Introduction
Fungal infections have increased over recent decades and now cause substantial global morbidity and mortality. Candida species are among the leading etiologic agents of invasive mycoses. Treatment is hampered by increasing antifungal resistance and by the limited tolerability of current drugs1. The risk increases with broad-spectrum antibiotic exposure, indwelling catheters, total parenteral nutrition, immunosuppression after cancer therapy or transplantation, and disruption of mucosal barriers2.
Among more than 150 Candida species, only a subset can grow at 37 °C, a prerequisite for human pathogenicity3. Pathogenic species use immune evasion, adhesion with biofilm formation on host tissues or devices, and secretion of tissue-damaging enzymes such as proteases, phospholipases and hemolysins to establish infection4,5.
Although Candida albicans (C. albicans) has historically been the most frequently isolated species in both healthy individuals and patients (often exceeding 80% of isolates), the incidence of nonalbicans Candida (NCAC) species has increased significantly over the past two decades6. In particular, Candida glabrata (C. glabrata) accounts for approximately 15% of systemic Candida infections and shows an innate propensity toward antifungal resistance7.
Unlike C. albicans, C. glabrata lacks certain virulence factors, such as hyphal development and protease secretion8. Nevertheless, its high degree of drug resistance and ability to form biofilms make it a formidable pathogen in immunocompromised hosts9. Epidemiological surveys indicate a global increase in the prevalence of C. glabrata, notably in intensive care units (ICUs), where immunocompromised or critically ill patients are most susceptible10.
Another clinically significant species, Candida tropicalis (C. tropicalis), also represents a leading cause of candidaemia and invasive candidiasis, particularly in patients with hematological malignancies, solid tumors, or those undergoing transplantation11. C. tropicalis shares virulence features with C. albicans, including biofilm formation and the secretion of tissue-damaging enzymes such as proteases, phospholipases, and esterases12. Alarmingly, multidrug resistance in C. tropicalis is increasing, with high rates of resistance reported against azoles and, to a lesser extent, echinocandins13. Together, C. glabrata and C. tropicalis exemplify how nonalbicans species are increasingly implicated in life-threatening infections, underscoring the urgent need for novel therapeutic strategies8.
In light of these challenges, innovative drug delivery systems have garnered attention as a means to overcome fungal drug resistance. Chitosan nanoparticles (CS-NPs), derived from the naturally occurring polysaccharide chitin, possess favorable characteristics such as biocompatibility, biodegradability and inherent antimicrobial activity14. Their nanoscale size provides an extensive surface area for the loading of diverse compounds, ranging from classical chemotherapeutic agents (e.g. carboplatin) to repurposed antimicrobials (e.g. ribavirin) and phytochemicals such as Thymus vulgaris (T. vulgaris) extracts15. Encapsulating antifungal or anticancer drugs in CS-NPs can increase their solubility, improve targeted delivery to the pathogen or tumor site and achieve sustained release, thereby reducing systemic toxicity16. Additionally, the cationic nature of CS-NPs promotes electrostatic interactions with negatively charged microbial surfaces, potentially bypassing common resistance mechanisms such as efflux pumps and biofilm barriers17.
Recent research highlights the potential synergy of combining azole antifungals (such as fluconazole) with alternative agents, including ribavirin (RBV), carboplatin, and essential oils derived from T. vulgaris. While RBV is principally recognized for its broad-spectrum antiviral activity, in vitro studies suggest that it also has antifungal effects on certain Candida species and may enhance the efficacy of conventional antifungals18. Similarly, carboplatin, which is widely used in oncology, has some antifungal potential, especially when combined with agents such as amphotericin B or other antifungals19. Moreover, T. vulgaris essential oil (TV EO) has repeatedly been shown to possess potent antimicrobial and antifungal properties, which are attributable to its main active constituents (e.g. thymol and carvacrol), which disrupt fungal membranes and reduce biofilm formation20.
Despite encouraging findings, the use of RBV, carboplatin and T. vulgaris against fungal infections, particularly when combined with fluconazole and formulated as CS-NPs, remains insufficiently studied. This gap is critical, given the urgent clinical need for therapies capable of overcoming the adaptive mechanisms of C. glabrata and C. tropicalis8. Notably, genes such as SNQ2 and ERG11 (in C. glabrata) and CDR1 and MDR1 (in C. tropicalis) are implicated in antifungal resistance, making them prime targets for understanding how novel treatments may circumvent drug efflux or drug-target modifications. SNQ2 encodes an ATP-binding cassette transporter that exports antifungal agents, reducing intracellular concentrations and promoting azole resistance21,22. ERG11 encodes lanosterol 14α-demethylase, the primary azole target. Mutations in ERG11 can reduce drug binding, although in C. glabrata, they occur less often than resistance driven by transporter overexpression23,24. In C. tropicalis, overexpression of the ATP-binding cassette (ABC) transporter gene CDR1 facilitates fluconazole resistance via efflux. In contrast, the major facilitator superfamily (MFS) transporter MDR1 more consistently confers high-level resistance, particularly in conjunction with ERG11 mutations25,26,27.
CS-NPs show antifungal activity through several complementary mechanisms. Their cationic surface binds to the negatively charged fungal wall and membrane, which increases permeability and damages the envelope. They also disrupt biofilms and can blunt efflux-associated azole resistance, which increases intracellular drug levels28,29. The major constituents of T. vulgaris essential oil, thymol and carvacrol, destabilize fungal membranes in a ergosterol-dependent manner. These compounds inhibit biofilm formation and can act synergistically with licensed antifungals30,31,32,33. RBV has adjunct anti-Candida activity in vitro, where it reduces virulence traits and enhances azole efficacy. Anticancer agents, including platinum drugs, can impede Candida morphogenesis, which provides a rationale to test carboplatin in nanoparticle form34.
To our knowledge, this is the first evaluation of RBV, carboplatin, and TV EOs formulated as chitosan nanoparticles against clinical isolates of fluconazole-sensitive and fluconazole-resistant C. tropicalis and fluconazole-resistant C. glabrata. Previous work reported the antifungal and azole-enhancing effects of ribavirin, mainly in C. albicans, but not in CS-NP systems18. Thymol and carvacrol have been loaded into CS-NPs with antifungal activity35, but systematic testing of whole TV EO, ribavirin, or carboplatin in this platform against C. glabrata and C. tropicalis is lacking. The antifungal potential of carboplatin has been noted, but its use in CS-NP delivery has not been studied.
Here, we assessed the effects of CS-NP-encapsulated ribavirin (CS-R NPs), carboplatin (CS-C NPs), and T. vulgaris essential oil (CS-TV NPs) on fluconazole-stratified isolates via phenotypic susceptibility and gene expression readouts. The goal is to determine whether nanoparticle delivery shifts resistance behavior. We focused on C. glabrata SNQ2 and ERG11 and on C. tropicalis CDR1 and MDR1, which represent efflux and target-site mechanisms, respectively, and quantified how each formulation affects these pathways. Integrating these molecular endpoints with activity data addresses a clear translational gap and outlines a strategy to modulate resistance in nonalbicans Candida.
Materials and methods
Isolation and species identification of the required strains
A total of 20 clinical isolates, comprising C. tropicalis (n = 10) and C. glabrata (n = 10), were obtained from the strain bank of Tarbiat Modares University. Antifungal susceptibility profiling was conducted by determining the minimum inhibitory concentration (MIC) of fluconazole for each isolate, following the Clinical and Laboratory Standards Institute (CLSI) M27-A3 and CLSI M60 guidelines for yeast susceptibility testing36. To ensure the reliability of the comparative assessments, standard reference strains of C. tropicalis and C. glabrata PFCC 901847 were included as controls throughout the study. For antifungal testing, fluconazole was dissolved in DMSO (dimethyl sulfoxide) at a concentration of 2 mg/mL and subsequently diluted to final concentrations ranging from 128 to 0.25 µg/mL.
Culture of Candida isolates
Candida isolates were cultured on Sabouraud dextrose agar (SDA, Merck, Inc., Germany), prepared according to the manufacturer’s guidelines, and sterilized by autoclaving at 121 °C for 15 min. To prevent bacterial contamination, chloramphenicol (Sigma‒Aldrich, USA) was added to the SDA at a final concentration of 100 µg/mL.
Isolates were retrieved from the strain repository and suspended in sterile saline to achieve a cell density equivalent to the 0.5 McFarland standard (approximately 1.5 × 10⁸ CFU/mL). A 100 µL aliquot of each suspension was uniformly inoculated onto SDA plates via the streak plate technique to ensure isolated colony formation. The inoculated plates were incubated at 35 °C for 24–48 h to promote optimal fungal growth.
Preparation of CS-NPs
CS-R NPs, CS-C NPs and CS-TV NPs were synthesized via the ionic gelation method37. Briefly, chitosan (CS) with an average molecular weight of 50–190 kDa and a degree of deacetylation of 75–85% was dissolved alongside RBV, carboplatin, and Thymus vulgaris essential oil (TV EO) in 5 mM HEPES buffer (pH 7.4; Merck, Inc., Germany). The pH of the resulting CS solution was adjusted to 4.7–4.8 using 2 M sodium hydroxide, and the resulting solution was subsequently filtered through a 0.45 μm syringe filter to eliminate any undissolved particulates. The final concentrations of CS and the encapsulated compounds were standardized at 1 and 0.1 mg/mL, respectively. To ensure uniform dispersion and mitigate aggregation, the prepared solutions were continuously stirred at 300 rpm for 18 h via a magnetic stirrer.
Ionic crosslinking was initiated by introducing a pentasodium tripolyphosphate (TPP) solution, which was prepared by dissolving 20 mg of TPP (Merck, Inc., Germany) in 5 mL of 5 mM HEPES buffer (pH 7.4), yielding a 4 mg/mL solution. This formulation maintained a CS-to-TPP weight ratio of 5:1. The TPP solution was added dropwise to the CS–drug mixture under continuous stirring, facilitating NP formation via electrostatic interactions between the protonated amine groups of CS and the phosphate moieties of TPP. Although the reaction volume increased slightly to approximately 105 mL, this dilution had no detrimental impact on NP formation.
Following crosslinking, the NPs were isolated via high-speed centrifugation at 15,000 × g for 30 min, effectively separating the synthesized NPs from unbound components. The resulting pellet was resuspended in 10 mL of deionized water and subjected to further purification by overlaying onto a 10% (v/v) glycerol solution, followed by centrifugation at 10,000 × g for 15 min. This step removed residual unbound components and prevented NP aggregation. The purified NPs were subsequently lyophilized and stored as a dry powder at 4 °C for subsequent physicochemical characterization, drug release kinetics analysis, and bioactivity assessment.
Encapsulation efficiency
The encapsulation efficiency (EE%) of the CS-R NPs, CS-C NPs, and CS-TV NPs was assessed by quantifying the unencapsulated fraction of each drug present in the supernatant following centrifugation at 24,000 × g and 4 °C for 45 min. The concentration of the unencapsulated drug was determined spectrophotometrically at 475 nm, and the amount of encapsulated drug was subsequently calculated via the following equation:
$${text{Encapsulation efficiency }}left( % right), = ,left[ {left( {{text{Total initial drug amount}} – {text{Amount of drug released in the supernatant}}} right)/{text{Total initial drug amount}}} right], times ,{1}00.$$
Particle size and zeta potential analysis
The hydrodynamic diameter and zeta potential of the CS-NPs were determined via dynamic light scattering (DLS) and electrophoretic light scattering (ELS) via a Zetasizer system (Malvern Panalytical Zetasizer Nano). The measurements were conducted at 25 °C to ensure accuracy, and all samples were appropriately diluted in HEPES buffer (pH 7.4) prior to analysis to prevent multiple scattering effects.
Morphological analysis
The morphology and surface structure of the NPs were examined via transmission electron microscopy (TEM). A droplet of the suspension was deposited onto a carbon film-coated copper grid (200 mesh) via a micropipette. The sample was allowed to rest undisturbed for 5 min to facilitate adequate adsorption onto the carbon substrate. Following this incubation period, excess liquid was delicately removed by gently contacting the edge of the grid with a piece of filter paper, minimizing potential disruption to the sample. The grid was subsequently left to air dry at ambient temperature under controlled conditions to prevent structural alterations. The prepared sample was then subjected to TEM analysis.
Fibroblast culture and maintenance
Human foreskin fibroblasts were cultured in high-glucose Dulbecco’s modified Eagle’s medium (DMEM, Gibco, Inc., USA) supplemented with 10% fetal bovine serum (FBS, Gibco, Inc., USA) and 1% penicillin‒streptomycin (Gibco, Inc., USA). Prior to cell seeding, all culture reagents, including DMEM and FBS, were prewarmed to 37 °C. The cells were maintained at 37 °C in a humidified incubator with 5% CO₂.
Cytotoxicity evaluation of the CS-NPs via the MTT assay
The cytotoxic effects of CS-NPs on fibroblasts were quantitatively assessed via the MTT assay (Sigma‒Aldrich, Merck, Inc., Germany). Fibroblasts were seeded into 96-well plates at a density of 1 × 104 cells per well and incubated at 37 °C in a humidified atmosphere containing 5% CO₂ until a monolayer was established. Upon cell attachment, the culture medium was aspirated, and the cells were exposed to various concentrations of CS-R NPs, CS-C NPs and CS-TV NPs. Each concentration was tested in triplicate, ensuring statistical robustness. The control groups included untreated cells (negative control) incubated with serum-free medium. The cells were further incubated under standard conditions (37 °C, 5% CO₂) for 24 h to assess cytotoxicity. The 24 h endpoint aligned with the antifungal assay window, captured early cytotoxic responses, and reduced artifacts from nutrient depletion. At this time point, 10 µL of MTT solution (5 mg/mL) was added to each well, and the plate was incubated for an additional 4 h at 37 °C to facilitate formazan crystal formation. After incubation, the medium containing unreacted MTT was carefully aspirated, and 100 µL of dimethyl sulfoxide (DMSO) was added to each well to dissolve the intracellularly formed formazan crystals. The plate was gently agitated for 5–10 min in the dark at room temperature to ensure complete dissolution. The absorbance of the solubilized formazan product was recorded at 570 nm, with a reference wavelength of 630 nm, via a Bio-Rad 680 microplate reader (Bio-Rad Laboratories, USA). The percentage of viable cells relative to the untreated control was calculated via the following equation:
$${text{Cell viability}}left( % right), = ,left( {{text{OD control}}/{text{OD sample}}} right), times ,{1}00.$$
The CS-R NPs, CS-C NPs, and CS-TV NPs were dissolved in sterile water and tested in the concentration range of 10–0.0195 µg/mL to assess fibroblast cytotoxicity at therapeutically relevant exposures.
Assessment of drug release
The in vitro release profiles of the CS-R NPs, CS-C NPs and CS-TV NPs were investigated by incubating the NP suspensions in phosphate-buffered saline (PBS; pH 7.4) at 37 °C under controlled humidity. At 2, 4, 8, 16, 32 and 64 h, aliquots were withdrawn and immediately replaced with fresh PBS to maintain sink conditions. The concentration of the released drug in each collected sample was then quantified via UV–Vis spectrophotometry at 475 nm, employing a previously established standard calibration curve. The cumulative drug release was used to assess the release kinetics and derive key parameters such as the half-life of release and the time required for the complete discharge of the drug from the NP matrix.
Drug susceptibility testing
A standardized microdilution assay was conducted in 96-well microplates to assess the antifungal activity of carboplatin, RBV, TV EO, CS-R NPs, CS-C NPs and CS-TV NPs, with itraconazole employed as a positive control. Briefly, 100 μL of each stock solution was dispensed into the first well of each row, followed by serial twofold dilutions across subsequent wells. Candida cells in the logarithmic growth phase were harvested, resuspended in RPMI-1640 medium (Gibco, Inc., USA), and adjusted to a final inoculum concentration of 1–2 × 103 CFU/mL by adding 900 μL of this inoculum to each well containing the test compounds.
TV EO was dissolved in 2% Tween 80 and serially diluted to final concentrations ranging from 1024 to 2 µg/mL, while carboplatin was dissolved in sterile water at a concentration of 14 mg/mL and subsequently diluted to final concentrations ranging from 400 to 0.781 µg/mL. RBV was dissolved in sterile water at a concentration of 49 mg/mL and serially diluted to final concentrations ranging from 128 to 0.25 µg/mL for antifungal testing.
A sterile control well containing the test compounds without fungal inoculum was included to ensure the absence of contamination; a growth control well containing only the fungal inoculum without any test compound served to confirm the viability of the organism. The plates were incubated at 35 °C for 48 h, after which they were examined for turbidity. The MIC was defined as the lowest concentration of the test compound at which no visible growth (no turbidity) was observed. For quality assurance, C. albicans ATCC 10,231 and C. glabrata PFCC 901,847 were utilized as reference strains.
Dose ranges were set via CLSI M27-A3 and M60 guidance, compound solubility, and preliminary range-finding. The upper limits prevented precipitation and vehicle effects. The lower limits capture the expected MIC values.
We used itraconazole as the reference azole because it is clinically deployed, retains activity against C. glabrata and C. tropicalis in many contexts, and has well-defined resistance mechanisms that include efflux pump upregulation and ERG11 alterations. This makes it a suitable mechanistic comparator for resistance-modulation studies38,39,40.
Selectivity index (SI) calculation
The selectivity index (SI) was calculated to assess the therapeutic window of each nanoparticle formulation relative to its antifungal activity. The SI was defined as the ratio of the IC₅₀ value obtained from the fibroblast cytotoxicity assay to the MIC₅₀ value determined against each Candida strain via the following formula:
$${mathbf{SI}} = {text{ IC5}}0_{{left( {{text{fibroblasts}}} right)}} /{text{MIC5}}0_{{({text{Candida}})}} .$$
RNA extraction and cDNA synthesis
Total RNA was isolated via an RNX-Plus kit (Sinaclon, Iran) according to the manufacturer’s instructions. The quantity and purity of the extracted RNA were initially confirmed via 1% (w/v) agarose gel electrophoresis and further verified via a NanoDrop spectrophotometer (Thermo Fisher Scientific, USA).
cDNA synthesis was performed via a Sinaclon cDNA Synthesis Kit (Sinaclon, Iran) following the standard protocol. In brief, 1 µg of total RNA was reverse transcribed in a 10-µL reaction mixture containing dNTPs, oligo(dT), and random hexamers. The resulting cDNA was stored at − 20 °C until further experimentation.
Quantitative real-time PCR analysis
Quantitative real-time PCR (qPCR) was conducted via a SYBR Green-based system (RealQ Plus 2 × Master Mix Green, Amplicon) on a Roche LightCycler 96 platform (Roche Diagnostics, Germany). Each 15 µL reaction consisted of 7.5 µL of 2 × Master Mix, 0.1 µL of forward primer (10 µM), 0.1 µL of reverse primer (10 µM), 0.1 µL of cDNA template (adjusted on the basis of RNA input), and 0.4 µL of RNase-free water. The thermal cycling conditions included an initial denaturation step at 95 °C for 15 min, followed by 40 cycles of 95 °C for 15 s (denaturation) and 60 °C for 40 s (annealing/extension). Melt curve analysis was performed from 55 °C to 95 °C to confirm the specificity of amplification. Relative gene expression levels were determined via the Pfaffl method, with normalization against the housekeeping gene ACT1, and the control group was used as a reference for comparative analysis. Gene expression experiments used exposure levels close to the species-specific MIC50 for each free compound and its matched CS-NP formulation. This provided sufficient antifungal pressure to elicit transcriptional responses while preserving RNA integrity. All reactions were performed in triplicate to ensure reproducibility. The primers used in this study are listed in Table 1.
Statistical analysis
Data analysis was performed via SPSS version 27 (IBM, USA) and GraphPad Prism version 10 (GraphPad Software, USA). The normality of the data distribution was assessed via the Kolmogorov‒Smirnov test, with p values < 0.05 considered statistically significant. For normally distributed data, the results are presented as the means ± standard deviations (SDs) and were visualized via bar charts. IC50 values were calculated via nonlinear regression analysis in GraphPad Prism. Two-way ANOVA, followed by Dunnett’s multiple comparisons test, was used to compare differences between experimental groups. All experiments were performed in triplicate, with measurements taken at two distinct time points to ensure the reproducibility and robustness of the data.
Results
Species identification and antifungal susceptibility of Candida isolates
The antifungal susceptibility of Candida species, specifically C. glabrata and C. tropicalis, to fluconazole was assessed through MIC values. For C. glabrata, the MIC values demonstrated an MIC50 of 64 µg/mL and an MIC90 of 128 µg/mL, with resistance ranging from 64 to 128 µg/mL. Notably, all 10 C. glabrata strains tested exhibited complete resistance to fluconazole (100%). In contrast, for C. tropicalis, the MIC50 was 16 µg/mL, and the MIC90 was 32 µg/mL, with a resistance range from 1 to 32 µg/mL. Among the strains tested, 9 out of 10 (90%) exhibited resistance to fluconazole, whereas only one strain (10%) was classified as sensitive. Furthermore, both the standard strains of C. glabrata and C. tropicalis exhibited significant resistance (Table 2).
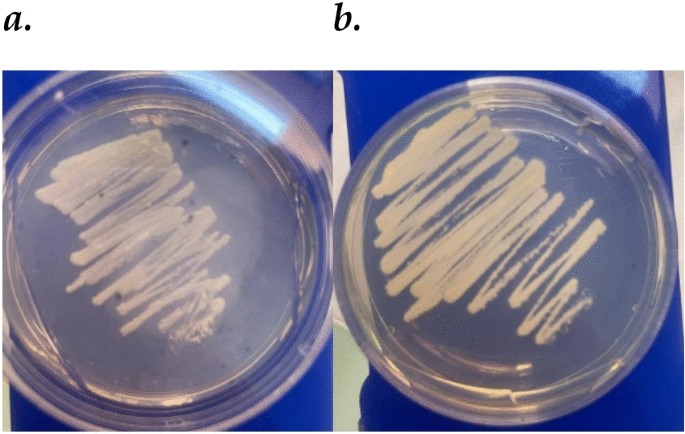
The morphological characteristics of Candida species isolates were assessed after culture on SDA at 35 °C for 48 h. C. tropicalis formed moist, creamy, off-white colonies with raised, convex, and occasionally irregular edges. Colonies grew rapidly and expanded several millimeters, and microscopic analysis revealed ovoid to elongated yeast cells with budding and occasional pseudohyphal formation. In contrast, C. glabrata produced smaller, more compact colonies with a smooth, shiny, translucent appearance. Its growth was rapid but less extensive. Microscopic examination revealed spherical to ovoid yeast cells with minimal pseudohyphal formation. Both species exhibited rapid growth, but their distinct colony and microscopic features allowed for differentiation, with C. tropicalis showing pronounced pseudohyphal growth and C. glabrata maintaining a uniform yeast form (Fig. 1).

Morphological characteristics of Candida species ((a) C. glabrata; (b) C. tropicalis) isolates on SDA after 48 h at 35 °C.
Physicochemical characterization of CS-NPs
Encapsulation efficiency
The encapsulation efficiency (EE%) of RBV, carboplatin, and TV EO within the CS-NPs was assessed, with the following outcomes: the CS-R NPs presented an EE% of 86% (± 0.7), the CS-C NPs presented an EE% of 81% (± 0.5), and the CS-TV NPs presented the highest EE% of 91% (± 0.6).
Particle size and zeta potential
The CS-TV NPs, CS-C NPs, and CS-R NPs presented hydrodynamic diameters of approximately 298 ± 10.13 nm, 288 ± 11.32 nm and 286 ± 8.27 nm, respectively, as determined by DLS. Furthermore, ELS measurements revealed zeta potentials of + 33 ± 2.14 mV for CS-TV NPs, + 34 ± 1.17 mV for CS-C NPs, and + 34 ± 3.18 mV for CS-R NPs (Table 3).
Morphological analysis of the NPs
TEM was performed to investigate the morphology and structural characteristics of the CS-TV NPs, CS-C NPs and CS-R NPs further. As shown in Fig. 2, the TEM images confirmed the formation of well-dispersed, spherical NPs with smooth surfaces. The observed particle sizes were slightly smaller than the hydrodynamic diameters measured by DLS, which is expected due to the absence of a hydration layer in TEM imaging. Compared with the CS-C NPs and CS-R NPs, the CS-TV NPs appeared to have a more uniform size distribution. Additionally, no significant aggregation was observed, suggesting good stability of the synthesized NPs.

TEM images of CS-NPs loaded with TV EO (a), carboplatin (b), and RBV (c).
Cytotoxicity of CS-NPs to fibroblast cells
The cytotoxicity of the CS-R NPs, CS-C NPs and CS-TV NPs at 24 h was assessed via their IC50 values. The IC50 of the CS-R NPs was 1.42 µg/ml (95% CI 1.25 to 1.60), indicating moderate cytotoxicity. CS-C NPs had an IC50 of 0.79 µg/ml (95% CI 0.72 to 0.85), demonstrating strong cytotoxicity at lower concentrations. Compared with the other treatments, the CS-TV NPs had the highest IC50 of 20.31 µg/ml (95% CI 17.36 to 24.55), suggesting a weaker cytotoxic effect. The goodness of fit was high for all the treatments, with R-squared values above 0.97 (Fig. 3). The control group exhibited high cell viability, with no significant cytotoxicity observed, confirming the reliability and accuracy of the experimental findings.

MTT assay evaluating the cytotoxicity of CS-NPs loaded with RBV, carboplatin, and TV EO on fibroblasts at 24 h. The graph displays the cell viability (%) at different concentrations (µg/ml) of CS-R NPs (●), CS-C NPs (■), and CS-TV NPs (▲). A dose-dependent decrease in cell viability was observed for all the treatments, with the CS-C NPs resulting in the lowest cell viability at higher concentrations.
In vitro drug release profile
The cumulative release profiles of the three CS-NP formulations (CS-R NPs, CS-C NPs and CS-TV NPs) are shown in Fig. 4. The data demonstrate a time-dependent increase in drug release, with measurements taken at 2, 4, 8, 16, 32 and 64 h and three replicates for each time point (N = 3). At 64 h, the CS-R NPs presented a mean cumulative release of 52.33% (SD = 1.53), whereas the CS-C NPs presented a mean release of 53.33% (SD = 4.73). In contrast, the CS-TV NPs presented the greatest cumulative release, reaching a mean of 60.00% (SD = 1.00) at the same time point (Fig. 4).

Cumulative drug release profile of CS-R NPs (●), CS-C NPs (■), and CS-TV NPs (▲) over 64 h. The data show the release kinetics of the three formulations, with CS-TV NPs exhibiting the highest cumulative release, followed by CS-C NPs and CS-R NPs, indicating differential release behavior. The results are expressed as a percentage of cumulative release over time (hours).
Antifungal activity of encapsulated drugs
The antifungal efficacy of RBV, carboplatin, TV EO and CS-NPs, including CS-R NPs, CS-C NPs, and CS-TV NPs, was assessed against C. glabrata and C. tropicalis strains. In C. glabrata, RBV presented an MIC50 of 8 µg/mL and an MIC90 of 16 µg/mL, whereas carboplatin presented an MIC50 of 25 µg/mL and an MIC90 of 50 µg/mL. TV EO had an MIC50 of 64 µg/mL and an MIC90 of 128 µg/mL. In comparison, the CS-R NPs, CS-C NPs and CS-TV NPs presented significantly lower MIC50 values of 0.078, 0.156 and 1.25 µg/mL, respectively, and MIC90 values of 0.156, 0.3125 and 2.5 µg/mL, respectively (Table 4).
For C. tropicalis, RBV demonstrated a notably lower MIC50 (4 µg/mL) and MIC90 (8 µg/mL) than in C. glabrata. Carboplatin yielded MIC50 and MIC90 values of 25 and 50 µg/mL, respectively. TV EO had identical MIC50 and MIC90 values of 64 and 128 µg/mL, respectively. Notably, the CS-NP formulations again exhibited superior antifungal activity, with MIC50 values of 0.078, 0.156 and 0.625 µg/mL and MIC90 values of 0.156, 0.3125 and 1.25 µg/mL for the CS-R NPs, CS-C NPs and CS-TV NPs, respectively (Table 4).
Fibroblast cytotoxicity is reported as the IC₅₀ (µg/mL) at 24 h. The selectivity index (SI) was calculated as the IC₅₀ in fibroblasts divided by the MIC₅₀ for each nanoparticle formulation.
Selectivity index
The SI was calculated to compare the cytotoxicity of the nanoparticle formulations in fibroblasts with their antifungal activity against Candida spp. (Table 4). Among the tested formulations, CS-TV NPs presented the highest SI values, with SI ≈ 16.2 for C. glabrata and 32.5 for C. tropicalis. The CS-R NPs had moderate SI values of approximately 18.2 for both species. In contrast, the CS-C NPs presented lower SI values (≈ 5.1 for both species), mainly because of their relatively low IC₅₀ in fibroblasts.
Gene expression analysis of treated Candida isolates
Figure 5a presents the relative SNQ2 gene expression in C. glabrata following various treatment regimens, normalized to a control value of 1.0. Among the free compounds, RBV (8 µg/mL) had the greatest downregulatory effect (0.3233 ± 0.02517), followed by carboplatin (25 µg/mL; 0.4600 ± 0.03606) and TV EO (64 µg/mL; 0.5433 ± 0.03512). Notably, the CS-NP formulations—CS‐R, CS‐C, and CS‐TV—further reduced SNQ2 expression (0.0620 ± 0.002646, 0.07533 ± 0.005033 and 0.1000 ± 0.01000, respectively), underscoring the augmented suppression of this gene achieved by nanoencapsulation.

Relative expression levels of SNQ2 (a) and ERG11 (b) in C. glabrata and of CDR1 (c) and MDR1 (d) in C. tropicalis strains treated with RBV, carboplatin, TV EO, CS-R NPs, CS-C NPs, and CS-TV NPs.
The relative ERG11 gene expression under the same treatments is highlighted in Fig. 5b. CS‐R elicited the most pronounced inhibition (0.072 ± 0.003), followed closely by CS‐C (0.084 ± 0.003). Moreover, compared with the control, CS‐TV significantly downregulated ERG11 expression (0.210 ± 0.020). Among the free formulations, carboplatin (0.343 ± 0.035) and RBV (0.417 ± 0.006) yielded moderate reductions in ERG11, whereas TV EO alone had the smallest effect (0.460 ± 0.036).
Figure 5c shows the relative CDR1 gene expression in C. tropicalis following exposure to either free compounds or their CS-NP formulations, normalized to a control value of 1.0. Among the free treatments, RBV (4 µg/mL) resulted in the most pronounced reduction in CDR1 (0.2533 ± 0.03055), followed by carboplatin (25 µg/mL; 0.3500 ± 0.03606) and TV EO (64 µg/mL; 0.4233 ± 0.03215). In contrast, the CS-NPs had a substantially stronger downregulatory effect, with the CS‐R NPs achieving the greatest decrease (0.01533 ± 0.003512), followed by the CS‐C NPs (0.04300 ± 0.003606) and CS‐TV NPs (0.05333 ± 0.002082).
The relative MDR1 gene expression in C. tropicalis under the same set of treatments is depicted in Fig. 5d. Among the free compounds, carboplatin (25 µg/mL) elicited the greatest suppression (0.4400 ± 0.01732), followed by RBV (4 µg/mL; 0.5800 ± 0.01000) and TV EO (64 µg/mL; 0.6467 ± 0.03786). However, nanoparticle (NP) formulations consistently outperformed their free counterparts, yielding notably lower MDR1 levels. Both the CS‐R NPs and the CS‐C NPs achieved robust downregulation (0.08867 ± 0.003215 and 0.08833 ± 0.003055, respectively), whereas the CS‐TV NPs also markedly reduced this value (0.1900 ± 0.04000).
Discussion
The present study evaluated the impact of CS-NPs loaded with RBV, carboplatin, and TV EO on the expression of SNQ2 and ERG11 in resistant C. glabrata and of CDR1 and MDR1 in resistant and sensitive C. tropicalis strains. Antifungal susceptibility testing revealed significant resistance patterns among the clinical isolates. All tested C. glabrata strains (100%) demonstrated complete resistance to fluconazole, with MIC50 and MIC90 values of 64 µg/mL and 128 µg/mL, respectively. The C. tropicalis strains presented slightly lower resistance (90%), with MIC50 and MIC90 values of 16 µg/mL and 32 µg/mL, respectively. These findings underscore the urgent clinical challenge posed by high antifungal resistance rates, particularly against fluconazole, supporting the necessity of exploring alternative antifungal strategies.
The increasing incidence of candidiasis, coupled with the emergence of drug-resistant strains, has been a major global health challenge in recent decades41. In addition to high morbidity and mortality rates, drug resistance exacerbates treatment difficulties in Candida species. Specifically, resistance to fluconazole in C. glabrata and C. tropicalis highlights the urgency of developing alternative treatments for these pathogens, as observed in previous studies42,43.
In our study, the physicochemical characterization of CS-NPs indicated successful synthesis, efficient drug encapsulation, and favorable physical properties. The NPs formed through electrostatic interactions between positively charged CS and negatively charged TPP were stable and well dispersed37. The particle size measurements ranged from 286 to 298 nm, and the strongly positive zeta potentials (+ 33 to + 34 mV) indicated formulation stability. These properties are crucial for NP-based drug delivery, ensuring prolonged circulation and controlled drug release44. These findings align with previous results where zeta potential values above + 30 mV contributed to enhanced suspension stability, preventing aggregation and ensuring uniform NP distribution45,46. Additionally, the size of CS-NPs can be influenced by the concentration and ratio of CS to TPP, with higher CS concentrations leading to larger NP sizes due to increased molecular interactions and cross-linking density47,48. The stability and uniform size distribution observed here further support the suitability of these formulations for drug delivery applications. Although the nanoparticles had a mean hydrodynamic diameter near 300 nm, this range is consistent with the mucoadhesive polymer carriers used in biomedical delivery. FDA guidance treats materials at the nanoscale when size-dependent behavior is present even above 100 nm and up to approximately 1 µm. Reports of chitosan mucoadhesive nanoparticles commonly fall between 200 and 500 nm49,50.
Analysis of the physicochemical properties suggested that the effectiveness of CS-NPs relies on their size, surface charge, and stability. NPs in the 150–200 nm range often exhibit increased bioavailability and enhanced cellular uptake51, which may explain the improved therapeutic effects observed in the present study. The aggregation behavior of NPs during storage is another critical factor for practical application; in this study, the NPs remained stable, with minimal increases in size over time, indicating their potential for long-term storage and use52.
The EE% was notably high for all formulations, particularly for CS-TV NPs (91%), suggesting effective encapsulation of TV EO. TEM analyses further confirmed the formation of uniformly spherical NPs with smooth surfaces. These characteristics are consistent with previous reports on CS-NPs used for drug encapsulation53,54. Morphology can significantly impact NP stability and interactions with biological membranes, influencing drug release kinetics and therapeutic efficacy55,56. The prolonged release kinetics observed are likely due to the strong electrostatic interactions between CS and the encapsulated agents, a phenomenon described in similar NP systems45,57.
Consistent with prior studies, CS-NPs often display a spherical morphology, appropriate size ranges, and high drug encapsulation potential. For example, Knaul et al. (1999) reported that CS-TPP NPs can exhibit a broader FTIR peak, suggesting increased hydrogen bonding, which is critical for enhancing drug loading efficiency and stability58.
Cytotoxicity assays on fibroblasts demonstrated significant differences in biocompatibility among the formulations. Compared with the CS-R NPs (IC50 = 1.42 µg/mL) and CS-C NPs (IC50 = 0.79 µg/mL), the CS-TV NPs exhibited the lowest cytotoxicity (IC50 = 20.31 µg/mL), indicating superior safety. This finding is particularly relevant because NP size and surface charge can influence toxicity, with smaller, positively charged particles often showing increased toxicity and improved cellular interactions59,60. Previous studies have also demonstrated the low toxicity of CS-based NPs, making them promising candidates for clinical applications61. Notably, the minimal toxicity of TPP confirmed the safety of the CS-NP formulations, especially when the TPP concentration was carefully controlled62. Together, these results suggest that CS-TV NPs could offer a safer alternative for antifungal therapy, reducing adverse effects while maintaining therapeutic potency. This observation aligns with studies indicating that certain essential oil-based formulations exhibit lower toxicity and strong antimicrobial effects, making them promising for therapeutic applications63.
Although some NP formulations containing polycations can cause necrotic cell membrane damage and high cytotoxicity64, our results show that CS-based NPs maintain both biocompatibility and antifungal potency, lowering the risk of adverse effects.
In vitro drug release profiles revealed a sustained, controlled release pattern, with CS-TV NPs showing the highest cumulative drug release (60%) after 64 h compared with that of CS-R NPs (52.33%) and CS-C NPs (53.33%). This biphasic release, an initial burst followed by sustained release, has been widely reported for CS-NPs65,66. This profile increases therapeutic efficacy while reducing dosing frequency, potentially minimizing side effects linked to conventional antifungal drugs67.
Antifungal efficacy studies revealed that nanoencapsulation significantly enhanced antifungal activity against both Candida species. Compared with the free drugs, the CS-NP formulations considerably reduced the MIC50 and MIC90 values. For example, CS-R NPs presented much lower MIC50 values (0.078 µg/mL) than free RBV (ranging from 4 to 8 µg/mL), which aligns with the known capacity of CS-NPs to improve drug solubility and stability, facilitating more effective intracellular drug accumulation68. Such a reduction in MIC values underscores the ability of CS-NPs to overcome fungal resistance mechanisms, a critical advantage for treating multidrug-resistant fungal infections.
A key component of the enhanced antifungal activity of CS-TV NPs is TV EO, which is rich in bioactive compounds (e.g. thymol and carvacrol). Prior work has demonstrated that these phenolic compounds have potent antifungal effects on various pathogenic fungi, including Candida species69. Their antifungal action is partly attributed to interference with fungal cell wall enzymes (e.g. chitin synthase and glucanases), resulting in cell wall disruption70. Moreover, thymol and carvacrol can interact with the fungal cytoplasmic membrane, increasing its permeability and triggering leakage of cellular contents, leading to cell death71,72. Thymol also inhibits the growth of azole- and amphotericin B–resistant Candida, highlighting its efficacy against drug-resistant infections73,74. Notably, thymol can disrupt biofilm formation in Candida, particularly in sessile forms, which are relatively resistant to conventional antifungal treatments75. These mechanisms are consistent with our findings, as incorporating TV EO into CS-NPs substantially improved antifungal efficacy against resistant Candida strains.
Our study also identified RBV as a promising candidate for enhancing antifungal activity against C. glabrata and C. tropicalis. Although RBV has shown antifungal effects, especially against C. albicans, C. parapsilosis, and C. tropicalis, its efficacy against C. glabrata remains limited, with MICs over 24.16 µg/mL for most C. glabrata strains76. However, its enhanced effectiveness in overcoming fluconazole resistance in Candida biofilms suggests that RBV is a valuable adjunct therapy for chronic and drug-resistant fungal infections. In particular, RBV appears to inhibit hyphal growth in Candida, reinforcing the need for more research on its antifungal mechanisms18.
Carboplatin demonstrated MIC50 and MIC90 values of 25 µg/mL and 50 µg/mL, respectively, in our study, which aligns with data from Routh et al. who reported complete inhibition of germ tube formation in C. albicans at 50 µg/mL77.
The SI was applied to relate cytotoxicity to antifungal activity. CS-TV NPs presented the highest SI values, indicating a wide therapeutic window and strong selectivity for Candida spp. over fibroblasts. This pattern aligns with their low MIC values and high IC₅₀ values. The CS-R NPs presented moderate SI values, whereas the CS-C NPs presented the lowest values, mainly due to increased cytotoxicity.
Compared with standard azole therapy, where nonalbicans Candida spp. often display efflux-mediated resistance and biofilm tolerance, the CS-NP formulations achieved lower MIC₅₀ values and higher SI values, with the greatest gains for the CS-TV NPs. These results point to a broader in vitro therapeutic window than the corresponding free agents and are consistent with reports that CS-NPs disrupt membranes, inhibit biofilms, and temper efflux phenotypes14,16,17,49. The membrane-active properties of thymol and carvacrol can also offset azole limitations, which supports their use in nanoparticle delivery systems71,78. While in vivo pharmacokinetics remain to be defined, the data suggest that CS-NPs, particularly CS-TV, could serve as adjuncts or alternatives when azoles underperform against C. glabrata and C. tropicalis.
Gene expression analyses provided further mechanistic insight, revealing substantial downregulation of resistance-associated genes following treatment with CS-NPs. In C. glabrata, SNQ2 and ERG11 expression decreased significantly after the CS-R NP, CS-C NP, and CS-TV NP treatments compared with the free compounds. Similarly, in C. tropicalis, CDR1 and MDR1 expression was significantly suppressed by NP formulations, particularly CS-R NPs. These results suggest that NPs can counteract resistance at the molecular level, potentially restoring antifungal susceptibility in resistant strains. Earlier studies have shown that NP-based formulations disrupt efflux pump activity and interfere with fungal resistance pathways79,80. This molecular counteraction aligns with previous findings, in which the inhibition of multidrug transporters was proposed as a strategy to combat antifungal resistance in Candida81,82,83.
The ability of thymol to disrupt fungal cell membranes, possibly by interacting with ergosterol, likely contributes to the reduced expression of SNQ2, ERG11, CDR1, and MDR1 upon NP treatment.84. Additionally, thyme oil has been reported to generate reactive oxygen species (ROS) within fungal cells, impairing essential cellular functions and enhancing antifungal efficacy85. These combined mechanisms reinforce the potential of CS-NPs loaded with RBV, carboplatin, and TV EO as promising strategies against resistant Candida strains.
Conclusion
The results of this study clearly demonstrate that CS-NPs loaded with RBV, carboplatin, and TV EO not only enhance antifungal efficacy but also offer improved biocompatibility, sustained drug release, and significant suppression of fungal resistance gene expression. These findings collectively support the further development and investigation of CS-NPs as promising antifungal therapeutic strategies against resistant Candida infections.
Despite these promising results, this study has several limitations. The in vitro nature of these experiments may not fully replicate the in vivo environment, and further studies are needed to validate these findings in animal models. Additionally, the long-term stability and potential toxicity of the CS-NPs require further investigation. Owing to budget constraints, comprehensive dose‒response testing across multiple isolates was incomplete, and cytotoxicity was assessed at a single 24 h time point.
Future research should include in vivo efficacy and safety studies with pharmacokinetic and biodistribution analyses. Expanded stability testing and multiple time point cytotoxicity and antifungal assays are needed. Mechanistic studies that define how CS-NPs suppress resistance gene expression would add resolution. Evaluation in combination with standard antifungals could reveal additive or synergistic effects and guide translational applications.
Data availability
The dataset underlying this study’s findings can be accessed upon reasonable request to the corresponding author. However, the data’s public availability is restricted due to ethical considerations and privacy protections.
References
-
Mudenda, S. Global burden of fungal infections and antifungal resistance from 1961 to 2024: Findings and future implications. Pharmacol. Pharm. 15(4), 81–112 (2024).
-
Rayens, E., Norris, K. A. & Cordero, J. F. Mortality trends in risk conditions and invasive mycotic disease in the United States, 1999–2018. Clin. Infect. Dis. 74(2), 309–318 (2022).
-
Vila, T. et al. Oral candidiasis: A disease of opportunity. J. Fungi 6(1), 15 (2020).
-
Oliver, J. C. et al. Candida spp. and phagocytosis: Multiple evasion mechanisms. Antonie Van Leeuwenhoek 112(10), 1409–1423 (2019).
-
Hassan, Y., Chew, S.Y. and Than, L.T.L. Candida glabrata: Adopting pathogenicity and resistant mechanisms for survival (2021).
-
Kumar, S. et al. Overview on the infections related to rare Candida species. Pathogens 11(9), 963 (2022).
-
Kim, J. & Sudbery, P. Candida albicans, a major human fungal pathogen. J. Microbiol. 49, 171–177 (2011).
-
Hassan, Y., Chew, S. Y. & Than, L. T. L. Candida glabrata: Pathogenicity and resistance mechanisms for adaptation and survival. J. Fungi 7(8), 667 (2021).
-
Lim, C.S.-Y. et al. Candida and invasive candidiasis: Back to basics. Eur. J. Clin. Microbiol. Infect. Dis. 31, 21–31 (2012).
-
Duggan, S. & Usher, J. Candida glabrata: A powerhouse of resistance. PLoS Pathog. 19(10), e1011651 (2023).
-
Dos Santos, M. M. & Ishida, K. We need to talk about Candida tropicalis: Virulence factors and survival mechanisms. Med. Mycol. 61(8), myad075 (2023).
-
Treviño-Rangel, R. D. J. et al. Phenotypical characterization and molecular identification of clinical isolates of Candida tropicalis. Rev. Iberoamericana Micol. 35(1), 17–21 (2018).
-
Keighley, C. et al. Candida tropicalis—A systematic review to inform the World Health Organization of a fungal priority pathogens list. Med. Mycol. 62(6), myae040 (2024).
-
Jha, R. & Mayanovic, R. A. A review of the preparation, characterization, and applications of chitosan nanoparticles in nanomedicine. Nanomaterials 13(8), 1302 (2023).
-
Haider, A. et al. Advances in chitosan-based drug delivery systems: A comprehensive review for therapeutic applications. Eur. Polym. J. 210, 112983 (2024).
-
Goyal, S. et al. Chitosan based nanocarriers as a promising tool in treatment and management of inflammatory diseases. Carbohydr. Polym. Technol. Appl. 7, 100442 (2024).
-
Huang, T. et al. Using inorganic nanoparticles to fight fungal infections in the antimicrobial resistant era. Acta Biomater. 158, 56–79 (2023).
-
Zhang, M. et al. Antifungal activity of ribavirin used alone or in combination with fluconazole against Candida albicans is mediated by reduced virulence. Int. J. Antimicrob. Agents 55(1), 105804 (2020).
-
Afeltra, J. & Verweij, P. Antifungal activity of nonantifungal drugs. Eur. J. Clin. Microbiol. Infect. Dis. 22, 397–407 (2003).
-
Benchallal, F. et al. Thymus vulgaris essential oil enclosed in β-cyclodextrin as drug delivery system: A promising alternative for antibiotic usage in combating Salmonella. Proc. Indian Natl. Sci. Acad. 90, 1–14 (2024).
-
Whaley, S. G. et al. Relative contribution of the ABC transporters Cdr1, Pdh1, and Snq2 to azole resistance in Candida glabrata. Antimicrob. Agents Chemother. https://doi.org/10.1128/aac.01070-18 (2018).
-
Torelli, R. et al. The ATP-binding cassette transporter–encoding gene CgSNQ2 is contributing to the CgPDR1-dependent azole resistance of Candida glabrata. Mol. Microbiol. 68(1), 186–201 (2008).
-
Vu, B. G. & Moye-Rowley, W. S. Azole-resistant alleles of ERG11 in Candida glabrata trigger activation of the Pdr1 and Upc2A transcription factors. Antimicrob. Agents Chemother. 66(3), e02098-e2121 (2022).
-
Nakayama, H. et al. In vitro and in vivo effects of 14α-demethylase (ERG11) depletion in Candida glabrata. Antimicrob. Agents Chemother. 45(11), 3037–3045 (2001).
-
El-Kholy, M. A. et al. Analysis of CDR1 and MDR1 gene expression and ERG11 substitutions in clinical Candida tropicalis isolates from Alexandria, Egypt. Braz. J. Microbiol. 54(4), 2609–2615 (2023).
-
Jin, L. et al. MDR1 overexpression combined with ERG11 mutations induce high-level fluconazole resistance in Candida tropicalis clinical isolates. BMC Infect. Dis. 18, 1–6 (2018).
-
Pandey, N. et al. Overexpression of efflux pump transporter genes and mutations in ERG11 pave the way to fluconazole resistance in Candida tropicalis: A study from a North India region. J. Glob. Antimicrob. Resist. 22, 374–378 (2020).
-
Shree Abirami, N. S., Pillai, D. S. & Shanmugam, R. The antifungal activity of chitosan nanoparticle-incorporated probiotics against oral candidiasis. Cureus 16(9), e70093 (2024).
-
Muangsawat, S. et al. Effects of efflux pumps on antifungal activity of chitosan against Candida albicans. J. Oral Microbiol. 16(1), 2357976 (2024).
-
Kowalczyk, A. et al. Thymol and thyme essential oil—new insights into selected therapeutic applications. Molecules 25(18), 4125 (2020).
-
Kowalczyk, A. Essential oils against Candida auris—A promising approach for antifungal activity. Antibiotics 13(6), 568 (2024).
-
Touati, A. et al. Essential oils for biofilm control: Mechanisms, synergies, and translational challenges in the era of antimicrobial resistance. Antibiotics 14(5), 503 (2025).
-
Qi, X. et al. Mechanisms of antifungal and mycotoxin inhibitory properties of Thymus vulgaris L. essential oil and their major chemical constituents in emulsion-based delivery system. Ind. Crops Products 197, 116575 (2023).
-
Yousfi, H. et al. Repurposing of ribavirin as an adjunct therapy against invasive Candida strains in an in vitro study. Antimicrob. Agents Chemother. https://doi.org/10.1128/aac.00263-19 (2019).
-
Vitali, A. et al. Antifungal carvacrol loaded chitosan nanoparticles. Antibiotics 11(1), 11 (2021).
-
Clinical and Laboratory Standards Institute Wayne. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts (Clinical and Laboratory Standards Institute Wayne, 2008).
-
Calvo, P. et al. Novel hydrophilic chitosan-polyethylene oxide nanoparticles as protein carriers. J. Appl. Polym. Sci. 63(1), 125–132 (1997).
-
Sanguinetti, M. et al. Mechanisms of azole resistance in clinical isolates of Candida glabrata collected during a hospital survey of antifungal resistance. Antimicrob. Agents Chemother. 49(2), 668–679 (2005).
-
Dudakova, A. et al. Molecular tools for the detection and deduction of azole antifungal drug resistance phenotypes in Aspergillus species. Clin. Microbiol. Rev. 30(4), 1065–1091 (2017).
-
Sharifzadeh, A. et al. Synergistic anticandidal activity of menthol in combination with itraconazole and nystatin against clinical Candida glabrata and Candida krusei isolates. Microb. Pathog. 107, 390–396 (2017).
-
Rezai, M. et al. Successful treatment with caspofungin of candiduria in a child with Wilms tumor; review of literature. J. Mycol. Med. 27(2), 261–265 (2017).
-
Yang, Z. et al. Fluconazole inhibits cellular ergosterol synthesis to confer synergism with berberine against yeast cells. J. Glob. Antimicrob. Resist. 13, 125–130 (2018).
-
Choudhury, S. Routine susceptibility testing for Candida albicans isolated from blood stream infections. Pathology 50(4), 462–464 (2018).
-
Ferrari, M. Cancer nanotechnology: Opportunities and challenges. Nat. Rev. Cancer 5(3), 161–171 (2005).
-
Nair, R. S. et al. An evaluation of curcumin-encapsulated chitosan nanoparticles for transdermal delivery. AAPS PharmSciTech 20(2), 69 (2019).
-
Nallamuthu, I., Devi, A. & Khanum, F. Chlorogenic acid loaded chitosan nanoparticles with sustained release property, retained antioxidant activity and enhanced bioavailability. Asian J. Pharm. Sci. 10(3), 203–211 (2015).
-
Fan, W. et al. Formation mechanism of monodisperse, low molecular weight chitosan nanoparticles by ionic gelation technique. Colloids Surf. B 90, 21–27 (2012).
-
Al-Nemrawi, N., Alsharif, S. & Dave, R. Preparation of chitosan-TPP nanoparticles: The influence of chitosan polymeric properties and formulation variables. Int. J. Appl. Pharm. 10(5), 60–65 (2018).
-
Zhao, K. et al. The mucoadhesive nanoparticle-based delivery system in the development of mucosal vaccines. Int. J. Nanomed. 17, 4579–4598 (2022).
-
Guidance, D. Considering Whether an FDA-Regulated Product Involves the Application of Nanotechnology (FDA, 2011).
-
Sabourian, P. et al. Effect of physico-chemical properties of nanoparticles on their intracellular uptake. Int. J. Mol. Sci. 21(21), 8019 (2020).
-
Morris, G. A. et al. The effect of prolonged storage at different temperatures on the particle size distribution of tripolyphosphate (TPP)–chitosan nanoparticles. Carbohyd. Polym. 84(4), 1430–1434 (2011).
-
Logigan, C.-L. et al. Poly (ethylene glycol) methyl ether acrylate-grafted chitosan-based micro-and nanoparticles as a drug delivery system for antibiotics. Polymers 16(1), 144 (2024).
-
Gerami, S. E. et al. Preparation of pH-sensitive chitosan/polyvinylpyrrolidone/α-Fe2O3 nanocomposite for drug delivery application: Emphasis on ameliorating restrictions. Int. J. Biol. Macromol. 173, 409–420 (2021).
-
Verma, S. et al. Shape dependent therapeutic potential of nanoparticulate system: Advance approach for drug delivery. Curr. Pharm. Des. 30(33), 2606–2618 (2024).
-
Fabozzi, A. et al. Design of non-spherical poly (lactic-co-glycolic acid) nanoparticles functionalized with hyaluronic acid for active tumor targeting. J. Drug Deliv. Sci. Technol. 87, 104836 (2023).
-
Wu, J. et al. Preparation and biological activity studies of resveratrol loaded ionically cross-linked chitosan-TPP nanoparticles. Carbohyd. Polym. 175, 170–177 (2017).
-
Knaul, J. Z., Hudson, S. M. & Creber, K. A. Improved mechanical properties of chitosan fibers. J. Appl. Polym. Sci. 72(13), 1721–1732 (1999).
-
Ozbek, O., Ulgen, K. O. & IleriErcan, N. The toxicity of polystyrene-based nanoparticles in Saccharomyces cerevisiae is associated with nanoparticle charge and uptake mechanism. Chem. Res. Toxicol. 34(4), 1055–1068 (2021).
-
Barbalinardo, M. et al. Surface charge overrides protein corona formation in determining the cytotoxicity, cellular uptake, and biodistribution of silver nanoparticles. ACS Appl. Bio Mater. 8(6), 5032–5043 (2025).
-
Iacob, A. T. et al. Recent biomedical approaches for chitosan based materials as drug delivery nanocarriers. Pharmaceutics 13(4), 587 (2021).
-
Lino, P. R. et al. Systematic modification and evaluation of enzyme-loaded chitosan nanoparticles. Int. J. Mol. Sci. 22(15), 7987 (2021).
-
Aljaafari, M. N. et al. An overview of the potential therapeutic applications of essential oils. Molecules 26(3), 628 (2021).
-
Monnery, B. D. et al. Cytotoxicity of polycations: Relationship of molecular weight and the hydrolytic theory of the mechanism of toxicity. Int. J. Pharm. 521(1–2), 249–258 (2017).
-
Behl, G. et al. Gallic acid loaded disulfide cross-linked biocompatible polymeric nanogels as controlled release system: Synthesis, characterization, and antioxidant activity. J. Biomater. Sci. Polym. Ed. 24(7), 865–881 (2013).
-
Manimekalai, P., Dhanalakshmi, R. & Manavalan, R. Preparation and characterization of ceftriaxone sodium encapsulated chitosan nanoparticles. Int. J. Appl. Pharm. 9(6), 10 (2017).
-
Banerjee, D., Burkard, L. & Panepinto, J. C. Inhibition of nucleotide biosynthesis potentiates the antifungal activity of amphotericin B. PLoS ONE 9(1), e87246 (2014).
-
Tsai, Y.-C. et al. Natural polysaccharide-based nanoparticles enhance intracellular delivery and cytotoxicity of Antrodia camphorata in breast cancer cells. Int. J. Mol. Sci. 26(17), 8420 (2025).
-
Sharifzadeh, A., Shokri, H. & Katiraee, F. Anti-adherence and anti-fungal abilities of thymol and carvacrol against Candida species isolated from patients with oral candidiasis in comparison with fluconazole and voriconazole. Jundishapur J. Nat. Pharm. Prod. 16(1), e65005 (2021).
-
Stojanović-Radić, Z. et al. Antifungal activity of Thymus vulgaris L. essential oil against Candida species causing otomycosis: Anti-virulence activity, synergistic potential, and cell wall effect. Chem. Biodivers. https://doi.org/10.1002/cbdv.202501145 (2025).
-
Ahmad, A., Van Vuuren, S. & Viljoen, A. Unravelling the complex antimicrobial interactions of essential oils—the case of Thymus vulgaris (Thyme). Molecules 19(3), 2896–2910 (2014).
-
Pedroso, M. B. et al. Antifungal activity and mechanism of action of monoterpenes against Botrytis cinerea. Ciência e Agrotecnol. 48, e018823 (2024).
-
Ahmad, A. et al. Fungicidal activity of thymol and carvacrol by disrupting ergosterol biosynthesis and membrane integrity against Candida. Eur. J. Clin. Microbiol. Infect. Dis. 30, 41–50 (2011).
-
Shariati, A. et al. Natural compounds: A hopeful promise as an antibiofilm agent against Candida species. Front. Pharmacol. 13, 917787 (2022).
-
Khan, M. S. A., Ahmad, I. & Cameotra, S. S. Carum copticum and Thymus vulgaris oils inhibit virulence in Trichophyton rubrum and Aspergillus spp. Braz. J. Microbiol. 45, 523–531 (2014).
-
Locke, J. B. et al. Characterization of in vitro resistance development to the novel echinocandin CD101 in Candida species. Antimicrob. Agents Chemother. 60(10), 6100–6107 (2016).
-
Routh, M. M., Chauhan, N. M. & Karuppayil, S. M. Cancer drugs inhibit morphogenesis in the human fungal pathogen, Candida albicans. Braz. J. Microbiol. 44, 855–859 (2013).
-
Nair, A. et al. Nanoparticles—Attractive carriers of antimicrobial essential oils. Antibiotics 11(1), 108 (2022).
-
Gurunathan, S., Lee, A. R. & Kim, J. H. Antifungal effect of nanoparticles against COVID-19 linked black fungus: A perspective on biomedical applications. Int. J. Mol. Sci. 23(20), 12526 (2022).
-
Hetta, H. F. et al. Nanotechnology-based strategies to combat multidrug-resistant Candida auris infections. Pathogens 12(8), 1033 (2023).
-
Bhattacharya, S., Sobel, J. D. & White, T. C. A combination fluorescence assay demonstrates increased efflux pump activity as a resistance mechanism in azole-resistant vaginal Candida albicans isolates. Antimicrob. Agents Chemother. 60(10), 5858–5866 (2016).
-
Hans, S., Fatima, Z. & Hameed, S. Insights into the modulatory effect of magnesium on efflux mechanisms of Candida albicans reveal inhibition of ATP binding cassette multidrug transporters and dysfunctional mitochondria. Biometals 34(2), 329–339 (2021).
-
Gil, F. et al. Extracts from Argentinian native plants reverse fluconazole resistance in Candida species by inhibiting the efflux transporters Mdr1 and Cdr1. BMC Complement. Med. Ther. 22(1), 264 (2022).
-
Mota, K. S. D. L. et al. Antifungal activity of Thymus vulgaris L. essential oil and its constituent phytochemicals against Rhizopus oryzae: Interaction with ergosterol. Molecules 17(12), 14418–14433 (2012).
-
Moazeni, M. et al. In vitro antifungal activity of Thymus vulgaris essential oil nanoemulsion. J. Herbal Med. 28, 100452 (2021).
Funding
This study was extracted from Shima Aghaei’s thesis and financially supported by the Research Council of Tarbiat Modares University, Tehran, Iran, in 2025.
Ethics declarations
Ethics approval
This study is experimental research approved as a thesis with an ethical code number of “IR.MODARES.REC.1401.226” at Tarbiat Modares University, Tehran, Iran.
Statement
To ensure the highest quality of writing, this article used Wordtune software to refine the language, improve clarity, enhance readability, and perform spelling and grammar checks. Finally, the entire article was reviewed and approved by the authors.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Aghaei, S., Yadegari, M.H., Rajabibazl, M. et al. Chitosan nanoparticles loaded with ribavirin, carboplatin, and thyme oil downregulate SNQ2, ERG11, CDR1, and MDR1 in Candida strains. Sci Rep 15, 42633 (2025). https://doi.org/10.1038/s41598-025-26841-0
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-26841-0