- Research
- Open access
- Published:
Biotechnology for Biofuels and Bioproducts volume 18, Article number: 94 (2025) Cite this article
Abstract
Background
Oleaginous microorganisms are promising lipid producers that accumulate an abundance of lipids from different carbon sources. However, the cost of the carbon source in the culture medium is a significant component of the total substrate cost. In this study, lignocellulose from corncob hydrolysate (CBH) was used instead of glucose as a low-cost medium for Schizochytrium fermentation.
Results
Eicosapentaenoic acid (EPA) content was 7.31%, after 110 h of fermentation, when the total sugar concentration of CBH was 80 g/L, which was greater than that of pure glucose medium. Replacing 40% of freshwater with fermentation wastewater (FW) resulted in biomass, lipid titer, and EPA titer of 42.16 g/L, 23.05 g/L, and 1.72 g/L, respectively. Compared with the initial CBH medium, the lipid and EPA titers in the 7.5-L bioreactor employing the FW recycling strategy using CBH as a carbon source increased by 12.10% and 9.26%, respectively.
Conclusions
Corncob hydrolysate can be used as a potential low-cost and effective carbon source for EPA production by Schizochytrium sp. The recycling of FW provides a reference for reducing freshwater consumption and environmental pollution and realizing green and economic recycling fermentation.
Introduction
Eicosapentaenoic acid (EPA) is a polyunsaturated fatty acid (PUFA) required for proper bodily function. It cannot be produced by the human body itself and must be obtained from exogenous foods. It inhibits platelet aggregation, prevents thrombosis, lowers blood fat, and prevents Alzheimer’s disease [1, 2]. In addition, it significantly inhibits inflammation and certain cancers [3, 4]. EPA was largely produced by marine fisheries for a long time [5]. However, it is unable to meet the growing market demand due to over-exploitation of marine resources, marine pollution, and other reasons. Numerous studies have demonstrated that marine oleaginous microorganisms can serve as a substitute source for the sustainable PUFA synthesis [6]. Schizochytrium sp., a lipid-producing marine microbe, has been widely exploited in scientific studies and industrial PUFA production [especially docosahexaenoic acid (DHA)] owing to its fast growth rate, simple cultivation, and high PUFA content [7]. However, the cost of fermentation substrates largely determines the economy of the microbial production of EPA, seriously hindering its industrial application. The cost of the carbon source in the medium accounts for approximately 80% of the substrate cost and 60% of the total fermentation cost [8]. Thus, inexpensive carbon sources must replace glucose in Schizochytrium sp. fermentation to reduce the cost of lipid production.
Lignocellulosic biomass, such as wood, grass, agricultural and forest residues, is the most abundant renewable resource on Earth [9]. Lignocellulose is composed primarily of cellulose, hemicellulose, and lignin. After pretreatment, these components can be transformed into sugars, indicating the significant potential of lignocellulose as a feedstock for biofuels production via microbial culture. In addition to improving fermentation economy, lignocellulose can aid in the growth of a circular economy that is environmentally friendly. Currently, only a few studies exist on the production of lipids by Schizochytrium sp. using lignocellulose as a carbon source. Olsen et al. [10] produced DHA by fermenting Aurantiochytrium limacinum SR21 with the hydrolysate of Norway spruce as a carbon source; the lipid content reached 44.00% (w/w), of which 1/3 was DHA. A mutant strain of Aurantiochytrium sp. with strong tolerance to the lignocellulose hydrolysate was obtained through multiple rounds of domestication when the sugarcane hydrolysate was used as a carbon source; the lipid titer and DHA titer reached 11.84 g/L and 3.15 g/L, respectively [11]. Wang et al. [12] fermented Schizochytrium sp. HX-308 in corn stover hydrolysate, and the lipid titer reached 21.20 g/L, which was 58.60% lower than that of the pure glucose group. Therefore, it is necessary to develop a method for efficient lipid production by fermenting Schizochytrium sp. using lignocellulose feedstock as the carbon source.
Biofermentation is a promising technology widely used in the food, medical, and biofuel industries. Nevertheless, fermentation generates an abundance of fermentation wastewater (FW) and requires large volumes of water resources. Fermentation wastewater usually contains high concentrations of organic pollutants, which are difficult and costly to treat, as well as cause environmental pollution when deposited without control [13].
Reports have suggested that the production of 1000 tons of microalgal lipids by fermentation will require 15,000 tons of freshwater and produce 13,000 tons of wastewater [7]. Wastewater resource exploitation has been the subject of numerous studies recently. Wastewater recycling technologies have been developed to produce several valuable compounds that reduce water consumption and fermentation costs [14,15,16]. Mineral elements and organic biomolecules (peptides, amino acids, nucleotides, and polysaccharides) are present in FW of Schizochytrium [7]. Song et al. [17] produced DHA and arachidonic acid (ARA) by the cross-recovery of Aurantiochytrium and Mortierella alpina FW. The titer of DHA and ARA was almost the same as that of the control group, reaching 30.40 g/L and 5.13 g/L, respectively. Fermentation reduced the water consumption by 50%. Wastewater recycling provides a new method for microbial fermentation to reduce costs and environmental pollution.
A Schizochytrium sp. ALE-D50, which has excellent EPA production characteristics, was screened using an adaptive evolution strategy of high temperature and oxygen stress [18]. The biomass, lipid, and EPA titer reached 35.33 g/L, 21.47 g/L, and 1.54 g/L, respectively, which had excellent production performance.
The objective of this study was to devise an environmentally friendly fermentation approach for recycling resources and enhancing the economic viability of the industrial production of EPA-rich lipids. First, the composition and total sugars content of the corncob hydrolysate (CBH) were analyzed, and the impact of typical degradation products [furfural, 5-hydroxymethylfurfural (5-HMF), and acetic acid] on the growth of Schizochytrium and lipid production was evaluated. The effects of CBH and glucose media with varying concentrations of total sugars on the growth, lipid accumulation, and fatty acid composition of Schizochytrium were compared. Subsequently, FW was used to replace freshwater to explore its effect on EPA production by Schizochytrium. Finally, the process was scaled up in a 7.5 L bioreactor using a fed-batch fermentation strategy. This study is highly significant in reducing the cost of raw substrate materials for fermenting Schizochytrium, as well as in minimizing water resource consumption and environmental pollution caused by wastewater discharge.
Materials and methods
Microorganism and culture media
The Schizochytrium variant ALE-D50 preserved in the laboratory was used for the biosynthesis of EPA. The strain was preserved in 20% (v/v) glycerol at − 80 ℃. The seed and fermentation media were identical to those used in our previous study [19]: seed medium, 30 g/L glucose, 10 g/L peptone, 5 g/L yeast extract, 0.05 g/L vitamin B1, 0.05 g/L vitamin B6, 0.0005 g/L vitamin B12, 15 g/L sea salt. Fermentation medium: 100 g/L glucose, 5.6 g/L peptone, 20 g/L C5H8NO4Na-H2O, 2.5 g/L KH2PO4, 7.2 g/L MgSO4, 12.8 g/L Na2SO4, 0.4 g/L CaCl2, 0.1 g/L vitamin B1, 0.1 g/L vitamin B6, 0.001 g/L vitamin B12, and 17.5 g/L sea salt.
For the CBH medium, the glucose in the fermentation medium was replaced with CBH, and the other components were the same as those in the fermentation medium.
Culture conditions
A seed culture was prepared by transferring 1 mL of the strain stored in a glycerol tube to a 250-mL shake flask containing 50 mL of seed medium and incubating at 28 ℃ and 200 rpm for 48 h. The seed cultures after three generations (10%, v/v) were inoculated into the fermentation medium and incubated at 28 ℃ and 200 rpm.
Fed-batch fermentation was conducted in a 7.5-L bioreactor with an initial working volume of 3 L using a pulse-feeding strategy. After three generations of seed cultures, the seed culture solution (10%, v/v) was inoculated into the fermentation medium. During fermentation, the samples were collected every 12 h to determine the sugar concentration, biomass, lipid titer, EPA titer, and other parameters. Moreover, when the sugar substrate concentration was below 20 g/L, 400 g/L of sterile glucose or CBH (equivalent to 400 g/L glucose) was added to the fermentation broth to maintain the sugar substrate at approximately 40 g/L. Fermentation conditions were 300 rpm, 2 vvm, and 28 ℃. Next, 2 mol/L HCl or 2 mol/L NaOH was added to the fermentation broth to adjust and stabilize the pH of the fermentation broth.
Corncobs hydrolysate and fermentation wastewater preparation
Air-dried corncobs were pulverized using a pulverizer and sieved to collect 40–80 mesh particles. Next, corncob powder was added into a 5-L three-necked flask containing 1% (w/w) sulfuric acid at a solid-to-liquid ratio of 1:10 and pretreated at 150 ℃ for 30 min. After pretreatment, the liquid and solid fractions were separated via vacuum filtration. The solid portion was washed with tap water, filtered, and air-dried to approximately 50% moisture content for enzymatic hydrolysis. Cellulose, hemicellulose, and lignin contents in corncobs were determined by National Renewable Energy Laboratory (NREL) method [20]. The corncob feedstock contained 46.14% (w/w) cellulose, 27.62% (w/w) hemicellulose, and 15.39% (w/w) lignin.
The enzymatic hydrolysis followed the method described by Mukasekuru et al. and was modified accordingly [21]. The air-dried solids (50% moisture, w/w) were mixed with citrate buffer (50 mM) to achieve a solid content of 5% wt. Enzymatic hydrolysis was carried out using a commercially available cellulase preparation, and the diluted enzyme solution was added at an enzyme concentration of 2 FPU/g dry substrate. Enzymatic hydrolysis was conducted at 50 ℃ and 180 rpm for 72 h. After 72 h of enzymatic hydrolysis, the enzymatically digested slurry was centrifuged (8000g, 10 min) and concentrated to titer the CBH with a glucose concentration of approximately 400 g/L.
Schizochytrium sp. was shake flask cultured for 110 h or fed-batch cultured for 120 h, with CBH as the carbon source, to obtain FW. After fermentation, the fermentation broth was centrifuged at 8000g for 15 min, the cells and supernatant were collected, and the supernatant was used as FW.
Analytical methods
The content of glucose and xylose in corncob hydrolysates was determined by high-performance liquid chromatography (HPLC) with a refractive index detector (RID-6A) and Sugar-Pak I (6.5 mm × 300 mm, 10 μm). The mobile phase consisted of ultrapure water with a flow rate of 0.6 mL/min. Furfural, 5-HMF, and acetic acid were analyzed by HPLC (Agilent Technologies, USA) using a UV/Vis (SPD-20A) detector and Aminex HPX-87H column (300 mm × 7.8 mm). The mobile phase comprised 5 mmol/L H2SO4 at a constant flow rate of 0.5 mL/min at 30℃. The detection wavelengths were 250 nm and 280 nm, respectively.
Next, the components of FW were determined. The total nitrogen in the wastewater was determined using the Kjeldahl method. The concentrations of the mineral ions (Na+, K+, Mg2+, and Ca2+) were determined using inductively coupled plasma spectrometry (ICP-OES Optima 8000; Perkin Elmer, USA). Radio frequency power was 1150 W, auxiliary gas flow rate was 0.5 L/min, pump speed was 45 rpm, cooler flow rate was 12 L/min, and the sampling depth was 5 mm.
The FW was diluted with double-distilled water, followed by centrifugation at 8000g for 10 min. The supernatant was collected and filtered using a 0.22-μm microporous membrane. Anions such as Cl−, SO42−, PO43− were measured by ion chromatography (CIC-D150; Shine, China) equipped with an AS-LH-AC-3 column (250 mm × 4.6 mm).
Next, FW was diluted with 10% trichloroacetic acid (TCA) in equal volumes. TCA can denature and precipitate proteins in FW, thus effectively removing protein interference. Subsequently, the mixture was filtered through a double layer of filter paper and centrifuged at 10,000g for 30 min. The amino acid content in wastewater was determined by HPLC equipped with a C18 column (4.6 × 250 mm, 5 μm).
The biomass was determined using the freeze-drying weighing method. Fermentation broth (30 mL) was centrifuged at 8000g for 10 min, and the bacterial precipitate was collected, freeze-dried in vacuum for 60 h, and weighed. Afterward, 3.20 mL of 6 mol/L HCl solution was added to 0.40g of the freeze-dried bacterial powder and incubated in a 75 °C water bath for 30 min to rupture the cells. The extraction was repeated for three to four times with 8 mL of n-hexane as the extraction solvent, dried in a rotary vacuum, and the lipid titer was quantitatively determined. The fatty acid composition was determined using a gas chromatograph according to the method described by Zhao et al. [22].
Statistical analysis
All experimental data are presented as mean ± standard deviation from three biologically independent replicates. Student’s t-test and one-way analysis of variance were performed using the SPSS software (version 20.0). The difference between the groups was expressed by English letters, and the difference between different letters was significant (p < 0.05). The significance levels are expressed as follows: *p < 0.05, **p < 0.01, and ***p < 0.001.
Results and discussion
Sensitivity of Schizochytrium sp. ALE-D50 to different degradation products of lignocellulose
Degradation products that strongly inhibit microorganisms are produced during the pretreatment of lignocellulosic biomass [23]. These inhibitors are primarily furfural derivatives and organic acids. The main components in the concentrated CBH were 414.57 g/L glucose, 256.20 g/L xylose, 1.75 g/L acetic acid, 0.76 g/L furfural, and 0.83 g/L 5-HMF (Table 1). The sensitivity of Schizochytrium sp. was evaluated by measuring the inhibitory substances produced by the lignocellulosic biomass and treatment.
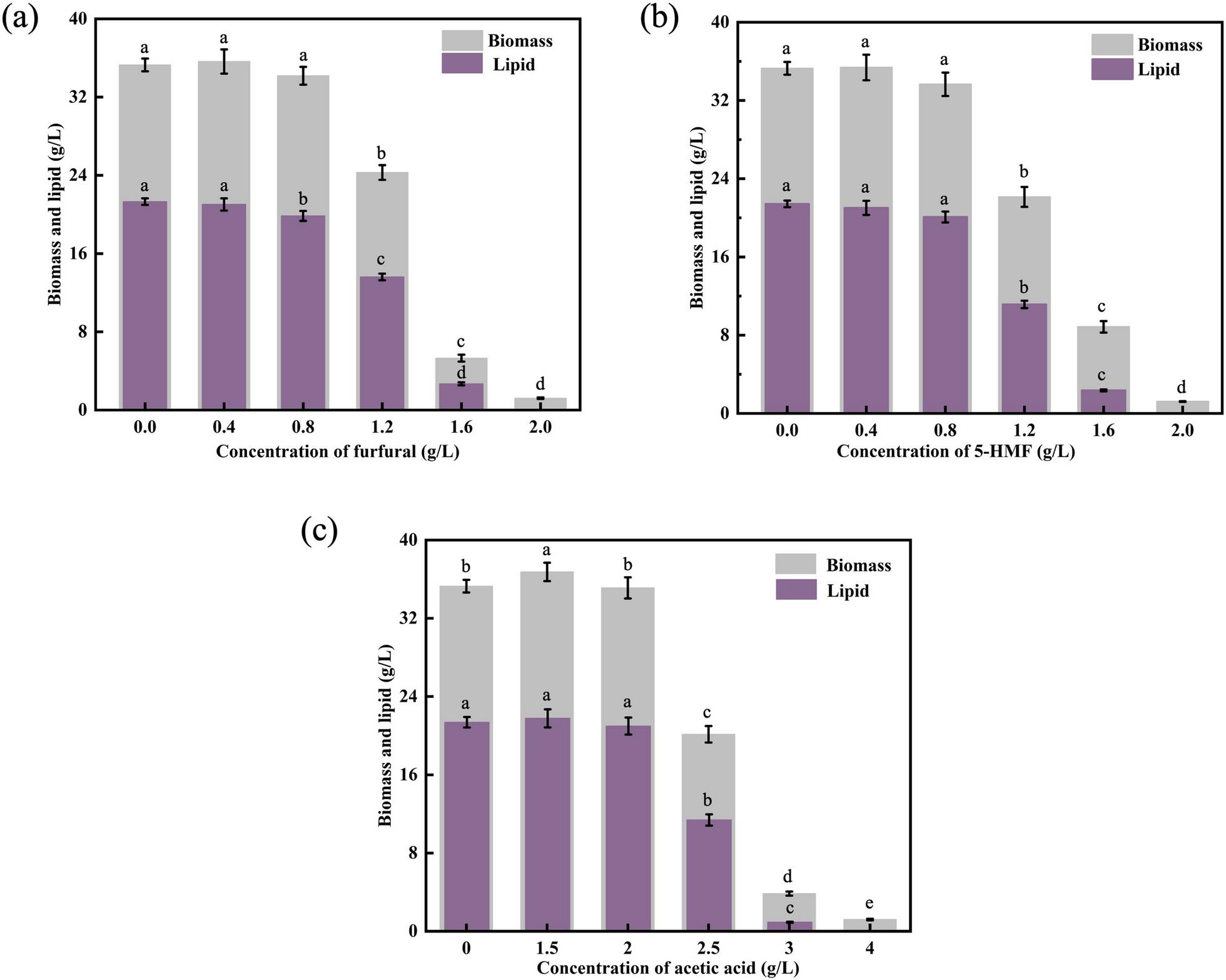
The synthesis of biomass and lipid synthesis progressively decreased with an increase in furfural concentration (Fig. 1a). Cell growth and lipid accumulation were not significantly affected by furfural concentrations of up to 0.8 g/L. Furfural clearly reduced biomass growth and lipid accumulation when the concentration reached 1.2, 1.6, or 2.0 g/L. The presence of 2 g/L furfural almost completely inhibited growth and lipid production. Similar results were obtained with 5-HMF (Fig. 1b). The production of lipids and biomass gradually declined as the concentration of 5-HMF increased. At a 5-HMF concentration of 0.8 g/L, no significant inhibitory effects were observed on the growth or lipid accumulation of Schizochytrium. However, when the concentration increased to 1.6 g/L, biomass production declined sharply by 69.03%, accompanied by an 89.32% reduction in lipid yield. Notably, at 2 g/L 5-HMF, cellular lipid accumulation was completely suppressed. Furfural derivatives are the main microbial inhibitors in lignocellulose hydrolysate after acidic hydrolysis, which can inhibit the glycolysis pathway, destroy redox balance, and induce oxidative stress to produce excessive reactive oxygen species [24,25,26]. The findings demonstrated that the essential enzymes for fatty acid production in Trichosporon fermentans were citrate lyase and malic enzymes, and furfural hindered both of their activities [27]. It is suggested that furfural inhibits fatty acid synthesis in Schizochytrium via a similar mechanism. At 0.5 g/L furfural, Rhodosporidium toruloides and Lipomyces starkeyi demonstrated total inhibition of growth, whereas Trichosporon cutaneum displayed considerable inhibition of growth [28]. In contrast, Schizochytrium sp. ALE-D50 demonstrated a better tolerance to furfural and 5-HMF.

Effects of different concentrations of inhibitors on biomass and lipid titer of Schizochytrium sp. a Furfural; b 5-HMF; and c acetic acid. Data represent the mean ± standard deviation (n = 3). 5-HMF: 5-hydroxymethyl furfural

Fermentation characteristics of Schizochytrium sp. under different concentrations of glucose and CBH medium. a Biomass and total sugar residue; b lipid; c EPA content; and d EPA titer. TFAs: total fatty acids. Data represent the mean ± standard deviation (n = 3)
Acetic acid is the most prevalent organic acid in lignocellulose hydrolysate. Acetic acid inhibits the development of yeast cells by increasing the levels of reactive oxygen species or decreasing intracellular pH [29]. However, when the concentration of acetic acid in the medium was 1.5 g/L, the biomass and lipid titer of Schizochytrium sp. ALE-D50 increased to 36.74 g/L and 21.76 g/L, respectively (Fig. 1c). Oleaginous microorganisms use acetic acid as the carbon source to produce lipids [30]. Acetic acid can be converted to acetyl-CoA by acetyl-CoA synthase, an important precursor of lipid biosynthesis. The lipid content of oleaginous yeasts Apiotrichum brassicae and Pichia kudriavzevii reached 68% when acetic acid was employed as the carbon source [31]. In addition, Schizochytrium produced DHA via fermentation using acetic acid as the sole carbon source. This resulted in biomass and DHA titers considerably higher than those of glucose, at 146.00 g/L and 23.00 g/L, respectively [32]. The lipid production of strain Schizochytrium sp. HX-308 increased by 17.6% in the presence of 1.5 g/L acetic acid [12]. Nevertheless, the growth of Schizochytrium was greatly delayed, and lipid synthesis was stopped when the acetic acid content of the solution exceeded 2.5 g/L.
Optimization of CBH concentration of Schizochytrium sp. ALE-D50
Because the concentrated CBH contained approximately 400 g/L of glucose, Schizochytrium sp. could be cultured using it as a carbon source. The effects of CBH and pure glucose on the fermentation of Schizochytrium sp. ALE-D50 were compared using a glucose medium with the same total sugars content. The feasibility of using CBH as a carbon source by Schizochytrium sp. ALE-D50 fermentation was explored. As sugar concentration increased, the biomass and lipid production of ALE-D50 increased gradually (Fig. 2). The biomass and lipid titers in the CBH medium with 80 g/L total sugar concentration were achieved at 36.89 g/L and 22.01 g/L, respectively. These values were nearly identical to those in 80 g/L glucose medium (37.16 g/L) and lipid titer (22.85 g/L) (p > 0.05). When the sugar concentration in the CBH medium exceeded 80 g/L, a downward trend in biomass and lipid production was observed, although this change was not statistically significant (p > 0.05). This phenomenon may indicate a potential inhibitory effect when the sugar concentration in the medium is too high. An increase in sugar concentration alters the C:N ratio in the medium, thereby significantly impacting the growth and lipid accumulation of Schizochytrium [6].
Interestingly, CBH shows a potential comparable to glucose as a carbon source for EPA biosynthesis in Schizochytrium. As shown in Fig. 2c, at 80 g/L total sugar concentration, EPA content in CBH medium reached 7.31%, an increase of 7.02% compared with glucose (p < 0.05). Numerous toxic byproducts, including furfural and acetic acid, are generated, causing oxidative stress in cells during lignocellulose pretreatment. PUFAs contain unsaturated double bonds and are excellent antioxidants. Therefore, Schizochytrium produces more PUFAs to protect itself from oxidative damage under oxidative stress conditions. As illustrated in Fig. 2d, at a sugar concentration of 80 g/L, the EPA titer in the CBH medium (1.61 g/L) was slightly higher than that in glucose medium (1.56 g/L) (p > 0.05). These findings suggest that CBH may be a potential alternative carbon source for Schizochytrium sp. ALE-D50. Thus, a CBH medium with a total sugar content of 80 g/L was selected for subsequent experiments.
Components of fermentation wastewater
The use of CBH as a carbon source can reduce the economic cost of the EPA titer by fermenting Schizochytrium. However, a huge volume of FW is discharged during fermentation. Wastewater is enriched with vital mineral elements necessary for the cellular composition and is involved in the cellular metabolism of Schizochytrium. Furthermore, it contains elevated levels of organic biomolecules, such as peptides, amino acids, and polysaccharides. The utilization of wastewater resources is expected to diminish wastewater discharge, lower the economic burden of fermentation, and mitigate environmental pollution.
The primary components of FW were first analyzed and compared with those of the CBH fermentation medium to verify the suitability of FW for replacing freshwater in a fermentation medium (Table 2). The difference in pH between CBH and FW media was insignificant. The cations in the FW were Na+ 10.20 g/L, K+ 0.61 g/L, Mg2+ 1.30 g/L, and Ca2+ 0.03 g/L, and the contents of anions were Cl− 7.26 g/L, SO42− 9.91 g/L, and PO43− 0.16 g/L. After fermentation, the concentrations of SO42− and PO43− decreased significantly, and these ions were absorbed to promote cell growth [33], whereas other ions, such as Na+ and K+, maintained the osmotic pressure of the medium [34]. Moreover, the amino acid composition analysis revealed that the concentrations of proline and aspartic acid in the FW were 3.49 times and 0.93 times higher, respectively, than those in the CBH medium (Supplementary Table S1). In contrast, glutamate, arginine, and other amino acids are reduced in FW due to active assimilation by Schizochytrium during fermentation. A comprehensive analysis revealed that the composition of FW indicates the potential practicality of recycling FW. To assess the suitability of FW, various ratios of FW (0%, 20%, 40%, 60%, and 80%) were employed to substitute freshwater in laboratory-scale shake flask fermentation. The effects of replacing freshwater with FW on the fermentation performance of Schizochytrium were evaluated.
Evaluation of fermentation performance of Schizochytrium sp. using fermentation wastewater
The effect of varying levels of FW on the fermentation efficiency of Schizochytrium is illustrated in Fig. 3. When the addition amount of FW is 40%, the introduction of FW increased biomass (Fig. 3a), reaching a maximum value of 42.16 g/L, which was 14.29% higher than the control (p < 0.05). This phenomenon can be attributed to the elevated levels of Na+ and SO42− present in the FW, with sodium sulfate identified as a favorable sodium salt for the productive cultivation of Schizochytrium [35]. In addition, increased osmotic pressure has been recognized as advantageous for cellular growth [36]. Nevertheless, the biomass reduced when the added amount was greater than 40%. At 80% addition, the biomass decreased by 10.76% compared to that in the control group. High salt conditions (high NaCl concentrations) can cause cell death [37], and FW contains high concentrations of Na+ and Cl−. Simultaneously, the increase of FW will lead to the accumulation of furfural, 5-HMF, and acetic acid in the CBH medium, which may potentially inhibit the growth and lipid accumulation of Schizochytrium sp. As shown in Fig. 3a, when the addition amount of FW was increased from 0 to 40%, the lipid titer showed a slight upward trend, in which the lipid titer of the 40% addition group reached 23.05 g/L, which was 4.73% higher than that of the control group (22.01 g/L) (p > 0.05). In contrast, when the amount of FW added was 80%, the lipid titer decreased by 24.94% compared with the control group (p < 0.05). The EPA titer displayed a decreasing trend in the range of 60% to 80% (Fig. 3b) and reached a maximum value of 1.72 g/L with 40% addition. The percentage of EPA in the overall TFAs exhibited a notable decrease when the FW concentration was in the range of 60–80% (Fig. 3c). The proportion of EPA in the TFAs was only 4.72% when FW was 80%, indicating a 54.87% reduction compared to the control group. Meanwhile, with FW added at contents of 60% or higher, PUFAs content in TFAs decreased, whereas saturated fatty acids (SFAs) content increased (Fig. 3d). When FW was added in excess, the salinity (NaCl) increased, which caused the cells to exhibit the characteristics of salt stress, and the fatty acid composition tended toward SFAs. This finding is consistent with that reported by Bagul et al. [38]. Simultaneously, Sun et al. [39] demonstrated that high salt stress was beneficial for Schizochytrium to accumulate lipids rich in SFAs. Furthermore, certain harmful components that impede cell growth and lipid synthesis may accumulate during fermentation, whereas more inhibitory byproducts, such as furfural and acetic acid, accumulate in the FW [38]. Overall, when 40% FW was used, the biomass, lipid production, and EPA production reached 42.16 g/L, 23.05 g/L, and 1.72 g/L, respectively. The results indicated that the FW could partially replace freshwater in the CBH medium (by approximately 40%) and reduce wastewater discharge.

Effects of different fermentation wastewater additions on the fermentation characteristics of Schizochytrium sp. a Biomass and lipid titer; b EPA titer; c fatty acid profile; d SFAs and PUFAs content. Data represent the mean ± standard deviation (n = 3)
The effect of varying the number of cycles on the fermentation characteristics of ALE-D50 was examined to develop a more economically efficient fermentation process. As shown in Fig. 4a, the biomass of the first cycle increased by 14.29% compared with that of the initial fermentation; however, the biomass decreased significantly with an increase in the number of cycles. The biomass of the second and third cycles decreased by 6.93% and 37.54%, respectively, compared to that of the initial fermentation. A similar trend was observed for the lipid titers. This decrease in biomass and lipid titer may be attributed to elevated levels of self-inhibitory biomolecules, including peptides, proteins, and polysaccharides, produced by cell autolysis in the FW from multiple cycles, which might have inhibited cell growth [7]. In addition, the repeated recycling of fermentation wastewater increased the content of furfural, 5-HMF, and acetic acid in the medium. The EPA titer decreased significantly with an increasing number of cycles compared with the initial fermentation (Fig. 4b). No significant difference was observed between an EPA titer of 1.72 g/L in the first cycle and that in the initial fermentation. The EPA titers in the second and third cycles were 42.86% and 68.32% lower, respectively, than those in the initial fermentation. Similarly, the percentage of EPA in TFAs decreased as the number of cycles increased (Fig. 4b). Thus, multiple cycles of FW fermentation exert a significant negative effect on the growth and lipid accumulation of Schizochytrium. Furthermore, the PUFA content demonstrated a significant downward trend with an increase in cycle time, whereas the SFA content exhibited a corresponding increase (Supplementary Fig. S1). Thus, it is feasible to recycle 40% of FW, which can effectively reduce wastewater discharge and production costs; however, the recycling times should be controlled to within two times.

Effects of different recycle numbers on the fermentation performance of Schizochytrium sp. a Biomass and lipid titer; b EPA titer and EPA content. Data represent the mean ± standard deviation (n = 3)
Effects of CBH and fermentation wastewater on growth and EPA production of Schizochytrium sp.
The growth dynamics and lipid synthesis in fed-batch fermentation were determined using a 7.5-L bioreactor to further evaluate the potential of corncob hydrolysates and FW as alternative carbon sources and partial freshwater for EPA production (Fig. 5). Total sugar, biomass, lipid production, and EPA production were measured at 12 h intervals, and when the sugar concentration in the fermenter was below 20 g/L, it was maintained at approximately 40 g/L by supplementing a high concentration of CBH solution (400 g/L). As shown in Fig. 6, a notable increase in biomass and lipid titers in CBH and FW was observed from 0 to 96 h, and biomass and lipid production remained essentially unchanged with the extension of fermentation time (after 96 h). Significant variations in biomass and lipid production were observed between the CBH and recycling fermentation methods. Specifically, at 120 h, the recycling fermentation of the FW achieved maximum biomass and lipid titers of 100.28 and 54.58 g/L, respectively, which were 21.83% and 12.10% higher than the corresponding values of the corncob hydrolysate. The titer of EPA by recycle fermentation reached 4.01 g/L during fermentation, which was 9.26% higher than that of CBH fermentation. The results demonstrated that the accumulation of substances in FW enhanced lipid production and cell growth.

Different fermentation strategies for EPA production from corncob hydrolysate. a conventional fermentation strategy using CBH as the sole carbon source; b cyclic fermentation strategy using CBH as the sole carbon source combined with FW

Comparative analysis of fermentation performance of Schizochytrium sp. in traditional fermentation and cyclic fermentation. a Conventional fermentation strategy; b cyclic fermentation strategy. Data represent the mean ± standard deviation (n = 3)
Researchers have recently investigated lignocellulosic biomasses such as corn stover, bagasse, and spruce as alternative carbon sources for lipid production by Schizochytrium sp. fermentation (Table 3). Compared to other types of lignocellulosic biomass, CBH substantially increased biomass and lipid accumulation in Schizochytrium sp. ALE-D50. Therefore, CBH has essential application prospects as an economical raw material for fatty acid production by Schizochytrium sp. Dioscorea zingiberensis starch saccharification liquid is glucose-rich wastewater produced by saponin extraction. Direct discharge causes environmental pollution and wastes resources. Bao et al. [6] used the starch saccharification liquid of Dioscorea zingiberensis tubers as a substitute carbon source for Schizochytrium sp. and obtained a surprising fermentation effect. The biomass and lipid titer reached 44.85 g/L and 14.58 g/L, respectively, when the sugar concentration was 90 g/L, which not only reduced resource waste, but also reduced lipid production cost. Fermentation wastewater, which is difficult to treat and pollutes the environment, is the major byproduct of lipid fermentation. Therefore, recycling FW has attracted increasing attention. Yin et al. [34] used 20% FW and 80% algal residue extract to culture Schizochytrium, and both biomass and lipid production achieved good results. The results showed that compared with traditional medium, the biomass and lipid titer of strain were increased by 12.26% and 24.56%, respectively. Meanwhile, Chen et al. [7] employed a mixture of 80% FW and 20% seawater, resulting in a biomass and lipid titer of Schizochytrium at 130.10 g/L and 39.00 g/L, respectively, realizing the recycling of FW. Recycling the FW reduces the costs of fermentation and wastewater discharge. The FW of the CBH is recycled so that the lipid titer and the EPA titer are further improved, and the economic cost is reduced.
To further evaluate the advantages of reusing the CBH combined with FW to produce EPA, for which an economic analysis was conducted. In this study, a rough estimate of the cost of processing 1 ton of corncobs was prepared using cellulase hydrolysis to convert the corncobs to glucose (Supplementary Table S2). After pretreatment and enzymatic hydrolysis, the glucose yield per ton of corncob was 259.51 kg. At present, the unit price of glucose is about 0.6 US$/kg, and the procurement cost of equivalent glucose is 155.71 US$. The cost of CBH as a carbon source was 80.69% of that of glucose. In addition, through the FW recycling technology, the wastewater treatment cost can be reduced by 40%, and the total cost can be further reduced. Using CBH as a carbon source combined with recycling of FW not only promoted EPA accumulation in Schizochytrium fermentation but also reduced EPA production cost.
In conclusion, the fermentation strategy of using CBH as an alternative carbon source for culturing Schizochytrium and combining it with wastewater recycling has potential application value and provides a novel method to further reduce the cost of EPA production and wastewater treatment, and realize green recycling fermentation.
Conclusions
This study reports the potential use of CBH as an effective carbon source for EPA biosynthesis in Schizochytrium sp.; replacing freshwater with up to 40% FW yielded the best results among all tested ratios. The FW recycling strategy using CBH as the carbon source was beneficial for the accumulation of EPA in Schizochytrium sp. and increased its potency. This approach lowers production expenses and waste management costs and mitigates environmental contamination. Pilot-scale fermentation process validation should be conducted in the future to test the industrial application potential of this strategy. In addition, alternative materials with more cost advantages can be screened to optimize the medium components and improve the fermentation process parameters to further improve the EPA fermentation potency.
Data availability
No datasets were generated or analyzed during the current study.
References
-
Innes JK, Calder PC. Marine omega-3 (N-3) fatty acids for cardiovascular health: an update for 2020. Int J Mol Sci. 2020. https://doi.org/10.3390/ijms21041362.
-
Wood AHR, Chappell HF, Zulyniak MA. Dietary and supplemental long-chain omega-3 fatty acids as moderators of cognitive impairment and Alzheimer’s disease. Eur J Nutr. 2022;61(2):589–604. https://doi.org/10.1007/s00394-021-02655-4.
-
Rasha F, Kahathuduwa C, Ramalingam L, Hernandez A, Moussa H, Moustaid-Moussa N. Combined effects of eicosapentaenoic acid and adipocyte renin-angiotensin system inhibition on breast cancer cell inflammation and migration. Cancers. 2020;12(1):1–17. https://doi.org/10.3390/cancers12010220.
-
Chiu CF, Hsu MI, Yeh HY, Park JM, Shen YS, Tung TH, Huang JJ, Wu HT, Huang SY. Eicosapentaenoic acid inhibits KRAS mutant pancreatic cancer cell growth by suppressing hepassocin expression and stat3 phosphorylation. Biomolecules. 2021;11(3):1–16. https://doi.org/10.3390/biom11030370.
-
Barta DG, Coman V, Vodnar DC. Microalgae as sources of omega-3 polyunsaturated fatty acids: biotechnological aspects. Algal Res. 2021. https://doi.org/10.1016/j.algal.2021.102410.
-
Bao Z, Zhu Y, Zhang K, Feng Y, Chen X, Lei M, Yu L. High-value utilization of the waste hydrolysate of Dioscorea zingiberensis for docosahexaenoic acid production in Schizochytrium sp. Bioresour Technol. 2021;336: 125305. https://doi.org/10.1016/j.biortech.2021.125305.
-
Chen ZL, Yang LH, He SJ, Du YH, Guo DS. Development of a green fermentation strategy with resource cycle for the docosahexaenoic acid production by Schizochytrium sp. Bioresour Technol. 2023;385: 129434. https://doi.org/10.1016/j.biortech.2023.129434.
-
Ma W, Zhang Z, Yang W, Huang P, Gu Y, Sun X, Huang H. Enhanced docosahexaenoic acid production from cane molasses by engineered and adaptively evolved Schizochytrium sp. Bioresour Technol. 2023;376: 128833. https://doi.org/10.1016/j.biortech.2023.128833.
-
Dahmen N, Lewandowski I, Zibek S, Weidtmann A. Integrated lignocellulosic value chains in a growing bioeconomy: status quo and perspectives. G C B Bioenergy. 2019;11(1):107–17. https://doi.org/10.1111/gcbb.12586.
-
Olsen PM, Kosa G, Kluver M, Kohler A, Shapaval V, Horn SJ. Production of docosahexaenoic acid from spruce sugars using Aurantiochytrium limacinum. Bioresour Technol. 2023;376: 128827. https://doi.org/10.1016/j.biortech.2023.128827.
-
Qi F, Zhang M, Chen Y, Jiang X, Lin J, Cao X, Huang J. A lignocellulosic hydrolysate-tolerant Aurantiochytrium sp. mutant strain for docosahexaenoic acid production. Bioresour Technol. 2017;227:221–6. https://doi.org/10.1016/j.biortech.2016.12.011.
-
Wang LR, Zhang ZX, Wang YZ, Li ZJ, Huang PW, Sun XM. Assessing the potential of Schizochytrium sp. HX-308 for microbial lipids production from corn stover hydrolysate. Biotechnol J. 2022;17(5): e2100470. https://doi.org/10.1002/biot.202100470.
-
Wang Q, Li X, Liu W, Zhai S, Xu Q, Huan C, Nie S, Ouyang Q, Wang H, Wang A. Carbon source recovery from waste sludge reduces greenhouse gas emissions in a pilot-scale industrial wastewater treatment plant. Environ Sci Ecotechnol. 2023;14: 100235. https://doi.org/10.1016/j.ese.2022.100235.
-
Yao S, Xiong L, Chen X, Li H, Chen X. Comparative study of lipid production from cellulosic ethanol fermentation wastewaters by four oleaginous yeasts. Prep Biochem Biotech. 2021;51(7):669–77. https://doi.org/10.1080/10826068.2020.1852416.
-
Laltha M, Sewsynker-Sukai Y, Gueguim Kana EB. Simultaneous saccharification and citric acid production from paper wastewater pretreated banana pseudostem: optimization of fermentation medium formulation and kinetic assessment. Bioresour Technol. 2022;361: 127700. https://doi.org/10.1016/j.biortech.2022.127700.
-
Nunui K, Boonsawang P, Chaiprapat S, Charnnok B. Using organosolv pretreatment with acid wastewater for enhanced fermentable sugar and ethanol production from rubberwood waste. Renew Energ. 2022;198:723–32. https://doi.org/10.1016/j.renene.2022.08.068.
-
Song X, Ma Z, Tan Y, Zhang H, Cui Q. Wastewater recycling technology for fermentation in polyunsaturated fatty acid production. Bioresour Technol. 2017;235:79–86. https://doi.org/10.1016/j.biortech.2017.03.034.
-
Ou Y, Qin Y, Feng SS, Yang HL. Dual stress factors adaptive evolution for high EPA production in Schizochytrium sp. and metabolomics mechanism analysis. Bioproc Biosyst Eng. 2024;47(6):863–75. https://doi.org/10.1007/s00449-024-03013-4.
-
Ou Y, Li YQ, Feng SS, Wang Q, Yang HL. Transcriptome analysis reveals an eicosapentaenoic acid accumulation mechanism in a Schizochytrium sp. mutant. Microbiol Spectr. 2023;11(3):1–15. https://doi.org/10.1128/spectrum.00130-23.
-
Chang YH, Chang KS, Hsu CL, Chuang LT, Chen CY, Huang FY, Jang HD. A comparative study on batch and fed-batch cultures of oleaginous yeast Cryptococcus sp. in glucose-based media and corncob hydrolysate for microbial oil production. Fuel. 2013;105:711–7. https://doi.org/10.1016/j.fuel.2012.10.033.
-
Mukasekuru MR, Kaneza P, Sun H, Sun FF, He J, Zheng P. Fed-batch high-solids enzymatic saccharification of lignocellulosic substrates with a combination of additives and accessory enzymes. Ind Crop Prod. 2020;146: 112156. https://doi.org/10.1016/j.indcrop.2020.112156.
-
Zhao B, Li Y, Mbifile MD, Li C, Yang H, Wang W. Improvement of docosahexaenoic acid fermentation from Schizochytrium sp. AB-610 by staged pH control based on cell morphological changes. Eng Life Sci. 2017;17:981–8. https://doi.org/10.1002/elsc.201600249.
-
Tan X, Wu X, Wang XZ, Tao YH, Li YB, Tang S, Zhang Q, Sheng YQ. Chitosan-assisted dilute acid pretreatment with enhanced prehydrolysate fermentability and enzyme digestibility of pretreated substrates. Int J Biol Macromol. 2025;310: 143231. https://doi.org/10.1016/j.ijbiomac.2025.143231.
-
Jilani SB, Olson DG. Mechanism of furfural toxicity and metabolic strategies to engineer tolerance in microbial strains. Microb Cell Fact. 2023;22(1):221. https://doi.org/10.1186/s12934-023-02223-x.
-
Allen SA, Clark W, McCaffery JM, Cai Z, Lanctot A, Slininger PJ, Liu ZL, Gorsich SW. Furfural induces reactive oxygen species accumulation and cellular damage in Saccharomyces cerevisiae. Biotechnol Biofuels. 2010;3: 2.
-
Vasilakis A, Rigos EM, Giannakis N, Diamantopoulou P, Papanikolaou S. Spent mushroom substrate hydrolysis and utilization as potential alternative feedstock for anaerobic co-digestion. Microorganisms. 2023;11(2):1–17. https://doi.org/10.3390/microorganisms11020532.
-
Huang C, Li YY, Liu LP, Wu H, Zong MH, Lou WY. Kinetics and mechanism analysis on microbial oil production by Trichosporon fermentans in rice straw hydrolysate. Ind Eng Chem Res. 2014;53(49):19034–43. https://doi.org/10.1021/ie502185u.
-
Chen X, Li Z, Zhang X, Hu F, Ryu DD, Bao J. Screening of oleaginous yeast strains tolerant to lignocellulose degradation compounds. Appl Biochem Biotechnol. 2009;159(3):591–604. https://doi.org/10.1007/s12010-008-8491-x.
-
Watanabe K, Nishijima M, Mayuzumi S, Aki T. Utilization of sugarcane bagasse as a substrate for lipid production by Aurantiochytrium sp. J Oleo Sci. 2022;71(10):1493–500. https://doi.org/10.5650/jos.ess22206.
-
Perez-Garcia O, Escalante FM, De-Bashan LE, Bashan Y. Heterotrophic cultures of microalgae: metabolism and potential products. Water Res. 2011;45:11–36. https://doi.org/10.1016/j.watres.2010.08.037.
-
Burgstaller L, Loffler S, De Marcellis L, Ghassemi K, Neureiter M. The influence of different carbon sources on growth and single cell oil production in oleaginous yeasts Apiotrichum brassicae and Pichia kudriavzevii. N Biotechnol. 2022;69:1–7. https://doi.org/10.1016/j.nbt.2022.02.003.
-
Shafiq M, Zeb L, Cui G, Jawad M, Chi Z. High-density pH-auxostat fed-batch culture of Schizochytrium limacinum SR21 with acetic acid as a carbon source. Appl Biochem Biotechnol. 2020;192(4):1163–75. https://doi.org/10.1007/s12010-020-03396-6.
-
Wang Y, Ho SH, Cheng CL, Guo WQ, Nagarajan D, Ren NQ, Lee DJ, Chang JS. Perspectives on the feasibility of using microalgae for industrial wastewater treatment. Bioresour Technol. 2016;222:485–97. https://doi.org/10.1016/j.biortech.2016.09.106.
-
Yin FW, Guo DS, Ren LJ, Ji XJ, Huang H. Development of a method for the valorization of fermentation wastewater and algal-residue extract in docosahexaenoic acid production by Schizochytrium sp. Bioresour Technol. 2018;266:482–7. https://doi.org/10.1016/j.biortech.2018.06.109.
-
Chen W, Zhou P, Zhu Y, Xie C, Ma L, Wang X, Bao Z, Yu L. Improvement in the docosahexaenoic acid production of Schizochytrium sp. S056 by replacement of sea salt. Bioproc Biosyst Eng. 2016;39(2):315–21. https://doi.org/10.1007/s00449-015-1517-1.
-
Hu XC, Ren LJ, Chen SL, Zhang L, Ji XJ, Huang H. The roles of different salts and a novel osmotic pressure control strategy for improvement of DHA production by Schizochytrium sp. Bioproc Biosyst Eng. 2015;38:2129–36. https://doi.org/10.1007/s00449-015-1452-1.
-
Emily H, Rahul K, Seetharaman V, Gilmour D, Phillip W. The search for a lipid trigger: The effect of salt stress on the lipid profile of the model microalgal species Chlamydomonas reinhardtii for biofuels production. Current Biotechnol. 2016;5:305–13. https://doi.org/10.2174/2211550105666160322234434.
-
Bagul VP, Annapure US. Effect of sequential recycling of spent media wastewater on docosahexaenoic acid production by newly isolated strain Aurantiochytrium sp. ICTFD5. Bioresour Technol. 2020;306: 123153. https://doi.org/10.1016/j.biortech.2020.123153.
-
Sun XM, Ren LJ, Bi ZQ, Ji XJ, Zhao QY, Huang H. Adaptive evolution of microalgae Schizochytrium sp. under high salinity stress to alleviate oxidative damage and improve lipid biosynthesis. Bioresour Technol. 2018;267:438–44. https://doi.org/10.1016/j.biortech.2018.07.079.
-
Patel A, Liefeldt S, Rova U, Christakopoulos P, Matsakas L. Co-production of DHA and squalene by thraustochytrid from forest biomass. Sci Rep. 2020;10:1992. https://doi.org/10.1038/s41598-020-58728-7.
-
Nguyen HC, Su CH, Yu YK, Huong DT. Sugarcane bagasse as a novel carbon source for heterotrophic cultivation of oleaginous microalga Schizochytrium sp. Ind Crop Prod. 2018;121:99–105. https://doi.org/10.1016/j.indcrop.2018.05.005.
-
He Y, Zhu X, Ning Y, Chen X, Sen B, Wang G. Saturated and polyunsaturated fatty acids production by Aurantiochytrium limacinum PKU#Mn4 on Enteromorpha hydrolysate. Mar Drugs. 2023;21(198):1–13. https://doi.org/10.3390/md21040198.
Acknowledgements
This work was supported by the Key Experimental Open Project of Ministry of Education of Industrial Biotechnology (no. KLIB-KF202304), Taishan Industrial Experts Programme (NO.tscx202306112).
Funding
The work was financially supported by the Key Experimental Open Project of Ministry of Education of Industrial Biotechnology (no. KLIB-KF202304), Taishan Industrial Experts Programme (NO.tscx202306112).
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ou, Y., Qin, Y., Wang, Y. et al. Co-utilization of corncob hydrolysate and fermentation wastewater for eicosapentaenoic acid production by Schizochytrium sp.. Biotechnol. Biofuels Bioprod. 18, 94 (2025). https://doi.org/10.1186/s13068-025-02692-9
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1186/s13068-025-02692-9