
Introduction
Enzymatic hydrolysis is among the most promising emerging technologies for polyester recycling. It pledges a full environmentally friendly, and effective management of the rising global plastic waste1,2 due to its mild operative conditions coupled with the advantage of substrate specificity useful for recycling of blended materials. The first enzymes active on polyesters were isolated from Actinobacteria and Fungi (Fusarium solani pisi in the 1970s3. In fact, since at least aliphatic polyesters such as cutin are ubiquitous in nature, numerous organisms are equipped with polyester hydrolysing enzymes. In the last decades, a plethora of ecosystems has been screened to identify new enzymes hydrolyzing polyesters4 with compost and marine environment as few of the most prolific sources. However, only some dozens of enzymes are currently known to efficiently decompose plastics actively5 justifying a continuous effort in the research of unique ecosystems, such as carnivorous plants, still unexplored in this regard6. In the last decade, enzyme based processes for the recovery of valuable building blocks from the most common commercial polyester poly(ethylene terephthalate) (PET), and a biodegradable alternative, namely as poly(butylene adipate –co– terephthalate) (PBAT) have been reported to allow resynthesis7. However, there is still a need for more efficient enzymes for this purpose.
It has been 125 years after the publication of Insectivorous plants8 by Charles Darwin and already almost 600 plants are now categorized under this class. The broad investigation is obviously due to their extraordinary ability of deriving nutrients not only from soil, but also from animals, practicing predation by means of evolved dedicated structures and mechanisms. Nepenthes and Sarracenia, well-documented taxa, passively capture their preys, mainly insects, through pitchers, where pepsin and other proteases are known to carry out prey digestion8. Enzymes are thought, in fact, to synergistically act to compensate the high “costs” of the carnivory9. Although the complete genome is not yet fully translated, transcriptomic fingerprints are available for some species of both genera to provide information about the pool of produced enzymes10,11 (proteases, RNAse, nuclease, phosphatase).
While the main application of carnivorous plant derived enzymes has been traditionally limited to medicine, their potential in environmental biotechnology has not yet been exploited10. Nevertheless, prey digestion mechanism is known to rely on signal transduction and symbiosis with microorganisms, inhabitants of the pitchers12,13,14. In addition, low prey specificity (therefore wide substrates ranges) together with broad pH and temperature optima ranges, imply a large variety of enzymes among which some could possibly be active on polyesters.
In this work, hydrolysis of the by far most common commercial polyester poly(ethylene terephthalate) (PET), and a biodegradable alternative, poly(butylene adipate –co– terephthalate) (PBAT) by the pitcher fluid of the carnivorous plants Nepenthes alata and Sarracenia purpurea was investigated. Although enzymatic hydrolysis by means of microbial cutinases of these polyesters has been widely studied15,16 the potential of enzymes from carnivorous plants has not yet been assessed for plastic degradation. The study therefore aims at searching for polyester active enzymes in the plant pitchers. Different conditions were screened also via proteomic tools to provide a more detailed picture of the putative responsible catalysts for polyesters depolymerization17,18. The possibility of applying the identified biocatalysts in plastic treatment would then result from several subsequent steps involving a multidisciplinary collaboration: from identification to isolation, characterization and optimization of the activity of these biocatalysts on the most prominently used synthetic polyesters through genetic engineering.
Materials and methods
Samples and chemicals
N. alata, S. purpurea and Tenebrio molitor (mealworm) were purchased from OBI (Austria). N. alata and S. purpurea were bought at juvenile growth stage and kept for the duration of the experiment in the same substrate. T. molitor was purchased and maintained as larvae. PET and PBAT were purchased from GoodFellow (UK) and received from BASF (Germany) respectively. Other chemicals including terephthalic acid (TPA) for calibrations, methyl jasmonate (JA) (> 98%), dithiothreitol (DTT) and iodoacetamide (IAA), Bradford reagent and bovine serum albumin (BSA), as well as triethylammonium bicarbonate buffer (TEAB) were supplied by Sigma-Aldrich at analytic purity level. Trypsin was purchased from New England Biolabs. Recombinant Nepenthes distillatoria Aspartic proteinase nepenthesin-1 expressed in Escherichia coli was purchased at Biozol (Eching b. München). H.insolens cutinase was from Novozymes.
Incubation of polyesters with carnivorous plants
Standard sized and weighed amorphous films of PET and PBAT (3 cm x 0.5 cm) (Table S1, ESI) were washed in three steps as described in Bertolini et al.16 prior to incubation. A first rinsing with Triton X-100 (2 g∙L− 1) was followed by a further wash in Na2CO3 and a final step in ultrapure water.
The pitchers of the plants were fed with additives (JA and/or mealworm), polymers (one film strip per pitch) and ultrapure water, according to the scheme simplified in Table 1, to evaluate the effect of the different combinations on polymer degradation in a time frame of 5 weeks. The incubation was performed in sunlight in greenhouses and plants were continuously watered and humified. The washed and dried polymer strips were placed in each carnivorous plant pitcher, together with 5 mL of ultrapure water. The pitchers were covered with parafilm to minimize external contaminations of the system. The different conditions ranged from polymer only, to mealworm-added and JA fed pitchers. JA standard was prepared by diluting JA in ultrapure water (concentration of 10 µg∙mL− 1). The concentration of JA in the pitcher was 0.2 µg∙mL− 119,20. From each pitcher 2 mL samples were taken once per week, until final harvesting of the total content after 5 weeks. Figure 1 summarizes the main experimental procedures, further discussed in the next paragraphs.

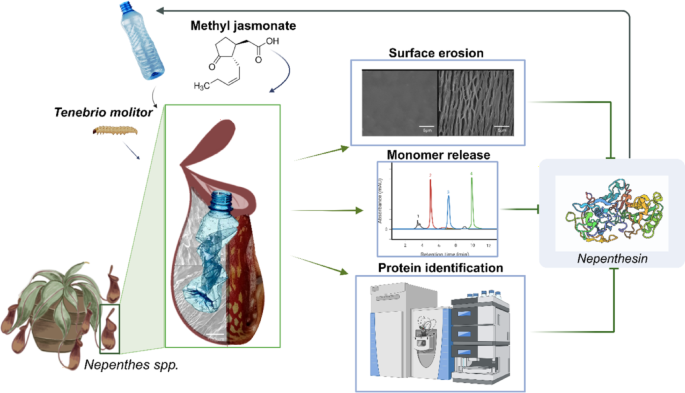
Schematic representation of samples/materials and experimental methods used in the research. Nepenthes spp., Tenebrio molitor, plastic samples, jasmonic acid. Analysis: surface characterizations, monomer release quantification and protein quantification and identification.
Analysis of polymers and hydrolysis products
The remaining recoverable polymers after incubation with the plants were isolated and washed with 8 M urea to remove residuals from the incubation. This was followed by further cleaning using the procedure described above. The surface chemical changes of the dried final polymers were determined through Fourier Transform-Infrared Spectroscopy (FT-IR, PerkinElmer Spectrum 100 Spectrometer) in ATR mode. Spectra were recorded from 4000 cm− 1 to 650 cm− 1, at a resolution of 2 cm− 1 for 40 scans. A reference table was used to build the functional group profile of each sample. Surface erosion patterns were analyzed through Scanning Electron Microscopy (SEM) acquired using a FEI Quanta 250FEG (Thermo Fisher Scientific, Hillsboro, OR) under high vacuum condition and a high tension ranging from 5 kV to 10 kV. Before acquisition through Everhart-Thornley-Detector in secondary electron (SE) mode, each sample was sputter-coated with gold (10 nm layer) under Argon atmosphere in a Scancoat six (Edwards, Burgess Hill, UK) to improve electrical conductivity.
To analyze soluble hydrolysis products, the collected liquid sample was diluted in methanol (1:1) for enzyme precipitation, acidified and filtered (0.2 μm polyamide filters) before High Performance Liquid Chromatography (HPLC, Agilent Technologies, 1260 Infinity supplied with reversed phase column C18) analysis as previously described15. TPA was quantified using a UV detector at the wavelength of 245 nm. The instrument was set at 0.85 mL∙min− 1 flow rate of methanol and 0.1% ormic acid. The same calibration was used to quantify the monomer concentration according to the peak intensity.
Statistical analysis was performed to assess differences in TPA concentrations between experimental conditions. Levene’s test was used to confirm homogeneity of variances, and the Shapiro-Wilk test was applied to evaluate normality. Since both conditions were met, ANOVA was conducted to test for significant differences between groups, followed by Tuke’s Honest Significance Difference (HSD) post-hoc test for pairwise comparison. Statistical significance was set at α < 0.05.
Filter-aided sample preparation (FASP) for proteomics
Following a first concentration step (centrifugation through molecular cut-off of 10 kDa, 4 °C, 3700 rpm, 10 min), total protein concentration in pitcher-fluid was determined through Bradford assay. Samples were diluted in Bradford buffer and compared to standards in ultrapure water as reference. An amount of 10 µL of each sample/standard/control were placed in a 96-well plate, added with 200 µL of Bradford reagent. After 5-minute incubation while shaking at room temperature, the absorbance at 595 nm was measured through a Tecan plate reader.
FASP digestion protocol was then applied to prepare the samples for proteomics analysis. All steps were performed under laminar flow workbench to avoid external contamination. Each specimen was reduced through 30-minute incubation with DTT 5 mM at 37 °C, followed by an additional 30-minute incubation with IAA 15 mM (no light condition, room temperature) to alkylate the extracts. The samples were then centrifuged (5 min, 12700 rpm, room temperature), in a spin column (MW cut-off of 100 kDa) primed with 400 µL of 8 M urea in 50 mM TEAB buffer. The samples were added in 400 µL-step per 20 min each. A final wash was performed through 300 µL of 8 M urea in 50 mM TEAB buffer. The column was further washed with three subsequent steps of 300 µL 50 mM TEAB buffer and afterwards transferred to a low-bind tube, where proteolytic digestion was carried out through trypsin in a 1:25 ratio protease: protein. The column was then incubated in 50 mM TEAB buffer 100 µL at 37 °C overnight. The digested peptides were eluted by centrifugation at 12,700 rpm for 5 min into the low bind tube, vacuum dried at 40 °C and stored for further analysis at -20 °C.
LC-MS/MS analysis
The samples were added with 10 µL of 0.1% trifluoroacetic acid solution. A 10-minute sonication step in a water bath at room temperature was performed prior to LC-MS/MS analysis as described by Yu et al.21. Briefly, peptides were separated on an Ultimate 300 RSLC nano system (Thermo Fisher Scientific, USA) equipped with an Acclaim PepMap precolumn (15 mm length, 300 μm inner diameter, 5 μm particle size, 100 Ǻ pore size; Thermo Fisher Scientific) and an Acclaim PepMap C18 analytical column (250 mm length, 75 μm inner diameter, 3 μm particle size; Thermo Fisher Scientific). A gradient of eluents A (0.1% v/v formic acid) and B (80% v/v acetonitrile with 0.1% v/v formic acid), ranging from 4% B to 35% B in 120 min and further to 55% B in 15 min, was used at a constant flow rate of 0.3 µL∙min− 1. A washing step with 90% B followed. Peptides were analyzed on a Q Exactive Plus Hybrid Quadrupol-Orbitrap (Thermo Fisher Scientific) mass spectrometer. The 25 most intense precursor ions in the range of 400–500 m/z with charge states of 2–4 exceeding 100 counts/s were fragmented for 25 ms. Spectra of fragmented ions were collected for 110 ms in the range of 100 m/z – 1800 m/z. HeLa protein digests were used as quality control standards.
Protein identification and quantification
A custom protein database, consisting of reviewed hydrolase family proteins (EC3), the reference proteome of T. molitor (UniProt-ID: UP000719412), and all proteins associated with Nepenthes (as of Oct. 10th 2023), was downloaded from UniProt22. Additionally, a proteome predicted from sequencing data of Nepenthes gracilis obtained from Saul et al.23 and the Common Repository of Adventitious Proteins (cRAP)24 were added. The proteome of T. molitor was included to enable identification of mealworm-derived proteins, that might have influenced the results, while the inclusion of all hydrolase family proteins (EC3) allows for the consideration of microbial proteins which could potentially contribute to the degradation process. The human proteome (UniProt-ID: UP000005640) was used as database for the HeLa standards. Decoys were added to the databases through Philosopher (v5.1.1)25. Sample files were searched against the database using the default workflow for label-free quantification in FragPipe (v21.0)26. In brief, peptide spectrum matches (PSM) were searched with strictly tryptic digestion, allowing 2 missed cleavages, a peptide length of 7–50 amino acids, and a peptide mass of 500 Da – 5000 Da. Precursor and fragment ion mass tolerance was set to 20 ppm and identified PSMs and proteins were filtered at a false discovery rate (FDR) of 1%. During quantification the m/z and retention time tolerance was set to 10 ppm and 0.4 min respectively. Only features with a minimum of 3 scans were quantified. The match between runs was enabled with a maximum of 10 donor runs, a retention time tolerance of 1 min, and an FDR of 1%.
Differential abundance analysis
Data processing and analysis was performed in R (v4.2.1)27. Proteins identified as contaminants or not present in at least 66% of replicates in a condition and time point were removed from the analysis and protein intensities were log2 transformed. Differential abundance analysis was performed using the R-package limma (v3.54.2)28. Comparisons were made between conditions with high TPA release and conditions with low TPA release or without added polyester films (Table S2). Proteins that were present in at least 50% of these comparisons and overabundant (log2FC > 2) in at least 50% of them were considered putative polyester degraders. Structural proteins like actin, tubulin and chaperones among others, were dismissed, as they were not considered of being directly involved in the hydrolysis of polyesters. Data is available as an additional supplementary information file.
Homology analysis
Sequences of identified putative polyester degrading proteins were compared to enzymes (see Table S3 in ESI) which have been reported to hydrolyze synthetic polyesters using blastp (v2.15.0)29. Structural similarity of these proteins was investigated by superimposing the protein structures in ChimeraX (v1.8)30. Protein structures were obtained from Protein Data Bank (PDB)31 if no 3D model was available the structure was predicted using the Google Colab notebook of AlphaFold (v2.3.2)32.
Molecular docking
Binding of the polyesters PET and PBAT was analyzed by performing molecular docking in AutoDock Vina33 using the DockingPie plugin (v1.2.1)34 for PyMOL (v3.0.3)35. A maximum of 40 binding poses was analyzed per ligand with an energy range of 3 kcal∙mol− 1 and exhaustiveness set to 8. Model ligands of PET and PBAT were generated from SMILES notations in Avogadro (v1.2.0)36 and the structures were optimized in the general AMBER force field (GAFF)37. Binding affinities of the model ligands to the putative polyester degrader were compared to the binding affinity of a reported, well-binding ligand, and to that of the polyesters to Polyester hydrolase Leipzig 7 (PDB: 7NEI), an enzyme capable of efficiently hydrolyzing PET38. Protein-ligand interactions of reasonable poses were analyzed in PLIP (v2.3.0)39.
Hydrolysis of PET and PBAT with recombinant Nepenthes Aspartic proteinase nepenthesin-1
Esterase activity of recombinant nepenthesin-1 was determined on para-nitrophenylbutyrate (p-NPB) as model substrate monitoring absorbance at 405 nm as previously described40. Hydrolytic activity of the recombinant nepenthesin-1 was tested on PET and PBAT films (3 cm x 0.5 cm size). Therefore, nepenthesin-1 was diluted in ultrapure water and incubated at 37 °C, 150 rpm for 72 h in 2 mL microcentrifuge tubes at a final concentration of 0.015 mg∙mL− 1. The same polymers were incubated with H. insolens cutinase at the same concentration and incubation conditions. Hydrolysis was assessed through the release of TPA and surface characterization of the films through Scanning Electron Microscopy.
Results
Hydrolysis of polyesters by Nepenthes and Sarracenia pitcher fluids
In a first step, hydrolysis of polyesters films made from PET and from PBAT in the pitcher fluid of N. alata was studied based on the release of TPA. TPA concentration was found to increase in all samples consistently within the considered time frame: the highest TPA values corresponded to week 5, the final time point also due to the lifespan of the pitch (approximately 2 months). There was no overlapping and influence of TPA signal (retention time 5.7 min in the applied system) with tyrosine´s signal.
The highest concentration of TPA released from PBAT and PET of 1.61 mM and 1.27 mM, respectively, was recorded in Nepenthes alata for conditions in presence of alive mealworm (Tm_a) and JA (Fig. 2, Table S4). Interestingly, in the presence of JA and Tm_a around 10 times more TPA was released into the pitcher liquid compared to the respective polymer incubated alone in the pitcher (PET and PBAT, p < 0.001).
For PET, even addition of dry mealworms (Tm_d) led to a 3-fold increase of TPA (p < 0.01). However, for PBAT, this effect was not observed, as PBAT/Tm_d showed only a moderate increase compared to PBAT alone (p < 0.001). On the other hand, the presence of JA approximately doubled the release of TPA (0.66 mM vs. 0.37 mM, p < 0.001). In addition, the combined condition (JA + dry mealworm) already led to better depolymerization of PET, as TPA content in this set-up was significantly higher than the only-mealworm condition (p < 0.001). However, for PBAT, the combined condition (Tm_d/JA) did not result in a statistically significant increase in TPA content compared to the only-mealworm condition (Tm_d, p > 0.05). The samples with polyester only led to a comparatively low amount of TPA, however for the controls containing hormone only or mealworm only no TPA was detected. These results indicate that not only a mechanical stimulus (body of the mealworm), but also with the presence of the hormone JA could trigger the secretion of enzymes responsible for the hydrolysis of polyesters.

Terephthalic acid (TPA) (mM) released from the hydrolysis of polyester films made from poly(ethylene terephthalate) (PET) and from poly(butylene adipate-co-terephthalate) (PBAT) incubated in the fluid of N. alata pitchers monitored based on the release of of TPA (expressed in mM). The influence of the presence of jasmonic acid (JA) and alive (Tm_a) and dry (Tm_d) T. molitor was assessed. (A) PET films. At the last time point, concentrations for PET/Tm_a/JA, PET/Tm_d/JA, and PET/Tm_a are not significantly different from each other but are significantly different from the other PET conditions (p < 0.001); (B) PBAT films. At the last time point, concentrations are significantly different between treatments (p < 0.05).
The polyesters were also hydrolyzed in S. purpurea pitcher fluids though to a lower extent when compared to N.alata (Fig. S1 and Table S5). Highest amounts of TPA were detected for PBAT (polymer only) and for PBAT/JA (polymer in presence of JA) for PBAT and PET/Tm_a/JA (alive mealworm and JA) for PET samples. Also the contribution of the hormone seemed to have a lower impact than in N. alata (12 times increase of TPA upon hormone introduction vs. only 2-fold in S. purpurea).
With the exception of PBAT/Tm_d and PET/Tm_d, all polymer-containing groups showed significant differences in TPA concentrations between N. alata and S. purpurea. Non-polymer-containing groups (Control, JA, Tm_a, Tm_a/JA, Tm_d) were not significantly different between the two plants. For the polymer-only conditions (PET and PBAT), S. purpurea released significantly more TPA than N. alata (p < 0.001). In contrast, all other polymer-containing conditions resulted in significantly higher TPA concentrations in N. alata compared to S. purpurea (p < 0.001).
In S. purpurea, the condition PET/Tm_a/JA consistently showed significantly higher TPA concentrations compared to PET, PET/JA, and PET/Tm_d (p < 0.05). For PBAT, both PBAT/Tm_a and PBAT/Tm_d had significantly lower TPA concentrations compared to PBAT/JA (p < 0.05). However, no significant differences were observed between most other conditions. These results suggest that while S. purpurea exhibits some variability in TPA release across conditions, its enzymatic activity is less pronounced compared to N. alata.
These findings provide a first evidence that pitcher fluids from N. alata and S. purpurea possess hydrolytic enzyme activity toward synthetic polyesters, with N. alata exhibiting significantly higher enzymatic efficiency. The hydrolysis of PET and PBAT, monitored via TPA release, was also enhanced by the presence of jasmonic acid (JA) and T. molitor (Tm), particularly in N. alata, where JA and alive Tm proved a synergistic effect. In these plants, the hormone has therefore an activating effect on the production of hydrolases, as the depolymerization was significantly higher when JA was present. In contrast, S. purpurea showed a more moderate response, but both plant digestive fluids showed a polyester degrading action, highlighting a previously underexplored natural system with potential relevance for plastic recycling.
As shown in previous studies7 hydrolysis of same size PET and PBAT films with 5 µM pure commercial enzyme (H.insolens cutinase) yielded 17 mM and 8 mM TPA, respectively, within 72 h of incubation at 70°C7 which helps the accessibility of the films to the enzyme. Being able to detect monomers within a few weeks of incubation at room temperature in a complex mixture, such as the plant digestive fluid, represents groundbreaking evidence of hydrolytic potential in such systems.
Fourier transform infrared spectroscopy (FT-IR) and scanning electron microscopy (SEM) analysis of partially hydrolyzed polyesters
Surface characterization of the after-treatment films via both FT-IR and SEM was used to bring further evidence of the incubation derived film erosion. The analysis was carried out for those polymers that were available after incubation. Most of the samples showed a gelatinous layer that was removed through extensive washing in urea prior to analysis. PET spectra of N.alata-incubated polymers are plotted in Fig. S2 (ESI). While there is almost complete superimposition of the original spectrum and only polymer condition spectrum, the normalized absorbance shifts progressively from the original profile in the incubated samples, especially PET/Tm_a/JA, in accordance with the HPLC analysis for this sample. The difference is visible in all the main peaks of the spectra, while it is mainly relevant in the 1710 cm− 1, corresponding to aryl ester stretching. Other groups, whose decrease indicates the hydrolysis are 1240 cm− 1, and 1090 cm− 1, corresponding respectively to C = O carbonyl stretching, C-O stretching of ester group and vibrations of ester C-O bond. Similarly, changes in the PBAT surface upon incubation in N.alata pitcher fluids (Fig. S3) were seen such as a reduction of the peak at 1710 cm− 1, mostly marked in PBAT/Tm_a/JA. Moreover, a decrease in relative absorbance was recorded at 1250 cm− 1, 1160 cm− 1, 1100 cm− 1 area, as C = O stretching and CH plane bending.
Longitudinal sections of the plants (Fig. S4 in ESI) as well as the residual polymers already analyzed through FT-IR, were further characterized via SEM. Despite a similar extent of hydrolysis according to TPA release, for both polymers, SEM pictures (Figs. 3 and S5) indicated a different mechanism of hydrolysis for PBAT and PET. For PBAT, massive erosion with cavities was seen after 5 weeks of incubation, indicating vertical progressing of hydrolysis. On the other hand, for PET a more homogenous pattern was seen. Hydrolysis patterns are known to be not only polymer dependent. In fact, different types of polyester hydrolases show different erosion modes. As an example, lipases preferentially catalyze vertical hydrolysis, compared to cutinase from Fusarium solani41. Here, the most visible degradation profile was observed in PBAT/Tm_a (alive mealworm, no hormone). Figures S6, 7 (ESI) highlights the same conditions incubated in S. purpurea pitchers. Similar findings were acknowledged in Quartinello et al.6. Consistently with the data from the recovered monomers, the films were degraded to a minor extent.

1000 X and 10,000 X SEM micrographs of PBAT films incubated in N. alata pitchers for five weeks.
Despite the fact that FT-IR and SEM provides a qualitative analysis, the results align with the previous HPLC derived consideration. Overall, all analysis results suggested more pronounced hydrolysis of polyesters in N. alata pitchers. Therefore, in the next step, studies towards the responsible proteins were conducted only for this plant.
Proteomics
Proteomic analyses were performed exclusively on N. alata pitcher fluids, as the HPLC results for S. purpurea fluids showed few significant differences between conditions and released far less TPA compared to Nepenthes. Additionally, the lack of available reference proteomes for Sarracenia species, further limited the feasibility of proteomic analysis for these samples.
Proteins were quantified through Bradford before and after the concentration through Vivaspin columns. Outputs are summarized in table S6.
In HeLa quality control standards, 3752 proteins were identified in line with the range reported in literature, 2700-600042,43,44,45,46. After filtering out contaminants and not consistent proteins across replicates, a total of 736 proteins were associated to Nepenthes pitcher fluids.
TPA release data indicated most efficient hydrolysis of polyesters in the presence of alive T. molitor (PET/Tm_a/JA; PET/Tm_a; PBAT/Tm_a; PBAT/Tm_a/JA). Therefore, these samples were compared to those containing dry T. molitor (PET/Tm_d, PBAT/Tm_d) at every time point (see Table S2 for a list of comparisons). Aspartic proteinase nepenthesin-1 (Uniprot-ID: Q766C3) was present in 83% of the comparisons and overabundant in 70%, while carboxypeptidase (Uniprot-ID: A0A140GML6) was present in 50% and overabundant in 63% thereof. When also including live mealworm conditions Tm_a and Tm_a/JA in the comparison, carboxypeptidase no longer met the criteria suggesting that its presence is influenced by the presence of mealworm rather than by the polyesters. However, after accounting for FDR by the Benjamini-Hochberg method, also p-values of nepenthesin rose from between 0.004 and 0.05 to values between 0.1 and 0.5. Hence, no significant influence of the different feed stimuli (alive or dry mealworm, JA addition or type of polyester) on the abundance of nepenthesin or other proteins was observed.
Molecular docking
The high abundance of nepenthesin in samples with elevated TPA release prompted further analysis. Alignment of amino acid sequences with well-known polyester hydrolyzing enzymes such as Leaf and Branch compost cutinase (LCC) and H. insolens cutinase (HiC) did not reveal any homology of nepenthesin (Table S3, ESI). Similarly, no structural similarity was observed when superimposing the 3D-structures.
For molecular docking, pepstatin, competitive aspartic protease inhibitor47 was chosen as baseline due to its strong binding to nepenthesin’s active site. Pepsin (PDB: 1PSO) was chosen as reference (sequence identity with nepenthesin: 34.7%, E-value: 3∙10− 11, RMSD: 1.788 Å for 186 aligned residues, indicating conserved active sites and high structural similarity (see Fig. S8)) to validate the docking procedure. Re-docking rigid pepstatin, co-crystallised with pepsin, to pepsin resulted in a binding affinity of -15.96 kcal∙mol− 1, which was reduced by half (-7.91 kcal∙mol− 1) when enabling ligand flexibility. Docking pepstatin to nepenthesin positioned the hydroxyl group of the statin residue in the third position of pepstatin in close proximity to the aspartic acid residues of the active site, which is consistent with the reported mechanism of inhibition48 (Fig. S9). The binding affinity of pepstatin to nepenthesin was − 7.47 kcal∙mol− 1. As an example of known effective hydrolysis activity, the polyesters PET and PBAT were docked to PHL7 resulting in binding affinities of -4.50 kcal∙mol− 1 and − 4.75 kcal∙mol− 1, respectively. The validity of the binding poses was confirmed by interactions previously described in literature39,49,50,51,52,53 including π-stacking of aromatic rings with F36 and W156, hydrophobic interactions of L210 and I179 and the proximity of the ester bond to S131 of the catalytic triad (Fig. S10).
The docking analysis of PET and PBAT to nepenthesin revealed several binding poses with ester bonds within 3 Å − 6 Å of catalytic aspartate residues (D35 and D237). These poses were stabilized by several hydrophobic interactions and hydrogen bonds. Residues F201 and Y244 on either end of the binding cleft were observed to engage in π-stacking with the aromatic rings of the polyesters (Fig. 4). Binding affinities were comparable to pepstatin, ranging from − 7.7 kcal∙mol− 1 to -6.8 kcal∙mol− 1 for PET and − 6.8 kcal∙mol− 1 to -6.3 kcal∙mol− 1 for PBAT. The lower binding affinities compared to PHL7 could be explained by the design of the active sites. PHL7’s active site is located on the surface of the enzyme, while nepenthesin’s long binding cleft is partially covered by a hairpin flap allowing more interactions with surrounding residues.
In aspartic proteases, the hydrolytic activity towards peptide bonds is mediated through a water molecule, activated by the active site aspartic acid residues54. The close proximity of the ester bonds to the active site could make them susceptible to nucleophilic attack by such an activated water molecule, resulting in the hydrolysis of the polyesters. This theory is supported by a 2021 study of Bose and Zhao55 who already demonstrated that a synthetic enzyme, mimicking the aspartic protease mechanism, was capable of hydrolysing the aromatic esters p-nitrophenyl acetate and p-nitrophenyl formate. Therefore, nepenthesin could indeed be responsible for hydrolysis of polyesters in N.alata pitcher fluid.

Binding of PET and PBAT to nepenthesin. (A) A slice through the binding cleft of nepenthesin with a bound PET chain (yellow), showing surface hydrophobicity (hydrophobic residues in red, hydrophilic in blue). Rendered in ChimeraX. (B) Front view of nepenthesin with a docked PET (purple) and active site ASP residues (pink). (C and D) Binding-interaction of docked of PET (purple) and PBAT (teal) to nepenthesin, with active site ASP residues in pink. Distances from the active site to the ester bond are shown in pink lines, H-bonds in blue dashed lines, hydrophobic interactions in black dotted lines, and π-stacking in green dotted lines. Rendered in PyMOL.
Hydrolysis of PET and PBAT by nepenthesin-1
Since the aspartic proteinase nepenthesin was highly abundant in conditions leading to better hydrolysis of polyesters in pitcher N. alata fluids, hydrolysis of the polyesters by recombinant nepenthesin-1 was investigated. Nepenthesin-1 from N.distillatoria was used in these experiments, as the enzyme from N. alata was not commercially available. Nevertheless, the nepenthesin identified in the proteomic study and the one from N.distillatoria share 43.7% sequence identity with 99% query coverage and an E-value of 9∙10− 73, supporting its use as representative enzyme. Nepenthesin-1 showed an esterase activity on p-NPB of 794 U∙mg− 1 and retained circa 20% of the activity after 72 h incubation at 37 °C (Table S7, ESI). Under these conditions, incubation of PET and PBAT with nepenthesin-1 lead to significant depolymerization (See HPLC chromatograms and histogram in Figs. S11–14), with TPA concentration of 0.28 mM TPA for PET and 0.72 mM TPA for PBAT. In comparison, the well-studied H. insolens cutinase7 incubated at the identical protein concentration yielded 0.21 mM TPA for PET and 0.28 mM for PBAT. No monomers were found in the controls (polymers with only water or only enzyme solution without polymers). The residual films were also washed (same protocol as for the preparation) and characterized in surface morphology through SEM. While PET surfaces showed limited signs of erosion (Fig. S15), PBAT exhibited a similar degradation pattern as seen after incubation in N. alata pitchers (Fig. S16, see also Fig. 2). These results find consistency with previous research on polyester hydrolysis by means of serine proteases56,57 although at the best of the authors´ knowledge, not updated extensive characterizations of aspartic proteases activity on polyesters exist. Nepenthesin, in particular, has already established biotechnological application for mass spectrometry preparation, as an example9. Its protease activity regulation, as well as its specific cleavage site, have been measured58 but at the moment, without any focus on polyesters as substrate. However, some works related nepenthesin proteolytic activity with the presence of insects (Drosophila melanogaster)58 either because of indirect acidification of the pitcher fluid, which would activate the protease, or through signaling due to mechanical stimuli19. Among the highest TPA concentrations were in fact those samples incubated with hormone and mealworms, which additionally supports the mediated activation of pitcher enzymes.
Conclusion
Tracing monomers of depolymerization confirmed hydrolysis of polyesters in pitcher fluids of the carnivorous plants N. alata, and S. purpurea. Investigation of the proteins present in N. alata pitcher fluids under different conditions suggested nepenthesin as at least a candidate responsible for the hydrolysis reaction. This was also indicated by docking of this aspartic protease with the polyesters PET and PBAT. In addition, purified recombinant nepenthesin was incubated on PET and PBAT to confirm its role in polyester depolymerization and indeed hydrolysis of the polyesters was seen through released soluble monomers in the hydrolysates and by SEM analysis of surface erosion patterns. The activity of the enzyme on these polyesters was comparable with the activity of the well-studied cutinase from H. insolens. Yet, in contrast to cutinases/esterases there is no/very little information in the literature about hydrolysis of these aromatic polyesters by proteases. The possibility of identifying a responsible for polyester hydrolysis paves the way for immediate practical application, such as the expression and utilization of pure nepenthesin in polyester recycling. This also indicates that under-explored natural niches, like carnivorous plants, could be a source of enzymes with polyester hydrolyzing activity, confirming the potential of bioexploration besides genetic enzyme improvement.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable requestThe mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD06190759.
References
-
European bioplastics. December 2023 Bioplastics Market Development Update 2023. 2022–2023. (2023).
-
Geyer, R., Jambeck, J. R. & Law, K. L. Production, use, and fate of all plastics ever made. Sci. Adv. 3, 25–29 (2017).
-
Purdy, R. E. & Kolattukudy, P. E. Hydrolysis of plant cuticle by plant pathogens. Properties of cutinase I, cutinase II, and a nonspecific esterase isolated from fusarium Solani Pisi. Biochemistry 14, 2832–2840 (1975).
-
Zhu, B., Wang, D. & Wei, N. Enzyme discovery and engineering for sustainable plastic recycling. Trends Biotechnol. 40, 22–37 (2022).
-
Buchholz, P. C. F. et al. Plastics degradation by hydrolytic enzymes: the plastics-active enzymes database—PAZy. Proteins Struct. Funct. Bioinforma. 90, 1443–1456 (2022).
-
Quartinello, F. et al. Together is better: the rumen microbial community as biological toolbox for degradation of synthetic polyesters. Front. Bioeng. Biotechnol. 9, 1–11 (2021).
-
Siracusa, C. et al. There and back again: recovery of terephthalic acid from enzymatically hydrolyzed polyesters for resynthesis. https://doi.org/10.1021/acssusresmgt.4c00430 (2025).
-
Charles Darwin. Insectivorous Plants (D. Appleton, 1895).
-
Miguel, S., Hehn, A., Bourgaud, F. & Nepenthes State of the Art of an inspiring plant for biotechnologists. J. Biotechnol. 265, 109–115 (2018).
-
Ravee, R., Salleh, F. I. M. & Goh, H. H. Discovery of digestive enzymes in carnivorous plants with focus on proteases. PeerJ 2018 (2018).
-
Srivastava, A., Rogers, W. L., Breton, C. M., Cai, L. & Malmberg, R. L. Transcriptome analysis of sarracenia, an insectivorous plant. DNA Res. 18, 253–261 (2011).
-
Buch, F. et al. Secreted pitfall-trap fluid of carnivorous nepenthes plants is unsuitable for microbial growth. Ann. Bot. 111, 375–383 (2013).
-
An, C., Il, Fukusaki, E. I. & Kobayashi, A. Aspartic proteinases are expressed in pitchers of the carnivorous plant nepenthes Alata Blanco. Planta 214, 661–667 (2002).
-
Ellison, A. M. & Gotelli, N. J. Energetics and the evolution of carnivorous plants – Darwin’s ‘most wonderful plants in the world’. J. Exp. Bot. 60, 19–42 (2009).
-
Yang, Y. et al. Complete bio-degradation of poly (butylene adipate-co-terephthalate) via engineered cutinases. Nat. Commun. 14, (2023).
-
Weinberger, S. et al. Enzymatic degradation of poly (Ethylene 2,5-furanoate) powders and amorphous films. Catalysts 7, (2017).
-
Tesei, D. et al. Shotgun proteomics reveals putative polyesterases in the secretome of the rock-inhabiting fungus Knufia Chersonesos. Sci. Rep. 10, 1–15 (2020).
-
Robinson, S. L., Piel, J. & Sunagawa, S. A roadmap for metagenomic enzyme discovery. Nat. Prod. Rep. 38, 1994–2023 (2021).
-
Pavlovič, A. & Mithöfer, A. Jasmonate signalling in carnivorous plants: copycat of plant defence mechanisms. J. Exp. Bot. 70, 3379–3389 (2019).
-
Nakamura, Y., Reichelt, M., E Mayer, V. & Mithöfer, A. Jasmonates trigger prey-induced formation of ‘outer stomach’ in carnivorous sundew plants. Proc. R Soc. B Biol. Sci. 280, 1–6 (2013).
-
Yu, H. et al. Identification of rabbit oviductal fluid proteins involved in Pre-Fertilization processes by quantitative proteomics. Proteomics 19, e1800319 (2019).
-
The UniProt Consortium. UniProt: the universal protein knowledgebase in 2023. Nucleic Acids Res. 51, D523–D531 (2023).
-
Saul, F. et al. Subgenome dominance shapes novel gene evolution in the decaploid pitcher plant nepenthes gracilis. Nat. Plants. 9, 2000–2015 (2023).
-
Dawson, C., Smith, T. & camprotR Processing, analysing and visualising CCP proteomics data. (2023).
-
Leprevost, F. V. et al. Philosopher: a versatile toolkit for shotgun proteomics data analysis. Nat. Methods 17 869–870. https://doi.org/10.1038/s41592-020-0912-y (2020).
-
University of Michigan, Department of Pathology, N. lab. FragPipe. (2024).
-
R Core Team. R: A language and environment for statistical computing. (2022).
-
Ritchie, M. E. et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015).
-
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
-
Goddard, T. D. et al. UCSF chimerax: meeting modern challenges in visualization and analysis. Protein Sci. 27, 14–25 (2018).
-
Berman, H. M. et al. The protein data bank. Nucleic Acids Res. 28, 235–242 (2000).
-
Jumper, J. et al. Highly accurate protein structure prediction with alphafold. Nature 596, 583–589 (2021).
-
Eberhardt, J., Santos-Martins, D., F Tillack, A. & Forli, S. AutoDock Vina 1.2.0: new Docking methods, expanded force field, and python bindings. J. Chem. Inf. Model. 61, 3891–3898 (2021).
-
Rosignoli, S. & Paiardini, A. DockingPie: a consensus Docking plugin for PyMOL. Bioinformatics 38, 4233–4234 (2022).
-
Schrödinger, L. L. C. The PyMOL molecular graphics system. (2015).
-
Hanwell, M. D. et al. Avogadro: an advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 4, 17 (2012).
-
Wang, J., Wolf, R. M., Caldwell, J. W., Kollman, P. A. & Case, D. A. Development and testing of a general amber force field. J. Comput. Chem. 25, 1157–1174 (2004).
-
Sonnendecker, C. et al. Low carbon footprint recycling of post-consumer PET plastic with a metagenomic polyester hydrolase. ChemSusChem 15, (2022).
-
Adasme, M. F. et al. PLIP 2021: expanding the scope of the protein–ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 49, W530–W534 (2021).
-
Siracusa, C. et al. Activity of an anaerobic thermoanaerobacterales hydrolase on aliphatic and aromatic polyesters. 1–10. https://doi.org/10.3389/fbioe.2024.1520680 (2025).
-
Zumstein, M. T. et al. Enzymatic hydrolysis of polyester thin films at the nanoscale: effects of polyester structure and enzyme Active-Site accessibility. Environ. Sci. Technol. 51, 7476–7485 (2017).
-
Jieyuan, J., Kolawole, J. & Kevin, M. O. C. Non-Biased enrichment does not improve quantitative proteomic delineation of reovirus T3D-Infected HeLa cell protein alterations. Front. Microbiol. 3, (2012).
-
Liao, P., Li, Y., Li, H. & Liu, W. Organellar proteome analyses of ricin toxin-treated HeLa cells. Toxicol. Ind. Health. 32, 1166–1178 (2016).
-
Guo, X. et al. A multiprotease map of the HeLa proteome for comprehensive Proteomics *. Mol. Cell. Proteom. 13, 1573–1584 (2014).
-
Cox, J. & Mann, M. Is Proteom. New. Genomics? Cell 130, 395–398 (2007).
-
Huang, Z. et al. Proteomic datasets of HeLa and SiHa cell lines acquired by DDA-PASEF and diapasef. Data Br. 41, 107919 (2022).
-
Bekalu, Z. E., Dionisio, G. & Brinch-Pedersen, H. Molecular properties and new potentials of plant nepenthesins. Plants 9, 570 (2020).
-
Rich, D. H. et al. Inhibition of aspartic proteases by Pepstatin and 3-methylstatine derivatives of Pepstatin. Evidence for collected-substrate enzyme Inhibition. Biochemistry 24, 3165–3173 (1985).
-
Tournier, V. et al. An engineered PET depolymerase to break down and recycle plastic bottles. Nature 580, 216–219 (2020).
-
Crnjar, A., Griñen, A., Kamerlin, S. C. L. & Ramírez-Sarmiento, C. A. Conformational selection of a Tryptophan side chain drives the generalized increase in activity of PET hydrolases through a ser/ile double mutation. ACS Org. Inorg. Au. 3, 109–119 (2023).
-
Sulaiman, S. et al. Isolation of a novel cutinase homolog with polyethylene Terephthalate-Degrading activity from Leaf-Branch compost by using a metagenomic approach. Appl. Environ. Microbiol. 78, 1556–1562 (2012).
-
Richter, P. K. et al. Structure and function of the metagenomic plastic-degrading polyester hydrolase PHL7 bound to its product. Nat. Commun. 14, 1905 (2023).
-
Barclay, A. & Acharya, K. R. Engineering plastic eating enzymes using structural biology. Biomolecules 13, 1407 (2023).
-
Dunn, B. M. Structure and mechanism of the Pepsin-Like family of aspartic peptidases. Chem. Rev. 102, 4431–4458 (2002).
-
Bose, I. & Zhao, Y. Selective hydrolysis of Aryl esters under acidic and neutral conditions by a synthetic aspartic protease mimic. ACS Catal. 11, 3938–3942 (2021).
-
Lim, H. A., Raku, T. & Tokiwa, Y. Hydrolysis of polyesters by Serine proteases. Biotechnol. Lett. 27, 459–464 (2005).
-
Aer, L. et al. Optimization of polyethylene terephthalate biodegradation using a self-assembled multi-enzyme cascade strategy. J. Hazard. Mater. 476, 134887 (2024).
-
Buch, F., Kaman, W. E., Bikker, F. J., Yilamujiang, A. & Mithöfer, A. Nepenthesin protease activity indicates digestive fluid dynamics in carnivorous nepenthes plants. PLoS One. 10, 1–15 (2015).
-
Perez-Riverol, Y. et al. The PRIDE database at 20 years: 2025 update. Nucleic Acids Res. 53, D543–D553 (2025).
Acknowledgements
The COMET center: acib: Next Generation Bioproduction is funded by BMK, BMDW, SFG, Standortagentur Tirol, Government of Lower Austria und Vienna Business Agency in the framework of COMET – Competence Centers for Excellent Technologies. The COMET-Funding Program is managed by the Austrian Research Promotion Agency FFG. This research was also funded by the European Union’s Horizon2020 research and innovation program under grant agreement No 953073 and the project UPLIFT (sUstainable PLastIcs forthe Food and drink packaging indusTry).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Siracusa, C., Gritsch, S., Vielnascher, R. et al. Carnivorous plants can decompose the polyesters poly(ethylene terephthalate) and poly(butylene adipate terephthalate). Sci Rep 15, 29059 (2025). https://doi.org/10.1038/s41598-025-14331-2
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-14331-2