
Introduction
The sustainable intensification of agriculture to meet global food demands while minimising environmental impact is one of the key challenges of the 21st century1. Conventional agricultural practices, which rely heavily on chemical fertilisers and pesticides, often lead to soil degradation, biodiversity loss, and environmental contamination2. These limitations highlight the urgent need for innovative approaches that can enhance crop productivity while preserving ecosystem integrity.
At the same time, space exploration poses additional requirements for autonomous and highly reliable food production, stimulating research in closed-loop life-support systems (Lasseur et al.). Within this context, ESA’s MELiSSA program has developed bioregenerative systems that recycle waste into valuable resources for crop production, including the cultivation of the cyanobacterium Limnospira indica (formerly Spirulina), a nutrient-dense food source rich in proteins, vitamins, antioxidants, and iron3,4,5. To further improve productivity and resilience without external inputs, MELiSSA also explores microbial-based biostimulants, substances or microorganisms that stimulate plant processes to enhance nutrient use efficiency, abiotic stress tolerance, and crop quality3,6,7.
Microbial biostimulants are attracting increasing attention for their ability to modulate plant-associated microbiota and deliver bioactive compounds, thereby improving growth, nutrient uptake, and stress resilience3,6,7. Their activity often relies on reshaping rhizosphere and root-associated communities, which play essential roles in plant health through processes such as nitrogen fixation, phosphate solubilisation, and the synthesis of phytohormones, including auxins and gibberellins8,9. Recent work has shown that biostimulant-induced modulation of these microbial networks can substantially enhance crop performance under both optimal and suboptimal conditions10,11,12.
Delivery route is a critical determinant of biostimulant efficacy13. Soil amendment introduces compounds into the root zone, where they interact with the rhizosphere microbiota, influence nutrient cycling, and modulate root colonisation14. In contrast, foliar application delivers compounds directly to the leaf surface, allowing rapid uptake and signalling, with potential downstream effects on root physiology and exudation15. However, systematic comparisons between these routes remain scarce, especially under controlled dose-equivalent conditions.
Here, we investigate the effects of L. indica-derived fractions on Solanum lycopersicum (tomato), a major crop valued for its nutritional quality and widely used as a model in biostimulant research16,17,18. Tomato was chosen because polysaccharides such as chitosan have already been shown to exert biostimulant effects19,20,21,22, and because its well-characterised, highly dynamic microbiome is particularly sensitive to soil, developmental stage, and external inputs, making it an ideal system to assess biostimulant-driven shifts in plant performance and root-associated communities23,24,25,26.
Although cyanobacteria-based biostimulants are increasingly studied due to their bioactive metabolites such as polysaccharides27, there remains a lack of mechanistic evidence linking Limnospira-derived fractions to issues of different culture conditions to plant phenotypes, microbiome dynamics (rhizosphere versus root), and solution chemistry under different application routes28,29,30,31. Importantly, culture conditions have been shown to markedly affect the biochemical composition of L. indica, particularly with respect to polysaccharide biosynthesis. For example, nitrate limitation induces a pronounced metabolic shift, leading to enhanced accumulation of polysaccharides, including extracellular exopolysaccharides (EPS)3.
In this study, we address these gaps by (i) comparing two L. indica-derived fractions, culture media (CM) and biomass extracts (BME), produced under nitrate-sufficient (N+) and nitrate-deficient (N−) conditions; (ii) applying dose-equivalent treatments via soil amendment and foliar application; (iii) monitoring tomato growth traits (height, branching, leaf, flower, and fruit number) over time; and (iv) characterizing treatment- and time-dependent shifts in rhizosphere and root-associated microbiota using 16S rRNA gene sequencing, and their associations with plant performance.
Results
We conducted a comprehensive characterisation of the different extract solutions used for biostimulation experiments in order to understand their composition and properties, which can influence their effectiveness in promoting plant growth through direct intake of bioactive components from L. indica extracts or through microbiome impact. By analysing factors such as growth curves of L. indica, carbohydrate and protein content, as well as polysaccharide composition, we can assess the qualities of the biostimulant extracts, their impact on plant characteristics and the associated microbiome impact.
L. indica culture fraction characterisation
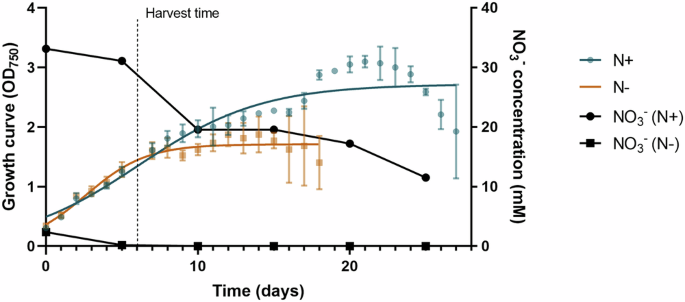
Limnospira indica growth curve in the presence or absence of nitrate and associated carbohydrates and proteins content was evaluated.
L. indica presented the ability to grow in the presence and absence of nitrate in its environment (Fig. 1). However, we observed a higher optical density, revealing a more effective growth, when the environment was in the presence of nitrate, and, therefore, optimal. Indeed, while the end of the exponential phase is reached at 20 days with an OD750 = 3 in N+ condition, it is reached at 9 days with an OD750 = 2 only in N− condition. Remaining ({{rm{NO}}}_{3}^{-}) in N− condition is observed between days 0 and 5, corresponding to the remaining nitrate from the mother cultures.

Nitrate concentration in both CM is represented in black (N+ = black circle, N− = black square).
In addition, the total carbohydrate and protein content was assessed to evaluate the impact and overall composition of L. indica when grown in the presence (N+) and absence (N−) of nitrate at the end of the exponential growth, with respective OD750 (N+) = 1.8 and OD750 (N−) = 1.2 (Fig. 2).

A Protein content in percentage of biomass dried weight (% of DW). B Total carbohydrate content within the biomass (capsular and internal polysaccharides) in percentage of dried weight of biomass (% DW). C Total released carbohydrate content present in CM (released polysaccharides), relative to the dried weight of biomass. The values are shown as mean ± SEM. Statistical significance was tested using a t-test. n = 3. DW dry weight, CPS capsular polysaccharides, RPS released polysaccharides. *p-value < 0.05, **p-value < 0.01, ***p-value < 0.001.
The protein content of L. indica when grown in the presence of nitrate (N+) was 76% of DW (Fig. 2A), while capsular and internal carbohydrates observed were 8% of DW (Fig. 2B). In parallel, when grown in the absence of nitrate N−, protein content of L. indica dropped to 31% of DW (Fig. 2A), and the capsular and internal carbohydrate content increased to 57% of DW (Fig. 2B). Last, the amount of excreted polysaccharides is increased by a factor of 3 when grown in the absence of nitrate from 0.05% of DW in N+ to 0.15% DW in N− (Fig. 2C).
Numerous studies have investigated the role of nitrogen in the growth of L. indica32,33,34,35,36. One notable aspect of L. indica’s biology is its requirement for nitrogen in its environment, which has been shown to have a fundamental impact on its growth37. Indeed, it has been demonstrated that the absence of nitrate in the CM can significantly impair the growth of L. indica32,36. This is likely due to the crucial role that nitrogen plays in the metabolic processes and cellular development of cyanobacteria38. Removing the combined nitrogen sources in non-diazotrophic cyanobacteria has an impact on the culture growth38. Moreover, in nitrogen-deprived conditions, photosynthesis persists, leading to the degradation of phycocyanin as a nitrogen source39. The reduction in chlorophyll, phycocyanin and phycobilisome (PBS) content causes a noticeable shift in cell pigmentation, transitioning from the typical blue-green to a yellow-green hue, a process referred to as bleaching or chlorosis39,40. Concurrently, carbon is accumulated in the form of both extracellular and intracellular polysaccharides, with glycogen serving as the primary intracellular storage compound41,42,43,44. This accumulation reaches a level where cellular sedimentation occurs due to increased biomass density. Additionally, it has been observed that nitrogen stress can lead to the modulation of secondary metabolite production in L. indica, which in turn can affect its overall growth and development45,46,47.
The importance of nitrogen for the growth of L. indica is further underscored by the fact that it is an essential nutrient for a wide range of species, including plants used in agricultural and horticultural applications3,48. Nitrogen is a crucial component of various biomolecules, such as proteins, nucleic acids, and chlorophyll, all of which play vital roles in the growth and development of photosynthetic organisms49.
The composition of internal and excreted polysaccharides was further evaluated to better characterise their monosaccharide profiles under nitrate-sufficient and nitrate-deficient conditions.
To evaluate the composition of polysaccharides synthesised by L. indica, the monosaccharide profiles of both intracellular (biomass-associated, capsular and internal) and extracellular (released into the culture medium) fractions were analysed. The relative abundances of monosaccharides were determined after acid hydrolysis and analysis through mass spectrometry (Fig. 3).

Left, internal (intracellular and capsular) polysaccharides within Limnospira indica‘s biomass (BME N+ and BME N−) and right, extracellular (excreted) polysaccharides within Limnospira indica‘s CM (CM N+ and CM N−). n = 5, mean of replicates.
Overall, the intracellular polysaccharides were dominated by Glucose and Galactose, which constituted the majority of the carbohydrate fraction within the biomass of N+ and N− conditions, with respectively 58% and 97% of detected monosaccharides. Other monosaccharides, including arabinose+xylose, fructose+rhamnose, ribose and mannose, were detected and mainly present in the BME N+ condition. Within the BME, N+ arabinose+xylose were present at 7%, fucose+rhamnose at 23%, ribose at 10% and mannose at 6%. In parallel, BME N− was composed of arabinose+xylose and fucose+rhamnose at respectively 1%, 2%. Ribose and Mannose were not detected.
The extracellular polysaccharides exhibited a distinct composition. Glucose and galactose remain the most observed monosaccharides in both N+ and N− conditions, with 55% and 29% of detected monosaccharides. Fucose+rhamnose were more represented compared to their proportion in the biomass, with 42% and 56% in N+ and N−, respectively. Arabinose+xylose and ribose were less abundant in CM N+ present at 3% and 0% respectively, compared to the biomass (N+), while they were more abundant in CM N− present at 13% and 1% respectively, compared to the biomass (N−).
The monosaccharide composition observed in nitrate-depleted biomass by ref. 50 was also primarily composed of glucose and galactose in the control (N+) (52%) or by ref. 51 with 60%. However,50 reported a higher concentration of rhamnose (30%) against only 9% by51. In contrast, under nitrate starvation, glucose remained the predominant monosaccharide in both studies (83% for ref. 50 and 94% for ref. 51), aligning with our results. Similarly,52 found that glucose was the only monosaccharide detected in nitrate-starved Arthrospira biomass.
As a result, polysaccharides from internal and excreted polysaccharides were mainly composed of glucose and galactose. Other detected monosaccharides showed a more balanced distribution. These findings indicate that L. indica modulates its extracellular polysaccharide composition, in response to environmental factors, leading to metabolic shifts due to the C:N ratio in a nitrate-deficient environment, as observed in several cyanobacterial strains53. Nitrate is retrieved from photosynthesis-linked proteins (phycobilisom, phycocyanin, chlorophyll) and carbon is accumulated in the form of both extracellular and intracellular polysaccharides40,42,44. Therefore, a decrease in nitrogen availability alters the C:N ratio, resulting in an excess of carbon. This excess carbon is subsequently utilised in the synthesis of polymers, including EPS. Previous studies have shown that EPS production is particularly enhanced when the C:N ratio reaches 10:153,54.
Biostimulation of S. lycopersicum
Different parameters were evaluated in order to characterise plant growth and biostimulant solutions. These parameters included the total height of the plant (Fig. 4), the number of ramifications (Supplementary Fig. 1), leaves (Supplementary Fig. 1), flowers (Fig. 5A, B) and early fruits (Fig. 5C, D). Importantly, the suckers on the plants were not removed during the observation period. Furthermore, the count of suckers was found not to be significantly different across the plants, indicating a uniform pattern of suckers development and direct impact on other parameters.

Evolution under amendment (A, C) and foliar (B, D) biostimulant applications over 35 days (A, B) and at day 35 (C, D). The values are shown as mean ± SEM. Statistical significance was tested using a t-test, n = 5. CM culture media biostimulant, BME biomass extract biostimulant, NC negative control.

Under amendment (A, C) and foliar (B, D) biostimulant application after 35 days. The values are shown as mean ± SEM. Statistical significance was tested using a t-test, n = 5. CM culture media, BME biomass extract, NC negative control.
Regarding the plant’s size (Fig. 4), when the first 20 days, does no clear impact of the biostimulant extracts used on the plant, the observations made at day 35 show significant differences between the conditions tested. Solutions prepared from L. indica biomass grown in the absence of nitrates (BME N−) increased plant height relative to water control when used as watering solution (amendment) compared to the water control (Fig. 4C), while CM from L. indica grown in the absence of nitrates (CM N−) induced a diminution of S. lycopersicum size. In parallel, CM of L. indica grown in the presence of nitrates (CM N+) appeared to significantly increase the plant size when applied directly on the leaves compared to the water control, while L. indica biomass grown in the presence of nitrates (BME N+) induced an inhibition of the plant size.
When looking at the ramifications and the leaves (Supplementary Fig. 1A) of S. lycopersicum (Supplementary Fig. 1C), it is observed that the amendment administration does not impact the number of ramifications nor leaves with any of the biostimulant solutions tested. However, when applied through the foliar application process (Supplementary Fig. 1B, D), BME of L. indica grown in the presence (BME N+) and absence of nitrates (BME N−) was shown to inhibit the number of ramifications observed and therefore the leaves compared to other biostimulant solutions.
In parallel, the number of flowers has been observed after 35 days of growth and revealed an interesting effect of biostimulant on S. lycopersicum (Fig. 5). Indeed, while applied by amendment, CM of L. indica grown in absence of nitrate (CM N−) has a significant impact on S. lycopersicum flowers production (Fig. 5A) compared to the other biostimulant solutions including water control. However, foliar application of biostimulant solutions did not present any difference in terms of flower production amongst all conditions tested (Fig. 5B). Last, the number of fruits has been observed after 35 days of growth and revealed an interesting effect of biostimulant on S. lycopersicum (Fig. 5). Indeed, when applied through amendment application, early fruit production was observed only with CM N− biostimulation (Fig. 5C). In addition, when applied through foliar application, early fruits were observed in 3 biostimulant conditions: CM N−, BME N+, and BME N−, where only N− solutions presented a significant increase in fruit early phenology (Fig. 5D).
Based on these results, it appears that the application of L. indica presents a great interest for biostimulation. More precisely, CM N− biostimulant presents interesting results both in amendment and foliar application. The flowering stage of the plant seems to be particularly impacted by the application of CM N− and BME N− biostimulants, where the amendment application of CM N− significantly increased the number of flowers produced by S. lycopersicum. Following the flowering stage, the early reproductive phenology (fruits) seems to be increased by both applications, with a maximal impact of BME N− applied on the leaves.
Biostimulant impact on rhizosphere and endosphere microbiota over time
Samples analysed in targeted genomics were samples extracted from the growth plants experiment at T0 (0 days), T20 (20 days) and T35 (35 days) of biostimulant application (see section “Biostimulation of Solanum lycopersicum”).
The NMDS profile of the pairwise community dissimilarity (Supplementary Fig. 2) demonstrated clustering patterns for each experiment based on the batch in which the experiment was conducted. Control groups were not consistently associated, indicating dissimilarities among them, and thereby not allowing comparison across the three experiments. Consequently, the results are structured across all eight datasets (Table 3): CMRhizoA, CMRootsA, CMRhizoF, CMRootsF, BMRhizoA, BMRootsA, BMRhizoF, and BMRootsF.
Although this dataset structure is not without limitations, it aligns with the study’s focus on evaluating the differential impact of treatments between water control and culture-derived fraction of L. indica from N+ and N− cultures (from both CM and BME). This structure facilitates testing the hypothesis: Does the biostimulant produced from nitrate-deficient (N−) Limnospira indica culture significantly impact the microbiota, and therefore explain the phenotypic results observed in section “Biostimulation of Solanum lycopersicum”?
Based on the phenotypic results obtained in S. lycopersicum seedlings growth experiments (see section “Biostimulation of Solanum lycopersicum”), the following analysis is mainly focused on datasets of CM analysis in the presence (N+) and absence (N−) of nitrate and further referred to as CMRhizoA, CMRootsA, CMRhizoF, and CMRootsF. Indeed, the primary interest in biostimulants lies in their impact on flower and fruit production rather than on plant size. Datasets of BME analysis are presented in supplementary data for clarity, yet are discussed in the following sections.
The PERMANOVA analysis significantly highlighted the influence of both time and conditions for CM and BME treated samples (conditions × time) on the bacterial community structure in all experiments conducted, within the rhizosphere and endosphere microbiota (Supplementary Table 1). When accounting for inter-batch variation using the strata argument, significant differences were also observed between biostimulant conditions (Supplementary Table 2). In amendment applications, CM N− differed significantly from the negative control in both rhizosphere and endosphere microbiota (Supplementary Table 3). In contrast, for foliar applications, no differences were detected between conditions in both the rhizosphere and root microbiota. The dissimilarity observed among control samples indicates that a degree of inter-batch variation is inherent to our system. While such variability may reduce the robustness of statistical inference and complicate the generalisation of treatment effects, the use of batch-constrained PERMANOVA helps mitigate this influence by restricting permutations within each batch. Importantly, the consistent detection of treatment-related differences across independent batches suggests that the observed effects are not artefacts of intra-batch variability but reflect reproducible biological responses. The α-diversity of the bacterial community of treated samples with biostimulant was measured via the Shannon index in the different conditions (Supplementary Fig. 3). All statistical analyses (p-values) are presented in Supplementary data (Supplementary Table 4). The analysis revealed that a CM N− amendment application induced a significant diminution of the α-diversity of the bacterial community in the rhizosphere microbiota at T20 compared to the water control (either at the same time and at T0). No difference was observed amongst all 3 conditions at T35, but a similar significant increase between T20 and T35 is noted in water control, CM N+ and CM N− conditions. In parallel, the α-diversity of the bacterial community in endosphere microbiota, revealed a clear significant increase over time for all the conditions tested. When applied through foliar application, the rhizosphere microbiota presented a significant increase from T0 to T35 in both CM N+ and CM N− conditions. In the endospheric microbiota, we observed a significant increase over time (T0–T20–T35) in all conditions tested. At T35, the α-diversity is higher in CM N− compared to water control and CM N+. Similar results are observed in samples biostimulated with BME (N+ and N−) through both applications. In addition, the endosphere microbiota in foliar application of BME N+ and BME N− presented an α-diversity higher at T35 BME N+ and BME N−.
Previous studies have shown a similar evolution of the α-diversity over time. Indeed, potato plants have shown a rhizosphere microbiota with a higher community diversity during the flowering stage, while the seedling and early stage showed a lower diversity55,56,57. Nevertheless, while the bacterial community has an impact on plant growth58,59, plants’ released metabolites also affect the microbial community both positively and negatively, depending on the development stage of the plant and on the metabolites produced. Saccharides positively impact microbiome, while antimicrobial compounds have an expected negative impact on the microbial diversity60. Regarding foliar application, it seems obvious that the microbial community evolution results from the production of the plant.
Alongside α-diversity, the distinctness of the microbial communities highlights the impact of the biostimulant on the said communities. Depending on the biostimulant used and application processes, the distinctness revealed the apparition of unique genera as shown on the Venn diagrams (Supplementary Fig. 4) and resumed in Table 1. At T35, most of the observed genera are shared in all conditions tested (from 27% to 65%) apart from CMRhizoF, where only 18% correspond to shared genera from all conditions. Nevertheless, regarding unique genera observed in each condition, more unique genera are observed in samples biostimulated with N− solutions except CMRhizoA and BMRootsA in both treatments. Similar results are observed when BME is used as a biostimulant solution.
The application of microbial-based biostimulants has been shown to increase the abundance and diversity of bacteria involved in nutrient cycling, such as nitrogen-fixing bacteria and phosphate-solubilising bacteria61. This microbial distinctiveness likely contributes to the phenotypic variations observed in S. lycopersicum growth, such as early reproductive phenology seen in see section “Biostimulation of Solanum lycopersicum”.
Furthermore, the composition of microbiota phyla after 20 and 35 days of biostimulation, has been analysed and is detailed in supplementary data (Supplementary Figs. 5 and 6). After a biostimulation with all conditions, the endospheric microbiota was dominated by four major taxa: α-, β-, γ–proteobacteria, and Bacteroidota. In the rhizosphere microbiota, the same four taxa were identified, in addition to Firmicutes and Acidobacteria. No bacterial group was exclusive to any particular condition. Overall, α–proteobacteria and β–proteobacteria predominantly characterised the endosphere microbiota, while α–proteobacteria and γ–proteobacteria were more abundant in the rhizosphere. These patterns highlight shifts in dominant phyla and genera under biostimulation. Nevertheless, while these taxa are associated with plant growth-promoting functions in the literature, their roles in our system cannot be directly inferred from taxonomic enrichment alone, and such functional interpretations should be considered hypothesis-generating pending direct experimental validation. Indeed, this taxon combination has already been observed in Arabidopsis thaliana and potato plants55,62 and has already been described as impacting plant growth. Alphaproteobacteria are known to include plant growth-promoting bacteria (PGPB), with several genera recognised as symbionts in legumes63,64. Betaproteobacteria play a key role in the nitrogen cycle, facilitating plant-available nitrogen forms while limiting CO2 fixation. They promote plant growth by fixing nitrogen in various plant species and enable the oxidation of ammonium to nitrite and nitrate, an essential reaction for plant development65. Gammaproteobacteria also contribute to plant growth by producing compounds such as auxins, gibberellins, cytokinins, and ethylene, as well as having an impact on nitrogen fixation and mineral solubilisation10,63,66. Lastly, Bacteroidota, identified as plant growth-promoting rhizobacteria (PGPR), demonstrates an ability to colonise plant roots successfully. Although their specific effects on plants are yet to be better described, they are known to significantly colonise during the early plant growth stage67,68. Further metagenomic analysis or assays (e.g. nifH qPCR, enzyme activities) would be required to directly validate the functional roles of these taxa in our system.
The 10 most represented genera of each CM biostimulated condition are displayed in the Circos graphs (Fig. 6), where relationships between different genera and treatment conditions are shown over time.

Circos graphs139 representing the relative abundance of rhizosphere microbiota (A, C) and endosphere microbiota (B, D) at T0, T20 and T35 following the application of L. indica CM-based biostimulant via amendment (A, B) and foliar (C, D) treatment. nper treatment = 5, nper plant = 5.
In both the CMRhizoA and CMRhizoF experiments, the four predominant genera are Rhodanobacterium, Devosia, Chitinophaga, and Dokdonella. These four key genera in the biostimulation profiles have well-known plant growth-promoting properties. Rhodanobacter was prominent across both amendment and foliar treatments, with its relative abundance comparable between amendment and foliar treatments. This genus has been recognised for its plant growth-promoting properties, particularly as an endophytic bacterium in S. lycopersicum, where it enhances biotic and abiotic stress tolerance69. Additionally, Rhodanobacter is a nitrogen-fixing bacterium70 that produces indoleacetic acid (IAA), which stimulates root growth, thereby facilitating nitrogen uptake from the soil70,71. This ability to promote nitrogen and carbon recycling within the soil ecosystem further underscores its ecological significance and potential applications in sustainable agriculture72. Our study revealed a similar relative abundance of 18 ± 1% of Rhodanobacter at T0 and T20 in all conditions except when CM N− was applied through foliar application, where the relative abundance dropped to 13%. Furthermore, Rhodanobacter relative abundance decreased at 13 ± 2% at T35 except when CM N− in both applications, where it dropped to 10%. Its abundance, mainly in the first days of our experiment, revealed its beneficial impact on the early plant growth through plant nitrogen fixation and stress tolerance. Its decline by T35, particularly under CM N− conditions, reflects its temporal appearance within the microbial community.
Similarly, Devosia has shown an equivalent abundance across biostimulant treatments, suggesting its involvement in root development and nitrogen cycling throughout the experiment. Known for its nitrogen-fixing capabilities70, Devosia contributes to plant growth by possibly producing phytohormones that enhance root development. This genus has been linked with increased plant growth, improved mycorrhization, and nitrogen accumulation73, suggesting its promising role in enhancing crop productivity74. Our study presented an initial increase of Devosia relative abundance from T0 (2% in CMRhizoA and 1% in CMRhizoF) to T20 (5 ± 1%), displaying stable abundance from T20 onwards across treatment conditions.
Chitinophaga exhibited a significant decline from T0 to T35. The sharp decline over time of some genera, such as Chitinophaga, could reveal a competitive interaction within the rhizosphere, as previously linked to reduced bacterial α-diversity. Previous studies have demonstrated that the presence of Ralstonia solanacearum in the rhizosphere can reduce the α-diversity of the bacterial community, with genera like Bacillus and Chitinophaga being negatively impacted58. This decline in Chitinophaga abundance under certain conditions may thus be indicative of competitive or antagonistic interactions within the rhizosphere microbial community. Our studies highlighted a relative abundance of Chitinophaga of 6% in CMRhizoA and 15% in CMRhizoF, decreasing drastically to 4 ± 1% at T20, and 3% at T35 in all conditions of both experiments.
Lastly, Dokdonella displayed slightly different results between foliar and amendment application, with increased abundance over time in specific conditions, pointing to its potential sensitivity to biostimulants. Dokdonella species are aerobic rods isolated from soil, and certain species within this genus can perform aerobic nitrate reduction, which may play a role in soil nutrient cycling75. Our study has exhibited an increase in relative abundance from T0 (2%) to T20 (4%), followed by stable relative abundance through T35 in amendment application. When the biostimulants were applied through foliar application, the water control revealed an increase from T0 (3%) to T35 (8%). This increase was also observed in CM N+ with a maximum of 6%. CM N− condition relative abundance appeared to increase as others at T20 (6%), but a slight decrease to T35 (4%) was observed.
Overall, the presence and behaviour of these bacterial genera in the biostimulation context reveal their roles in nutrient cycling, stress tolerance, and plant growth promotion. The temporal evolution of the relative abundance of bacterial genera in this study highlights dynamic shifts in microbial community composition under biostimulation conditions. This study highlights the dynamic behaviour of key plant growth-promoting bacterial genera (Rhodanobacter, Devosia, Chitinophaga, and Dokdonella) under biostimulation conditions with amendment and foliar treatments over time. The observed microbial dynamics provide insights into how biostimulants can shape the rhizosphere microbiome and optimise conditions for sustainable agricultural practices.
In both the CMRootsA and CMRootsF experiments, the five predominant genera of interest are Massilia, Rhodanobacter, Acidovorax, Shinella, and Asticcacaulis. Similar to observations in the rhizosphere, these genera are recognised for their plant growth-promoting properties and are frequently highlighted in biostimulation studies for their beneficial effects.
The relative abundance evolution over time of Massilia appears similar between amendment and foliar treatments. This genus is associated with plant growth promotion, particularly in stressful environments. Massilia has been shown to enhance plant resilience to salinity stress, helping with nutrient accumulation and root colonisation in corn plants76. Additionally, Massilia has been linked to the promotion of lateral root growth and biomass accumulation, especially in flowering plants like maize, where it associates with root functional genes involved in reproduction77. Our study revealed a marked decrease in Massilia relative abundance over time, starting with a high relative abundance of 32 ± 2% at T0, decreasing to 13 ± 2% at T20, until 3 ± 1% at T35 in both application methods. Therefore, considering previous studies on this genus, its presence could suggest a shift in the plant growth stage with the arrival of the flowering stage. In addition, the presence of Massilia in the microbial community may also be beneficial in supporting early plant growth, as suggested by co-occurrence with protists that positively impact the bacterial community structure and nitrogen uptake78,79,80 suggesting its importance at the early growth stage.
Rhodanobacter, a well-known plant growth promoter69,70 exhibited a higher relative abundance in amendment treatments compared to foliar treatments, with a similar temporal pattern in both. Rhodanobacter has shown a high initial relative abundance at T0 (15% in CMRootsA and 9% in CMRootsF) and decreased strongly at T20 (7% in CMRootsA and 0.4% in CMRootsF). At T35, Rhodanobacter relative abundance was in both treatments at 3 ± 1%.
Acidovorax displayed a distinct response depending on the biostimulant application method. While this genus is traditionally known as a biotrophic pathogen, several Acidovorax strains are commensal or beneficial, producing hormones and secondary metabolites that promote plant growth and help to compete with pathogens81,82. In the amendment experiment (CMRootsA), Acidovorax relative abundance remained stable from T0 to T20 (5 ± 1%), whereas it increased with the application of CM N+ and CM N− biostimulants at T35, reaching respectively 9% and 7%. On the contrary, in the foliar treatment, Acidovorax abundance decreased over time, starting from 11% decreasing to 7 ± 1% in water control and CM N+. Especially low levels have been observed at T20 (0.3%) and T35 (1%) in the CM N− condition relative to other treatments.
As observed with Acidovorax, the relative abundance of Shinella presented a distinct response depending on the biostimulant application method. In the amendment experiment with water control, Shinella abundance decreased steadily from T20 (6%) to T35 (1%), while it showed a slight increase at T20 (9%) followed by a decrease at T35 (6%) in the CM N+ and CM N− conditions. A similar pattern was observed in the foliar treatment with all biostimulant conditions, with a relative abundance of 1% at T0, increasing to 7 ± 2% at T20 and finally decreasing to 3 ± 1%. Known for its production of phytohormones, Shinella can positively influence plant growth and development by enhancing root formation and seed germination83,84. In particular, strains of Shinella have been shown to mitigate microcystin (MC) accumulation in plants, reducing toxin levels in roots and eliminating it from leaves and fruits, which suggests potential applications for enhancing plant safety in contaminated environments85. In our study, this behaviour could derive from the competitiveness of microbial communities exacerbated by the biostimulation treatments.
Finally, Asticcacaulis’s relative abundance showed distinctive behaviour based on the biostimulant applied, but presented similar results when applied to the soil or leaves. This genus has been identified as a plant growth-promoting bacterium in crops like sugar beet, where it enhances seedling growth and supports biofilm formation that can protect plants against environmental stress and diseases86,87. Indeed, Asticcacaulis showed an increase in abundance in the control from T0 (2%) to T35 (5.5 ± 1.5%) but a decrease from T20 (6%) to T35 (3%) in the CM N+A, CM N+F and CM N−A conditions. Interestingly, CM N−F biostimulants made the relative abundance of Asticcacaulis stable at 3% at T20 and T35. The higher abundance of Asticcacaulis in amendment-treated roots may suggest that this application method is more valuable for supporting beneficial biofilms in the rhizosphere.
The results provide a detailed characterisation of the genera identified within the rhizosphere and endosphere microbiota following biostimulation with L. indica CM as a biostimulant. These findings indicate that biostimulation with CM and its excreted metabolites, mainly polysaccharides, may favour specific genera in the plant microbiota contributing to enhanced plant growth. These genera have also been observed in the microbiota of plants biostimulated with BME from L. indica (Supplementary Fig. 7). A similar distribution and evolution of these genera has been similar in rhizosphere microbiota. However, the root microbiota appeared to differ from the results observed with the CM biostimulant. Massalia was the most impacted genus with an important decrease of the relative abundance from T0 to T35 (23% to 11% in amendment and 41% to 26% in foliar application) with water control, and a significant increase to 45% in CM N+ when applied on the leaves and CM N− when applied on the soil at T35. These results underscore the significance of genus-specific abundance in the plant growth promotion process, genera that are overly abundant or, on the contrary, less prevalent may diminish the beneficial impact on plant growth promotion.
To further delve into biostimulation impact on genera in the microbiota of both rhizosphere and roots, heatmaps were realised for each experiment (Figs. 7 and 8: RhizoA, RootsA, RhizoF, RootsF). These heatmaps displayed respectively 95 genera in five RGs, 63 genera in four RGs, 43 genera in four RGs and 52 genera in five RGs statistically significantly affected, identified using nbGLM analysis.

Rhizosphere (A) and endosphere (B) microbiome evolution following the application of L. indica CM-based biostimulant via amendment treatment. Colours represent normalised abundance values. RG response group. nper treatment = 5, nper plant = 5.

Rhizosphere (A) and endosphere (B) microbiome evolution following the application of L. indica CM-based biostimulant via foliar treatment. Colours represent normalised abundance values. RG response group. nper treatment = 5, nper plant = 5.
In the CMRhizoA experiment (Fig. 7A), RG1 comprise genera predominantly found at T0 with an increased prevalence at T20 under CM N+ and CM N− conditions. RG2 included genera primarily present at T20 in CM N+ and CM N−. RG3 included genera found consistently in controls (all time points), as well as in CM N+ and CM N− at T35. Within RG4, the most representative genera were observed across all conditions at T35. Finally, RG5 was characterised by genera present under all conditions at T35 but more predominantly in the CM N− condition.
In the CMRhizoF experiment (Fig. 8A), RG1 comprise genera predominantly found in all conditions at T20 and T35. RG2 comprises genera present at T35 in all conditions and slightly overrepresented in CM N−. RG3 included genera found at T20 in all conditions. Finally, in RG4, the most representative genera were observed at T0.
In the CMRootsA experiment (Fig. 7B), RG1 comprises genera predominantly found in the control at T35. RG2 is composed of genera present at T35 in all conditions and overrepresented in CM N−. RG3 included genera found massively at T0. Finally, in RG4, the most representative genera were observed across all time points in CM N+ and CM N−.
In the CMRootsF experiment (Fig. 8B), RG1 comprises genera predominantly found in all conditions at T20 and T35. RG2 is composed of genera present at T35 in water control and CM N− conditions. RG3 included genera found only at T0. RG4 included a single genus, Bosea, found in CM N+ condition at T35. Finally, in RG5, the most representative genera were massively observed at T35 in the CM N− condition.
Our analysis revealed that when CM from L. indica is used as a biostimulant, several genera are prevalent compared to the water control groups over time. Overly expressed genes appeared in biostimulated conditions with CM N− biostimulant at T35. Amongst them genus were particularly highlighted Acidovorax, Aeromicrobium, Agrobacterium, Aliidongia, Anaerocolumna, Anaerotaenia, Azospirillum, Bauldia, Brevundimonas, Caldicoprobacter, Castellaniella, Chryseobacterium, Citrifermentans, Dechlorobacter, Desulfitobacterium, Devosia, Empedobacter, Enterobacter, Ferrovibrio, Ferruginibacter, Ferruginibacter, Fimbriiglobus, Fimbriimonas, Flavipsychrobacter, Flavobacterium, Flectobacillus, Fluviicola, Gellertiella, Geomonas, Geotalea, Hydrocarboniphaga, Lacisediminimonas, Lelliottia, Luteimonas, Lysobacter, Methylobacterium, Methylomicrobium, Nemorincola, Niabella, Nitrobacter, Paludibaculum, Paracoccus, Patulibacter, Pedobacter, Pedomicrobium, Prolinoborus, Pseudescherichia, Pseudoduganella, Pseudoxanthomonas, Reyranella, Rurimicrobium, Sphingopyxis, Stenotrophobacter, Stenotrophomonas, Tepidisphaera, Terrimonas, Tsuneonella, Tuberibacillus, Urbifossiella, Variovorax, and Xenophilus. Most of these genera have been observed in tomato rhizosphere or endosphere microbiota in previous studies10,88. Additionally, a large proportion of these genera were also characterised as impacting during tomato flowering, which corresponds to our T3510. Genus revealed by the heatmaps at T35 in CM N− conditions (both applications) was statistically different in endosphere microbiota compared to the rhizosphere microbiota (Fig. 7, B and Fig. 8, B). More importantly, RGs highly represented in this analysis corresponded to genera present in N− conditions. These genera are represented in Fig. 9.

CM culture media biostimulant, N− nitrate-deficient culture, N+ nitrate-sufficient culture.
A clear increase of Pseudoduganella and Afipia is observed in the case of a foliar application of CM N− (Fig. 9). These gram-negative bacteria are known for their potential as plant growth promoters71,89. In the meantime, Enterobacter, a known plant growth promoter90,91 presented a clear overabundance in the CM N− condition in foliar application compared to the CM N+ biostimulant and water control with the same application, where it is not represented at all. Nevertheless, Entorobacter was observed in amendment application (all conditions), but with a minimal abundance compared to the CM N− condition applied on the leaves. Last, Variovorax has been observed in the majority of CM N+ conditions with foliar application. Its abundance was reduced in the CM N− condition with foliar application, and was even less present in all conditions represented in Fig. 9.
Amongst the genera observed in Fig. 9, the most significantly abundant genera observed in CM N− with amendment or foliar treatment are represented in Table 2 with their known impact on plant growth.
The study demonstrates that biostimulants derived from L. indica CM significantly influence the rhizosphere and endosphere microbiota, particularly under CM N− conditions at T35. Key genera have been highlighted and prove their roles in promoting plant growth and stress resilience. Biostimulation brings up distinct microbial patterns, with several genera previously associated with S. lycopersicum microbiota and flowering stages. These results underscore the potential of L. indica-based biostimulants to enhance crop productivity by modulating beneficial microbial communities.
Relation between phenotypic observation and genomic results on biostimulant impact
The integration of phenotypic and taxonomic results highlights a clear link between biostimulant-induced microbiota modulation and plant growth responses in S. lycopersicum. The observed phenotypic differences in plant height and flower count under various biostimulant treatments align with the shifts in microbial community composition and diversity identified, revealing the occurrence of interesting PGPB92,93.
Phenotypically, the use of nitrate-deficient culture of L. indica (BME N− or CM N−) as a biostimulant through amendment application significantly enhanced plant height, and flower production, at T35 compared to the water control, while foliar application of CM of L. indica containing nitrate (CM N+) increased plant size. Nevertheless, biostimulants containing nitrate (CM N+) presented an inhibitory effect on plant size. These results suggest that the composition of the biostimulant influences its efficacy, notably through its impact on the microbiota associated with plant roots and rhizosphere. Genomic data corroborate these findings, revealing significant shifts in microbial community composition and diversity over time. The α-diversity analysis (Shannon index) demonstrated a notable increase in microbial diversity from T0 to T35 under CM N− and BME N− treatments, suggesting enhanced microbial community complexity. This increase in diversity is consistent with the presence of unique genera, particularly under nitrate-deficient conditions, as evidenced by the Venn diagram analysis (Supplementary Fig. 4). PERMANOVA (Supplementary Table 1) further confirmed the significant influence of both time and biostimulant conditions on microbial community structure, supporting the temporal nature of the phenotypic responses.
In addition, key microbial genera enriched under biostimulation treatments, such as Rhodanobacter, Devosia, Azospirillum, and Flavipsychrobacter, are recognised for their roles in nitrogen fixation, phytohormone production, and stress resilience10,69,70,71,73,74,94. More importantly, Devosia and Azospirillum are associated with improved nitrogen availability and root development, suggesting better plant development and therefore aligning with the enhanced flower production observed under CM N− biostimulation10. Similarly, Rhodanobacter and Flavipsychrobacter contribute to auxin production and nutrient uptake, which may explain the increased plant height under BME N+ treatments69,70,71,94. These genera, enriched predominantly at T35, likely play a role in supporting the later stages of plant growth and flowering as observed by ref. 10 and ref. 55.
Temporal dynamics of microbial community composition further elucidate the relationship between plant phenotypes and microbiota. Early-stage dominance of genera such as Massilia and Rhodanobacter at T0-T20 suggests their role in plant establishment and early growth78,79,80. By T35, the enrichment of genera like Devosia and Azospirillum corresponds with the onset of flowering and biomass accumulation10,55. These temporal patterns reinforce the importance of microbiota establishment over time by benefiting from the biostimulation with L. indica.
The demonstrated efficacy of nitrate-deficient L. indica cultures (CM N−, BME N−) as biostimulants emphasises the necessity of optimising formulations to selectively enrich plant growth-promoting microbial genera. Notably, biostimulants derived from CM N− cultures are rich in polysaccharides, as observed in section “L. indica culture fraction characterisation”. These polysaccharides have been reported to play a major role in promoting beneficial interactions between plants and microorganisms95,96. Indeed, polysaccharides serve as a crucial energy source for bacteria, facilitating their proliferation and activity, and also contribute to antifungal and pathogen-suppressing effects in plants97. Our data highlight clear nitrate-dependent shifts in both internal and excreted polysaccharide profiles. While these observations strongly suggest a potential mechanistic role of polysaccharides in shaping plant-microbiome interactions, we acknowledge that this linkage remains hypothesis-generating, as direct functional assays were beyond the scope of the present study. Nonetheless, these findings provide a solid foundation for future work aimed at experimentally testing the contribution of specific polysaccharide fractions to plant responses and microbiota modulation. Furthermore, microbial activity associated with these biostimulants has been suggested to enhance nutrient availability and uptake in plants, thereby promoting growth98,99. Additionally, polysaccharides have been linked to improvements in soil structure by increasing soil aggregation and water retention, fostering a more conducive environment for root development100. The presence of polysaccharides in biostimulants can also stimulate the production of EPS by beneficial bacteria, further reinforcing plant-microbe interactions and enhancing plant resilience to environmental stresses. Altogether, this enriched environment with polysaccharides shows promising properties to prevent and cope with different abiotic stresses9,10,94,101,102.
Nevertheless, while polysaccharides play a significant role, other bioactive compounds and free metabolites present in the L. indica culture fraction may also contribute to plant-microbe interactions and stress mitigation. Metabolites such as organic acids, volatile organic compounds, amino acids, and phenolic compounds can influence microbial community composition by selectively promoting the growth of beneficial microorganisms or inhibiting pathogens3,103,104,105,106. Organic acids, for instance, are known to facilitate nutrient solubilisation, improving the bioavailability of essential elements such as phosphorus and iron107. Additionally, amino acids, vitamins and volatile organic compounds may serve as signalling molecules, modulating microbial activity and inducing plant defence responses108,109,110,111,112. Certain secondary metabolites, including flavonoids and phytohormone-like compounds, could further enhance plant resilience to abiotic stresses by regulating oxidative stress responses and improving water-use efficiency.
Altogether, while polysaccharides represent a key component of these biostimulants, the combined action of multiple metabolites likely contributes to their overall efficacy. Future studies should aim to understand the relative contributions of these various compounds to optimise biostimulant formulations for enhanced plant growth and stress resilience.
Discussion
Most studies investigating the biostimulant potential of cyanobacteria have essentially focused on biomass or on specific biomass fractions, such as polysaccharides, proteins, or lipid extracts. By contrast, very few studies have examined the potential role of the residual culture medium, which can accumulate a variety of extracellular metabolites secreted during growth113. The downstream management of this culture medium represents a critical bottleneck in the economics of cyanobacteria cultivation, as large volumes are generated and their disposal or recycling impacts production costs. Several studies have therefore considered either the reuse of CM for successive batches114,115 or the substitution of synthetic media with wastewater streams as a more sustainable input115,116. In this context, our work provides novel insights by evaluating, for the first time, the biostimulant potential of the residual culture medium from L. indica cultures, particularly under conditions of nitrogen deprivation.
Furthermore, to our knowledge, there are no or very few studies that have addressed this research by modifying the metabolism of L. indica during cultivation in such a way as to induce metabolic shifts that cause a drastic change in intracellular and extracellular metabolites. In the present study, we demonstrate that both the biomass and residual culture medium (CM N−, BME N−) of L. indica cultivated under nitrogen deficiency show markedly higher biostimulant activity than when it is grown under nitrate-replete conditions. Nitrogen deprivation induced a substantial increase in polysaccharide synthesis, including extracellular polymeric substances (EPS), and altered the protein-to-polysaccharide ratio. Such shifts are consistent with previous reports in other cyanobacteria and microalgae, where nitrogen stress promotes carbon flux redistribution toward carbohydrate storage and EPS production117.
Despite the relatively small number of plants tested per treatment (n = 5), we observed a significant increase in floral production (+49.8% compared to the water control; p-value <0.05, Student’s t-test) in plants treated with the CM N− fraction. These promising results highlight the potential of residual culture medium as a low-cost, sustainable biostimulant. Future studies should aim to increase replication, extend the experimental period, and assess fruit yield and quality. Indeed, microalgal or cyanobacteria treatments, whether in the form of aqueous or ethanolic extracts, crude biomass, polysaccharide fractions, or foliar suspensions, have already been shown to modulate tomato fruit composition, including pigments (carotenoids, lycopene), antioxidants (phenolics), vitamins (e.g. vitamin C), mineral accumulation (Fe, Zn), and sugars118,119,120. These results strongly suggest that extending the evaluation beyond floral induction to fruit biochemical profiling would provide a more comprehensive picture of the benefits of L. indica extracts.
Another key finding of our study concerns the plant-associated microbiome. Interestingly, we showed that the profiling of the rhizosphere and endosphere bacteriome revealed a significant change in alpha diversity, particularly with CM N− and BME N− in both applications. The enrichment in polysaccharides under N− conditions appears to enhance specific plant-microbiota interactions, as evidenced in this study. We observed an increase in plant growth-promoting bacteria (PGPB), including Acidovorax (increased by 4% from T0 to T35 through soil amendment), Devosia (increased by 3% from T0 to T25 through soil amendment, and 4% through foliar application), and Massalia (increased by 22% from T0 to T35 through soil amendment), following biostimulation with the CM N− fraction.
These data suggest a direct and an indirect action of the bioactive compounds depending on the mode of application (soil amendment versus foliar application) and the extract tested (residual CM versus BME from L. indica cultures in the presence or nitrate-deficient condition). Polysaccharides, particularly those present in residual CM, are probably involved in the effects observed, both at the phenotypic level and at the microbial level associated with the rhizosphere and endosphere. Indeed, the polysaccharides have already been reported to play a major role in promoting beneficial interactions between plants and microorganisms95,96. Polysaccharides serve as a crucial energy source for bacteria, facilitating their proliferation and activity, and contribute to antifungal and pathogen-suppressing effects in plants97. Furthermore, microbial activity associated with these biostimulants has been suggested to enhance nutrient availability and uptake in plants, thereby promoting growth98,99. Additionally, polysaccharides have been linked to improvements in soil structure by increasing soil aggregation and water retention, fostering a more conducive environment for root development100. The presence of polysaccharides in biostimulants can also stimulate the production of EPS by beneficial bacteria, further reinforcing plant-microbe interactions and enhancing plant resilience to environmental stresses. Altogether, this enriched environment with polysaccharides shows promising properties to prevent and cope with different abiotic stresses9,10,94,101,102.
Beyond their nutritive role, polysaccharides also function as elicitors of plant defence responses121. Compounds such as laminarin, alginates, fucoidans, and carrageenans have been shown to activate defence signalling pathways, modulate secondary metabolism, and alter root exudation profiles, thereby reshaping soil microbial communities122,123. These dual effects, direct metabolic priming of plants and indirect modulation of the microbiome, suggest that polysaccharides are central mediators of the biostimulant activity observed in our study. As demonstrated by ref. 121, ethanol-precipitated purified extracts of polysaccharides from Cyanobacteria, including L. indica, show effects on defence and metabolic markers in tomatoes.
Nevertheless, polysaccharides are unlikely to act alone. Cyanobacteria CM contain a wide array of metabolites, including organic acids, amino acids, phenolic compounds, volatile organic compounds (VOCs), and phytohormone-like molecules, all of which can influence plant physiology and microbiome composition. Organic acids are well-documented for their roles in nutrient solubilization, particularly phosphorus and iron107. Amino acids, vitamins, and VOCs can act as signalling molecules that modulate microbial activity and induce systemic plant responses108,111,112. Secondary metabolites such as flavonoids or indole-like compounds may further enhance abiotic stress resilience by regulating oxidative balance and water-use efficiency3. Thus, the combined action of multiple metabolite classes, rather than polysaccharides alone, likely underpins the biostimulant efficacy of the CM N− fraction.
Taken together, our findings support a model in which L. indica extracts act through both direct effects on plant physiology and indirect effects mediated via microbiome modulation. Future research should aim to disentangle the relative contributions of these compound classes using fractionation approaches (e.g. purified EPS versus low-molecular-weight metabolites). Integrative approaches combining metabolomics of exudates, plant proteo-transcriptomics, and meta-omics of associated microbiota (metaproteomics, metagenomics, metatranscriptomics) will be essential to elucidate the complex tripartite interactions between plant, bacteria-derived compounds, and the microbiome. In parallel, functional assays (e.g. nifH qPCR, enzyme activities) would be beneficial to directly test the functions associated with the genera identified in the taxonomic analysis. Additionally, stable isotope tracing (13C, 15N) could be used to follow the transfer of bacteria-derived carbon and nitrogen into plant tissues and microbial communities. Finally, time-course experiments across hours, days, and full crop cycles would provide valuable insights into the persistence and dynamics of these interactions.
In conclusion, this study highlights that cyanobacteria biostimulants are not limited to biomass but also include the residual culture medium. Exploring this often-overlooked component reveals significant biostimulant potential, particularly when cyanobacteria are cultivated under nitrogen stress, a condition that redirects metabolic fluxes toward enhanced production of polysaccharides, including EPS. These compounds act not only directly on plant physiology but also indirectly by reshaping the associated microbiome, thereby strengthening plant growth and resilience.
The demonstration that polysaccharide-enriched residual culture medium stimulates flowering and promotes beneficial interactions within the rhizosphere opens new perspectives for sustainable agriculture. By valorising a fraction often regarded as waste, this approach offers a dual advantage: reducing costs linked to spent medium management while increasing the profitability of cyanobacteria production. Thus, the use of residual culture medium as a biostimulant emerges as both an economically attractive and environmentally sustainable strategy, enhancing the overall appeal of Spirulina cultivation.
Methods
Limnospira indica culture
Limnospira indica strain (PCC8005 from Paster Institute) was maintained in 250 mL Erlenmeyer flasks in a photosynthetic growth chamber (MLR-352H-PE, Panasonic, Japan) at 30 °C, 140 rpm shaking, and continuous illumination of 60 μmol. photons. m−2. s−1 (Li-193SA; Li-Cor BioSciences, USA). Two media were used: Cogne Modified Zarrouk medium (ref. 124) (N+ hereafter) and Cogne Modified Zarrouk medium nitrate-depleted (N− hereafter). Volume was increased in 2-litre glass bottles for biostimulant extract preparation.
Biostimulant solutions preparation
Four biostimulant fractions containing potential bioactive compounds from L. indica were extracted following the procedure (Fig. 10):
-
CM from a fresh culture of L. indica (OD = 1,7) grown in N+ and N− conditions were retrieved by centrifugation (3800 rpm for 20 min) after a 20 min sonication bath (Elmasonic S 100H) in order to break the vacuoles and enable efficient centrifugation.
-
Biomass from a fresh culture of L. indica (OD = 1.7) growth in N+ and N− conditions was retrieved by centrifugation (3800 rpm for 20 min) after a 20 min sonication bath. The biomass was resuspended in the same volume of Milli-Q water as the initial culture volume and boiled for 1 h. The supernatant is retrieved by centrifugation (3800 rpm for 20 min).

L. indica cultures (N+ and N−) are centrifuged, the supernatant is retained for CM solutions, and the biomass is further processed by boiling to produce the BME. Realised with Procreate®.
Both N+ and N− were harvested together at OD = 1.7 (corresponding to a DW of 1.4 in the N+ condition and 1.6 in the N− condition, Supplementary Fig. 8) before N− culture started the plateau phase and declined. At this stage, both culture growth was similar (Fig. 1). An independent culture per condition was done before each experiment, described in the subsection “Solanum lycopersicum seedling growth biostimulation”. Biostimulant solutions are used immediately after preparation, and are stored at 4 °C for the course of the experiment.
These solutions were diluted in Milli-Q water as follows:
-
For amendment treatment: culture supernatant solution (2.5% (v/v)), biomass solution (2.5% (v/v)) or water only, with Tween-20® (0.01% (v/v)) in all solutions.
-
For foliar treatment: culture supernatant solution, biomass solution (25% (v/v)) or water only, with Tween-20® (0.01% (v/v)) in all solutions.
Due to volume application constraints on the leaves (leaf area without applications on the soil), the biostimulant solution has been concentrated (10×), resulting in a 2.5% concentration when applied by amendment against a 25% concentration when applied by foliar application. The pH of the working solutions (mean ± SD) was: CM N+, 10.26 (±0.05); CM N−, 10.51 (±0.1); BME N+, 8.85 (±0.03); BME N− 8.74 (±0.02); Negative control, 9.84 (±0.02). In addition, Tween-20®, as a surfactant, was used to increase the cells’ permeability, allowing biostimulant compounds to penetrate the plants’ tissues125,126.
L. indica culture fraction characterisation
For carbohydrate quantification, L. indica CM (supernatant) were first washed on ultrafiltration units (Vivaspin® 100 KDa, Sartorius®) to eliminate salts present in the medium and concentrate EPS in MQ-water. The concentrated supernatants were mixed with 3 volumes of absolute ethanol to precipitate polysaccharides. In parallel, L. indica cells were freeze-dried (FINN-AQUA®, LYOVAC GT 2E) before being incubated for 2 hrs at 100 °C in HCl 0.6 M to collect capsular and internal polysaccharides.
Carbohydrate assay was performed using (ref. 127) methods as follows. 10 μL of phenol 80% (80:20 v/v in ethanol) solution and 1 mL of 98% H2SO4 were respectively added to 400 μL of the samples. A standard curve was generated with D-glucose as follows: 0μg. mL−1, 12.5 μg.mL−1, 25 μg. mL−1, 50 μg. mL−1, 100 μg. mL−1, 150 μg. mL−1. After 10 min of incubation at room temperature, the optical density was read at 490 nm.
Safety hazards: Phenol may cause damage to organs through prolonged or repeated exposure. Phenol is a mutagen and may cause damage to organs through prolonged or repeated exposure. Sulfuric acid, H2SO4, may irritate and burn the skin and eyes, and may lead to blindness. Exposures may cause headache, nausea and vomiting.
For Protein quantification, L. indica biomass was filtered on a 0.2 μm (Sartorius®, cellulose nitrate filter) using a vacuum pump. L. indica biomass was freeze-dried (FINN-AQUA®, LYOVAC GT 2E). Dried samples were resuspended in a lysis extraction buffer from the Preomics iST Sample preparation kit®. Samples were vortexed at room temperature for 20 min, followed by a 10 min incubation at 95 °C at 1000 rpm. Then, a sonication by probe step (3 × 10 sec, amplitude 20%, IKA U50 sonicator) at 4 °C was performed. Cell debris was removed by centrifugation (15 min, 1100 rpm, 4 °C). Proteins were recovered within the supernatant. Protein assays were performed using the Bradford method128. The standard curve was done by using BgG in MQ-water as follows: 0 μg μL−1, 1 μg μL−1, 2 μg μL−1, 5 μg μL−1, 8 μg μL−1, 10 μg μL−1. Colourimetric assay was performed using Bradford reagent, and the optical density was read at 595 nm, immediately after the reagent mix.
Last, nitrate assays were performed following129 protocol. 125μL of sample were mixed thoroughly with 800μL of 5% (w/v) salicylic acid in concentrated H2SO4. After 20 min of incubation at room temperature, 9.5 mL 2N NaOH were added slowly before cooling at room temperature. The standard curve was generated with NaNO3 as follows: 20 mM, 10 mM, 2.5 mM, 1.25 mM, 0.6 mM, 0.3 mM, and 0.15 mM. Samples and standards were read within 20 min at 410 nm.
Safety hazards: Sulfuric acid, H2SO4, may irritate and burn the skin and eyes, and may lead to blindness. Exposures may cause headache, nausea and vomiting. Sodium hydroxide, NaOH, may cause severe skin burns and eye damage.
Composition of polysaccharides from L. indica
Polysaccharides from supernatant (released polysaccharides, RPS) and biomass (capsular and endopolysaccharides) of both nitrate-deficient (N−) and nitrate-rich (N+) cultures composition in monosaccharides was analysed. Polysaccharides from the supernatant were concentrated using ultrafiltration units (Vivaspin® 100 KDa, Sartorius®) (centrifugation at 4000 × g for 10 min) before being washed 3× with MQ-water. In parallel, L. indica cells were freeze-dried (FINN-AQUA®, LYOVAC GT 2E). The following saccharides were used as standard: D-Glucuronic acid, D-Glucose, D-Mannose, D-Xylose, D-Galactose, D-Arabinose, L-Rhamnose, D-Galacturonic acid, D-Ribose, Fucose, N-acetylglucosamine, N-acetyl galacturonic. All saccharides are diluted in ULC-MS water at a concentration of 50 mg/mL.
Concentrated RPS, dried biomass and 20 μL of standard solutions were hydrolysed with 2 mL of 4 M TFA (Trifluoroacetic acid) at 98 °C for 5 h. The samples were evaporated to dryness, and the pellets were resuspended in 500 μL of ULC MS water. Samples were filtered through a 0.2 μm membrane (Whatman® Mini-UniPrep®).
Liquid chromatography was performed using a Sciex Exion LC HPLC. All samples were separated on a XBridge® BEH Amide column (130 Å, 2.1 × 150 mm, 5 μm) (Waters Corporation®, 186006590). The solvent system consisted of two mobile phases as follows: mobile phase A (H2O, formic acid 0.1%) and mobile phase B (Acetonitrile 99.9%, formic acid 0.1%). The column operated at a flow rate of 0.2 mL min−1 at 40 °C. The gradient elution program was as follows: 0–1 min, 15% B; 1–5 min, 15–17% B; 5–9 min, 17–20% B; 9–12 min, 20–25% B; 12–12.5 min, 25–35% B; 12.5–13.5 min, 35% B; 13.5–14 min, 35–15% B; 14–15 min, 15% B.
Mass spectrometry was performed on a high-resolution mass spectrometer (SCIEX ZenoTof™ 7600), acquiring data in full scan ion mode and tandem MS/MS using an electrospray ionisation (ESI) source. Fragmentation was performed using collision-induced dissociation (CID). MS was operated using a positive mode (collision energy voltage 10 V, electrospray voltage 5500 V, declustering potential 50 V) with a source temperature of 200 °C. MSMS was performed using a negative mode as well (collision energy voltage 40 V, declustering potential 50 V, CE spread 20 V).
Analyses were performed using SCIEX OS software (version 3.3.1.43).
Safety hazards: Trifluoroacetic acid, TFA, is a corrosive chemical that may severely irritate and burn the skin and eyes, with possible eye damage. Breathing Trifluoroacetic Acid may irritate the nose, throat and lungs.
S. lycopersicum seedling growth biostimulation
The GreenHab environment was prepared to suit the needs of S. lycopersicum at the two-leaf stage for 5 weeks. The experiment was conducted in a greenhouse (Mammoth PRO + DR 240 L) under controlled conditions with a 16 h light/8h darkness cycle and an average temperature over time of 26 °C. Two General Electric 400 W XO HPS were used for the illumination. The temperature and the air flow inside the GreenHab were regulated by one air extractor (VK 200 U 780 m3/h) and by three fans continuously turned ON. The light was switched ON from 7 a.m. to 8 p.m. each day, and the intensity during the day was 440 μmol photons m−2 s−1.
The tomato plants S. lycopersicum (variety MoneyMakers) (ordered at the C.A.R.A.H, Ath, Belgium) were received at a 2-leaf stage, 3 weeks after being sown, n = 5. The sample size was defined to satisfy statistical validations while fitting our GreenHab volume constraints. The plants were grown in the greenhab for 2 days before being transferred to fresh soil (DCM Potground for Professionals, pH = 6) in 10 cm diameter pots. Pots were circular (top ({rm{varnothing }}) 8.5 cm, bottom ({rm{varnothing }}) 7.5 cm, height 7.0 cm) with an internal geometric volume of ≈0.352 L. All plants were potted using soil from the same batch. Plants were watered with Milli-Q water for 7 days before the experiment, every 2 days for amendment application and every day for foliar treatment. Foliar treatments were prepared at a tenfold higher concentration than the soil amendment, minimising application volume and improving leaf uptake while preventing runoff.
For amendment treatment, biostimulant solutions were administered (100 mL of 2.5% solution for the first 19 days, then 200 mL of 2.5% solution for the following 15 days) on the soil by carefully avoiding any contact with the leaves (Fig. 11).

Biostimulants were applied on the leaves or the soil, and samples were collected after 0, 10, 20 and 35 days. Realised with Procreate®.
For foliar treatment, biostimulant solutions were administrated by spraying them onto the leaves every day (5 mL of 25% solution for the first 19 days, then 10 mL of 25% solution for the following 15 days) by carefully avoiding the soil (soil was shielded using a pot-sized cardboard cover) while Milli-Q water (100 mL for the 19 first days, then 200 mL) was applied on the soil (Fig. 11).
Biostimulant solution for amendment treatment and watering solution for foliar treatment were top-applied to the soil surface within a ≈2–3 cm radius of the stem, solutions were not mixed into the substrate. Pots sat on saucers and were inspected after dosing; no leachate was observed, indicating full retention in the substrate. The irrigation followed a standard greenhouse regime applied equally across treatments.
Plants characteristics (size of the plant, number of branches, leaves, floral buds, flowers, and early fruits) are collected after 0, 10, 20 and 35 days, after transplanting, reaching stage 6 of the BBCH scale (ref. 130 and Supplementary Fig. 9). This time point was selected as it corresponds to the onset of the reproductive stage in tomato, a phenological important transition where biostimulants may exert significant effects. Here, because measurements were taken at 35 days, the flower stage and fruit production responses capture early reproductive phenology (flowering onset/initial fruit set). In parallel, the roots and the rhizosphere soil are collected for bacterial sequencing analysis (16S sequencing). Due to space management, the samples have been divided into 3 batch experiments (following each other in time). NMDS and PERMANOVA analysis have been performed on T0 from each batch in order to determine our ability to directly compare batches together (Supplementary Fig. 10 and Supplementary Table 5). Our analysis revealed that T0 where different, and for full transparency, batch analyses are kept separate. Samples were further analysed by experiment, represented in the following table:
Biostimulant impact on rhizosphere and endosphere microbiota
In this study, the rhizosphere denotes soil present around the root system, adhering to the root system (root-associated soil, centre of the bulk soil). The root fraction comprises microorganisms associated with the roots, including both surface-attached (rhizoplane) and internal (endophytic) communities. To capture both compartments, we intentionally did not perform surface sterilisation. Accordingly, this fraction represents the combined root-associated community rather than a strictly sterilised endosphere.
DNA extraction, amplification and 16S rRNA amplicon sequencing were performed as follows: Samples (Table 3) of the rhizosphere soil and roots were collected in sterile conditions (under flame) for targeted genomic analysis after 0, 20 and 35 days. Roots were washed, in sterile conditions, in 3 sterile PBS baths to remove the remaining soil. Two millilitres of PBS were added to the primary cut-off roots before crushing them using the UltraTurax. Samples were centrifuged at max speed for 15 min, supernatant was kept at −40 °C. DNeasy Power Soil kit (Qiagen®) was performed using the pellet for the roots sample and 250 mg of soil (without remaining roots) for the rhizosphere samples, following the manufacturer’s instructions. Subsequent DNA was quantified using the QuantiFluor® ONE dsDNA System (Promega®).
To characterise the bacterial communities, the entire 16S rRNA gene was amplified using the 16S Barcoding Kit 24 V14 (SQK-16S114.24, Oxford Nanopore Technologies), allowing for broad taxonomic resolution. The DNA was amplified by PCR using specific 16S primers131:
27F (5‘-AGAGTTTGATCMTGGCTCAG-3’) and 1492R (5‘-CGGTTACCTTGTTACGACTT-3’),
which spans nearly the full length of the 16S rRNA gene132 and contains 5’ tags which facilitate the ligase-free attachment of Rapid Sequencing Adaptors added in the library. DNA amplification was performed using 20 ng of DNA. The PCR conditions were set according to the kit’s protocol, with an initial denaturation of 1 min at 95 °C followed by 25 cycles of denaturation (20 s at 95 °C), annealing (30 s at 55 °C) and extension (2 min at 65 °C), and a final extension 5 min at 65 °C. Barcodes were ligated to the amplicons according to the kit’s protocol during the PCR, allowing multiplexing of samples. 80 ng of amplified DNA were purified using the manufacturer’s instructions, and DNA quantification was performed using the QuantiFluor® ONE dsDNA System (Promega®). Purified 16S rRNA gene amplicons were prepared for sequencing using the 16S Barcoding Kit SQK-16S114.24.
The sequencing library was prepared by following the manufacturer’s instructions using 50 ng of purified DNA combined from 24 samples per flow-cell. The sequencing was carried out on the GridION Mk1 using flow cells (FLO-MIN114.8) with a pore count of above 1200. The sequencing run was conducted for 20 h with a minimum read length of 1000 bp, a high-accuracy basecalling, a sequencing score of 9 and a minimum barcoding score of 60.
The taxonomic identification was performed with the EPI2ME tool (Oxford Nanopore), workflow wf-16s for the identification of the origin of single reads from 16S amplicon sequencing. The basecalling and demultiplexing were done using MinKNOW (24.02.16) and Dorado (7.3.11, integrated in MinKNOW), as well as the workflow WF_16S v1.5.0, which uses minimap2 (v2.26-r1175). The following parameters have been used: classification method = minimap2, Database = ncbi-16S–18S, taxonomic rank = G, minimum reads length = 1200, maximum reads length = 1800, minimum score quality = 9. After identification, all identifications with fewer than ten reads or identified as “incertae sedis”133 are removed from the analysis.
Quality of the sequencing has been verified through rarefaction curves analysis (Supplementary Fig. 11 and Supplementary Table 6) using a Python script (in Supplementary Script 1)134. The α-diversity (Shannon index) was calculated on each rarefied sample set (Limma RGui package, using RStudio version 2024.04.2), excluding samples with less than 40,000 reads for rhizosphere samples and less than 4000 for endosphere samples, resulting in the exclusion of one unique sample (labelled in red in Supplementary Table 7). The β-diversity was first assessed through non-metric MultiSimensional Scaling (NMDS) profile of the pairwise community dissimilarity (Bray–Curtis) indices of the 16S sequencing data was realised using the PAST software (version 4.17)135,136. Venn diagrams were produced using Rgui on RStudio134,137 to determine the genus distributions of different conditions. β-diversity was evaluated using dissimilarity and multivariate analyses by using the Rgui package vegan (version 2.6-8) using RStudio138. Relative abundance is represented in the present study using the Circos online tool (https://mk.bcgsc.ca/tableviewer/visualize/), allowing highlight of similarities and differences from the 10 most representative genera per condition139. Last, to test the significance of the factors analysed (Condition, Time), PERMANOVA tests were performed on the Bray–Curtis dissimilarity using 100,000 permutations using RStudio. In addition, PERMANOVA with the strata argument was applied to account for inter-batch variation, and pairwise comparisons between conditions were assessed using the pairwiseAdonis method.
The identification and validation of response groups (RG) were performed following134 analysis method. The total of genus identified amongst the 8 datasets analysed in the study (Table 3) as significantly affected genus by the condition are plotted on respective heatmaps. These genera were defined by the deviance analysis on the rarefied counts under nbGLM (negative binomial distribution and Generalised Linear Model), revised by 1000 resampling iterations of the residual variance by the utilisation of the Rgui package mvabund140. RGs were defined with a hierarchical cluster dendrogram (Euclidean distance and average clustering) using the Rgui package vegan138. The statistical validity of these groups was assessed by comparing the RG clustering with a null model by Monte-Carlo simulation (Supplementary Fig. 12) by using all the genera to reinforce the randomisation process.
Data availability
All raw data are available in the NCBI repository, under project accession PRJNA1328183: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1328183. Python code used for rarefaction is provided in the Supplementary data.
Code availability
Python code used for rarefaction is provided in Supplementary data.
References
-
Tilman, D., Balzer, C., Hill, J. & Befort, B. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 108, 20260–4 (2011).
-
Tripathi, S., Srivastava, P., Devi, R. S. & Bhadouria, R. Influence of synthetic fertilizers and pesticides on soil health and soil microbiology. In: Agrochemicals Detection, Treatment and Remediation (ed. Prasad, M. N. V.) 25–54 (Butterworth-Heinemann, 2020).
-
Renaud, C., Leys, N. & Wattiez, R. Photosynthetic microorganisms, an overview of their biostimulant effects on plants and perspectives for space agriculture. J. Plant Interact. 18, 2242697 (2023).
-
Lasseur, C. et al. MELISSA: Overview of the Project and Perspectives. SAE Technical Paper 2005-01-3066 (SAE International, Warrendale, 2005).
-
Lasseur, C. et al. Melissa: the European project of closed life support system. Gravit. Space Res. 23, 2010 (2010).
-
Kapoore, R. V., Wood, E. E. & Llewellyn, C. A. Algae biostimulants: a critical look at microalgal biostimulants for sustainable agricultural practices. Biotechnol. Adv. 49, 107754 (2021).
-
Yakhin, O. I., Lubyanov, A. A., Yakhin, I. A. & Brown, P. H. Biostimulants in plant science: a global perspective. Front. Plant Sci. 7, 2049 (2017).
-
Adeleke, B. S., Babalola, O. O. & Glick, B. R. Plant growth-promoting root-colonizing bacterial endophytes. Rhizosphere 20, 100433 (2021).
-
Hamid, B. et al. Bacterial plant biostimulants: a sustainable way towards improving growth, productivity, and health of crops. Sustainability 13, 2856 (2021).
-
Novello, G. et al. The impact of nitrogen-fixing bacteria-based biostimulant alone or in combination with commercial inoculum on tomato native rhizosphere microbiota and production: an open-field trial. Biology 13, 400 (2024).
-
Olanrewaju, O. S. & Babalola, O. O. The rhizosphere microbial complex in plant health: a review of interaction dynamics. J. Integr. Agric. 21, 2168–2182 (2022).
-
Pantigoso, H. A., Newberger, D. & Vivanco, J. M. The rhizosphere microbiome: plant–microbial interactions for resource acquisition. J. Appl. Microbiol. 133, 2864 (2022).
-
Li, J., Van Gerrewey, T. & Geelen, D. A meta-analysis of biostimulant yield effectiveness in field trials. Front. Plant Sci. 13, 836702 (2022).
-
Mendes, R., de Barros, I., D’Andréa, P. A., D’Andréa-Kühl, M. S. C. & Rodrigues, G. S. A multi-attribute approach to evaluating the impact of biostimulants on crop performance. Front. Plant Sci. 14 (2023).
-
Fernández, V. & Eichert, T. Uptake of hydrophilic solutes through plant leaves: current state of knowledge and perspectives of foliar fertilization. Crit. Rev. Plant Sci. 28, 36–68 (2009).
-
Ofori, P. A. et al. Greenhouse tomato production for sustainable food and nutrition security in the tropics. In: Tomato—From Cultivation to Processing Technology (IntechOpen, 2022).
-
Nicola, S., Tibaldi, G., Fontana, E., Crops, A.-V. & Plants, A. Tomato production systems and their application to the tropics. Acta Hortic. 821, 27–34 (2009).
-
Beecher, G. R. Nutrient content of tomatoes and tomato products. Proc. Soc. Exp. Biol. Med. 218, 98–100 (1998).
-
Colman, S. L. et al. Chitosan microparticles improve tomato seedling biomass and modulate hormonal, redox and defense pathways. Plant Physiol. Biochem. 143, 203–211 (2019).
-
Hernández, V., Botella, M. Á, Hellín, P., Fenoll, J. & Flores, P. Dose-dependent potential of chitosan to increase yield or bioactive compound content in tomatoes. Horticulturae 8, 1152 (2022).
-
Stasińska-Jakubas, M. & Hawrylak-Nowak, B. Protective, biostimulating, and eliciting effects of chitosan and its derivatives on crop plants. Molecules 27, 2801 (2022).
-
Reyes-Pérez, J. J. et al. Chitosan application improves the growth and physiological parameters of tomato crops. Horticulturae 11, 878 (2025).
-
Liu, X., Sun, R., Hu, S., Zhong, Y. & Wu, Y. Aromatic compounds releases aroused by sediment resuspension alter nitrate transformation rates and pathways during aerobic-anoxic transition. J. Hazard. Mater. 424, 127365 (2022).
-
Flores, S. S. et al. The tomato’s tale: exploring taxonomy, biogeography, domestication, and microbiome for enhanced resilience. Phytobiomes J. 8, 5–20 (2024).
-
Mejia, G. et al. Rhizosphere microbiome influence on tomato growth under low-nutrient settings. FEMS Micrbiol. Ecol. 101, fiaf019 (2024).
-
Mutale-joan, C. et al. Microalgae-cyanobacteria-based biostimulant effect on salinity tolerance mechanisms, nutrient uptake, and tomato plant growth under salt stress. J. Appl. Phycol. 33, 3779–3795 (2021).
-
Santini, G., Biondi, N., Rodolfi, L. & Tredici, M. R. Plant biostimulants from cyanobacteria: an emerging strategy to improve yields and sustainability in agriculture. Plants 10, 643 (2021).
-
Curiel-Alegre, S. et al. Evaluation of biostimulation, bioaugmentation, and organic amendments application on the bioremediation of recalcitrant hydrocarbons of soil. Chemosphere 307, 135638 (2022).
-
Garza-Alonso, C. A. et al. Strawberry biostimulation: from mechanisms of action to plant growth and fruit quality. Plants 11, 3463 (2022).
-
Tekaya, M. et al. Foliar application of fertilizers and biostimulant has a strong impact on the olive (Olea europaea) rhizosphere microbial community profile and the abundance of arbuscular mycorrhizal fungi. Rhizosphere 19, 100402 (2021).
-
Şesan, T. E. et al. Effects of foliar treatment with a trichoderma plant biostimulant consortium on Passiflora caerulea L. yield and quality. Microorganisms 8, 123 (2020).
-
Deschoenmaeker, F. et al. Impact of different nitrogen sources on the growth of Arthrospira sp. PCC 8005 under batch and continuous cultivation—a biochemical, transcriptomic and proteomic profile. Bioresour. Technol. 237, 78–88 (2017).
-
Deschoenmaeker, F. et al. Nitrogen depletion in Arthrospira sp. PCC 8005, an ultrastructural point of view. J. Struct. Biol. 196, 385–393 (2016).
-
Li, Z. et al. Effects of culture conditions on the performance of Arthrospira platensis and its production of exopolysaccharides. Foods 11, 2020 (2022).
-
Mousavi, M., Mehrzad, J., Najafi, M. F., Zhiani, R. & Shamsian, S. A. A. Nitrate and ammonia: two key nitrogen sources for biomass and phycocyanin production by Arthrospira (Spirulina) platensis. J. Appl. Phycol. 34, 2271–2281 (2022).
-
Levantesi, C. et al. Filamentous Alphaproteobacteria associated with bulking in industrial wastewater treatment plants. Syst. Appl. Microbiol. 27, 716–727 (2004).
-
Geada, P. et al. 17-Cyanobacterial toxins as a high value-added product. In: Microalgae-based Biofuels and Bioproducts. Woodhead Publishing Series in Energy (eds. Gonzalez-Fernandez, C. & Muñoz, R.) 401–428 (Woodhead Publishing, 2017).
-
Forchhammer, K. & Selim, K. A. Carbon/nitrogen homeostasis control in cyanobacteria. FEMS Microbiol. Rev. 44, 33–53 (2020).
-
Richaud, C., Zabulon, G., Joder, A. & Thomas, J.-C. Nitrogen or sulfur starvation differentially affects phycobilisome degradation and expression of the nblA gene in Synechocystis strain PCC 6803. J. Bacteriol. 183, 2989–2994 (2001).
-
Forchhammer, K. & Schwarz, R. Nitrogen chlorosis in unicellular cyanobacteria—a developmental program for surviving nitrogen deprivation. Environ. Microbiol. 21, 1173–1184 (2019).
-
Bertocchi, C., Navarini, L., Cesàro, A. & Anastasio, M. Polysaccharides from cyanobacteria. Carbohydr. Polym. 12, 127–153 (1990).
-
Laroche, C. Exopolysaccharides from microalgae and cyanobacteria: diversity of strains, production strategies, and applications. Mar. Drugs 20, 336 (2022).
-
De Philippis, R. & Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 22, 151–175 (1998).
-
De Porcellinis, A., Frigaard, N.-U. & Sakuragi, Y. Determination of the glycogen content in cyanobacteria. J. Vis. Exp. 17, 56068 (2017).
-
Gründel, M., Scheunemann, R., Lockau, W. & Zilliges, Y. Impaired glycogen synthesis causes metabolic overflow reactions and affects stress responses in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 158, 3032–3043 (2012).
-
Hasunuma, T. et al. Dynamic metabolic profiling of cyanobacterial glycogen biosynthesis under conditions of nitrate depletion. J. Exp. Bot. 64, 2943–2954 (2013).
-
Monshupanee, T. & Incharoensakdi, A. Enhanced accumulation of glycogen, lipids and polyhydroxybutyrate under optimal nutrients and light intensities in the cyanobacterium Synechocystis sp. PCC 6803. J. Appl. Microbiol. 116, 830–838 (2014).
-
Gopalakrishnan, S. et al. Plant growth promoting rhizobia: challenges and opportunities. 3 Biotech 5, 355–377 (2015).
-
Dubey, R. S. & Mohammad, P. Physiological mechanisms of nitrogen absorption and assimilation in plants under stressful conditions. In: Handbook of Plant and Crop Physiology 483–516 (CRC Press, 2014).
-
Depraetere, O. et al. Harvesting carbohydrate-rich Arthrospira platensis by spontaneous settling. Bioresour. Technol. 180, 16–21 (2015).
-
Cuellar-Bermudez, S. P. et al. Quantification of extracellular and biomass carbohydrates by Arthrospira under nitrogen starvation at lab-scale. Algal Res. 68, 102907 (2022).
-
Xia, J. L., Nie, Z. Y. & Levert, J. M. Changes in content, constituents and distribution of constitutive and excreted sugars of spirulina (Arthrospira) maxima in nutrient-limited batch cultures. In: Algae and Their Biotechnological Potential: Proceedings of the 4th Asia-Pacific Conference on Algal Biotechnology, 3–6 July 2000 in Hong Kong (eds. Chen, F. & Jiang, Y.) 135–146 (Springer Netherlands, Dordrecht, 2001).
-
Van Camp, C., Fraikin, C., Claverie, E., Onderwater, R. & Wattiez, R. Capsular polysaccharides and exopolysaccharides from Gloeothece verrucosa under various nitrogen regimes and their potential plant defence stimulation activity. Algal Res. 64, 102680 (2022).
-
Suresh Kumar, A., Mody, K. & Jha, B. Bacterial exopolysaccharides—a perception. J. Basic Microbiol. 47, 103–117 (2007).
-
Hou, Q. et al. Rhizosphere microbial diversity and community dynamics during potato cultivation. Eur. J. Soil Biol. 98, 103176 (2020).
-
İnceoğlu, Ö, Al-Soud, W. A., Salles, J. F., Semenov, A. V. & van Elsas, J. D. Comparative analysis of bacterial communities in a potato field as determined by pyrosequencing. PLoS One 6, e23321 (2011).
-
Micallef, S. A., Channer, S., Shiaris, M. P. & Colón-Carmona, A. Plant age and genotype impact the progression of bacterial community succession in the Arabidopsis rhizosphere. Plant Signal. Behav. 4, 777–780 (2009).
-
Santos, L. F. & Olivares, F. L. Plant microbiome structure and benefits for sustainable agriculture. Curr. Plant Biol. 26, 100198 (2021).
-
Preston, G. M. Plant perceptions of plant growth-promoting Pseudomonas. Philos. Trans. R. Soc. B Biol. Sci. 359, 907 (2004).
-
Chaparro, J. M., Badri, D. V. & Vivanco, J. M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 8, 790–803 (2014).
-
Bulgarelli, D., Schlaeppi, K., Spaepen, S., Ver Loren van Themaat, E. & Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 64, 807–838 (2013).
-
Schlaeppi, K., Dombrowski, N., Oter, R. G., Ver Loren van Themaat, E. & Schulze-Lefert, P. Quantitative divergence of the bacterial root microbiota in Arabidopsis thaliana relatives. Proc. Natl. Acad. Sci. USA 111, 585–592 (2014).
-
Barbaccia, P. et al. Plant growth-promoting activities of bacteria isolated from an anthropogenic soil located in Agrigento province. Microorganisms 10, 2167 (2022).
-
Ahmad, I., Pichtel, J. & Hayat, S. Plant-Bacteria Interactions: Strategies and Techniques to Promote Plant Growth (John Wiley & Sons, 2008).
-
Maheshwari, P. & Sankar, P. M. Culture-independent and culture-dependent approaches in symbiont analysis: In proteobacteria, Chap. 42. In: Microbial Symbionts, Developments in Applied Microbiology and Biotechnology (ed. Dharumadurai, D.)743–763 (Academic Press, 2023).
-
Antoun, H. & Prévost, D. Ecology of plant growth promoting rhizobacteria. In Siddiqui, Z. A. (ed.) PGPR: Biocontrol and Biofertilization 1–38 (Springer Netherlands, Dordrecht, 2006).
-
Lyu, D. et al. Plant growth-promoting rhizobacteria (PGPR) with microbial growth broth improve biomass and secondary metabolite accumulation of Cannabis sativa L. J. Agric. Food Chem. 71, 7268 (2023).
-
Fracchia, F. et al. Colonization of naive roots from populus tremula × alba involves successive waves of fungi and bacteria with different trophic abilities. Appl. Environ. Microbiol. 87, e02541–20 (2021).
-
Lee, S. A. et al. Complete genome sequence of the plant growth-promoting endophytic bacterium Rhodanobacter glycinis T01E-68 isolated from tomato (Solanum lycopersicum L.) plant roots. Microbiol. Soc. Korea 55, 422–424 (2019).
-
Nam, J. H., Thibodeau, A., Qian, Y. L., Qian, M. C. & Park, S. H. Multidisciplinary evaluation of plant growth promoting rhizobacteria on soil microbiome and strawberry quality. AMB Express 13, 18 (2023).
-
Zafar, M. et al. Effect of plant growth-promoting rhizobacteria on growth, nodulation and nutrient accumulation of lentil under controlled conditions. Pedosphere https://doi.org/10.1016/S1002-0160( (2012).
-
Zhang, X. et al. The impact of cell structure, metabolism and group behavior for the survival of bacteria under stress conditions. Arch. Microbiol. 203, 431–441 (2021).
-
Zhang, C. et al. A tripartite bacterial-fungal-plant symbiosis in the mycorrhiza-shaped microbiome drives plant growth and mycorrhization. Microbiome 12, 13 (2024).
-
Chhetri, G. et al. Devosia rhizoryzae sp. nov., and Devosia oryziradicis sp. nov., novel plant growth promoting members of the genus Devosia, isolated from the rhizosphere of rice plants. J. Microbiol. 60, 1–10 (2022).
-
Villamil, M. B. et al. Microbial signatures in fertile soils under long-term N management. Front. Soil Sci. https://doi.org/10.3389/fsoil.2021.765901 (2021).
-
Krishnamoorthy, R. et al. Arbuscular mycorrhizal fungi and associated bacteria isolated from salt-affected soil enhances the tolerance of maize to salinity in coastal reclamation soil. Agric. Ecosyst. Environ. 231, 233–239 (2016).
-
Wang, D. et al. Lateral root enriched Massilia associated with plant flowering in maize. Microbiome 12, 124 (2024).
-
Asiloglu, R. et al. Protist-enhanced survival of a plant growth promoting rhizobacteria, Azospirillum sp. B510, and the growth of rice (Oryza sativa L.) plants. Appl. Soil Ecol. 154, 103599 (2020).
-
Carvalhais, L. C., Dennis, P. G. & Schenk, P. M. Plant defence inducers rapidly influence the diversity of bacterial communities in a potting mix. Appl. Soil Ecol. 84, 1–5 (2014).
-
Xu, A., Liu, C., Zhao, S., Song, Z. & Sun, H. Dynamic distribution of Massilia spp. in sewage, substrate, plant rhizosphere/phyllosphere and air of constructed wetland ecosystem. Front. Microbiol. https://doi.org/10.3389/fmicb.2023.1211649 (2023).
-
Han, S. et al. Systemic responses of barley to the 3-hydroxy-decanoyl-homoserine lactone producing plant beneficial endophyte Acidovorax radicis N35. Front. Plant Sci. 7, 1868 (2016).
-
Siani, R., Stabl, G., Gutjahr, C., Schloter, M. & Radl, V. Acidovorax pan-genome reveals specific functional traits for plant beneficial and pathogenic plant-associations. Microb. Genom. 7, 000666 (2021).
-
Shi, P. et al. Multiple metabolic phenotypes as screening criteria are correlated with the plant growth-promoting ability of rhizobacterial isolates. Front. Microbiol. 12, 747982 (2022).
-
Vincze, É-B., Becze, A., Laslo, É & Mara, G. Beneficial soil microbiomes and their potential role in plant growth and soil fertility. Agriculture 14, 152 (2024).
-
Haida, M. et al. Microcystin-degrading bacteria reduce bioaccumulation in Fragaria vulgaris and enhance fruit yield and quality. Environ. Sci. Pollut. Res. Int. 31, 54502–54524 (2024).
-
Okazaki, K. et al. Community analysis-based screening of plant growth-promoting bacteria for sugar beet. Microbes Environ. 36, ME20137 (2021).
-
Li, Y. et al. Biofilms formation in plant growth-promoting bacteria for alleviating agro-environmental stress. Sci. Total Environ. 907, 167774 (2024).
-
Zhang, X. et al. Effects of different cultivation media on root bacterial community characteristics of greenhouse tomatoes. Front. Microbiol. 14, 1182347 (2023).
-
Huang, Z. et al. Bacterial inoculants improved the growth and nitrogen use efficiency of Pyrus betulifolia under nitrogen-limited conditions by affecting the native soil bacterial communities. Appl. Soil Ecol. 170, 104285 (2022).
-
Kämpfer, P., Ruppel, S. & Remus, R. Enterobacter Radicincitans sp. nov., a plant growth promoting species of the family Enterobacteriaceae. Syst. Appl. Microbiol. 28, 213–221 (2005).
-
Khalifa, A. Y., Alsyeeh, A.-M., Almalki, M. A. & Saleh, F. A. Characterization of the plant growth promoting bacterium, Enterobacter cloacae MSR1, isolated from roots of non-nodulating Medicago sativa. Saudi J. Biol. Sci. 23, 79 (2015).
-
Berendsen, R. L., Pieterse, C. M. J. & Bakker, P. A. H. M. The rhizosphere microbiome and plant health. Trends Plant Sci. 17, 478–486 (2012).
-
Mekonnen, H. & Kibret, M. The roles of plant growth promoting rhizobacteria in sustainable vegetable production in Ethiopia. Chem. Biol. Technol. Agric. 8, 15 (2021).
-
Posada, L. F. et al. Endophytic microbial diversity associated with commercial cultivar and crop wild relative banana variety could provide clues for microbial community management. Microbiol. Res. 287, 127862 (2024).
-
Cohen, R. A., Walker, K. & Carpenter, E. J. Polysaccharide addition effects on rhizosphere nitrogen fixation rates of the California cordgrass, Spartina foliosa. Wetlands 29, 1063–1069 (2009).
-
Zhu, H., Mo, Z., Wang, Y. & Su, J. The accumulation of polysaccharides in Dendrobium officinale is closely related to rhizosphere bacteria. Int. Microbiol. 28, 993–1004 (2024).
-
Mahmoud, Y. A.-G., El-Naggar, M. E., Abdel-Megeed, A. & El-Newehy, M. Recent advancements in microbial polysaccharides: synthesis and applications. Polymers 13, 4136 (2021).
-
Chen, E., Yang, C., Tao, W. & Li, S. Polysaccharides produced by plant growth-promoting rhizobacteria strain Burkholderia sp. BK01 enhance salt stress tolerance to Arabidopsis thaliana. Polymers 16, 145 (2024).
-
Saberi Riseh, R., Ebrahimi-Zarandi, M., Gholizadeh Vazvani, M. & Skorik, Y. A. Reducing drought stress in plants by encapsulating plant growth-promoting bacteria with polysaccharides. Int. J. Mol. Sci. 22, 12979 (2021).
-
Ghasemzadeh, Z. et al. The role of extracellular polysaccharide produced by Bradyrhizobium strain in root growth, improvement of soil aggregate stability and reduction of soil detachment capacity. Rhizosphere 27, 100771 (2023).
-
Peng, Y. et al. Pseudoxanthomonas sp. JBR18, a halotolerant endophytic bacterium, improves the salt tolerance of Arabidopsis seedlings. Plant Physiol. Biochem. 207, 108415 (2024).
-
Ulrich, K., Kube, M., Becker, R., Schneck, V. & Ulrich, A. Genomic analysis of the endophytic stenotrophomonas strain 169 reveals features related to plant-growth promotion and stress tolerance. Front. Microbiol. 12, 687463 (2021).
-
Liu, M. et al. Cyanobacteria blooms potentially enhance volatile organic compound (VOC) emissions from a eutrophic lake: field and experimental evidence. Environ. Res. 202, 111664 (2021).
-
Kultschar, B., Llewellyn, C., Kultschar, B. & Llewellyn, C. Secondary Metabolites in Cyanobacteria (IntechOpen, 2018).
-
Gademann, K. & Portmann, C. Secondary metabolites from cyanobacteria: complex structures and powerful bioactivities. Curr. Org. Chem. 12, 326–341 (2008).
-
du Jardin, P. Plant biostimulants: definition, concept, main categories and regulation. Sci. Hortic. 196, 3–14 (2015).
-
Lugtenberg, B. & Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 63, 541–556 (2009).
-
Fincheira, P. & Quiroz, A. Microbial volatiles as plant growth inducers. Microbiol. Res. 208, 63–75 (2018).
-
Rodríguez, A. A., Stella, A. M., Storni, M. M., Zulpa, G. & Zaccaro, M. C. Effects of cyanobacterial extracellular products and gibberellic acid on salinity tolerance in Oryza sativa L. Saline Syst. 2, 7 (2006).
-
Havaux, M. et al. Vitamin B6 deficient plants display increased sensitivity to high light and photo-oxidative stress. BMC Plant Biol. 9, 130 (2009).
-
Zeier, J. New insights into the regulation of plant immunity by amino acid metabolic pathways. Plant Cell Environ. 36, 2085–2103 (2013).
-
Rogan, C. J. et al. Transporter-mediated depletion of extracellular proline directly contributes to plant pattern-triggered immunity against a bacterial pathogen. Nat. Commun. 15, 7048 (2024).
-
Dai, L. et al. Effects of the supernatant of Chlorella vulgaris cultivated under different culture modes on lettuce (Lactuca sativa L.) growth. Front. Nutr. 11, 1437374 (2024).
-
Jwa, E. et al. Recycling of nutrient medium to improve productivity in large-scale microalgal culture using a hybrid electrochemical water treatment system. Water Res. 246, 120683 (2023).
-
Christenson, L. & Sims, R. Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts. Biotechnol. Adv. 29, 686–702 (2011).
-
Arora, N., Lo, E., Legall, N. & Philippidis, G. P. A Critical review of growth media recycling to enhance the economics and sustainability of algae cultivation. Energies 16, 5378 (2023).
-
Pereira, S. et al. Complexity of cyanobacterial exopolysaccharides: composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly. FEMS Microbiol. Rev. 33, 917–941 (2009).
-
Augšpole, I., Sivicka, I., Kampuss, K., Semjonovs, P. & Missa, I. Biologically active compounds in tomato fruits under the application of water–ethanol spirulina, dunaliella and chlorella microalgae extracts on plants’ leaves. Int. J. Plant Biol. 15, 1338–1352 (2024).
-
Mostafa, M. M., Hammad, D. M., Reda, M. M. & El-Sayed, A. E.-K. B. Water extracts of Spirulina platensis and Chlorella vulgaris enhance tomato (Solanum lycopersicum L) tolerance against saline water irrigation. Biomass. Convers. Biorefinery 14, 21059–21068 (2024).
-
Gitau, M. M., Farkas, A., Ördög, V. & Maróti, G. Evaluation of the biostimulant effects of two Chlorophyta microalgae on tomato (Solanum lycopersicum). J. Clean. Prod. 364, 132689 (2022).
-
Rachidi, F., Benhima, R., Kasmi, Y., Sbabou, L. & Arroussi, H. E. Evaluation of microalgae polysaccharides as biostimulants of tomato plant defense using metabolomics and biochemical approaches. Sci. Rep. 11, 930 (2021).
-
Ali, O., Ramsubhag, A. & Jayaraman, J. Biostimulant properties of seaweed extracts in plants: implications towards sustainable crop production. Plants 10, 531 (2021).
-
Adedayo, A. A. & Babalola, O. O. The potential of biostimulants on soil microbial community: a review. Front. Indus. Microbiol. https://doi.org/10.3389/finmi.2023.1308641 (2023).
-
Cogne, G., Lehmann, B., Dussap, C.-G. & Gros, J.-B. Uptake of macrominerals and trace elements by the cyanobacterium Spirulina platensis (Arthrospira platensis PCC 8005) under photoautotrophic conditions: culture medium optimization. Biotechnol. Bioeng. 81, 588–593 (2003).
-
Rachidi, F., Benhima, R., Sbabou, L. & Arroussi, H. E. Microalgae polysaccharides bio-stimulating effect on tomato plants: growth and metabolic distribution. Biotechnol. Rep. 25, e00426 (2020).
-
Jamur, M. C. & Oliver, C. Permeabilization of cell membranes. In: Immunocytochemical Methods and Protocols (eds. Oliver, C. & Jamur, M. C.) 63–66 (Humana Press, Totowa, 2010).
-
DuBois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A. & Smith, F. Colorimetric method for determination of sugars and related substances. Nature. https://doi.org/10.1021/ac60111a017 (1956).
-
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
-
Cataldo, D. A., Maroon, M., Schrader, L. E. & Youngs, V. L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 6, 71–80 (1975).
-
Tilly, N., Aasen, H. & Bareth, G. Fusion of plant height and vegetation indices for the estimation of barley biomass. Remote Sens. 7, 11449–11480 (2015).
-
Vanhee, M. et al. Implementation of full-length 16S nanopore sequencing for bacterial identification in a clinical diagnostic setting. Diagn. Microbiol. Infect. Dis. 108, 116156 (2024).
-
Heuer, H., Krsek, M., Baker, P., Smalla, K. & Wellington, E. M. Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 63, 3233–3241 (1997).
-
Redelings, B. D. & Holder, M. T. Taxonomic supertree construction with incertae sedis taxa. In: Bioinformatics and Phylogenetics: Seminal Contributions of Bernard Moret (ed. Warnow, T.) 151–173 (Springer International Publishing, Cham, 2019).
-
Delacuvellerie, A., Cyriaque, V., Gobert, S., Benali, S. & Wattiez, R. The plastisphere in marine ecosystem hosts potential specific microbial degraders including Alcanivorax borkumensis as a key player for the low-density polyethylene degradation. J. Hazard. Mater. 380, 120899 (2019).
-
Hammer, O. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron 4, 9 (2001).
-
Delacuvellerie, A. et al. Microbial biofilm composition and polymer degradation of compostable and non-compostable plastics immersed in the marine environment. J. Hazard. Mater. 419, 126526 (2021).
-
Ritchie, M. E. et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015).
-
Wang, Y., Naumann, U., Wright, S. T. & Warton, D. I. Mvabund—an R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 3, 471–474 (2012).
-
Krzywinski, M. et al. Circos: an information aesthetic for comparative genomics. Genome Res. 19, 1639–1645 (2009).
-
Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 14, 927–930 (2003).
-
Kim, I., Nguyen, M. H., Lee, S., Choi, B. & Shin, K. Variation of soil bacterial communities in forest soil contaminated with chainsaw lubricants. Microorganisms 12, 508 (2024).
-
Thrash, J. C., Pollock, J., Torok, T. & Coates, J. D. Description of the novel perchlorate-reducing bacteria Dechlorobacter hydrogenophilus gen. nov., sp. nov. and Propionivibrio militaris, sp. nov. Appl. Microbiol. Biotechnol. 86, 335 (2009).
-
Demin, K. A., Prazdnova, E. V., Minkina, T. M. & Gorovtsov, A. V. Sulfate-reducing bacteria unearthed: ecological functions of the diverse prokaryotic group in terrestrial environments. Appl. Environ. Microbiol. 90, e01390–23 (2024).
-
Mingo, F. S., Diekert, G. & Studenik, S. Enrichment of Desulfitobacterium spp. from forest and grassland soil using the O-demethylation of phenyl methyl ethers as a growth-selective process. Microbiology 162, 224–235 (2016).
-
Sitte, J. et al. Microbial links between sulfate reduction and metal retention in uranium- and heavy metal-contaminated soil. Appl. Environ. Microbiol. 76, 3143–3152 (2010).
-
Khoso, M. A. et al. Bacillus altitudinis AD13-4 enhances saline–alkali stress tolerance of alfalfa and affects composition of rhizosphere soil microbial community. Int. J. Mol. Sci. 25, 5785 (2024).
-
Quijia Pillajo, J., Chapin, L. J., Quiroz-Moreno, C. D., Altland, J. E. & Jones, M. L. Nutrient availability and plant phenological stage influence the substrate microbiome in container-grown Impatiens walleriana ‘Xtreme Red’. BMC Plant Biol. 24, 176 (2024).
-
Ravin, N. V. et al. Genome analysis of fimbriiglobus ruber SP5T, a planctomycete with confirmed chitinolytic capability. Appl. Environ. Microbiol. 84, e02645–17 (2018).
-
Stegelmeier, A. A., Rose, D. M., Joris, B. R. & Glick, B. R. The use of PGPB to promote plant hydroponic growth. Plants 11, 2783 (2022).
-
Mohite, J. A. et al. Exploring the potential of methanotrophs for plant growth promotion in rice agriculture. Methane 2, 361–371 (2023).
-
Chen, M.-H. et al. Aliidongia dinghuensis gen. nov., sp. nov., a poly-β-hydroxybutyrate-producing bacterium isolated from Pinus massoniana forest soil. Int. J. Syst. Evolut. Microbiol. 67, 212–217 (2017).
-
Dedysh, S. N., Beletsky, A. V., Kulichevskaya, I. S., Mardanov, A. V. & Ravin, N. V. Complete genome sequence of paludibaculum fermentans P105T, a facultatively anaerobic acidobacterium capable of dissimilatory Fe(III) reduction. Microbiol. Resour. Announc. 10, 10.1128/mra.01313–20 (2021).
-
Jacobs-Hoffman, I. & Hills, P. N. Effects of the commercial biostimulant BC204 on the rhizosphere microbial community of Solanum lycopersicum L. South Afr. J. Bot. 143, 52–60 (2021).
-
Ganesh, J., Singh, V., Hewitt, K. & Kaundal, A. Exploration of the rhizosphere microbiome of native plant Ceanothus velutinus—an excellent resource of plant growth-promoting bacteria. Front. Plant Sci. 13, 979069 (2022).
-
Salazar-Garcia, G., Balaguera-Lopez, H. E. & Hernandez, J. P. Effect of plant growth-promoting bacteria Azospirillum brasilense on the physiology of radish (Raphanus sativus L.) under waterlogging stress. Agronomy 12, 726 (2022).
-
Li, C.-H. et al. Biocontrol of verticillium wilt and colonization of cotton plants by an endophytic bacterial isolate. J. Appl. Microbiol. 113, 641–651 (2012).
-
Li, C.-H., Zhao, M.-W., Tang, C.-M. & Li, S.-P. Population dynamics and identification of endophytic bacteria antagonistic toward plant-pathogenic fungi in cotton root. Microb. Ecol. 59, 344–356 (2010).
-
Liu, G.-H. et al. Nitrogen fixation and transcriptome of a new diazotrophic geomonas from paddy soils. mBio 14, e02150–23 (2023).
-
Jeon, B. J. et al. Plant growth-promoting effects of a novel Lelliottia sp. JS-SCA-14 and comparative genome analysis. Front. Plant Sci. 15, 1484616 (2024).
-
Ulrich, K. et al. Physiological and genomic characterisation of Luteimonas fraxinea sp. nov., a bacterial species associated with trees tolerant to ash dieback. Syst. Appl. Microbiol. 45, 126333 (2022).
-
Zhang, D.-C. et al. Luteimonas terricola sp. nov., a psychrophilic bacterium isolated from soil. Int. J. Syst. Evolut. Microbiol. 60, 1581–1584 (2010).
-
Garcia Teijeiro, R., Belimov, A. A. & Dodd, I. C. Microbial inoculum development for ameliorating crop drought stress: a case study of Variovorax paradoxus 5C-2. N. Biotechnol. 56, 103–113 (2020).
-
Yim, B., Winkelmann, T., Ding, G.-C. & Smalla, K. Different bacterial communities in heat and gamma irradiation treated replant disease soils revealed by 16S rRNA gene analysis—Contribution to improved aboveground apple plant growth? Front. Microbiol. 6, 1224 (2015).
Acknowledgements
This article has been made possible through the author’s involvement in the MELiSSA project, ESA’s life support system program (https://www.esa.int/Enabling_Support/Space_Engineering_Technology/Melissa), and through the ARTEMiSS project (supported by ESA/PRODEX and BELSPO), providing the PhD grants of Cécile Renaud. This work was made possible through the Projet Phenix Biocontrol, funded by the European Regional Development Fund (FEDER) and the Walloon Region (RW). The Bioprofiling platform used for monosaccharides analysis was supported by the European Regional Development Fund and the Walloon Region, Belgium.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Renaud, C., Delacuvellerie, A., Largeteau, P. et al. Biostimulatory effects of Limnospira indica on the modulation of rhizosphere and endosphere microbiota and its impact on plant growth. npj Sustain. Agric. 4, 7 (2026). https://doi.org/10.1038/s44264-025-00115-8
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s44264-025-00115-8