Introduction
Restriction factors (RFs) are involved in the response to numerous viral infections. They are part of the innate immune system and most of them are both activators and targets of the interferon response (reviewed in1). They are of various origins, are usually efficient against a wide variety of viruses and can interfere at multiple steps of the viral replicative cycles. Among this wide variety of factors, interferon induced trans-membrane (IFITM) and serine incorporator (SERINC) proteins constitute 2 unrelated families of genes encoding membrane-associated proteins, some of which possess antiviral activities. SERINC3 and SERINC5 are integral membrane proteins that can be incorporated in the membrane of enveloped viral particles and limit their infectivity. Both proteins have been described as active against HIV-1 derived particles, among others, in the absence of the HIV-1 encoded Nef protein that can counteract their activities2,3,4,5. SERINC3 and SERINC5 interfere with cell and virus membrane fusion steps via different mechanisms and also with innate immune responses through IFN-I and NF-κB signalling (reviewed in6). The IFITM family of proteins comprises 5 genes all situated in a locus on human chromosome 11. Among them, only IFITM1, 2 and 3 proteins are considered “immunity-related” and have been shown to be effective RFs. They are membrane-associated proteins, and can interfere with numerous viruses (including HIV-1), enveloped or not, via different mechanisms (7, reviewed in1,8,9). Like SERINC proteins, they can interfere with the fusion step between the cellular and viral membranes early in the infection cycle of enveloped viruses and Nef has recently been reported to impair IFITM3 function10. However, unlike SERINC3 and SERINC5 that are incorporated on viral particles, IFITM1-3 block membrane fusion when expressed by the target cells.. They can also disturb other steps of viral cycles, including the production of viral particles and the incorporation of the viral glycoproteins during virus assembly (11,12,13, reviewed in1,8,9). IFITM1 possesses a cellular sub-localisation different from IFITM2 and IFITM3 (cell membrane vs endosomes), and this difference has been reported to account for the different activities of the proteins: IFITM1 would block viruses whose membrane fuses with the cell surface membrane whereas IFITM2 and IFITM3 would block the entry of viruses that use the endosomal pathway (14,15,16, reviewed in1). However a recent report indicate that coexpression of IFITM1 and IFITM3 partially relocalises IFITM1 to late endosomes and lysosomes where IFITM3 is present, suggesting that the respective roles of the 3 IFITM proteins may be more complex17. Altogether, the exact mechanisms of action are multiple, not all well understood and may sometimes appear different depending on the system used in the studies. In particular, RFs have initially been described with infectious replicative viruses, and their effects can vary when using non replicative pseudotyping systems.
In recent years, gene therapy projects have considerably expanded, with numerous potential applications investigated and new treatments marketed. Some of these promising products are based on HIV-1-derived lentiviral vectors which provide stable transduction of various cell types18. These vectors have been refined over the years to increase their safety and efficiency, removing all potentially harmful or unnecessary viral proteins from the HIV-1 element18,19,20. Among those suppressed factors are “accessory” proteins (namely Nef, Vif, Vpr and Vpu) that are not required for the infectious cycle of HIV-1 virus in some cell lines, and also not needed for single-cycle integration of non-replicative vectors. However, it was found that their role in vivo consists of (but is not necessarily limited to) counteracting the effects of several RFs (eg Nef, Vif and Vpu respectively can inhibit SERINC/IFITM3, APOBEG3G and BST2 proteins), permitting a productive HIV-1 infection2,3,10, reviewed in21). The removal of these accessory proteins from lentiviral vectors (LVs) therefore potentially lowers/hampers their efficacy to transduce cells expressing RFs.
LVs are highly efficient tools to transduce most cell lines, but their transduction efficacy is often much lower in primary cells. For clinical applications, VSV-G is currently the choice glycoprotein for pseudotyping LVs because of its broad tropism, high titres and resistance allowing purification without infectivity loss. However, several cell types of major interest, including B cells and resting HSPCs and T cells, have been shown to express low levels of LDL receptors, the cellular proteins used to mediate entry via VSV-G, leading to a strongly reduced LV transduction rate22. Different glycoproteins whose receptor is better expressed in these cells can be used to replace VSV-G (23,24,25,26,27,28, reviewed in 29). These glycoproteins, especially those of retroviral origin, often requires the addition of compounds so called “transduction adjuvants” that enhance transduction efficiency. Cationic compounds such as polybrene, protamine sulfate or semen-derived enhancer of virus infection (SEVI) can be added to increase transduction efficiency by decreasing the natural repulsion between negatively-charged cells and viral membranes and also by promoting viral aggregation, but some of these factors can be toxic to sensitive cells30,31,32. Peptides such as those derived from fibronectin (e.g. Retronectin), have dual binding domains that bring cell membrane and viral particles closer, but this strategy requires coating the peptides on the culture dishes and is more cumbersome to use than soluble adjuvants33. Other compounds act as transduction enhancers by enhancing membrane fusion through their amphiphilic properties or by enhancing the adhesion or aggregation of particles towards the cells, for instance poloxamer derivatives (e.g. LentiBoost) or amphipathic peptide such as Vectofusin34,35. Using fibronectin fragments or Vectofusin is particularly useful to obtain high levels of transduction with LVs pseudotyped with retroviral envelopes such as GALV-TR or RD11435,36. More recently, it has been recognized that cells difficult to transduce with LVs such as CD34 + hematopoietic progenitor cells, express high levels of IFITM3 RFs that could be blocked by several compounds. Rapamycin, a mTor inhibitor, caraphenol A, a resveratrol trimer and cyclosporin H, a derivative of cyclosporin A without cyclophilin inhibitory activity have been used to decrease IFITM3 endosomal expression and improve HSPCs lentiviral transduction37,38,39,40.
In this study, we developed a model system to systematically investigate the effect of the SERINC and IFITM RF expression in target cells on their transduction by LVs, depending on the nature of the envelope glycoprotein used and the transduction conditions used.
Results
1- Generation and characterisation of model cell lines
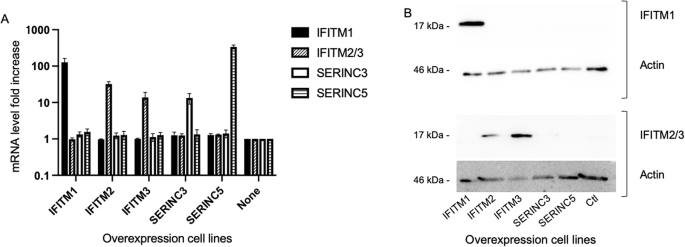
The model system is based on engineering the human HEK293T cell line to overexpress independently each of the RFs from IFITM and SERINC families, and measure the effects on lentiviral transduction efficiency. HEK293T cells are relevant models for this study as they are easy to grow and to transduce with low doses of LVs. They are also refractory to the interferon antiviral pathway activation during lentiviral transduction41. For all these reasons, the effect of each of the selected RFs can likely be tested independently of any other ones that could be otherwise induced by the transduction process. RF-overexpressing HEK293T stable cell lines were generated by lentiviral transduction using similar expression systems for each RF to obtain consistent study systems. The open reading frames (ORFs) of each of the 5 SERINC/IFITM RFs were expressed under the control of the short EF1alpha promoter with a common Kozak sequence upstream of the initiation codon and the stabilising WPRE sequence downstream of the ORF. The Hygromycin resistance gene was expressed from a separate promoter (PGK) within the LV. Following transduction with several doses of each RF-expressing LV, 293 T cells were selected with hygromycin and assessed for the number of integrated LV genomes. Populations of cells with an average vector copy number (VCN) of 1 were deemed comparable and were selected for the study. All cell lines had similar growth rates throughout the experiments and their viability was not affected, indicating that none of the overexpressed genes is susceptible to alter the results of the experiments through nonspecific mechanisms. Expression of the RF mRNAs of interest was specific in the different selected cell lines as expected (Fig. 1A). The protein expression level of IFITM proteins was detected specifically by Western blot in the IFITM-expressing cells but it was not possible in SERINC-expressing cells for which we had no antibody available for immunodetection Fig. 1B). The use of constructs with an identical structure (eg promoter, Kozak sequence and exact ORF sequence) and a VCN of 1 was chosen to compare the RF efficiency using conditions of expression as similar as possible. However qPCR data in Fig. 1A show variations in the mRNA fold increases for the 5 genes. These differences are in some part due to differences in the expression levels of the genes in the native HEK293T cells (indeed for similarly overexpressed mRNA levels, the fold increases will be higher if the initial mRNA content is lower) but are also due to the intrinsic properties of the nucleic acid and amino acid sequences that can affect their stability. Indeed, using the same primers, mRNA levels are slightly higher in the IFITM2 overexpressing cells compared to IFITM3 overexpressing cells, but western blot in Fig. 1B show more IFITM3 protein than IFITM2 probably in relation with a higher stability of IFITM3 protein.

RF levels in the overexpression HEK293T cell lines. (A). Quantification of RF mRNA expression in the overexpressing cell lines and control cells (Ctl) by qRT-PCR. Because of their very similar sequences (93,3% identity), IFITM2 and IFITM3 transcript levels were measured with the same primer pair. The results correspond to the means and standard deviations of two RNA extractions, each of them measured in duplicates. (B). Expression of IFITM proteins in the overexpressing cell lines detected by western blot. IFITM1 (Top) and IFITM2/3 (Bottom) were tested independently in all 6 cell lines and were detected only in the corresponding overexpressing cell lines (apparent MW for both 17 kDa, top panels). Total protein content in each well was checked by looking at actin content (MW 46 kDa, lower panels).
2- Variable sensitivity to RFs depending on the glycoprotein
We first tested the different cell lines for their sensitivity to transduction by LVs pseudotyped with the most commonly used glycoprotein in gene therapy, the vesicular stomatitis virus G glycoprotein (VSV-G), in the absence of any transduction adjuvant. As shown in Fig. 2, the 5 SERINC/IFITM RFs that were tested had only limited effect on the capacity of the cells to be transduced, with the maximal observed decrease in the transduction efficiency being around 25% in the cell line overexpressing IFITM3. Overexpression of IFITM2, a closely related protein, also led to a slight but significant decrease in the transduction sensitivity of the cells (approximately 10%). None of the other 3 RFs (IFITM1, SERINC3 and SERINC5) showed any effect in this assay. The observed effects were relatively small, but clearly statistically significant (p < 0.0001 for both in a paired t-test). We then tested the effects of the RFs on LVs pseudotyped with Cocal-G, a less well-studied glycoprotein, originating from the Cocal vesiculovirus, a close parent of VSV, and which is of notable clinical interest as it cannot be inactivated by normal human serum, unlike the VSV-G protein42. The results in Fig. 2 show that Cocal-G-pseudotyped LV proved very resistant to RF overexpression. Among the tested factors only IFITM3 could significantly reduce the transduction of the cells with Cocal-G LV, with a small effect limited to 13% (p = 0.006), clearly lower than that of VSV-G.

Effect of RF expression on cell transduction. The 6 model cell lines overexpressing a RF (or control) were transduced with a GFP-expressing LV pseudotyped with 4 different viral glycoproteins (indicated above each graph) and the transduction efficiency measured by flow cytometry. For each experiment, the transduction level of each RF-expressing cell line was normalised to that of the control cell line. The graphs represent the means + /- SD of independent experiments (VSV-G n = 28, Cocal-G n = 16 , Syn1 n = 21, BaEV n = 5). Significant differences between the control cell line and the RF expressing ones are indicated (t-test).
We also tested retroviral glycoproteins. First was Syncytin1 (Syn1), a human protein of retroviral origin involved in placenta formation, that can be used to pseudotype LVs27,43,44 and requires the use of Vectofusin for high levels of transduction27. Unlike what was observed with the 2 vesiculovirus glycoproteins, 4 of the 5 RFs tested noticeably reduced the transduction efficiency with Syn1. Only the SERINC3-expressing cell line showed no modification of its transducability. The strongest decrease in transduction was observed when IFITM3 was overexpressed (approximately 75% decrease), followed by IFITM2 (55%), IFITM1 (18%) and SERINC5 (approx. 10%). All these effects were highly statistically significant. We then determined the sensitivity profile of another glycoprotein of clinical interest for its ability to transduce CD34 + cells, the baboon endogenous retrovirus (BaEV) glycoprotein used to transduce T and B cells26,45. As shown in Fig. 2, the transduction rates of all 6 cell lines are similar with LV pseudotyped with BaEV Env, indicating that this glycoprotein is not affected by any of the 5 RFs that we tested.
3- Modulation of the RF effects by transduction adjuvants
The previous series of experiments was performed in the absence of any transduction adjuvants, which is mostly relevant for standard cell lines that can easily be transduced. However, LV transduction is often much less efficient on primary cells, requiring the use of additional compounds to achieve the required levels of vector copies in such cells. We tested if the RF effects that we identified were modified by the use of such transduction adjuvants. Among those, we tested polybrene, protamine sulfate, cyclosporin H and Vectofusin that cover different chemical families and modes of actions31,34,35,40. The tested adjuvants were added individually at the same time as the LV and the transduction rates of each cell line were determined using flow cytometry as described above. For each condition (ie combination of glycoprotein, RF and nature of the adjuvant) we quantified the effect of the RF as the decrease in infectivity its expression induces (ie effect = [Titre Ctl cell line] / [Titre RF cell line]). The statistical significance of the differences in RF efficiency measured with or without each transduction adjuvants was tested in paired experiments. As shown in Fig. 3, the sensitivity of the vesiculovirus-derived glycoproteins to the tested RFs was not modified by adding polybrene or protamine sulfate. Similarly, the results obtained with BaEV Env pseudotyped LV showed little changes. It should be noted that this may be due to the fact that these 3 glycoproteins are the more resistant to the effects of the RFs, making variations harder to detect. On the contrary, Syn1-pseudotyped LV were strongly affected by some of the adjuvants, mostly cyclosporin H and Vectofusin. Both adjuvants significantly and strongly decreased the inhibitory effects of IFITM3, and of IFITM2 in a more limited way, while IFITM1-mediated restriction was not affected. The effects of cyclosporin H were expected based on previous studies showing that cyclosporin H can block IFITM3 action in human CD34 + cells and THP-1 cell line40,46. The effects of Vectofusin are novel and appeared to be slighly lower than those of cyclosporin H.

Modulation of the effects of RFs on cell transduction in the presence of transduction adjuvants. The 6 model cell lines were transduced with eGFP-expressing LVs pseudotyped with different viral glycoproteins in the presence or absence of transduction adjuvants. The effect of each RF was measured for each condition as the fold decrease it induces in transduction (= [transduction efficiency Ctl] / [transduction efficiency RF]) and was plotted as mean + /- SD. Significant differences between the fold decreases measured with or without adjuvants in paired experiments are indicated (t-test). (graphs: 5 < n < 28; stats: 4 < n < 15).
Effects of cyclosporin H and Vectofusin on IFITM2/3 protein levels
Cyclosporin H is a powerful compound to enhance lentiviral transduction of CD34 + cells40,46. The addition of cyclosporin H during the transduction of CD34 + cells with a VSV-G-pseudotyped LV decreases their IFITM3 content as measured by intracellular staining and results in higher vector copy number integration (Fig. 4A). Vectofusin is known to enhance the transduction of CD34 + hematopoietic progenitor cells with VSV-G-pseudotyped lentiviral vectors35 but the effects are more modest than those of cyclosporin H (Fig. 4B) and therefore less likely to be detected by examining an effect on IFITM3 levels. To confirm the relationship between IFITM3 levels reduction capacity and transduction enhancement, the effects of cyclosporin H were compared to that of Vectofusin in the overexpression model 293 T cell lines. Total IFITM2/3 content was quantified by flow cytometry after intracellular staining for IFITM2, IFITM3 in overexpressing cells and control after treatment or not with cyclosporin H or Vectofusin for 17 h (Fig. 5). Both compounds induced a decrease in IFITM2/3 expression level, and the differences in mean fluorescence intensity (MFI) were statistically significant (t-test in paired experiments). Interestingly, the decrease was stronger with cyclosporin H, consistent with the more pronounced inhibition of IFITM3 restriction seen in Fig. 3. Of note, both effects seem specific since the total amount of LAMP1 protein, an endo-lysosomal protein that partially colocalises with IFITM3, was unaffected in the same conditions.

Enhancement of CD34 + cell transduction using adjuvants. (A) The effect of cyclosporin H (CsH) addition on IFITM2/3 expression during the transduction of CD34 + cells with two rounds of VSV-G-pseudotyped LV was determined by flow cytometry 24 h after the start of the transduction (left). The corresponding transduction efficiencies were quantified by measuring the VCN using ddPCR 5 days adter transduction (right). Cells were isolated from 3 independent donors. (B). Comparison between the effects of the addition of cyclosporin H and Vectofusin (VF) on CD34 + cell transduction efficiency with a single round of VSV-G pseudotyped LV. Transduction levels were determined by measuring the VCNs by ddPCR on cells isolated from 3 independent donors. Significant differences are indicated (t-test).

Effects of Vectofusin and cyclosporin H treatment on the expression levels of IFITM2/3 and LAMP1 proteins in IFITM2, IFITM3 overexpressing cell lines and controls. Cells were treated for 17 h with Vectofusin or cyclosporin H and total amounts of IFITM2/3 and LAMP1 proteins were determined by flow cytometry after intracellular staining. Cells were gated based on SSC / FSC criteria and the the MFI of the whole population was measured for each protein. (n = 4 for IFITM2/3, n = 3 for LAMP1). The effects of Vectofusin and cyclosporin H on MFI of each cell population were tested (paired t-test) and statistically significant differences are indicated.
Discussion
In this study, we generated over-expressing cell lines to investigate the effect of IFITM and SERINC proteins to restrict LV transduction. These 2 families of proteins had already been shown in some settings to affect the fusion step between cell and virus membranes that occurs early in infection, where they play “symmetric roles” with SERINC proteins acting while incorporated on the virus membrane while IFITM are present in the membranes of the target cells. While no effect of SERINC proteins was identified in the system we tested (ie when RFs are expressed by the target cells), in agreement with previous findings (reviewed in6), we found that IFITM3 was the most potent RF in the model among those tested, and also demonstrated that IFITM2, the product of a recent IFITM3 gene duplication, had similar, albeit weaker, effects than IFITM3. Interestingly, the extent of the transduction inhibition depended on the nature of glycoprotein used to pseudotype the LVs with unexpected differences between retroviral envelope glycoproteins. We confirmed that cyclosporin H can decrease intracellular IFITM3 protein content and relieve the restriction effects on LVs, but describe for the first time that Vectofusin, which is known as a fusion enhancing and particle aggregating adjuvant, has similar properties.
Among the 5 RFs tested, we report that IFITM3 proteins restrict the transduction with VSV-G -pseudotyped LVs as already reported40, but we also see an effect of IFITM2. IFITM2 arose from a recent gene duplication of IFITM3 and the 2 proteins have very similar sequences47. While many previous studies mostly concentrate on IFITM3 protein, the ancestral gene, IFITM2 effects are generally less studied but our data shows that IFITM2 retained some antiviral effects, although less potent than those of IFITM3. Among vesiculovirus glycoproteins, our assay shows that the Cocal-G protein is more resistant than its VSV counterpart to both IFITM2 and IFITM3 proteins. This adds up to other interesting properties of the Cocal-G variant for gene therapy, like its resistance to human serum inactivation42. Nowadays VSV-G protein is the gold standard for LV-based clinical applications, but our results may provide further justification to use Cocal-G-pseudotyped LVs in gene therapy especially since IFITM3 is known to be a major factor in the limitation to human CD34 + cell transduction with LVs38,39,40,46.
We also tested the effect of the expression of IFITM and SERINC RFs on LVs pseudotyped with 2 retroviral envelope proteins that are good candidates to efficiently transduce B cells26,27, unlike VSV-G whose receptor is not expressed in these cells22. Our results indicate that the sensitivity is independent of the viral family, with BaEV Env and Syncytin1-pseudotyped LVs showing very different behaviour. The BaEV Env is resistant to all 5 RFs, with no statistically significant effect detected, whereas Syn1-mediated transduction is impaired by all 3 IFITM proteins, with IFITM3 being the most efficient. This is all the more surprising that BaEV Env and Syn1 are using the same cellular proteins hASCT1 and hASCT2 as receptors for cell infection44,48 and therefore were expected to use the same mechanisms of cell entry. It is however possible that the 2 proteins have different intrinsic properties that affect their sensitivity to IFITM proteins. Despite being phylogenetically a retroviral envelope protein, Syn1 is an endogenous envelope gene whose ancestor was coopted approximately 25 million years ago in the primate lineage for its highly fusogenic properties, the characteristics responsible for its present physiological role49. Syn1 conservation of infectious properties is not subjected to any selection pressure, and its hyperfusogenicity property is very likely associated with a lower stability. Retroviral Env interactions with a receptor (or pH acidification) lead to conformational changes in Env proteins enabling fusion of virus and cell membranes. The hyperfusogenicity of Syn1 associated with its decreased overall stability could result in different properties compared to BaEV Env even though they reportedly use the same receptors. Also, it was recently shown that, among other mechanisms, IFITM3 blocks viral infection/transduction by mimicking SNARE proteins and increasing heterotypic endosome fusion (with lysosomes), to the detriment of homotypic fusion, thus promoting viral particles degradation13. If for example BaEV induces cell and virus membrane fusion quicker than Syn1, BaEV-pseudotyped particles would be less sensitive to IFITM3 restriction, in agreement with our observations.
Finally, we tested the effects of several transduction adjuvants on IFITM-mediated restriction. No strong effect was detected for either vesiculovirus glycoproteins or the BaEV Env. These 3 proteins are the ones showing the less sensitivity to the tested RFs, and this means it would be much harder to detect any anti RF effect of the adjuvants because the initial variations are very low. However, we did not detect any enhancing effect of the adjuvants, indicating that they are at best option for LV transduction in the presence of RFs. The situation is different for Syn1-pseudotyped LV, that are much more sensitive to IFITM proteins restriction. Polybrene and Protamine sulfate show very limited, if any, effect on IFITM proteins restriction. However, Vectofusin and cyclosporin H are able to reduce the inhibiting effect of both IFITM2 and IFITM3, with cyclosporin H being more effective. The restriction effect observed with IFITM1 is unaffected by either of these 2 compounds, in agreement with IFITM1 and IFITM2/3 having distinct mechanisms of viral restriction. The effect detected with cyclosporin H is in agreement with studies that showed it can impair IFITM3 restriction in human CD34 + cells and show that the effect and extends to another cell type40,46. The mechanisms at play seem to be similar, with cyclosporin H treatment leading to a decrease in total amount of IFITM2/3, therefore counteracting their antiviral effect. Surprisingly, we observed that Vectofusin, a peptide thought to promote essentially entry by facilitating membrane fusion and viral particles adhesion and aggregation, also leads to a decrease in the protein levels of IFITM2/3. The lower IFITM2/3 inhibiting effect of Vectofusin compared to cyclosporin H correlates with a weaker effect on total IFITM proteins content, but we cannot exclude that cyclosporin H could also impact LV transduction efficiency through other independent mechanisms. Noteworthily, the IFITM2/3-inhibiting effect carried by Vectofusin is consistent with the modest effect on the transduction of CD34 + cells with VSV-G-pseudotyped lentiviral vectors. However, a previous study showed that Vectofusin strongly improves the transduction of human CD34 + cells with vectors pseudotyped with the retroviral envelope GALV-TR presumably due to increased viral:cell membrane fusion, particle adhesion and aggregation, although no link to IFITM3 was made at the time35,50. Our data suggest that such enhancing effect of Vectofusin is linked to a greater sensitivity of some retroviral envelopes to restriction factors. The effects of Vectofusin and cyclosporin H seem rather specific since we did not detect any variation in the protein level of LAMP1, an endo-lysosomal protein that partially colocalises with IFITM2/3. The molecular mechanisms leading to IFITM2/3 depletion by these 2 very chemically distinct compounds are not yet characterised. Interestingly, a series of mTOR inhibitors have also been successfully used to decrease IFITM2/3 protein levels and enhance LV transduction in different cell models38,51. In particular, Caraphenol A addition enhances LV transduction of CD34 + cells while decreasing their IFITM2/3 content, similar to the effects of Vectofusin and cyclosporin H39. It would be very interesting to determine if these compounds use similar or independent mechanisms to alter IFITM2/3 content, and if additive effects could be reached. Further studies would be of great interest and could help improve both our understanding of cell intrinsic immunity and gene therapy protocols for cells resistant to LV transduction.
Methods
Procurement of human cells
The use of human cells was performed in accordance with international ethical guidelines and in accordance with french regulation applying to non-interventional research on human subjects. The use of human cell lines and the use of umbilical cord blood samples collected anonymously at the CHSF hospital in Evry, following uncomplicated birth and with the mother’s informed consent, were approved by the french ministry of higher education and research under reference codecoh DC 2019–3578.
Plasmids
pMD2G (#12,259) and pRSV Rev (#12,253) were obtained from Addgene. pMD2 Cocal-G was a gift from HP Kiem, Fred Hutchinson Cancer Research Center). pMDLG-pRRE Kana was derived from pMDLG-pRRE (obtained from Addgene #12,251) by replacing the Ampicillin resistance cassette by a Kanamycin resistance cassette. BaEV-RD was obtained from Dr FL Cosset (CIRI, ENS Lyon France). pCAG Syn1 was generated by replacing eGFP with the human syncytin1 ORF (Genbank NM_014590.4) in the pCAG eGFP plasmid (Addgene #89,684). pCCL EF1short eGFP WPRE was generated from a custom generated pRRL EF1short eGFP WPRE (Vector Builder) by replacing the RSV promoter domain in the composite 5’ LTR by the CMV promoter domain. LVs for RF expression were derived from a custom-generated pRRL EF1short eGFP WPRE PGK Hygro plasmid (Vector Builder) by replacing eGFP with the ORFs for the selected RF. SERINC3 ORF was obtained from Sino Biological (HG23919-U). IFITM1, IFITM2, IFITM3 and SERINC5 ORFs were custom-synthetised (Genecust) and are identical to Genbank NM_003641.5, NM_006435.3, NM_021034.3 and NM_001174072.3 respectively). The GLOBE-AS3 LV plasmid has been decribed elsewhere52.
Cell culture, LV production and transduction
LentiX 293 T cells (TaKaRa) were grown in a humid atmosphere at 37 °C and 5% CO2 in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% heat-inactivated foetal calf serum (Biowest), 100 mg/mL streptomycin, and 100 U/mL penicillin. For LV production, LentiX 293 T cells were seeded at 5 × 106 cells per 10 cm dish. The day after seeding, cells were transfected using 35 µL Fugene 6 (Promega) and 3 µg pMDLG-pRRE Kana (HIV-1 core proteins expression), 4.5 µg lentiviral transgene-expressing vector (pCCL EFlong eGFP (eGFP LV) or pRRL EFshort RF PGK Hygro, a dual expressing vector to concomitantly express a RF and hygromycin resistance (Hygro) , 1.1 µg pRev (HIV-1 Rev expression) and 1.5 µg of glycoprotein expressing plasmid (pMD2G for VSV-G expression, pCAG Syn1 for Syncytin1 expression, BaEV-TR for BaEV GP expression and pMD2 Cocal-G for Cocal-G protein expression). Cell medium was changed the day after transfection, and LV containing supernatants were collected 40–48 h later, filtered (0.45 µm) and stored at -80 °C before use.
Stable cell lines overexpressing RFs or controls were generated by transducing LentiX 293 T cells with varying doses of LVs encoding for individual RFs and hygromycin resistance. Cells were expanded 48 h post transduction and selected for 2 weeks using 300 u/mL hygromycin B (InvivoGen). After selection was completed, genomic DNA was extracted and VCNs of the transgene were measured in each population by ddPCR as described in53. Populations with one copy of transgene were used for overexpression experiments. Titration experiments were performed in 24-well plates seeded with 1 × 105 LentiX 293 T cells or derived populations the day before adding LVs (or 5 × 104 cells in 48-well plates). LV infectious titres (TU/mL) were determined by quantifying the proportion of eGFP expressing cells by flow cytometry 72 h post transduction, using conditions were 5–30% of the cells are eGFP-positive. In some cases, adjuvants were added for transduction following the manufacturer’s instruction. Adjuvant concentrations were experimentally determined in the laboratory based on efficacy and are in the range used by others (eg31,40) or recommender by suppliers. Polybrene (Sigma), protamine sulfate (Sigma) and Vectofusin (Miltenyi) were used at 8 µg/mL, cyclosporin H (Sigma) was used at a concentration of 8 µM. No cell toxicity due to these compounds was detected. CD34 + cells were isolated from cord blood samples following uncomplicated births by magnetic bead cell sorting according to the manufacturer’s instructions (Miltenyi). CD34 + cell experiments were performed as described in46 using one or two rounds of LV transduction. Briefly, CD34 + cells (1E + 06 cells/mL) were cultured in X-VIVO15 serum-free medium without phenol red (Lonza), supplemented with penicillin (100 U/mL) and streptomycin (100 mg/mL) (Gibco), c-kit ligand (50 ng/mL), FLT3-ligand (100 ng/mL), interleukin-3 (20 ng/mL) and thrombopoietin (50 ng/mL) (Miltenyi Biotech). After 18 to 24 h of culture, the VSV-G pseudotyped, eGFP-expressing LV was added (5 × 10exp7 ig/mL) with or without adjuvant (Vectofusin 12 µg/mL or cyclosporin H 8 µM). The medium was changed after 6 h and cells were cultured in X-VIVO15 medium containing cytokines and 10% heat-inactivated fetal bovine serum (FBS) (Cytiva, Pasching, Austria) for several days before quantifying transduction by ddPCR.
qRT-PCR
Total RNA were extracted using RNA Plus kit (Macherey Nagel) and reverse transcribed (RT) with SuperScript First Strand kit (Invitrogen). qRT-PCR was done using Luna® Universal qPCR Master Mix (NEB) on a QS3 machine (Applied Biosystems). Efficacy of the PCR reactions was checked for each primer pair, and we also verified there was no amplification when no RT reaction was performed. Transcript levels were normalised to the TFIID gene and the no-RF control cell line using the ∆∆Ct method. Primer sequences are provided in supplementary Table S1.
Western blotting
Cells were lysed in PBS—NP40 1%—DOC 0,1% supplemented with protease inhibitors (Complete EDTA-free, Roche) for 30 min on ice. Insoluble cell debris were removed by centrifugation (12 000 rpm 5 min at 4 °C). Samples were subjected to SDS-PAGE using Mini-PROTEAN TGX4-15% gels (BioRad) and proteins were transferred on PVDF membranes using a semi-dry system. Membranes were blotted with anti IFITM1 antibody (Proteintech 60,074–1-Ig, 1/5000) or anti IFITM2/3 antibody (Proteintech 66,081–1-Ig, 1/5000). These 2 antibodies have been chosen because they do not cross-recognise the other members of the IFITM family. After incubation with the primary antibodies, membranes were washed 3 times in PBS – Tween 0,1% and incubated with an HRP-conjugated anti-mouse IgG secondary antibody (Cytiva NA931). Membranes were washed and signal was revealed using HRP substrate (Amersham ECL Prime, Cytiva) and read using an Imager 680 (Life Technology). Exposition times were selected on the «integrated» images automatically provided by the Imager that combine the ECL signal with a normal light photo displaying the molecular weight marker («combined» jpeg files in the Supplementary data). Figure panels were generated from files containing only the ECL signal («native ECL» tif files in the Supplementary data), with contrast and brightness adjusted for better visibility («uncropped» » tif files in the Supplementary data). Anti actin antibody (Proteintech 66,009–1-Ig, 1/10,000) was used to check total protein content in each lane after membrane stripping (ReBlot Plus Strong, Millipore).
FACS staining
LentiX 293 T-derived cells were stained using FIX & PERM kit (Invitrogen) with a mouse anti IFITM2/3 antibody (Proteintech 66,081–1-Ig, 1/200) and a rabbit anti LAMP1 (CST #9091, 1/100) in some experiments. After washing, cells were incubated with secondary antibodies (Alexa 488 anti mouse antibody, 1/200, Life Technologies) or APC—anti mouse IgG1 (1/100, Biolegend) plus Alexa 488 anti rabbit antibody (1/50, Biolegend). Specificity of the staining was checked using no primary antibody and isotype controls.
Data availability
Data are available upon request and materials may be provided upon agreement with Inserm. The point of contact for any data or questions regarding the study is Marie Dewannieux, PhD, ART-TG, 30 Rue H. Desbruères, 91,100 Corbeil-Essones, France. marie.dewannieux@inserm.fr.
References
-
Majdoul, S. & Compton, A. A. Lessons in self-defence: Inhibition of virus entry by intrinsic immunity. Nat. Rev. Immunol. 22(6), 339–352 (2022).
-
Rosa, A. et al. HIV-1 Nef promotes infection by excluding SERINC5 from virion incorporation. Nature 526, 212–217 (2015).
-
Usami, Y., Wu, Y. & Göttlinger, H. G. SERINC3 and SERINC5 restrict HIV-1 infectivity and are counteracted by Nef. Nature 526, 218–223 (2015).
-
Leonhardt, S. A. et al. Antiviral HIV-1 SERINC restriction factors disrupt virus membrane asymmetry. Nat. Commun. 14(1), 4368 (2023).
-
Diehl, W. E. et al. Influence of different glycoproteins and of the virion core on SERINC5 antiviral activity. Viruses 13(7), 1279 (2021).
-
Yu, J. et al. SERINC5 counters retroviruses and non-retroviruses. Front. Cell Infect. Microbiol. 14, 1516806 (2024).
-
Lu, J. et al. The IFITM proteins inhibit HIV-1 infection. J. Virol. 85(5), 2126–2137 (2011).
-
Shi, G., Schwartz, O. & Compton, A. A. More than meets the I: The diverse antiviral and cellular functions of interferon-induced transmembrane proteins. Retrovirology 14(1), 53 (2017).
-
Wang, J. et al. The antiviral activity of interferon-induced transmembrane proteins and virus evasion strategies. Viruses 16(5), 734 (2024).
-
Agarwal, M., et al., Restriction of HIV-1 infectivity by interferon and IFITM3 is counteracted by Nef. bioRxiv, 2025.
-
Lee, W. J. et al. IFITM proteins inhibit HIV-1 protein synthesis. Sci. Rep. 8(1), 14551 (2018).
-
Ahi, Y. S. et al. IFITM3 reduces retroviral envelope abundance and function and is counteracted by glycoGag. MBio 11(1), 10–1128 (2020).
-
Rahman, K. et al. SNARE mimicry by the CD225 domain of IFITM3 enables regulation of homotypic late endosome fusion. EMBO J. 44(2), 534–562 (2025).
-
Weston, S. et al. A membrane topology model for human interferon inducible transmembrane protein 1. PLoS ONE 9(8), e104341 (2014).
-
Weston, S. et al. Alphavirus restriction by IFITM proteins. Traffic 17(9), 997–1013 (2016).
-
Smith, S. E. et al. Interferon-induced transmembrane protein 1 restricts replication of viruses that enter cells via the plasma membrane. J. Virol. 93(6), 10–128 (2019).
-
Wilt, I., et al., IFITM1 and IFITM3 cooperate to restrict virus entry in endolysosomes. bioRxiv, 2025: p. 2025.06.01.657267.
-
Naldini, L., Trono, D. & Verma, I. M. Lentiviral vectors, two decades later. Science 353(6304), 1101–1102 (2016).
-
Zufferey, R. et al. Multiply attenuated lentiviral vector achieves efficient gene delivery in vivo. Nat. Biotechnol. 15(9), 871–875 (1997).
-
Dull, T. et al. A third-generation lentivirus vector with a conditional packaging system. J. Virol. 72(11), 8463–8471 (1998).
-
Kmiec, D. & Kirchhoff, F. Antiviral factors and their counteraction by HIV-1: many uncovered and more to be discovered. J. Mol. Cell Biol. 16(2), mjae005 (2024).
-
Amirache, F. et al. Mystery solved: VSV-G-LVs do not allow efficient gene transfer into unstimulated T cells, B cells, and HSCs because they lack the LDL receptor. Blood 123(9), 1422–1424 (2014).
-
Frecha, C. et al. Stable transduction of quiescent T cells without induction of cycle progression by a novel lentiviral vector pseudotyped with measles virus glycoproteins. Blood 112(13), 4843–4852 (2008).
-
Frecha, C. et al. Efficient and stable transduction of resting B lymphocytes and primary chronic lymphocyte leukemia cells using measles virus gp displaying lentiviral vectors. Blood 114(15), 3173–3180 (2009).
-
Girard-Gagnepain, A. et al. Baboon envelope pseudotyped LVs outperform VSV-G-LVs for gene transfer into early-cytokine-stimulated and resting HSCs. Blood 124(8), 1221–1231 (2014).
-
Levy, C. et al. Baboon envelope pseudotyped lentiviral vectors efficiently transduce human B cells and allow active factor IX B cell secretion in vivo in NOD/SCIDgammac(-/-) mice. J. Thromb. Haemost. 14(12), 2478–2492 (2016).
-
Stable pseudotyped lentiviral particles and uses thereof. Inventor Anne Galy. Assigned to Institut National de la Sante et de la Recherche Medicale INSERM, Genethon, Universite D’Evry Val D’Essonne, Filed 2017–04–21 and publication of US11993781B2 on 2024–05–28.
-
Renner, A. et al. Development of KoRV-pseudotyped lentiviral vectors for efficient gene transfer into freshly isolated immune cells. Gene Ther. 31(7–8), 378–390 (2024).
-
Gutierrez-Guerrero, A., Cosset, F. L. & Verhoeyen, E. Lentiviral vector pseudotypes: Precious tools to improve gene modification of hematopoietic cells for research and gene therapy. Viruses 12(9), 1016 (2020).
-
Toyoshima, K. & Vogt, P. K. Enhancement and inhibition of avian sarcoma viruses by polycations and polyanions. Virology 38(3), 414–426 (1969).
-
Cornetta, K. & Anderson, W. F. Protamine sulfate as an effective alternative to polybrene in retroviral-mediated gene-transfer: Implications for human gene therapy. J. Virol. Methods 23(2), 187–194 (1989).
-
Wurm, M. et al. The influence of semen-derived enhancer of virus infection on the efficiency of retroviral gene transfer. J. Gene Med. 12(2), 137–146 (2010).
-
Hanenberg, H. et al. Colocalization of retrovirus and target cells on specific fibronectin fragments increases genetic transduction of mammalian cells. Nat. Med. 2(8), 876–882 (1996).
-
Delville, M. et al. A nontoxic transduction enhancer enables highly efficient lentiviral transduction of primary murine T cells and hematopoietic stem cells. Mol. Ther. Methods Clin. Dev. 10, 341–347 (2018).
-
Fenard, D. et al. Vectofusin-1, a new viral entry enhancer, strongly promotes lentiviral transduction of human hematopoietic stem cells. Mol. Ther. Nucleic Acids 2(5), e90 (2013).
-
Piovan, C. et al. Vectofusin-1 Promotes RD114-TR-Pseudotyped Lentiviral Vector Transduction of Human HSPCs and T Lymphocytes. Mol Ther Methods Clin Dev 5, 22–30 (2017).
-
Wang, C. X. et al. Rapamycin relieves lentiviral vector transduction resistance in human and mouse hematopoietic stem cells. Blood 124(6), 913–923 (2014).
-
Shi, G. et al. mTOR inhibitors lower an intrinsic barrier to virus infection mediated by IFITM3. Proc Natl Acad Sci U S A 115(43), E10069–E10078 (2018).
-
Ozog, S. et al. Resveratrol trimer enhances gene delivery to hematopoietic stem cells by reducing antiviral restriction at endosomes. Blood 134(16), 1298–1311 (2019).
-
Petrillo, C. et al. Cyclosporine H overcomes innate immune restrictions to improve lentiviral transduction and gene editing in human hematopoietic stem cells. Cell Stem Cell 23(6), 820–832 (2018).
-
Ferreira, C. B. et al. Lentiviral vector production titer is not limited in HEK293T by induced intracellular innate immunity. Mol. Ther. Methods Clin. Dev. 17, 209–219 (2020).
-
Trobridge, G. D. et al. Cocal-pseudotyped lentiviral vectors resist inactivation by human serum and efficiently transduce primate hematopoietic repopulating cells. Mol. Ther. 18(4), 725–733 (2010).
-
An, D. S., Xie, Y. M. & Chen, I. S. Y. Envelope gene of the human endogenous retrovirus HERV-W encodes a functional retrovirus envelope. J. Virol. 75(7), 3488–3489 (2001).
-
Lavillette, D. et al. The envelope glycoprotein of human endogenous retrovirus type W uses a divergent family of amino acid transporters/cell surface receptors. J. Virol. 76, 6442–6452 (2002).
-
Bernadin, O. et al. Baboon envelope LVs efficiently transduced human adult, fetal, and progenitor T cells and corrected SCID-X1 T-cell deficiency. Blood Adv. 3(3), 461–475 (2019).
-
Mormin, M. et al. Cyclosporin H improves the transduction of CD34(+) cells with an anti-sickling globin vector, a possible therapeutic approach for sickle cell disease. Hum. Gene Ther. 35(21–22), 896–903 (2024).
-
Compton, A. A. et al. Natural mutations in IFITM3 modulate post-translational regulation and toggle antiviral specificity. EMBO Rep. 17(11), 1657–1671 (2016).
-
Blond, J. L. et al. An envelope glycoprotein of the human endogenous retrovirus HERV-W is expressed in the human placenta and fuses cells expressing the type D mammalian retrovirus receptor. J. Virol. 74, 3321–3329 (2000).
-
Mallet, F. et al. The endogenous retroviral locus ERVWE1 is a bona fide gene involved in hominoid placental physiology. Proc. Natl. Acad. Sci. U S A 101(6), 1731–1736 (2004).
-
Ingrao, D. et al. Concurrent measures of fusion and transduction efficiency of primary CD34+ cells with human immunodeficiency virus 1-based lentiviral vectors reveal different effects of transduction enhancers. Hum. Gene Ther. Methods 25(1), 48–56 (2014).
-
Shi, G., et al., Rapalogs downmodulate intrinsic immunity and promote cell entry of SARS-CoV-2. J Clin Invest, 132(24) 2022.
-
Poletti, V. et al. Pre-clinical development of a lentiviral vector expressing the anti-sickling betaAS3 globin for gene therapy for sickle cell disease. Mol. Ther. Methods Clin. Dev. 11, 167–179 (2018).
-
Corre, G. et al. Lentiviral standards to determine the sensitivity of assays that quantify lentiviral vector copy numbers and genomic insertion sites in cells. Gene Ther. 29(9), 536–543 (2022).
Acknowledgements
The authors would like to thank FL Cosset (CIRI, ENS, Lyon, France) and HP Kiem (Fred Hutchinson Cancer Research Center) for the gift of reagents, and G Penverne for technical help. We are also grateful to the mothers and to the maternity staff of the”Centre Hospitalier Sud-Francilien” (Corbeil-Essonnes, France) for the gift of umbilical cord blood. The study was funded by a grant from Genopole (Evry, France) to MD (ATIGE 2020) and also supported by equipment funds from the french national research agency (ANR-20-F2II-0001, ANR-22-AIBB-006) and by help for vectors from ANR-22-PEBI-0008 (THERA-B). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Ethics declarations
Competing interests
The author A. Galy is inventor of a patent entitled “ Stable pseudotyped lentiviral particles and uses thereof “ published as US11993781B2. The other authors have no competing interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Dewannieux, M., Fofana, F., Hamon, K. et al. Combined effects of restriction factors and transduction adjuvants on lentiviral vector gene transfer efficacy. Sci Rep 15, 38643 (2025). https://doi.org/10.1038/s41598-025-22470-9
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-22470-9