45,46. The enzyme for Chl d synthesis is unknown, but it is hypothesized that Chl d can be synthesized from Chl a in a single step utilizing molecular oxygen47,48. These results suggest that bioengineering crops to produce Chl f or Chl d (once the enzyme is known) would only require the expression of a single gene.
However, efficient FR photon utilization in crops requires not only the synthesis of these chlorophylls but also their integration into remodeled photosynthetic complexes. This requires designing pigment binding sites selective for Chl d and f. In this respect, it is promising that experimental studies have shown that the proteins of in vitro assembled plant light-harvesting complexes can bind Chl d and Chl f at specific sites49,50. Fine tuning of the binding sites to enhance their affinity for specific Chls and further shift their absorption into the far-red is underway51, guided by the analysis of the Chl d and f binding sites of far-red photosystems18.
Furthermore, to ensure high light-harvesting and electron transport efficiency, the number and location of the far-red absorbing pigments should be carefully considered18,21. This is particularly important because the harvested energy must be quickly transferred to the reaction center to maximize conversion efficiency. To ensure a high trapping efficiency within the engineered plant photosystems, it may also be necessary to engineer Chl d/f binding sites within the reaction centers of both photosystems, creating red-shifted primary donors analogous to those found in A. marina or FaRLiP cyanobacterial photosystems. Interestingly, PSII from A. marina and FaRLiP cyanobacteria have evolved different strategies to perform charge separation with lower energy photons. A. marina PSII maximizes charge separation efficiency at the expense of increased recombination reactions, while FaRLiP PSII takes the opposite approach. In an engineered plant PSII containing Chl d/f as the primary donor, there is the potential to engineer an approach that is a hybrid of the two aforementioned scenarios. Although this aspect remains to be experimentally verified, the results here now show this would be of great value.
The question that initiated this study was whether the considerable effort needed in discovery and translation to implement longer wavelength chlorophylls in crops would be valuable. The clear answer from this analysis is that it would be immensely valuable. Indeed, if the daunting challenge of providing sufficient food in the second half of this century is to be met, all efforts are needed whether increasing photosynthetic efficiency, improved crop protection, environmental tolerance or agronomy42. Under ideal growing conditions, crop biomass production is determined by the amount of sunlight photons captured and the efficiency with which these are transduced into carbon assimilation52. Incorporating Chl d/f increases the number of utilizable photons for crops. Recent findings have identified many optimization targets to improve leaf photosynthetic conversion efficiency under high light conditions32,52,53, including manipulating CO2 diffusion54, increasing the content or the turnover number of Rubisco3,55,56, engineering photorespiratory bypasses57, and integrating algal, cyanobacterial or C4 CO2 concentration mechanisms into C3 plants58,59. However, in the field, photosynthesis, especially in the middle and lower canopy parts, is often light-limited. Enhancing the absorption and utilization of FR light by leaves can mitigate light limitation of the leaves in the middle and lower canopy. Therefore, combining the changes listed above that increase light-saturated photosynthesis with use of FR light would be synergistic, providing a greater scope for improving canopy photosynthetic efficiency and much needed increases in yield potential, implementable across most crops.
Methods
The photon flux density of FR at earth surface
The distributions of energy density (I; J m−2 s−1 nm−1) of a solar spectrum (ASTM G173-03, https://www.nrel.gov/grid/solar-resource/spectra-am1.5.html, The AM1.5 Global tilt) was used to calculate the photon flux density (Q; mol m−2 s−1) at each wavelength (Fig. 1):
$$Q=frac{Ilambda }{{hc}{N}_{A}}$$
(1)
where λ is wavelength (m); c the speed of light (3.00 × 108 m s−1); NA Avogadro’s number (6.02 × 1023 mol−1); and h Planck’s constant (6.626 ×10−34 J s). The receiving surface is defined in the standards as an inclined plane at 37° tilt toward the equator, facing the sun.
The light distribution in a soybean canopy
Stem positions, leaf lengths, widths, petiole lengths and angles for trifoliate leaves were measured from soybean plants (Glycine max L. Merr., Pioneer 93B15) growing on the University of Illinois South Farms. Plants were sampled at multiple developmental stages during the whole growing season from the day of year (DOY) 168 to 26760. Following standard agronomic practice, the row spacing of the canopy was 38 cm, and plant spacing within the rows 10 cm (Supplementary Fig. 7). From this the 3D architecture of the crop canopy was recreated. The surface of each leaf was divided into ‘pixels’ of approximately 1 cm2 60,61. Photon fluxes for each wavelength range (blue, green, red, and FR) were calculated for each leaf ‘pixel’ at hourly intervals from 06:00 – 18:00 on Aug 18th, 2023, in Champaign IL, US (40.11 N, 88.21 W) using a forward ray-tracing algorithm (FastTracer)62. At every hour, the direct and diffuse light entering and within the canopy is predicted together with scattered radiation due to reflection from, and transmission through, the leaves within the canopy. Leaf reflectance and transmission were specified separately for each wavelength category (blue, green, red, and FR) according to parameters listed in Table 1. The total absorbed photon flux density (Qabs, Eq. (2)) for each leaf ‘pixel’ was computed by summing the absorbed fluxes from all wavelength ranges.
$${Q}_{{abs}}={Q}_{{abs}{{_}}B}+{Q}_{{abs}{{_}}G}{+Q}_{{abs}{{_}}R}+{Q}_{{abs}{{_}}{FR}}$$
(2)
where ({Q}_{{abs_B}},,{Q}_{{abs_G}}{,{Q}}_{{abs_R}},{rm{and}},{Q}_{{abs_FR}}) are absorbed blue, green, red, and far-red light respectively. Atmospheric transmittance was set as 0.80 to estimate the incident direct and diffuse photon fluxes reaching the top of the canopy at each time point on a sunny day.
Leaf ecophysiology
The leaf‐level ecophysiology model63 used in this study couples the physical and biochemical processes that regulate photosynthesis and gas exchange. Specifically, the model simultaneously calculates the boundary layer conductance to vapor and heat, leaf temperature, transpiration (E), net CO2 assimilation (A), intercellular CO2 concentration (Ci), and stomatal conductance (gs) under different environmental conditions. It was implemented across all canopy “pixels” in the 3D model.
A of soybean leaf is described by the steady-state FvCB biochemical model photosynthetic CO2 assimilation64. Here, A is assumed to be constrained by either RuBP carboxylase-oxygenase (Rubisco) activity (({W}_{c})) or by the rate of ribulose-1,5-bisphosphate (RuBP) regeneration (({W}_{j})):
$$A=min left({W}_{c},,{W}_{j}right)left(1-frac{{varGamma }^{*}}{{C}_{i}}right)-{R}_{d}$$
(3)
$${W}_{c}=frac{{V}_{{cmax}}cdot {C}_{i}}{{C}_{i}+{K}_{c}(1+frac{O}{{K}_{o}})}$$
(4)
$${W}_{j}=frac{Jcdot {C}_{i}}{4.5{C}_{i}+10.5{varGamma }^{*}}$$
(5)
where Γ* is the chloroplast CO2 partial pressure when both Rd and CO2 assimilation are equal such that net A = 0, Rd is mitochondrial respiration. Vcmax is the maximum rate of carboxylation, Kc and Ko are the Michaelis-Menten constants of Rubisco for CO2 and O2, respectively, and O is the oxygen concentration. Photosynthesis is more likely limited by ATP availability rather than NADPH (Supplementary Note 1, Supplementary Figs. 8 and 9), hence we use the ATP limited Wj equation (Eq. (5)). The electron transport rate (J) is described by a non-rectangular hyperbolic function65:
$$J=frac{{Q}_{2}+{J}_{max }-sqrt{{left({Q}_{2}+{J}_{max }right)}^{2}-4theta ,{Q}_{2},{J}_{max }}}{2theta }$$
(6)
$${Q}_{2}={0.5Q}_{{abs}}{varPhi }_{{PSII}}$$
(7)
({Q}_{{abs}}) is the amount of absorbed photosynthetically active photon flux available for electron transport (Eq. (2)). ΦPSII, the maximum quantum yield of photosystem II; θ is a curvature factor. 0.5 represent the proportion of ({Q}_{{abs}}) that reaches photosystem II51. Parameters and values are listed in Supplementary Table 4.
Canopy photosynthesis and water use efficiency
Net canopy CO2 assimilation was calculated by summing A values of all leaves ‘pixels’ over the daylight hours (Eq. (8)).
$${Ac}=frac{sum ({A}_{i}cdot {S}_{i})}{{S}_{{ground}}}$$
(8)
where Sground represents the ground area occupied by the simulated canopy (Supplementary Fig. 7). Ai is net leaf CO2 assimilation rate of a leaf pixel (i), and Si is the corresponding leaf area of the leaf pixel. Canopy transpiration (Ec) was similarly calculated by summing transpiration rate of every leaf pixel (Ei) using Eq. (9)
$${Ec}=frac{sum ({E}_{i}cdot {S}_{i})}{{S}_{{ground}}}$$
(9)
Water use efficiency (WUE) of the canopy was calculated as the ratio of Ac to Ec (Eq. (10)):
$${WUE}=frac{{Ac}}{{Ec}}$$
(10)
Simulations were conducted in MATLAB 2019a (Mathworks, https://uk.mathworks.com).
The measurement of reflectance and transmittance spectra of Chlorophyll d
Acaryochloris marina MBIC 11017 was photoautotrophically grown in IMK medium at 25 °C at a constant irradiance of 20 μE m−2 s−1. Chl d was extracted from A. marina cells using 100% ice-cold methanol and incubated in dark at 4 °C for 15 min48, samples were then centrifuged, and the clear supernatant was collected for subsequent measurement. Transmittance spectra of Chl d in methanol were measured using at room temperature on a Cary 4000 UV–VIS spectrophotometer. Room-temperature absorption spectra of A. marina cells were measured on Cary 4000 UV–Vis-spectrophotometer equipped with an integrating diffuse reflectance sphere (DRA-CA-50, Labsphere) to correct for light scattering.
Both Acaryochloris cell suspensions and extracted Chl d in methanol were diluted to various concentrations for measuring their reflectance and transmittance spectra (Supplementary Fig. 1). Reflectance and transmittance values at an Optical Density of 5 (at 350–800 nm) were estimated using linear interpolation (interp1 function in MATLAB).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All reflectance and transmittance spectra data are provided in Supplementary Information file. Source data are provided with this paper.
Code availability
Code that supports the findings of this study are available at GitHub [https://github.com/yuwangcn/Chld_Chlf_in_soybean].
References
-
Van Dijk, M., Morley, T., Rau, M. L. & Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2, 494–501 (2021).
-
Hasegawa, T. et al. Extreme climate events increase risk of global food insecurity and adaptation needs. Nat. Food 2, 587–595 (2021).
-
Yoon, D.-K. et al. Transgenic rice overproducing Rubisco exhibits increased yields with improved nitrogen-use efficiency in an experimental paddy field. Nat. Food 1, 134–139 (2020).
-
De Souza, A. P. et al. Soybean photosynthesis and crop yield are improved by accelerating recovery from photoprotection. Science 377, 851–854 (2022).
-
Nürnberg, D. J. et al. Photochemistry beyond the red limit in chlorophyll f–containing photosystems. Science 360, 1210–1213 (2018).
-
Gan, F. et al. Extensive remodeling of a cyanobacterial photosynthetic apparatus in far-red light. Science 345, 1312–1317 (2014).
-
Gan, F., Shen, G. & Bryant, D. A. Occurrence of far-red light photoacclimation (FaRLiP) in diverse cyanobacteria. Life 5, 4–24 (2014).
-
Kühl, M., Chen, M., Ralph, P. J., Schreiber, U. & Larkum, A. W. A niche for cyanobacteria containing chlorophyll d. Nature 433, 820–820 (2005).
-
Chen, M. & Blankenship, R. E. Expanding the solar spectrum used by photosynthesis. Trends Plant Sci. 16, 427–431 (2011).
-
Antonaru, L. A., Cardona, T., Larkum, A. W. D. & Nürnberg, D. J. Global distribution of a chlorophyll f cyanobacterial marker. ISME J. 14, 2275–2287 (2020).
-
Consoli, G. et al. Locating the missing chlorophylls f in far-red photosystem I. Preprint at https://www.biorxiv.org/content/10.1101/2024.08.06.606606v1 (2024).
-
Mascoli, V., Bhatti, A. F., Bersanini, L., van Amerongen, H. & Croce, R. The antenna of far-red absorbing cyanobacteria increases both absorption and quantum efficiency of Photosystem II. Nat. Commun. 13, 3562 (2022).
-
Mascoli, V., Bersanini, L. & Croce, R. Far-red absorption and light-use efficiency trade-offs in chlorophyll f photosynthesis. Nat. Plants 6, 1044–1053 (2020).
-
Zamzam, N. et al. Femtosecond visible transient absorption spectroscopy of chlorophyll-f-containing photosystem II. Proc. Natl Acad. Sci. 117, 23158–23164 (2020).
-
Viola, S. et al. Impact of energy limitations on function and resilience in long-wavelength Photosystem II. Elife 11, e79890 (2022).
-
Tros, M. et al. Breaking the red limit: efficient trapping of long-wavelength excitations in chlorophyll-f-containing photosystem I. Chem. 7, 155–173 (2021).
-
Ulrich, N. J., Shen, G., Bryant, D. A. & Miller, S. R. Ecological diversification of a cyanobacterium through divergence of its novel chlorophyll d-based light-harvesting system. Curr. Biol. 34, 2972–2979.e4 (2024).
-
Elias, E., Oliver, T. J. & Croce, R. Oxygenic photosynthesis in far-red light: strategies and mechanisms. Ann. Rev. Phys. Chem. 75, 231–256 (2024).
-
Wolf, B. M. & Blankenship, R. E. Far-red light acclimation in diverse oxygenic photosynthetic organisms. Photosynthesis Res. 142, 349–359 (2019).
-
Long, S. P., Marshall-Colon, A. & Zhu, X. G. Meeting the global food demand of the future by engineering crop photosynthesis for yield potential. Cell 161, 56–66 (2015).
-
Srinivasan, V., Kumar, P. & Long, S. P. Decreasing, not increasing, leaf area will raise crop yields under global atmospheric change. Glob. Change Biol. 23, 1626–1635 (2017).
-
McCree, K. J. in Physiological plant ecology I: responses to the physical environment (eds, Lange, O. L., Nobel, P. S., Osmond, C. B., & Ziegler, H.) 41–55 (Springer Berlin Heidelberg, 1981).
-
Shibayama, M. & Akiyama, T. Seasonal visible, near-infrared and mid-infrared spectra of rice canopies in relation to LAI and above-ground dry phytomass. Remote Sens. Environ. 27, 119–127 (1989).
-
Monsi, M. & Saeki, T. über den Lichtfaktor in den Pflanzengesellschaften und seine Bedeutung für die Stoffproduktion. Jpn. J. Bot. 14, 22–52 (1953).
-
Monteith, J. Light distribution and photosynthesis in field crops. Ann. Bot. 29, 17–37 (1965).
-
FAOStat. FAOSTAT, http://faostat3.fao.org/faostat-gateway/go/to/home/E (2023).
-
Song, X.-P. et al. Massive soybean expansion in South America since 2000 and implications for conservation. Nat. Sustain. 4, 784–792 (2021).
-
McCree, K. J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 9, 26 (1971).
-
Savoy, B., Cothren, J. & Shumway, C. Soybean biomass accumulation and leaf area index in early‐season production environments. Agron. J. 84, 956–959 (1992).
-
Kneip, C. et al. Protonation state and structural changes of the tetrapyrrole chromophore during the Pr→ Pfr phototransformation of phytochrome: a resonance Raman spectroscopic study. Biochemistry 38, 15185–15192 (1999).
-
McGuirk, S. L. & Cairns, I. H. Relationships between soil moisture and visible–NIR soil reflectance: a review presenting new analyses and data to fill the gaps. Geotechnics 4, 78–108 (2024).
-
Ort, D. R. et al. Redesigning photosynthesis to sustainably meet global food and bioenergy demand. Proc. Natl Acad. Sci. 112, 8529–8536 (2015).
-
Miyashita, H. et al. Chlorophyll d as a major pigment. Nature 383, 402–402 (1996).
-
Tomo, T. et al. Characterization of highly purified photosystem I complexes from the chlorophyll d-dominated cyanobacterium Acaryochloris marina MBIC 11017. J. Biol. Chem. 283, 18198–18209 (2008).
-
Allakhverdiev, S. I. et al. Redox potential of pheophytin a in photosystem II of two cyanobacteria having the different special pair chlorophylls. Proc. Natl Acad. Sci. 107, 3924–3929 (2010).
-
Ritchie, H. & Roser, M. Crop yields have increased dramatically in recent decades, but crops like maize would have improved more without climate change (Our World in data, 2024).
-
Rockwell, N. C., Su, Y.-S. & Lagarias, J. C. Phytochrome structure and signaling mechanisms. Annu. Rev. Plant Biol. 57, 837–858 (2006).
-
Cheng, M.-C., Kathare, P. K., Paik, I. & Huq, E. Phytochrome signaling networks. Annu. Rev. Plant Biol. 72, 217–244 (2021).
-
Ort, D. R. et al. Redesigning photosynthesis to sustainably meet global food and bioenergy demand. Proc. Natl Acad. Sci. USA 112, 8529–8536 (2015).
-
Ainsworth, E. A. & Long, S. P. 30 years of free‐air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Glob. Change Biol. 27, 27–49 (2021).
-
Long, S. P., Ainsworth, E. A., Leakey, A. D., Nosberger, J. & Ort, D. R. Food for thought: lower-than-expected crop yield stimulation with rising CO2 concentrations. science 312, 1918–1921 (2006).
-
Kimball, B. A. Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations 1. Agron. J. 75, 779–788 (1983).
-
UN-FAO. How to feed the world in 2050, 35 (United Nations Food and Agricultural Organization, 2020).
-
Rogers, A. et al. Increased C availability at elevated carbon dioxide concentration improves N assimilation in a legume. Plant Cell Environ. 29, 1651–1658 (2006).
-
Ho, M.-Y., Shen, G., Canniffe, D. P., Zhao, C. & Bryant, D. A. Light-dependent chlorophyll f synthase is a highly divergent paralog of PsbA of photosystem II. Science 353, aaf9178 (2016).
-
Trinugroho, J. P. et al. Chlorophyll f synthesis by a super-rogue photosystem II complex. Nat. Plants 6, 238–244 (2020).
-
Fukusumi, T. et al. Non-enzymatic conversion of chlorophyll-a into chlorophyll-d in vitro: a model oxidation pathway for chlorophyll-d biosynthesis. FEBS Lett. 586, 2338–2341 (2012).
-
Schliep, M., Crossett, B., Willows, R. D. & Chen, M. 18O labeling of chlorophyll d in Acaryochloris marina reveals that chlorophyll a and molecular oxygen are precursors. J. Biol. Chem. 285, 28450–28456 (2010).
-
Elias, E., Brache, K., Schäfers, J. & Croce, R. Coloring outside the lines: exploiting pigment–protein synergy for far-red absorption in plant light-harvesting complexes. J. Am. Chem. Soc. 146, 3508–3520 (2024).
-
Elias, E., Liguori, N., Saga, Y., Schafers, J. & Croce, R. Harvesting far-red light with plant antenna complexes incorporating chlorophyll d. Biomacromolecules 22, 3313–3322 (2021).
-
Elias, E., Liguori, N. & Croce, R. At the origin of the selectivity of the chlorophyll-binding sites in Light Harvesting Complex II (LHCII). Int. J. Biol. Macromol. 243, 125069 (2023).
-
Zhu, X.-G., Long, S. P. & Ort, D. R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 61, 235–261 (2010).
-
Araus, J. L., Sanchez-Bragado, R. & Vicente, R. Improving crop yield and resilience through optimization of photosynthesis: panacea or pipe dream? J. Exp. Bot. 72, 3936–3955 (2021).
-
Flexas, J. Genetic improvement of leaf photosynthesis and intrinsic water use efficiency in C3 plants: why so much little success? Plant Sci. 251, 155–161 (2016).
-
Lin, M. T., Occhialini, A., Andralojc, P. J., Parry, M. A. & Hanson, M. R. A faster Rubisco with potential to increase photosynthesis in crops. Nature 513, 547–550 (2014).
-
Salesse-Smith, C. E. & Long, W. Y. SP Rubisco as a simple means to enhance photosynthesis and productivity now without lowering nitrogen use efficiency. N. Phytol. 245, 951–965 (2025).
-
South, P. F., Cavanagh, A. P., Lopez‐Calcagno, P. E., Raines, C. A. & Ort, D. R. Optimizing photorespiration for improved crop productivity. J. Integr. Plant Biol. 60, 1217–1230 (2018).
-
Ermakova, M., Danila, F. R., Furbank, R. T. & von Caemmerer, S. On the road to C4 rice: advances and perspectives. Plant J. 101, 940–950 (2020).
-
Hennacy, J. H. & Jonikas, M. C. Prospects for engineering biophysical CO2 concentrating mechanisms into land plants to enhance yields. Annu. Rev. Plant Biol. 71, 461–485 (2020).
-
Song, Q. F., Srinivasan, V., Long, S. P. & Zhu, X. G. Decomposition analysis on soybean productivity increase under elevated CO2 using 3-D canopy model reveals synergestic effects of CO2 and light in photosynthesis. Ann. Bot. 126, 601–614 (2020).
-
Wang, Y., Burgess, S. J., de Becker, E. M. & Long, S. H. P. Photosynthesis in the fleeting shadows: an overlooked opportunity for increasing crop productivity? Plant J. 101, 874–884 (2020).
-
Song, Q. F., Zhang, G. L. & Zhu, X. G. Optimal crop canopy architecture to maximise canopy photosynthetic CO2 uptake under elevated CO2 – a theoretical study using a mechanistic model of canopy photosynthesis. Funct. Plant Biol. 40, 109–124 (2013).
-
Drewry, D. T. et al. Ecohydrological responses of dense canopies to environmental variability: 1. Interplay between vertical structure and photosynthetic pathway. J. Geophys. Res. Biogeosci. 115 G04022 1-25 (2010).
-
Farquhar, G. D., Caemmerer, S. V. & Berry, J. A. A biochemical-model of photosynthetic CO2 assimilation in leaves of C3 species. Planta. 149, 78–90 (1980).
-
Caemmerer, S. V. Biochemical models of leaf photosynthesis (CSIRO Publishing, 2000).
Acknowledgements
We thank Edward Lochocki, Lynn Doran, Elena Pelech, Coralie Salesse-Smith, Cindy Chan and Jeff Hansen for their comments and advice on earlier versions of this manuscript. Award number 1-612289, S.L. is supported by Stanley O. Ikenberry Endowment. Grant number 714.018.001, R.C. and T.O. are supported by the Dutch organization for Scientific research (NWO) via a TOP grant.
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Christine Foyer and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, Y., Oliver, T.J., Croce, R. et al. Addition of longer wavelength absorbing chlorophylls into crops could increase their photosynthetic productivity by 26%. Nat Commun 16, 7933 (2025). https://doi.org/10.1038/s41467-025-62885-6
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41467-025-62885-6