- Research
- Open access
- Published:
- Pramod Sivan1,
- Deepika Dahiya2,
- Ylenia Jabalera3,
- Taru Koitto2,
- Raul Perez-Jimenez3,
- Ewa J. Mellerowicz4,
- Emma Master2,6 &
- …
- Francisco Vilaplana1,5
Biotechnology for Biofuels and Bioproducts volume 18, Article number: 70 (2025) Cite this article
Abstract
Background
Overcoming lignocellulose recalcitrance to enzymatic or chemical processing is a prerequisite for biorefinery applications. Expansins and loosenins are non-lytic proteins that could assist reducing this recalcitrance by disrupting the intermolecular contacts between plant cell wall components. Here, immunolocalization with fluorescence and transmission electron microscopy (TEM) were used to study the ability of a Bacillus subtilis expansin-like protein (BsEXLX1), a Phanerochaete carnosa loosenin protein (PcaLOOL12) and a fusion protein of PcaLOOL12 with the carbohydrate-binding module 63 (CBM63) of BsEXLX1 (i.e., PcaLOOL12-CBM63) to bind secondary cell walls (SCW) of aspen fibres, including fresh aspen wood, milled wood fibres (MWF) and MWF subjected to subcritical water extraction.
Results
The immunofluorescence labelling of fresh wood samples showed a weak signal for PcaLOOL12 and a strong signal for BsEXLX1 and PcaLOOL12-CBM63, suggesting the importance of CBM63 for protein adsorption to SCW components. TEM analysis after immunogold labelling revealed the presence of BsEXLX1 and PcaLOOL12-CBM63 in all secondary cell wall layers. Pretreatment of wood samples with the proteins reduced the binding of glucomannan- and glucuronoxylan-specific monoclonal antibodies. Similarly, protein adsorption to MWF was higher before subcritical water extraction. Together, these results suggest the adsorption of BsEXLX1 and PcaLOOL12-CBM63 to SCWs was mediated at least in part by their interaction with hemicelluloses.
Conclusions
Our study demonstrates that microbial expansin-related proteins can bind to the secondary walls of aspen wood through potential interaction of CBM63 with hemicelluloses.
Introduction
Cell growth in plants involves changes in the structural organization of the polymeric network of cellulose and matrix polysaccharides in primary cell walls [1, 2]. The family of α-expansin (EXPA) proteins are involved in acid-induced cell wall loosening and are responsible for structural changes in the primary cell wall crucial for wood formation [3, 4]. The related β-expansin protein family (EXPB), which include grass pollen allergens, are known to facilitate intracellular pollen tube invasion, whereas two other plant expansin subfamilies, expansin-like A (EXLA) and expansin-like B (EXLB) have not been so far functionally analysed [5]. Xylem cell growth is followed by the deposition of a thick multilayered secondary wall (SCW), characterized by aligned cellulose microfibrils embedded in a complex matrix of glucuronoxylan and glucomannan hemicelluloses, which become impregnated with polyphenolic lignin together with surrounding primary cell walls. This complex structure and chemistry of wood cell walls contribute to the highly recalcitrant properties of lignocellulose biomass against its degradation.
Microbial enzymes from wood-degrading microorganisms are well known for their ability to disassemble cell walls. They include glycoside hydrolases (cellulases, xylanases, mannanases, pectinases), polysaccharide lyases (pectate lyases, rhamnogalacturonan lyases), oxidoreductases (lytic polysaccharide monooxygenases, LPMOs), and ligninases (laccases, peroxidases, polyphenol oxidases) that can disintegrate the cell wall causing the decomposition of lignocellulosic biomass [6]. These enzymes are widely used for commercial applications requiring saccharification of complex cell wall polymers into monomers [7]. However, the potential application of lignocellulosic biomass components into functional materials require their modification without complete disassembly. In this regard, microbial expansin-related proteins appear as promising biotechnological tools to disrupt the assembly of cellulosic fibres through biophysical effects, leading to an increased accessibility of catalytic enzymes to the glycan substrates during the biochemical conversion of biomass [8,9,10].
Microbial expansin-like proteins (EXLXs) have been identified in plant pathogens and saprotrophs suggesting their role in plant biomass decomposition [8]. Structurally, EXLXs comprise an N-terminal six-stranded double-Ѱ beta-barrel (DPBB) domain (D1) homologous to GH45 glycoside hydrolase family (GH) and a C-terminal domain (D2) that belongs to type-A CBM family 63 [11]. Other expansin-related proteins can have additional domains, such as a family 2 carbohydrate-binding module (CBM2), family 5 glycoside hydrolase (GH5) domain [12, 13], or GH9 domain [14]. Similarly, swollenins retain the core EXLX structure but have an N-terminal CBM1 (a type-A CBM) and fibronectin III insertion [15]. By contrast, microbial expansin-related proteins that lack the C-terminal CBM63 are classified as loosenins and ceratoplatanins [16,17,18].
The biophysical mechanisms responsible for the relaxation of plant cell walls by expansins involve the disruption of noncovalent interactions between cellulose microfibrils and hemicelluloses, or between tight junctions between neighbouring microfibrils, leading to cellulose microfibril slippage causing cell wall creep [19, 20]. The ability of Bacillus subtilis expansin BsEXLX1 to induce extension of alkali-pretreated wheat coleoptiles and weaken filter paper was demonstrated, and subsequent mutations in the CBM63 domain demonstrated this domain was necessary for creep through selective interaction of the CBM63 with cellulose microfibrils [21]. Although lacking a CBM63, recombinantly produced loosenins from Phanerochaete carnosa (PcaLOOL) were shown to boost the enzymatic conversion of wood pulps and preparation of cellulose nanocrystals [22, 23], and increase the interfibril distances of wood-derived holocelluloses [24]. These studies on artificially modified substrates such as filter paper and wood pulps raised the question regarding the ability of microbial expansin-like proteins and loosenins to interact with cell wall polymers in native cell walls from hardwood trees. This study aims to evaluate the adsorption of expansin BsEXLX1, loosenin PcaLOOL12, and a fusion of PcaLOOL12 with CBM63 from BsEXLX1, to cell wall components of woody tissues from hybrid aspen using advanced microscopic techniques.
Materials and methods
Plant material
The fresh wood samples were collected from hybrid aspen (Populus tremula L. × tremuloides Michx. clone T89) trees grown for 9 weeks in the phenotyping platform (WIWAM Conveyor, custom designed by SMO, Eeklo, Belgium) [25]. To generate wood samples with partial removal of hemicelluloses, milled dry wood samples of hybrid aspen were sieved to 100–500 mm particle size and 1 g of sieved wood powder was subjected to subcritical water extraction (SWE) at 170 °C with buffered pH 5 for 60 min with an accelerated solvent extractor (ASE-300, Dionex, USA), according to method described elsewhere [26]. The non-extracted wood powder was used as control.
Chemical composition analysis of wood powder
The aspen wood powder and the residue after subcritical water extraction were subjected to monosaccharide analysis by a two-step sulfuric acid hydrolysis [27]. In brief, 1 mg of sample was incubated with 125 μL of 72% H2SO4 at room temperature for 3 h, then diluted with 1375 μL of deionized water, and further incubated at 100 °C for 3 h. The hydrolysates were diluted 10 times with MilliQ water, filtered through a 0.2-mm syringe filter (Chromacol 17-SF-02-N) and analysed by high-performance anion exchange chromatography with pulsed amperometric detection (HPAEC-PAD) (ICS-6000 DC, Dionex) equipped with a CarboPac PA1 column (4 × 250 mm, Dionex) at 30 °C using the eluent gradients previously reported [28]. Monosaccharide quantification was performed by standard calibration (Ara, Rha, Fuc, Xyl, Man, Gal, Glc, GalA, MeGlcA and GlcA). The Klason lignin content was estimated from the gravimetric calculation after acid hydrolysis.
Microbial expansin and loosenin cloning and production
The expansin-like protein from Bacillus subtilis (BsEXLX1, GenBank accession no. WP_003231419.1) was recombinantly produced in Escherichia coli strain BL21 (DE3) from the codon-optimized gene subcloned to pET21a( +) plasmid. For protein expression, cells were incubated in LB medium at 37 °C until the optical density (OD600) reached 0.6, and 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) was added for protein induction overnight. The cells were pelleted by centrifugation at 6000 g for 20 min, the pellets were resuspended in lysis buffer (50 mM Tris–HCl, NaCl 200 mM, pH 7.8, Halt Protease Inhibitor, Invitrogen), and sonicated at 30% amplitude for 5 min. The cell debris was separated by ultracentrifugation at 33,000 g for 1 h, and the supernatants were mixed with His GraviTrap affinity column (GE Healthcare), washed with buffer supplemented with 20 mM imidazole, and eluted with an elution buffer (50 mM Tris–HCl, NaCl 200 mM, pH 7.8, 300 mM imidazole). The proteins were dialyzed to remove imidazole and sodium dodecyl sulphate and the protein purification was verified by polyacrylamide gel electrophoresis (SDS-PAGE). The final protein concentration was calculated by measuring the absorbance at 280 nm in a Nanodrop 2000C.
The loosenin-like protein from Phanerochaete carnosa, PcaLOOL12 (GenBank code EKM51974.1) [29] was recombinantly produced in Komagataella phaffii strain SMD1168H in accordance with the manufacturer’s instructions (Invitrogen, Thermo Fisher Scientific) and as previously described [18]. The PcaLOOL12-CBM63 fusion protein comprised a PcaLOOL12 at the N-terminus and CBM63 from BsEXLX1 at the C-terminus of the protein. The two domains were connected by the linker sequence taken from BsEXLX1, including two last amino acids from BsEXLX1 domain 1 [18]. PcaLOOL12–CBM63 was recombinantly produced in K. phaffii as previously reported [18]. All the three recombinant proteins were purified with a His-tag on their C-terminal ends that served as a target for immunolabeling experiments.
Expansin and loosenin treatment of woody biomass
The fresh wood samples from the 38th and 39th internodes were cut into blocks of 2 × 2 × 10 mm (radial, tangential and longitudinal) size. The proteins were diluted to 1 mg mL−1 using sodium acetate buffer (pH 5.0 for BsEXLX1 and pH 6.0 for PcaLOOL12 and PcaLOOL12–CBM63). Between 5 and 6 wood blocks having a total weight of 6–7 mg or 6 mg of SWE residues and wood powder were transferred to 1.5 mL Eppendorf tubes containing microbial proteins loaded at 2% (w/w) and incubated at 25 °C for 48 h. Each treatment was carried out in triplicate.
Microscopic analysis of treated biomass
After incubation with recombinant proteins, the samples were briefly washed in buffer and fixed in a mixture of 0.1% glutaraldehyde and 4% paraformaldehyde in 50 mM sodium cacodylate buffer for 4 h at room temperature and left overnight at 4 °C. After fixation, the SWE residue and wood powder samples were embedded in 4% agarose [30], and the solidified agarose was cut into 3 × 4 × 3 mm cubes. Both wood blocks and agarose cubes were then washed in phosphate buffer (pH 7.0), dehydrated in graded series of ethanol for 20 min from 30 to 95% (15 min each) to pure ethanol (× 3) and embedded in LR white resin as described elsewhere [31].
For bright field and immunofluorescence microscopy, 2-µm-thick transverse sections were prepared from LR white embedded blocks with a diamond knife. For general histological evaluation of the wood tissues, semithin sections were stained with toluidine blue O (0.5% aqueous). To confirm the protein interaction into deeper areas, these sections were taken after removing of 0.2-mm-thick surface layer from the blocks. Sections were mounted on formvar-coated slides and treated with 50 mM glycine/phosphate buffered saline (PBS) solution for 15 min. The sections were washed with PBS buffer and suspended in blocking buffer (PBS containing 3% skim milk) for 30 min at room temperature. Sections were incubated in anti-His-tag monoclonal antibodies (6x-His Tag monoclonal antibody, Alexa Flour 488, Thermo Scientific, USA, 1:100 dilution in PBS buffer) for 1 h at room temperature. After washing in PBS buffer, the sections were mounted in Fluoroshield (Sigma, Germany) on a clean glass slide and were examined using a Leica DMi8 inverted microscope (Leica Biosystems, Germany) fitted using 499 nm excitation and 520 nm emission. Immunofluorescence negative control sections were processed with omission of the antibody.
For immunogold labelling, ultrathin sections of 90 nm thickness obtained using ultramicrotome (Reichert Ultracut S, Leica, Austria) were mounted on copper grids, which were suspended in buffer A (PBS containing 1% bovine serum albumin and 0.1% NaN3, pH 8.2) for 30 min at room temperature. For the recombinant protein localization, the grids were incubated with anti-His-Tag antibody with 10 nm colloidal gold (10 nm Ni-NTA-Nanogold, Nanoprobes, USA) for 45 min, washed in three changes in buffer A and MilliQ water, and dried. For localization of glucuronoxylan and glucomannan, the grids were incubated with LM10 (for glucuronoxylan) and LM21 (glucomannan) monoclonal antibodies obtained from Plant Probes (UK) diluted 1:20 (V:V) in buffer A for 45 min. After three washings with buffer A for 5 min each, the grids were incubated with goat anti-rat secondary antibody labelled with 10-nm colloidal gold particles (BB International, UK) for 1 h at room temperature. For the negative immunolocalization control, the sections were processed with omission of primary antibodies. Finally, the grids were washed in six changes of buffer A for 15 min each, followed by washing with distilled water. All labelled sections were post-stained with 5% uranyl acetate for 30 min, washed in running distilled water for 1 min. All sections were examined under a transmission electron microscope (TEM, FEI TALOS L120C) at an accelerating voltage of 100 kV. The gold particle density quantification (per mm2 area) after LM10 and LM21 labelling was based on 20 measurements from five images per each triplicated sample. All measurements were taken using ImageJ software (National Institutes of Health, USA).
Statistical analysis
Statistical analyses were performed in JMP Pro (v.16.0) software (SAS Institute Inc., Cary, NC, USA).
Results
Immunolocalization of BsEXLX1, PcaLOOL12 and PcaLOOL12-CBM63 in the cell walls of wood fibres
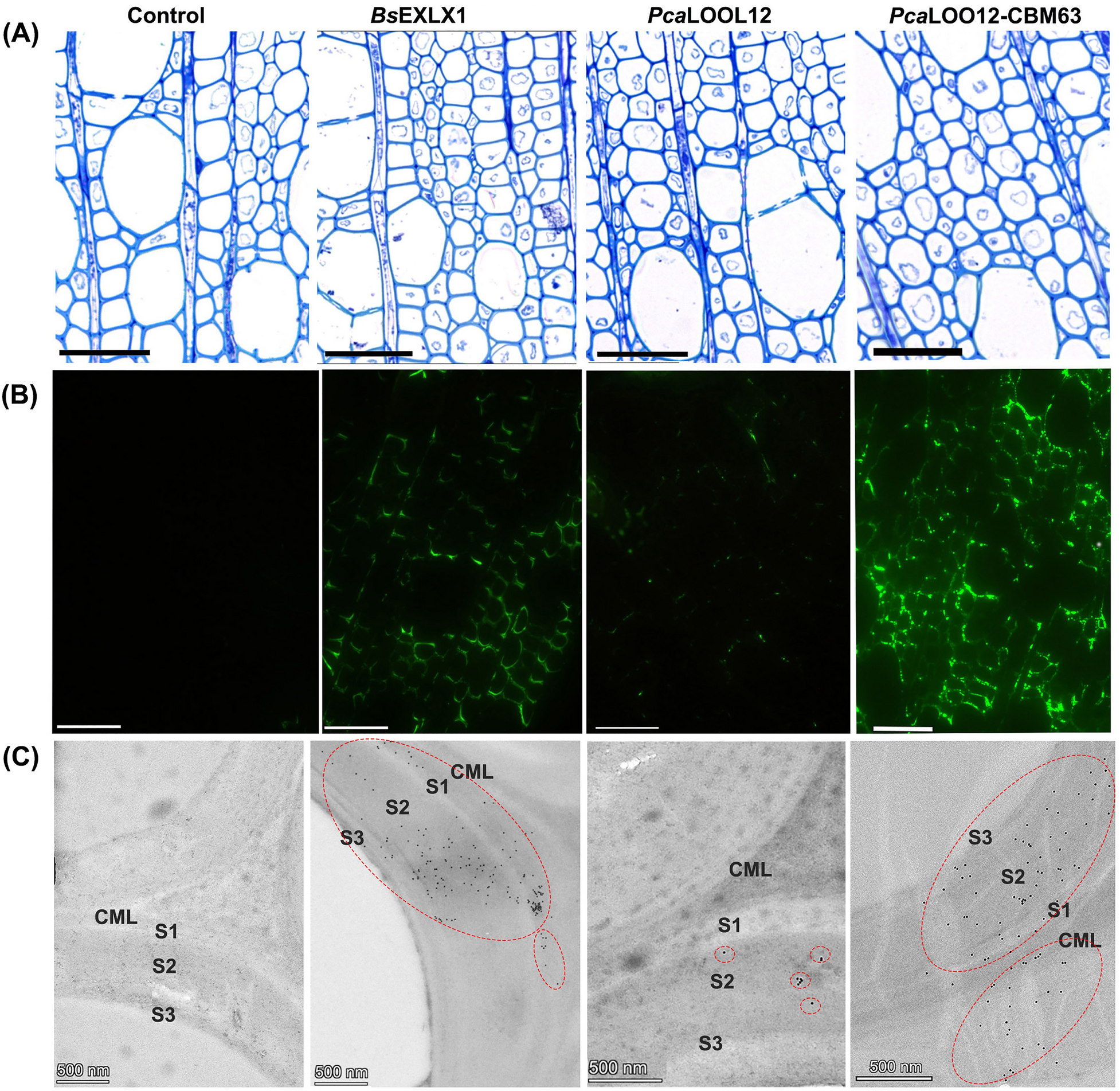
To evaluate the interaction of the microbial expansin-like (BsEXLX1), loosenin (PcaLOOL12) and PcaLOOL12-CBM63 fusion proteins with lignified wood cell walls, the anatomy of developing wood was first examined using TBO-stained semithin wood sections (Fig. 1A). The selected wood areas containing secondary wall-thickened and lignified wood fibres were subjected to immunolabeling experiments. Strong fluorescence signals from the fibre cell walls of BsEXLX1–treated samples in the selected wood zone area indicated the ability of the protein to interact with the lignified wood cell walls (Fig. 1B). For wood blocks treated with PcaLOOL12, a weak fluorescence signal was observed (Fig. 1B). By contrast, the samples treated with PcaLOOL12-CBM63 showed strong labelling. The differences in labelling among the three proteins were confirmed by transmission electron microscopy after immunogold labelling. A strong gold labelling for BsEXLX1 and PcaLOOL12-CBM63 was evident in the secondary fibre wall, while a weaker immunogold labelling was observed in the PcaLOOL12 treated wood (Fig. 1C). These results demonstrate the strong adsorption of BsEXLX1 and PcaLOOL12-CBM63 fusion protein into the fibre secondary walls, since the signals were not washed out during immunolabeling procedure. The signals from both BsEXLX1 and PcaLOOL12-CBM63 become more apparent in the secondary wall layers (S1, S2 and S3) compared to the compound middle lamellae (CML) (Fig. 1C). A reduced labelling in the CML could be due to a hindered binding of the proteins due to the higher lignin content and/or the different composition of CML versus secondary wall layers of hardwood fibres [32].

Light microscopy (A), immunofluorescence microscopy (B) and immunogold-transmission electron microscopy (C) images of transverse sections from fresh aspen wood samples treated with buffer (control), BsEXLX1, PcaLOOL12 and PcaLOOL12-CBM63 proteins. Sections subjected to toluidine blue staining for general histology (A) and immunolabeling with anti-His antibody for recombinant protein localization within cell wall (B, C). Scale bar = 50 um (A, B), 500 nm (C). Compound middle lamellae (CML), secondary wall layers (S1, S2 and S3). Red dotted circles indicate areas of dense gold particle distribution
Immunolocalization of hemicelluloses in the microbial protein-treated aspen wood fibres
The wood cell wall is characterized by spatial and temporal variation in the distribution of hemicelluloses and pectins. Therefore, to validate the possible relation between hemicellulose distribution and protein binding, we investigated the variation in the spatial distribution pattern of hemicelluloses in the secondary wall of aspen wood fibres. The localization of glucuronoxylan using LM10 antibody in a control tissue that was not incubated with microbial proteins revealed strong labelling in all the secondary wall layers (Fig. 2A), suggesting a homogenous distribution of glucuronoxylan in the secondary wall. A decrease in the gold particle distribution was observed in the S2 and S3 layers of BsEXLX1 and PcaLOOL12-CBM63 treated samples (Fig. 2A, B).

A Transmission electron microscopy (TEM) images from the transverse sections of control and protein-treated aspen wood, labelled with LM10 antibody for glucuronoxylan and LM21 antibody for glucomannan. B Density of gold particles (per mm2 area) in the control and protein-treated wood fibres using LM10 and LM21 labelling. *P ≤ 0.05; **P ≤ 0.01; ***P ≤ 0.001 for comparisons with control by Dunnett´s test. Red dotted circles indicate areas of dense gold particle distribution
The localization of glucomannan using LM21 antibody revealed relatively low labelling in S1 layer while S2 and S3 layer showed a higher density of gold particles, suggesting that glucomannans have certain spatial heterogeneity in the distribution pattern and/or in epitope exposure within the secondary wall layers of aspen wood fibres (Fig. 2A). The immunogold particle density measurements of LM21 labelling in control tissue (without protein treatment) was very low compared to LM10 labelling suggesting that glucuronoxylan is more abundant compared to glucomannan in the secondary wall of aspen wood fibres (Fig. 2A, B). Nevertheless, the LM21 labelling was clearly diminished following treatment with BsEXLX1 and PcaLOOL12-CBM63 treated samples (Fig. 2F–H).
The decreased LM10 and LM21 signals from protein-treated samples compared to controls indicate that these proteins mask the glucuronoxylan and glucomannan epitopes. Our results suggest that the CBM63 facilitates binding between these proteins and hemicelluloses, and such interaction possibly restricts the glucuronoxylan and glucomannan recognition by LM10 and LM21 antibodies.
Protein localization in wood fibres after partial removal of hemicelluloses
The immunolabeling patterns described above suggest that the main interactions between the proteins and the hemicelluloses in aspen were mediated by the CBM63. To evaluate this hypothesis, we partially removed the hemicelluloses from the aspen wood powder using subcritical water extraction. SWE also removes a minor amount of extractable lignin, potentially in the form of lignin–carbohydrate complexes (LCCs), without largely affecting the cellulose and lignin content in the SWE residue [26, 33]. The chemical analysis of the SWE residue compared to the starting material revealed a 30–50% reduction in the monosaccharides attributed to glucomannan and glucuronoxylan (Fig. 3A). This decrease in glucuronoxylan and glucomannan was also confirmed by the decrease in immunolocalization signals and gold particle density of LM10 and LM21 in the SWE residue (Fig. 3B, C).

A Chemical composition of aspen wood (AW) and residue (R) of subcritical water extraction. B Transverse sections from fibres in the aspen wood and residue labelled for glucuronoxylan with LM10 antibody and glucomannan with LM21 antibody. C Density of gold particles (per mm2 area) for LM10 and LM21 labelling in the aspen wood and residue of subcritical water extraction. ***P ≤ 0.001 for comparisons with AW by Student’s t-test. Red dotted circles indicate areas of dense gold particle distribution. Red dotted circles indicate areas of dense gold particle distribution
To evaluate the effect of hemicellulose removal on the binding ability of the recombinant proteins, immunogold localization was performed in the protein-treated wood powder (starting material) and SWE residue entrapped into agarose for easy handling during processing and embedment in LR white resin. As observed for the fresh wood samples, the protein-treated wood powder (without SWE extraction) showed a strong labelling for BsEXLX1 and the fusion protein PcaLOOL12-CBM63 but not for wild-type PcaLOOL12 (Fig. 4). After hemicellulose extraction, however, weak to no protein labelling was observed in the SWE residue (Fig. 4). These results support the hypothesis that hemicelluloses are needed for CBM63-mediated binding of BsEXLX1 and the fusion protein PcaLOOL12-CBM63 to lignified secondary walls of aspen wood fibre.

Ultrathin sections of agarose-embedded, recombinant protein (BsEXLX1, PcaLOOL12 and PcaLOOL12-CBM63)-treated wood powder and SWE residue, immunogold labelled with anti-His tag antibody. Compound middle lamellae (CML), secondary wall layers 1 (S1), 2 (S2) and 3(S3). Red dotted circles indicate areas of dense gold particle distribution
Discussion
So far, in situ studies of BsEXLX1 and other expansin-related proteins have focused on primary cell wall tissues. On the other hand, secondary cell walls found in trees represent the most abundant carbon sink and source of renewable materials for biorefinery applications. In hardwoods, the secondary wall polysaccharide matrix is composed of glucuronoxylan and glucomannans; however, to our knowledge there have been no reports on the possible roles of these components in expansin-like protein binding. Our study now provides experimental evidence for the role of hardwood hemicelluloses (glucuronoxylan and glucomannan) as binding sites for CBM63 in BsEXLX1 and the PcaLOOL12-CBM63 fusion protein in the secondary cell walls of aspen wood.
Consistent with earlier studies that demonstrate the importance of CBM63 to BsEXLX1 binding to wheat coleoptile primary cell walls [21], the immunolocalization studies reported herein showed that appending the CBM63 to the C-terminus of PcaLOOL12 considerably increased loosenin binding to secondary cell walls of aspen wood. The immunolocalization studies also indicated a possible effect of the hemicellulose composition on the binding of CBM63-containing proteins to the aspen samples. For example, the density of immunogold-labelled BsEXLX1 and PcaLOOL12-CBM63 was higher in secondary cell wall layers comprising glucuronoxylan and glucomannan compared to the CML region, which is characterized by comparatively high lignin and pectin contents as well as xyloglucan as the main hemicellulose [32]. Moreover, the labelling signal of LM10 and LM21 antibodies towards glucuronoxylan and glucomannan, respectively, decreased following pretreatment of the aspen samples with the CBM63-containing proteins, presumably through protein binding and masking the hemicellulose epitopes recognized by these antibodies. Consistent with this interpretation, partial hemicellulose removal by SWE substantially reduced the protein labelling signal observed in subsequently treated wood samples. Since SWE releases hemicelluloses from wood without substantially affecting lignin and cellulose contents [26, 33], this result also suggests that BsEXLX1 and PcaLOOL12-CBM63 preferentially bind hemicelluloses over cellulose and lignin. Binding of BsEXLX1 to hemicelluloses over other cell wall components has been observed using extracted wheat coleoptile primary cell walls [21], solid-state NMR analyses of Arabidopsis thaliana cell walls showing BsEXLX1 binding to cellulose in close proximity to xyloglucan [34], as well as commercial sources of xylan, lignin and cellulose [35, 36]. The importance of glucuronoxylan and glucomannan to BsEXLX1 and PcaLOOL12-CBM63 binding to the aspen wood samples now indicate these hemicelluloses could mediate expansin-related protein binding to the secondary cell walls of hardwood fibres.
Besides substrate selectivity, the depth of BsEXLX1 and PcaLOOL12-CMB63 penetration into the aspen wood samples was investigated. Earlier immunolocalization studies of BsEXLX1 in 0.5 mm-thick sections of celery petioles show strong surface binding of BsEXLX1 to different cell types of xylem and phloem [37]. Herein, the wood sections were considerably thicker (2 µm) and strong immunolabeling signals from BsEXLX1 and PcaLOOL12-CMB63 were observed 0.2 mm from the exposed sample surface. Whereas the tracheary elements in the primary xylem celery petioles are characterized by secondary walls in the form of spiral, annular and helical thickening in restricted cell wall areas [38], the tracheary elements in secondary xylem of higher woody plants have more complex pitted structure. Our study demonstrated that BsEXLX1 and PcaLOOL12-CBM63 are able to bind within the structurally complex secondary wall of hardwood fibres.
In conclusion, immunolocalization methods were used herein to evaluate the interaction of microbial expansin-like protein BsEXLX1 and loosenin PcaLOOL12, wild-type or fused with CBM63 (PcaLOOL12-CBM63), with wood fibres. We demonstrated the role of CBM63 in enhancing the binding of PcaLOOL12 to secondary cell wall components of aspen wood fibres. The masking effect of the proteins on glucuronoxylan and glucomannan labelling, and the weak immunolabeling of these proteins in wood fibres subjected to partial hemicellulose removal, suggested that CBM63-mediated binding occurs mainly through its interaction with hemicelluloses. Potential benefits of the protein binding to lignocellulose processing have not been tested but are worth investigating. In the case of enzymatic lignocellulose processing, careful consideration of protein loadings will likely be important to prevent potential competition of binding sites for expansin-related proteins and lytic enzymes.
Data availability
No datasets were generated or analysed during the current study.
References
-
Cosgrove DJ. Wall extensibility: its nature, measurement, and relationship to plant cell growth. New Phytol. 1993;124:1–23.
-
Cosgrove DJ. Structure and growth of plant cell walls. Nat Rev Mol Cell Biol. 2024;25:340–58.
-
McQueen-Mason S, Cosgrove DJ. Disruption of hydrogen bonding between plant cell wall polymers by proteins that induce wall extension. Proc Natl Acad Sci U S A. 1994;91(14):6574–8.
-
Gray-Mitsumune M, Blomquist K, McQueen-Mason S, Teeri TT, Sundberg B, Mellerowicz EJ. Ectopic expression of a wood-abundant expansin PttEXPA1 promotes cell expansion in primary and secondary tissues in aspen. Plant Biotechnol J. 2008;6:62–72.
-
Cosgrove DJ. Plant expansins: diversity and interactions with plant cell walls. Curr Opin Plant Biol. 2015;25:162–72.
-
Zerva A, Pentari C, Ferousi C, Nikolaivits E, Karnaouri A, Topakas E. Recent advances on key enzymatic activities for the utilisation of lignocellulosic biomass. Bioresour Technol. 2021;342:126058.
-
Giovannoni M, Gramegna G, Benedetti M, Mattei B. Industrial use of cell wall degrading enzymes: the fine line between production strategy and economic feasibility. Front Bioeng Biotechnol. 2020;8:356.
-
Cosgrove DJ. Microbial expansins. Annu Rev Microbiol. 2017;71:479–97.
-
Gourlay K, Hu J, Arantes V, Andberg M, Saloheimo M, Penttilä M, Saddler J. Swollenin aids in the amorphogenesis step during the enzymatic hydrolysis of pretreated biomass. Bioresour Technol. 2013;142:498–503.
-
Baker JO, King MR, Adney WS, Decker SR, Vinzant TB, Lantz SE, Nieves RE, Thomas SR, Li LC, Cosgrove DJ, Himmel ME. Investigation of the cell-wall loosening protein expansin as a possible additive in the enzymatic saccharification of lignocellulosic biomass. Appl Biochem Biotechnol. 2000;84–86:217–23.
-
Kerff F, Amoroso A, Herman R, Sauvage E, Petrella S, Filée P, Charlier P, Joris B, Tabuchi A, Nikolaidis N, Cosgrove DJ. Crystal structure and activity of Bacillus subtilis YoaJ (EXLX1), a bacterial expansin that promotes root colonization. Proc Natl Acad Sci. 2008;105(44):16876–81.
-
Georgelis N, Nikolaidis N, Cosgrove DJ. Biochemical analysis of expansin-like proteins from microbes. Carbohydr Polym. 2014;100:17–23.
-
Haddad Momeni M, Zitting A, Jäämuru V, Turunen R, Penttilä P, Buchko GV, Hiltunen S, Maiorova N, Koivula A, Sapkota J, Marjamaa K, Master ER. Insights into the action of phylogenetically diverse microbial expansins on the structure of cellulose microfibrils. Biotechnol Biofuels. 2024;17:56.
-
de Sandozequi A, Salazar-Cortés JJ, Tapia-Vázquez I, Martínez-Anaya C. Prevalent association with the bacterial cell envelope of prokaryotic expansins revealed by bioinformatics analysis. Protein Sci. 2022;31(5):e4315.
-
Saloheimo M, Paloheimo M, Hakola S, Pere J, Swanson B, Nyyssönen E, Bhatia A, Ward M, Penttilä M. Swollenin, a Trichoderma reesei protein with sequence similarity to the plant expansins, exhibits disruption activity on cellulosic materials. Eur J Biochem. 2002;269(17):4202–11.
-
Quiroz-Castañeda RE, Martínez-Anaya C, Cuervo-Soto LI, Segovia L, Folch-Mallol JL. Loosenin, a novel protein with cellulose-disrupting activity from Bjerkandera adusta. Microb Cell Fact. 2011;10:8.
-
Baccelli I. Ceratoplatanin family proteins: one function for multiple biological roles? Front Plant Sci. 2015;5:769.
-
Monschein M, Ioannou E, Koitto T, Al Amin LA, Varis JJ, Wagner ER, Mikkonen KS, Cosgrove DJ, Master ER. Loosenin-like proteins from Phanerochaete carnosa impact both cellulose and chitin fiber networks. Appl Environ Microbiol. 2023;89(1):e01863-e11822.
-
Cosgrove DJ. Catalysts of plant cell wall loosening. F1000Research. 2016;5:119.
-
McQueen-Mason SJ, Cosgrove DJ. Expansin mode of action on cell walls; analysis of wall hydrolysis, stress relaxation, and binding. Plant Physiol. 1995;107(1):87–100.
-
Georgelis N, Tabuchi A, Nikolaidis N, Cosgrove DJ. Structure function analysis of the bacterial expansin EXLX1. J Biol Chem. 2011;286(19):16814–23.
-
Dahiya D, Koitto T, Kutvonen K, Wang Y, Haddad Momeni M, de Ruijter S, Master ER. Fungal loosenin-like proteins boost the cellulolytic enzyme conversion of pretreated wood fiber and cellulosic pulps. Bioresour Technol. 2024;394:130188.
-
Jabalera Y, Dahiya D, Ordóñez Cencerrado CD, Caballero AJ, Zaldua N, Eceiza A, Master ER, Perez-Jimenez R. Impact of loosenins on the enzymatic preparation of cellulose nanocrystals. Carbohydr Polym. 2025;357:123469.
-
Dahiya D, Péter-Szabó Z, Senanayake M, Pingali SV, Leite WC, Byrnes J, Buchko GW, Sivan P, Vilaplana F, Master ER, O’Neill H. SANS investigation of fungal loosenins reveals substrate-dependent impacts of protein action on the inter-microfibril arrangement of cellulosic substrates. Biotechnol Biofuels Bioprod. 2025;18(1):27.
-
Wang W, Talide L, Viljamaa S, Niittylä T. Aspen growth is not limited by starch reserves. Curr Biol. 2022;32(16):3619-3627.e4.
-
Sivan P, Heinonen E, Gandla ML, Jiménez-Quero A, Özeren HD, Jönsson LJ, Mellerowicz EJ, Vilaplana F. Sequential extraction of hemicelluloses by subcritical water improves saccharification of hybrid aspen wood grown in greenhouse and field conditions. Green Chem. 2023;25:5634–46.
-
Saeman JF, Moore WE, Mitchell RL, Millett MA. Technique for the determination of pulp constituents by quantitative paper chromatography. Tappi J. 1954;37:336–43.
-
McKee LS, Sunner H, Anasontzis GE, Toriz G, Gatenholm P, Bulone V, Vilaplana F, Olsson L. A GH115 α-glucuronidase from Schizophyllum commune contributes to the synergistic enzymatic deconstruction of softwood glucuronoarabinoxylan. Biotechnol Biofuels. 2016;9:2.
-
Suzuki H, Vuong TV, Gong Y, Chan K, Ho CY, Master ER, Kondo A. Sequence diversity and gene expression analyses of expansin-related proteins in the white-rot basidiomycete, Phanerochaete carnosa. Fungal Genet Biol. 2014;72:115–23.
-
Brodelius P, Nilsson K. Entrapment of plant cells in different matrices: a comparative study. FEBS Lett. 1980;122:312–6.
-
Sivan P, Patel VR, Rajput KS, Rao KS. Distribution of tension wood like gelatinous fibers in the roots of Acacia nilotica (Lam.) Willd. Planta. 2014;240:1191–202.
-
Sivan P, Vilaplana F, Mellerowicz EJ. Cell wall polysaccharide matrix dynamics during wood development. In: Geitmann A, editor. Plant cell walls: research milestones and conceptual insights. Boca Raton: CRC Press; 2023. p. 412–40.
-
Martínez-Abad A, Giummarella N, Lawoko M, Vilaplana F. Differences in extractability under subcritical water reveal interconnected hemicellulose and lignin recalcitrance in birch hardwoods. Green Chem. 2018;20:2534–46.
-
Wang T, Park YB, Caporini MA, Rosay M, Zhong L, Cosgrove DJ, Hong M. Sensitivity-enhanced solid-state NMR detection of expansin’s target in plant cell walls. Proc Natl Acad Sci USA. 2013;110(41):16444–9.
-
Wang Q, Chen L, Lin H, Yu D, Shen Q, Wan L, Zhao Y. The binding, synergistic and structural characteristics of BsEXLX1 for loosening the main components of lignocellulose: lignin, xylan, and cellulose. Enzyme Microb Technol. 2016;92:67–75.
-
Georgelis N, Nikolaidis N, Cosgrove DJ. Bacterial expansins and related proteins from the world of microbes. Appl Microbiol Biotechnol. 2015;99:3807–23.
-
Tovar-Herrera OE, Rodríguez M, Olarte-Lozano M, Sampedro-Guerrero JA, Guerrero A, Pinto-Cámara R, Alvarado-Affantranger X, Wood CD, Moran-Mirabal JM, Pastor N, Segovia L, Martínez-Anaya C. Analysis of the binding of expansin Exl1, from Pectobacterium carotovorum, to plant xylem and comparison to EXLX1 from Bacillus subtilis. ACS Omega. 2018;3(6):7008–18.
-
Evert RF. Esau’s plant anatomy; meristems, cells, and tissues of the plant body: their structure, function, and development. Hoboken: John Wiley & Sons, Inc.; 2006. p. 255–316.
Acknowledgements
We acknowledge all scientific discussions enabled through the BioUPGRADE research network (https:// bioupgrade.eu). Authors are grateful to Umeå Centre for Electron Microscopy (UCEM) for providing facilities for sample preparation and technical support for Transmission Electron microscopy analysis.
Funding
Open access funding provided by Royal Institute of Technology. This project has received funding from the European Union’s Horizon 2020 research and innovation program under Grant Agreement No 964764. The content presented in this document represents the views of the authors, and the Commission is not responsible for any use that may be made of the information it contains.
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sivan, P., Dahiya, D., Jabalera, Y. et al. Carbohydrate-binding domain CBM63 of microbial expansin-like BsEXLX1 facilitates the adsorption of expansin-related proteins to hemicelluloses in plant secondary cell walls. Biotechnol. Biofuels Bioprod. 18, 70 (2025). https://doi.org/10.1186/s13068-025-02674-x
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1186/s13068-025-02674-x