Introduction
Globally, researchers are becoming more interested in exploring the potential of plant–soil–microbiome interactions to enhance soil health, plant growth, and crop productivity1. However, despite several breakthroughs in using soil microbes for plant benefits2, the ultimate result of attaining sustainable agriculture that promises food security for the population boom projected for the future still seems unrealistic. This is because, as established by the Committee on Science Breakthroughs 2030, the detailed understanding of the core microbiomes which is key to exploring plant-soil-microbiome interactions for enhanced agricultural productivity efficiently3, is still minimal. As a result, the agroecosystem, an actively complex environment with an enormous variety of interactions between the three factors; soil component-plant-and-microorganisms—has received significant attention4. This dynamic environment embodies several beneficial microorganisms (not yet culturable) that are potential tools to sustain food security amidst increasing global food demand5. An outstanding example of this ecosystem is a tiny region of the soil around plant roots, the rhizosphere notable for its essential role in shaping microbial community composition, diversity, and activity6. With the influence of different metabolites (exudates) such as organic compounds released by plants, this zone is a hot spot for diverse microbial communities with distinct activities and functions7. Meanwhile, the diversity and interplay between this rhizo-microbiome i.e. microbial community inhabiting the rhizosphere, contribute significantly to soil health and nutrient cycling which are key to the growth and quality of crops especially rhizome crops such as carrots8,9. The rhizosphere of carrot with an array of microbiomes10 falls among such habitats to look for promising microbial taxa, hence a study on the community structure and diversity will inform their intricate interactions that could be harnessed for sustainable agriculture and other biotechnological applications.
Carrot (Daucus carota L.) is among the important vegetable crops cultivated worldwide for its edibility, health benefits associated with its nutritional constituents such as β-carotene, vitamins, and essential minerals, and agricultural importance11. As a taproot that serves as the plant root, the development, health, and productivity of this vegetable crop are notably influenced by complex interactions between the soil components, soil microbes, and the plant itself12. In crop rotation systems, carrot is often explored to address critical challenges in agriculture, such as breaking pest cycles, minimizing soil-borne diseases, and loosening compacted soil for root growth and water infiltration13. These practices are remarkable in enhancing nutrient availability, and improving soil structure and fertility for subsequent plants14. The preceding crop in a rotation system deposits vital organic compounds and nutrients into the soil, which in turn, determines the structure and diversity of the microbial community in the soil15. These benefits make crop rotation increasingly attractive as an alternative to monoculture systems, which over time result in soil deterioration, accumulating pests and diseases, and declining agricultural produce16. For instance, leguminous crops such as soybean which naturally fixes atmospheric nitrogen, will enrich soil fertility and improve microbial diversity, thereby shaping the intricate triparty interactions within the rhizosphere of subsequent crops. Aside from boosting crop yields, this process reduces greenhouse gas emissions and enhances soil health, all of which support sustainable agriculture17. Despite the advantages, exploring the rhizosphere microbiome for sustainable agriculture remains a mirage without knowing their composition and diversity which is key to identifying their role in nutrient cycling, plant health, and disease control18. Furthermore, the influence of preceding crops on the composition and diversity of this rhizo-microbiome is critical for leveraging them to improve plant productivity with sustainable agricultural practices19.
Substantial advances have been made in understanding microbial communities in plant rhizosphere and their ecological role, still, research on the core rhizo-microbiome of carrot is scarce. In particular, how different cropping systems, including monoculture and rotation affect the composition and diversity of carrot rhizo-microbiome is largely understudied. The few previous works that particularly employed metagenomics approaches had diverse focuses including the impact of soil management practices and fertilization regimes1,10 in the carrot rhizosphere. The comparative soils (excluding an uncultivated bulk soil) sampled in these studies limit the findings from conveying the true picture of core microbiomes specifically associated with carrot rhizosphere. This lack of detailed data creates a knowledge gap about how crop rotation system drives the diversity and structure of the rhizo-microbiome community in carrots, which is crucial for developing strategies to optimize soil health and crop yield. The current study aims to fill this gap by comparing the taxonomic composition and structural diversity of microbiomes in the rhizosphere of carrots grown under a monoculture system with those grown after soybean cultivation, using bulk soil from an uncultivated land as control.
The study stems from the hypotheses that the microbial diversity in soybean-precedent carrot rhizosphere will be higher than in monoculture, due to increased nutrients and varying organic residue such as root exudates from precedent soybean in the former20,21. It is also hypothesized that certain taxa with potential plant benefits (like organic matter decomposition, phosphate solubilization, disease suppression, and nitrogen fixation) resulting from the preceded crop22, will be more abundant in the soybean-precedent carrot rhizosphere. To explore the hypotheses, the study sought answers to three fundamental questions: (1) What are the differences in the microbial community structure viz.; taxonomic composition and distribution at the genus, family, and phylum levels between monoculture and soybean-precedent carrot rhizosphere, and a distant bulk soil?; (2) Are there notable differences in the occurrence of specific groups of microbes within and across the three sampling points?, and (3) Are soil properties and microbial community composition across the three sampling points correlated in any way? The study, therefore, employs Next Generation Sequencing, specifically the shotgun metagenomics approach, to provide robust information on the core rhizo-microbiome associated with a carrot grown under monoculture and soybean-precedent cultivation. By establishing this baseline data, the study not only seeks to provide insight into the distinction of the core rhizo-microbiome of carrots, but it will also uncover how cropping systems drive the microbial community with implications for sustainable agricultural practice and food safety.
Materials and methods
Study site and soil sampling
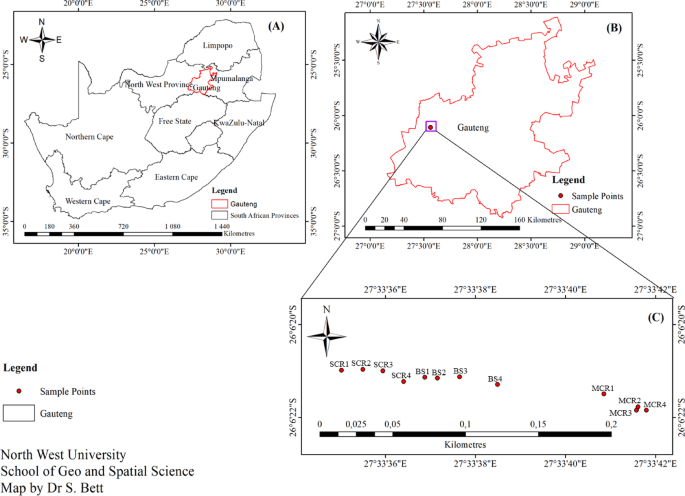
A total of twelve (12) samples consisting of 4 replicates each of rhizosphere soil from monoculture carrot (MCR), bulk soil from uncultivated land (BS), and rhizosphere soil from soybean-precedent carrot (SCR), were collected in Gauteng province of South Africa. The two carrot farmlands separately lying on a major commercial farm (26° 05′ 41.3″ S 27° 38′ 03.3″ E) at Tarlton, were selected for this study basically because of the distinct cropping histories (cropping systems) carried out on each with time. Both farmlands have been used mainly for organic farming practices for about eighteen (18) years using particular biofertilizers, Terramax and SoluPhos, and organic matter-based fertilizer (Humesoil). Also, the two farmlands were planted with the same carrot variety (Flamenco variety) during sample collection for this study. The monoculture carrot (MCR) farm has a history of continuous cropping, planting only carrots every season for over ten (10) years. The soybean-precedent carrot (SCR) farm has consistently been used for crop-rotation farming with soybean-carrot sequence history, i.e., carrot grown after soybean cultivation for about ten (10) years. The bulk soil (BS) from uncultivated (about 10 m from planting sites) was sampled for comparative purposes, to compare rhizosphere microbiome from the carrot farmlands with non-agricultural soil, serving as a reference point for existing microbial structure in soil without agricultural activity23. This was done to avoid the limitation of extrapolating general conclusions from the findings. The location of the sampling sites, Tarlton, is a suburb of Randfontein municipality in the West Rand District of Gauteng Province (Fig. 1), approximately 40 km southwest of Johannesburg. Randfontein consists of a mix of gently rolling hills and flat lands, with some elevated parts, situated on the Witwatersrand region known for its gold mining history. However, the study area has diversified into agriculture over the years, with a significant record of farming activities, including livestock farming and crop cultivation. This area experiences a climate with a mean temperature range of 15 °C to 35 °C and 0 °C to 20 °C in summer and winter, respectively. Precipitation in this area mostly occurs within the summer period (October-March), making an annual average range of 600–700 mm24.
About 600 g each of four (4) replicate soil samples were collected from SCR (26° 06′ 21.0″ S 27° 33′ 35.5″ E), BS (26° 06′ 21.1″ S 27° 33′ 36.9″ E), and MCR (26° 06′ 21.8″ S 27° 33′ 41.8″ E) separately. Rhizosphere soil sampling was done at the root development stage of carrots, 50 DAS (days after sowing), to capture the utmost microbial variability25. Using a pre-sterilized hand trowel to uproot the plant, loose soil on the taproot was gently shaken off, and the attached soil was brushed up with a sterile soft-bristled brush into a labelled sample bag. Rhizosphere soil from five random carrot plants was brushed up and pooled to make a composite sample per replicate. Bulk soil, five random points pooled as one sample, was collected within the upper layer (0–5 cm depth), because microbial composition in the region is relatively dense26. The samples were immediately transported in a cooler box to the Laboratory27. A part (500 g) of each sample was kept in the refrigerator at 4 °C while the other part was stored in the freezer at − 80 °C28.

Map showing the sampling sites and points with GPS coordinates. SCR, Rhizosphere soils from soybean-precedent carrot; BS, Bulk soils from uncultivated land; MCR, Rhizosphere soils from monoculture carrot.
Soil physicochemical properties analysis
Standard techniques were used to analyze the physicochemical properties of each soil sample collected from the carrot rhizospheres and comparable bulk soil. Firstly, each of the samples was well mixed, ground, and sieved (with a 2 mm sieve) after being air-dried to exclude wood, rubble, solid materials, and external particles29. The texture classification using a hydrometer was carried out to classify the soil according to the United States Department of Agriculture (USDA) particle sizes; 0.05 –0.002 mm for silt; 2.0–0.05 mm for sand; and < 0.002 mm for clay, as described by Enagbonma, Aremu30. Using a pH meter, the soil pH at a 1:2.5 ratio of soil to 1 M potassium chloride (KCl) was determined. The phosphorus contents of the soils were estimated by the spectrophotometric technique following the Bray 1 test, while the method of Walkley Black was adopted to analyze the organic carbon concentration31. Total nitrogen content including ammonium (NH4+) and nitrite (NO3−) was determined by a colorimetric method after extracting with 0.1 N potassium sulfate (K2SO4) method. Exchangeable sodium (Na), calcium (Ca), potassium (K), and magnesium (Mg) extracted (at pH 7) with a 1:10 ratio of soil to 1 M Ammonium acetate, were estimated with atomic absorption spectrophotometric (AAS) technique32.
Extraction of metagenomic DNA and sequencing
The metagenomic DNA molecules in the soil samples (0.25 g) were extracted following a stepwise protocol described by the manufacturer (Qiagen, Germany) of Dneasy® Power soil® DNA Isolation kit. The concentration and quality of the Extracted DNAs were examined with a Nanophotometer® NP80 Touch (Implen, Germany). All DNAs were appropriately labeled and stored at − 80 °C before sequencing. Using shotgun sequencing, a metagenomic dataset on each DNA sample was generated at the Novogene facility (NovogeneAIT Genomics Singapore Pte Ltd, Singapore). Briefly, the QC of the DNA samples including sample quantitation, integrity, and purity was further assessed at Novogene using a Qubit fluorometer, Agilent fluorometer, and Agarose Gel Electrophoresis Quantitation respectively. Using 1 µg of each sample, the DNA molecules were fragmented into segments (~ 350 bp) with a Covaris ultrasonic disruptor (Covaris, MA, USA), and a metagenomic library was constructed through adapters ligation, end repair, A-Tails addition, purification, and PCR amplification33. A QC check on the integrity of the construct and the size of the inserted fragment was done using the Advanced Analytical Technologies Incorporated (AATI) analysis34. The library was quantified with a Q-PCR for appropriate concentration (> 3 nM) and quality35. Afterward, appropriate libraries were sequenced with the Illumina X Plus (PE 150) system (NovogeneAIT Genomics Singapore Pte Ltd, Singapore).
Sequencing data analysis
The raw sequences (Fastq format) obtained from the Illumina sequencing platform were assessed for data quality, including the absence of external or artifact sequences, ambiguous sequences, and lengthy sequences (with a mean deviation of > 2)29. Poor or low-quality reads and adapters in the raw data were trimmed and filtered using the Fastp (v0.23.4)36, while contaminations from the host genomic DNA were removed with a Bowtie2 (v2.5.4) alignment37. Using the parameter settings for assembling, –presets meta-large (–end-to-end, –sensitive, -I 200, -X 400), Megahit (v1.1.2) was used to assemble the clean data38. Also, the MetaGeneMark (v2.10) platform was employed to determine the open reading frame (ORF) for each scaftig (> = 500 bp) using the default parameters39, and sequences with < 100 nt length were removed. The initial gene catalogue was obtained by demultiplexing the data with CD-Hit (v4.6.8), using -c 0.95, -G 0, -aS 0.9, − 1, -d 0 as parameter settings, as described by Lian, Huettel40. The number of genes that aligned with the initial gene catalogue of the clean data was determined with Bowtie2 (v2.5.4) before removing genes with < = 2 reads to achieve the unigene sequences. The abundance of the unigene in each sample was determined with the formula below:
$$:{G}_{k}=frac{{r}_{k}}{{L}_{k}}:.::frac{1}{{sum:}_{i=1}^{n}:frac{ri}{Li}}$$
where Gk is the abundance of unigene in the sample; r is the number of the gene reads on alignment; L is the length of the gene; and i is the initial value.
Taxonomic classification was analyzed by aligning the unigene sequences from each sample using DIAMOND (v2.1.9) with the blastp algorithm (parameter setting: 1e−5). The Micro_NR comprising sequences of all microbial domains (archaea, bacteria, virus, and fungi) was used as the corresponding database, and sequences with e-value 41. The eggNOG database was adopted for the functional category alignment, following the same sequence parameter in DIAMOND41. The algorithm of the Lowest Common Ancestor (LCA) in MEGAN (v6.8.20) was used to annotate each unigene (microbial species) sequence, which inferred the abundance of each microbial taxa (from kingdom to species level) and functional category in the soils42. Using at least 80% prevalence threshold in all the samples and a minimum of 0.01% relative abundance cut-off, on the R microbiome package (core_members() function), the core microbiome was determined across the treatments using the formula:
$$:text{P}text{r}text{e}text{v}text{a}text{l}text{e}text{n}text{c}text{e}:left(text{P}right)=:frac{Number:of:samples:with:taxon:detected:}{Total:number:of:samples}times:100$$
Statistical analysis
Using Duncan’s multiple range (post hoc) test, the difference in means from one-way analysis of variance (ANOVA) of a triplicate result of each soil physicochemical properties was computed for the level of significance p < 0.0543. The microbial community composition in the rhizospheric soil samples (MCR and SCR) and the comparable bulk soil (BS) was determined based on their average relative abundance in the four replicated samples from each sampling point. The relative composition of different taxa between the soil samples was determined with an abundance clustering heatmap using the R heatmap package with a z-score. Distribution differences of the microbiome (at the phyla level) between MCR, BS, and SCR were assessed with a principal component analysis (PCA) using R (ade4 package). The beta diversity of the microbiome between the points was estimated with the principal coordinate analysis (PCoA) based on Bray-curtis dissimilarity matrices of microbial taxa relative abundance44. Using the one-way analysis of similarity (ANOSIM) (R vegan package) with 999 permutations, the alpha diversity of the microbiome in individual sample groups (MCR, BS, and SCR) was determined by Shannon, Simpson (1 – D), and Chao1 diversity indexes calculated with appropriate formulas:
$$text{Shannon diversity index} (H^{prime}): :{H}^{{prime:}}=-{sum:}_{i=1}^{S}frac{ni}{N}text{ln}frac{ni}{N}$$
where (:S) is the total number of species observed; (:ni) is the number of individuals of species; (:N) is the total number of individuals in each sample. The higher the H′ values the higher the diversity.
$$text{Simpson diversity index} (1- D): :D=:{sum:}_{i=1}^{S}frac{ni:left(ni-1right)}{N:left(N-1right)}$$
where (:S) is the total number of species observed; (:ni) is the number of individuals of species i; (:N) is the total number of individuals in each sample. Index was expressed as 1 – D, where values close to 1 indicate higher diversity.
$$text{Chao1 diversity index}: :{text{C}text{h}text{a}text{o}1=S}_{obs}+:frac{{F}_{1}^{2}}{{2F}_{2}}$$
where (:S) is the total number of species observed; F1 is the number of species represented by a single individual; F2 is the number of species represented by two individuals.
The impact of edaphic factors on the microbiome distribution within sample groups was assessed by the redundancy analysis (RDA) using the soil properties in Table 1, with 999 Monte Carlo permutations for the significance test. CANOCO 5 (Microcomputer Power, Ithaca, NY, USA) was used to construct the RDA plot, and the correlation was determined with the Pearson’s correlation matrix generated with PAST v.4.1045. The quality sequences for the metagenomic-assembled genome (MAG) associated with soil samples from each of the sampling points; monoculture carrot rhizosphere (MCR), bulk soil from an uncultivated land (BS), and soybean-precedent carrot rhizosphere (SCR) are available at the NCBI SRA database under bioproject PRJNA1170280, PRJNA1170282, and PRJNA1174177 respectively.
Results
Physicochemical properties of monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil
The result of the physicochemical analysis on the soil samples depicted in Table 1 shows that the three sampling sites (MCR, BS, and SCR) consist of soil parameters at varying concentrations. The values of the parameters in all replicates before computing the averages are contained in (Table S1). The calcium (70.05 ± 1.28%) and moisture content (5.34 ± 0.00%) recorded for the rhizosphere soil of soybean-precedent carrot (SCR) are significantly higher than those (68.05 ± 0.44 and 4.33 ± 0.00% respectively) of monoculture carrot. The pH value, potassium, magnesium, sodium, Total N, NNH4, N-NO3 contents, clay, and sand proportion are not significantly different (p < 0.05) across all soil samples. However, the rhizosphere soil of soybean-precedent carrot (SCR) exhibits the highest percentage mean of sand (69.00 ± 3.92%) and clay (20.00 ± 3.16%) compared to monoculture carrot (MCR) and bulk soil (BS). Although the differences are not statistically significant, phosphorus content is the highest (216.25 ± 8.54 mg/kg) in BS followed by SCR (197.00 ± 10.46 mg/kg), while MCR recorded the least (190.00 ± 14.72 mg/kg). The variation in the mean values of the extractable macronutrients; K, Ca, Mg, and Na, recorded for all sampling points is not statistically significant.
Information on sequence data obtained from monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil
The average raw reads of the sequence data from each sampling point; MCR, BS, and SCR, contain 10.56, 10.66, and 11.58 gigabases respectively. After quality control, the clean base in the sequence reads at 0.1% error rate are 10.35, 10.41, and 11.32 with 93.86%, 93.70%, and 93.51% quality scores correspondingly. The percentage guanine-cytosine (% G-C) contents of the sequences are 66.24%, 65.82%, and 65.60% for MCR, BS, and SCR respectively. Also, the total length of sequence data generated from the soil samples of carrot rhizospheres and a distant bulk soil differs among the sampling points (MCR, BS, and SCR) with different scaftigs (Table S2). Generally, the sequence data consists mainly of bacterial sequences averaging 93.45% across the sampling points. In the SCR soil, bacterial community dominates, making up 93.69% of the sequence, followed by archaeal (1.38%), eukaryotic (0.02) %, and viral (0.01%), while other sequences (4.90%) remain unclassified. The sequences for BS and MCR contain 93.49 and 93.17% bacterial, 1.47 and 1.76% archaeal, 0.02 and 0.01% viral sequence with 5.01 and 5.04% remaining unclassified respectively. Only 0.01% of the entire metagenome in each of the two soil samples (BS and MCR) is for eukaryotes, separately (Table S3).
Structural composition of the core microbiome in monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil
In total, 197 microbial phyla belonging to the bacterial and viral kingdoms, as well as the domains archaea and eukaryota (consisting of fungi, plantae, and protista), are detected in the soil samples with varying abundances (Table S4). However, after filtering at ≥ 80% prevalence threshold with a minimum of 0.01% relative abundance cut-off, 35 core microbial phyla are recorded across the three sampling points at varying abundances. Three (8.57%) of the core microbiome are phylum; Nitrososphaerota, Euryarchaeota, and Thermoproteota belonging to the archaea domain, while only one viral phylum—Uroviricota is recorded. The soil sample from each sampling point is dominated by varying phyla belonging to the bacterial kingdom making up 88.57% of the microbial community. Most (77.42%) of the bacterial phyla are relatively enriched in the soybean-precedent carrot rhizosphere (SCR) compared to the bulk soil sample BS (6.45%) and the monoculture carrot rhizosphere MCR (12.90%). The phylum Actinomycetota recorded the highest abundance within each soil sample across the three sampling points (Table S5), however, the phylum was significantly enriched in MCR (Fig. 2). The Pseudomonadota abundance (14.69%) is significantly higher in SCR (at p < 0.05) than in BS (13.30%) and MCR (11.05%). The abundances of Chloroflexota and Spirochaetota in SCR (7.45% and 0.25%) and BS (7.35% and 6.40%) are higher than in MCR (6.40% and 0.16%), respectively. The abundances of Acidobacteriota, Planctomycetota, Myxococcota, Bacteriodota, Thermodesulfobacteriota, Armatimonadota, Rhodothermota, Calditrichota, Ignavibacteriota, and other five Candidatus (C. Eisenbacteria, C. Latescibacteriota, C. Zixiibacteriota, C. Aminicenantes) are not significantly different between SCR and BS but are higher in SCR than in MCR with significant differences (at p < 0.05) (Table S6).
The PCA result in Fig. 3 depicts the distribution pattern of microbiomes (phylum level) in the rhizospheres of monoculture (MCR) and soybean-precedent carrot (SCR) and a distant bulk soils (BS) with 69.13% and 30.86% variance in both axes 1 and 2 of the PCA respectively. The comparison of the distribution shows that the structural composition of the soybean-precedent carrot rhizosphere (SCR) forms a distinct cluster separated from BS and MCR. The result shows that Actinomycetota, Bacillota, and Pseudomonadota are significantly enriched in the monoculture carrot rhizosphere, distant bulk soil, and the soybean-precedent carrot rhizosphere respectively. Also, the microbial community in SCR is not only distinct but is significantly different from that in MCR (ANOSIM R > 0.5 and p < 0.05) (Fig. S1). Conversely, the difference between the community structure of BS and MCR sampling points is weak and not statistically significant (with ANOSIM R < 0.5 and p = 0.05) (Fig. S1).

Heatmap of relative abundance of the microbiome (phylum level) in monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil. The color saturation gradient represents the scale bar (with z-score) for the transformed relative abundance of each group. MCR, Rhizosphere soils from monoculture carrots; BS, Bulk soils from uncultivated land; and SCR, Rhizosphere soils from soybean-precedent carrots.

PCA plot on relative distribution of different phyla between monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil. MCR, Rhizosphere soils from monoculture carrots; BS, Bulk soils from uncultivated land; and SCR, Rhizosphere soils from soybean-precedent carrots.
At the family level, 48.57% of the total number (35) of microbial community encountered in this work are enriched in the soybean-precedent carrot rhizosphere (SCR) compared to the bulk soil (BS) and monoculture carrot rhizosphere (MCR) enriched with 37.14% and 14.29% respectively (Fig. 4). The family Nocardioidaceae is enriched, with the highest abundance, across the sampling points; MCR (15.15%), BS (13.22%), and SCR (12.07%). The family Nitrobacteraceae is significantly more abundant in SCR (with 0.92%) than in BS (0.76%) and MCR (0.57%) (at p < 0.05) (Table S5). Hyphomicrobiaceae (1.32%), Gaiellaceae (1.36%), Microbacteriaceae (0.92%), Gemmataceae (0.67%), Vicinamibacteraceae (0.43%), and Phyllobacteriaceae (0.38%) are more abundant in SCR than in MCR where they have 0.71, 1.11, 0.61, 0.48, 0.30, and 0.26% abundances respectively. However, the differences between their abundances in SCR and BS (1.07, 1.19, 0.78, 0.59, 0.39, and 0.34% respectively) are not statistically significant. Although, its relative abundances in SCR (0.24) and BS (0.20) are not significantly different, the family Pirellulaceae are notably not encountered in MCR.
In contrast, the highest number of bacterial genera (51.43%) in this work are enriched in MCR, whereas, only one genus—Desertimonas (2.86%) dominates BS, compared to the soybean-precedent carrot rhizospheric (SCR) dominated by 45.71% of the genera (Fig. 4). Among other genera, Bradyrhizobium, Hyphomicrobium, and Agromyces are enriched in soybean-precedent carrot rhizosphere (SCR) with abundances (0.78, 0.58, and 0.47%) that are higher than those in BS (0.63, 0.49, and 0.36%) and MCR (0.48, 0.36, and 0.23%) correspondingly. Five genera—Gaiella, Aeromicrobium, Luteitalea, Mesorhizobium, and Rhodoplanes recorded significantly higher abundances (1.19, 0.53, 0.43, 0.30, and 0.29%) in SCR compared to those (0.97, 0.39, 0.30, 0.21, and 0.15%) in MCR (p < 0.05), but not different in BS with 1.05, 0.48, 0.39, 0.27, and 0.26% abundance respectively (Table S5). Notably, the genus Rhodococcus occurs in SCR only, while Methyloceanibacter is encountered in SCR and BS with the highest abundance in SCR, but not in MCR.

Heatmap of relative abundance of the microbiome at (A) family and (B) genus level in monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil. The color saturation gradient represents the scale bar (with z-score) for the transformed relative abundance of each group. MCR, Rhizosphere soils from monoculture carrots; BS, Bulk soils from uncultivated land; and CR, Rhizosphere soils from soybean-precedent carrots.
Microbial diversity in monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil
The alpha diversity of the microbiome composition (at the genus level) in monoculture (MCR) and soybean-precedent carrot rhizosphere (SCR) and those of the distant bulk soil shows different range in the microbial community across sampling fields. Table 2 shows that the Simpson indexes for all the sampling points are significantly different at a p-value of 0.0388, with SCR having the highest index value (0.93 ± 0.01), followed by BS (0.92 ± 0.01) and MCR (0.90 ± 0.02). Although post hoc shows no significant pairwise difference, there is higher diversity in SCR with a Shannon index value of 6.18 ± 0.18 compared to BS (6.07 ± 0.13) and MCR (5.76 ± 0.21), showing significant differences in the overall ANOVA (p-value of 0.0497). Conversely, with differences in post hoc multiple comparison, BS recorded highest Chao1 index value (3407.03 ± 40.20), followed by SCR (3392.65 ± 51.28) and MCR having the least value (3345.15 ± 111.22). However, the overall ANOVA shows no significant difference in the Chao1 indexes of the microbiome diversity across sampling points (at p-value > 0.05).
Comparing the result of any pair of the replicate samples from each sampling point to another (MCR, BS, and SCR) using the PCoA, there is a distinct clustering in the community structure of the microbiome at the genus level (Fig. 5). The principal coordinate constructed in axis 1 (PCo1) captures most (69.13%) of the variation in the microbial diversity across the three sampling points. Based on ANOSIM, the diversity in community composition between SCR and MCR is statistically significant with p-value = 0.042, with a higher similarity level within each sampling point than between the two (R-value = 0.563). The microbial diversity within each of SCR and BS is higher than when compared between the both, but the variation is not statistically significant (p-value = 0.597; R-value = − 0.094). However, the difference in the diversity between BS and MCR lacks statistical significance (ANOSIM, p-value = 0.101; R-value = 0.292) (Fig. S2).

PCoA plot for beta diversity of the microbiome in monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil based on Bray–Curtis dissimilarity. The relative distance between points indicates the similarity or dissimilarity degree, where each point represents a soil sample, and the percentage on the axis explains the proportion of variance in that dimension. MCR, Rhizosphere soils from monoculture carrots; BS, Bulk soils from uncultivated land; SCR, Rhizosphere soils from soybean-precedent carrots.
Metabolic functions associated with monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil
Considering the eggNOG at the lowest hierarchical level (3), the metabolic pathways associated with plant-beneficial functionalities across the sampling fields are depicted in Fig. 6. Of the 21 pathways for different metabolic processes, 66.7% have the highest relative abundance in SCR, followed by MCR (19.0%), while BS is more enriched with only 14.3%. The relative abundance of 47.6% of the functions, including central carbon metabolism, ATP synthesis, bacteriocin production, resistance mechanisms, modifying enzyme, dioxygenase, nitrite reductase, lipolytic protein, carbon-nitrogen hydrolase, and carbon-phosphorus lyase activities, is significantly enriched in SCR (at p < 0.05) (Table S6). Except for ubiquitin-mediated proteolysis and nitrite oxidoreductase activities, which are significantly abundant in MCR and BS, respectively, there is no significant difference at p < 0.05 in the abundance of other functions across the sampling fields.

Plant-beneficial metabolic functions associated with monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil. SCR, Rhizosphere soils from soybean-precedent carrots; BS, Bulk soils from uncultivated land; MCR, Rhizosphere soils from monoculture carrots.
Impact of edaphic factors on microbial community in monoculture and soybean-precedent carrot rhizosphere and a distant bulk soil
Using the soil properties in Table 1, their effect on the microbial composition and diversity at the genus level in the monoculture (MCR) and soybean-precedent carrot rhizosphere (SCR) and those of the distant bulk soil are represented with a redundancy analysis (RDA) plot (Fig. 7). The plot shows that most (84.29%) of the variability is captured in Axis 1, and the relationship patterns between soil properties and microbial taxa are represented in Axis 2. The soil properties show varying influences on the bacterial community composition indicated by the Pearson correlation coefficients (r-values) at varying significance (p < 0.05), expressed in Fig. S3 and the corresponding vector length and direction (on RDA plot). The arrow of pH, for instance, pointing toward Desertimonas suggests a positive relationship, which is also indicated by a strong positive correlation coefficient (r = 0.98) in the matrix. The distribution of different genera in the direction or opposite of specific soil properties is associated with the soil condition at varying significance (at p < 0.05) (Fig. S3). Sand texture significantly (p < 0.05) influences the composition of different genera, showing a strong positive correlation with Sphingomonas (r-value = 1) and a strong negative correlation with Solirubrobacter, Knoelli, and Nitrososphaera (r = − 0.99 each). Organic carbon positively correlates with Norcardioides, Rubrobacter, Conexibacter, and Baekduia (r = 0.99 each); and negatively correlates with Microbacterium, Luteitalea, Mesorhizobium, and Rhodoplane (r = − 0.99 each) at a significant level (p < 0.05). Significantly, Total N positively correlates with Lamia and Intrasporangium with strong coefficients (r = 0.99 each); and negatively correlates with Gaiella (r = − 0.99 each). However, the compositions of some genera, including Streptomyces, Mycolicibacterium, Arthrobacter, and Rhodococcus, are not significantly affected by any soil property (p < 0.05). Summary of the RDA (Table S7) supports the results showing that silt, total nitrogen, organic carbon, and sand texture contributed 84.2%, 83.2%, 81.0%, and 75.8% impact to the composition and diversity of microbiome across the treatments, but none was statistically significant.

Redundancy analysis (RDA) plot showing the impact of soil parameters on the microbial composition in the carrot rhizospheres and bulk soil. The length of each vector depicts the strength of the influence by the soil parameter; *S-Value = Sum of extractable K, Ca, Mg, and Na (c.mol(+)/kg)(me%).
Discussion
Shotgun metagenomics approach was employed in this study to provide insight into how cropping systems affect the diversity and community structure of the core microbiomes associated with the carrot rhizosphere. The study compared the abundance and diversity of the rhizo-microbiome at various taxonomic levels in the rhizosphere of monoculture carrots, with what is obtainable in those grown under a crop rotation system following soybean cultivation, and distant bulk soil from uncultivated land. The emerging and efficient high throughput sequencing technique was used to generate robust data, from which the three hypothetical questions relating to the study were addressed.
The microbial community abundances at phylum, family, and genus in monoculture carrot rhizosphere (MCR), distant bulk soil from an uncultivated land (BS), and soybean-precedent carrot rhizosphere (SCR) shown in heatmaps (Figs. 2 and 4) were constructed to address the question (1) regarding differences in taxonomic composition. Complementarily, the result of alpha diversity (genus level) and the distribution pattern at the phylum level (in the PCA plot) of the microbiome explained the difference in their microbial community structure. Generally, the soil samples obtained across the sampling points were dominated by phyla of the bacterial domain, had three archaeal phyla: Nitrososphaerota, Euryarchaeota, and Thermoproteota, and contained only one viral phylum, Uroviricota. These findings support the report of Bagci, Suzuki1, who also recorded a dominance (at the phylum level) of the bacterial taxa with a few archaea in the rhizosphere of carrot plants. The archaeal and viral phyla in this study have been reported to be found inhabiting different plant rhizospheres in previous studies46,47,48,49. The dominance of bacteria (88.57%), at the phylum level, in this study indicates the crucial role of the bacterial community in the soil ecosystem50. The archaeal phyla were dominant in the monoculture carrot rhizosphere (MCR), while the only viral phylum was enriched in the bulk soil (BS), suggesting that the bulk and monoculture carrot rhizosphere soils were more suitable for these taxa. On the other hand, most of the bacterial phyla (77.42%), are enriched in the soybean-precedent carrot rhizosphere (SCR), indicating the suitability of soil used for soybean-carrot rotational cultivation for bacterial proliferation, owing to its improved nutrient and bio-character, resulting from complementary and mutually beneficial effects of the crops51. The indication that soybean-precedent carrot rhizosphere fosters a more abundant bacterial community compared to the monoculture system in this study aligns with previous reports on crop rotation systems that led to significant shifts in rhizobacterial communities, which improve soil health and plant yield52,53.
Consequently, the PCA result, which captured most (92.01%) of the variability of the data, clearly shows a disparity in the distribution pattern between the soybean-precedent carrot rhizosphere microbiome and those of monoculture carrot. The distribution of the microorganisms (phylum level) in the soybean-precedent carrot rhizosphere forms a distinct cluster significantly different from those in the monoculture carrot rhizosphere, indicating a unique community structure in the former. This could be attributed to the potential of soybean, a leguminous crop that preceded carrot planting, to fix nitrogen, increase carbon pools, and boost nutrient availability15,53,54, enhancing abundance of some specific microbiome in the niche. Whereas, the slight overlap in the community distribution in monoculture carrot rhizosphere (MCR) and bulk soil (BS) underscores some level of similarity between the two sampling points. The findings are inconsistent with the work of Zhao, Li55 which reported significant differences in the microbial structure between bulk and rhizosphere soils of two halophytes. This could be a result of depletion in available nutrients and soil fertility associated with long-term monoculture systems56. Furthermore, bacteria at the family level dominated the rhizosphere soil of soybean-precedent carrots with relatively high abundance compared to monoculture carrots. This could be attributed to the uniqueness of the microhabitat created by plant roots (through their exudates) and other environmental features that serve as enhancing factors that promote a diverse microbial community at the higher domain with distinctive functions57,58. The dominance of these taxa in the respective locations correlates with the relative complexity of each and the localization of the microbial community15,59,60,61. For instance, some archaea might not withstand the dynamics associated with root exudates and other biotic interactions in the rhizosphere62. Also, the viral abundance in the bulk soil emphasizes the influence of viruses in organizing the microbial community, possibly through predation on the bacterial population63.
The wide range covered by the vector lengths for monoculture carrot rhizosphere (MCR) in the PCoA plot can also be related to the high microbial community population associated with the niche. However, the few genera that dominated the soybean-precedent carrot rhizosphere (SCR) exhibited significantly higher relative abundances than those of monoculture carrot. This scenario highlights the dominance of unique communities based on selection pressure in the plant rhizosphere64. Additionally, based on a distance matrix (Bray–Curtis dissimilarity), the diversity analysis (beta) further establishes differences in microbial diversity at the genus level across the sampling points. The microbial community structure in the soybean-precedent carrot rhizosphere is unique compared to those of monoculture and comparable bulk soil, as shown by the distanced cluster on the PCoA plot. The similarity analysis (ANOSIM) supports this with a significant difference between SCR and MCR (R-value = 0.5625; p-value = 0.042), suggesting that SCR and MCR consist of a markedly different microbial community. Conversely, pairing BS and MCR showed a significant similarity (R-value = 0.29167; p-value = 0.101), indicating that the microbial community structure in the monoculture carrot rhizosphere shares some similarities with the bulk soil from uncultivated land65. This justifies the idea that agricultural management practices, particularly cropping systems including crop rotation and monoculture determine the structure of the rhizosphere microbiome, making it different from that of bulk soil66,67,68.
The second question was addressed by differences in the relative dominance and occurrence or absence of certain taxa at different taxonomic levels across the sampling points (SCR, BS, and MCR) inferred from the heatmap (Figs. 2 and 4). Also, alpha diversity, which explains variability within an ecosystem, was used to assess the comparative diversity based on the richness, evenness, and abundance of the microbial community within the soil sample. The result showed that the community structure of the rhizosphere of soybean-precedent carrot (SCR) is majorly dominated by numerous plant-soil beneficial microorganisms across different taxa with notably high relative abundances. Prominent among the dominating genera in this study are Bradyrhizobium, Mesorhizobium, Rhodococcus, Gaiella, and Hyphomicrobium, with growing pieces of evidence on their plant growth-promoting capabilities69,70,71,72,73. All dominating microorganisms across the taxonomic levels recorded in this study for soybean-precedent carrot (SCR) have been reported in other research reports as dominant in different regions of plant roots and agricultural soils, which offer beneficial activities to plants and the soil74,75,76. Evidently, like other previous studies, the present study also recorded arrays of metabolic functions (Fig. 6 and Table S6) significant to sustainable agriculture, with varying abundances across the sampling points. The enrichment of Actinomycetota, a phylum known for organic matter decomposition and nutrient cycling that enhances soil health, across all sampling points underscores the crucial role of the phylum in the soil ecosystem77. Several other phyla with different soil functions and plant benefits were enriched in the soybean-precedent carrot rhizosphere (SCR); for example, Pseudomonadota encountered in this study has been associated with essential metabolic activities of agricultural significance, including biomass and energy metabolism, and antibiosis activity at plant-root regions78. Also, Chloroflexota is known to play a key role in nutrient cycling through complex compound degradation79. The significant abundance of metabolic functions such as central carbon metabolism, ATP synthesis, bacteriocin production, resistance mechanisms, modifying enzyme, dioxygenase, nitrite reductase, lipolytic protein, carbon-nitrogen hydrolase, and carbon-phosphorus lyase activities in the SCR, suggests relatively enhanced plant–microbial interactions, that benefit carrot preceded by soybean, at the rhizosphere80. Studies indicate that central carbon metabolism, ATP synthesis, and carbon fixation, that is relatively enriched in the treatment, are metabolic pathways fundamental to microbial growth and biomass accumulation, directing nutrient availability and energy flow within the rhizosphere80,81. Bacteriocin production, resistance mechanisms, and modifying enzyme activity pathways contribute to microbial interactions and survival strategies, suppressing and competing with plant pathogens, in the rhizosphere soil82. Although their relative abundances are not significantly different across the fields (Table S6), secondary metabolite (antibiotics) synthesis and specific resistance mechanisms like efflux pump and polyketide synthesis activity that are more enriched in the rhizosphere, underscore the rhizo-microbiomes’ role in protecting soybean-precedent carrot against pathogens, through antimicrobial production83. Notably, an anaerobic group of bacteria belonging to Pirellulaceae family, which contributes significantly to nitrogen fixation through anaerobic ammonia-oxidation (Anammox)76, was found in the soybean-precedent carrot rhizosphere but not in monoculture carrot. This indicates that the carrot could have inherited this microbial group with anammox-mediated nitrogen cycling capability from the preceding soybean, to mitigate nitrogen loss84. This is evidenced by nitrite reductase activity, significant abundance, and nitrite oxidoreductase activity enrichment, two metabolic functions vital to nitrogen transformation under anaerobic conditions involving ammonium and nitrite conversion for plant use, particularly during nitrogen loss85. Similarly, an archaeal genus, Methyloceanibacter, which partakes in plant nutrient supply through organic and inorganic matter transformation81, was not encountered in monoculture carrots, indicating that rhizo-microbiomes of soybean-carrot rotation can contribute to more stable nutrient cycling compared with monoculture carrots. Also, with a variety of functional roles, including organic matter decomposition, known to them86,87, the comparatively large abundance of Hyphomicrobium and Rhodoplanes in the soybean-precedent carrot rhizosphere, corroborates the dominance of other soil and plant-benefiting microbial communities in this kind of soil treatment. Coupled with other less abundant enzymatic functions, including laccase, chitinase, and proteolysis activity present in this rotational treatment, the rhizosphere is significantly enriched with lipolytic protein activity (Table S6), which is essential for nutrient release and availability for plant uptake through organic decomposition88. More remarkable is the exclusive abundance of Rhodococcus in the former, suggesting that the crop rotation system, specifically the preceding soybean exudates, might have selectively selected them to play crucial roles in the carrot rhizosphere89. These genera are well-known for their biodegradability of a wide range of complex compounds, contributing to nutrient cycling, preserving soil health, and stimulating plant growth90,91. Deoxygenase activity, central to pathways involved in the breakdown of recalcitrant organic matter92, is among the metabolic functions recorded in the soybean-precedent carrot rhizosphere, with significant abundance. The abundance of other similar functions, including xenobiotic biodegradation and metabolism, laccases activity for complex plant polymer degradation93, and chitinase activity for chitin breakdown94, are not significantly different across the sampling fields, but are more enriched in rotational than monoculture treatment. However, the relative abundance of these metabolic functions explains the exclusive presence of complex compound degraders, Rhodococcus, in the soybean-precedent carrot rhizosphere, highlighting the better capacity of its specific microbiomes in recycling matter to enhance soil structure and fertility80. Jena, Maitra51 reported that residues from preceding soybean offer subsequent crops multifaceted capacity, including exuding organic acids, fixing soil nitrogen, shaping soil organic matter, and fostering beneficial soil microbes, in the rhizosphere. This justifies the significant abundance of nitrogen-fixing, soil-benefiting, and plant-enhancing legume-associated bacterial genera, Bradyrhizobium and Mesorhizobium, in the soybean-rotation carrot rhizosphere95. For instance, such carrots may benefit from increased nutrient availability from these two genera, Bradyrhizobium and Mesorhizobium, that are most probably inherited from their preceding legume crop53,96, explaining their higher abundance in the soybean-precedent treatment compared to monoculture carrots and the distant bulk soil97. These microbiomes’ relative abundance and integral roles in the rhizosphere of rotational carrot are further highlighted by the significant abundance of carbon-nitrogen hydrolase and carbon-phosphorus lyase activities98,99. Beyond organic matter degradation or anammox process, these enzymatic activities and that of thiosulfate sulfurtransferase enzyme, are important in supplying and cycling essential nutrients such as nitrogen, sulfur, and phosphorus, promoting plant growth and soil health in this kind of treatment99.
The claims of this study to address research question (2) are further supported by the alpha diversity assessment results with Shannon and Simpson indexes, indicating that the richness and evenness (p = 0.0497), and diversity (p = 0.0388) of the respective microbial community structures across the three locations are significantly different. This underscores the three sampling points as being evenly rich with significantly distinct microbial communities associated with diverse functions in the soil ecosystem. However, the rhizosphere of soybean-precedent carrot exhibited the highest species richness and evenness (based on the Shannon index) and diversity (based on the Simpson index), indicating a complex yet stable ecosystem. This is in line with a study by Jin, Lyu100 which found the bacterial richness and diversity in the rhizosphere of different vegetable rotations higher than continuous cropping substrates. It also corroborates the findings of Li, Li23 which showed that crop rotation-fallow systems significantly increase bacterial alpha diversity indices in paddy fields in a tillage year-dependent manner. However, this scenario does not correspond with the findings from a bioinformatics study on intercontinental sequence data of rhizospheric and bulk soil microbiomes by Ling, Wang81 who reported a reduction in the observed bacterial richness and Shannon diversity index in plant rhizosphere soil. This variation may be because the study did not cover the Apiaceae family to which carrot belongs. The deviation may speak of the uniqueness of the soybean-precedent carrot rhizosphere as shown by the contrasting result recorded for paddy soil—at the rhizosphere of rice cultivation in Ling, Wang81 study. Perhaps, the soybean-precedent carrot rhizosphere exhibits distinct biotic and abiotic properties, making it a suitable habitat for wider microbial taxa including aerobes and anaerobes. This further underlines the key role of heterogeneity in the rhizosphere under crop rotation, including residual nutrients, root exudates, and soil moisture, in shaping the diversity and community structure of microorganisms in the habitat81. Research has shown that crop rotation systems, particularly with soybeans, can significantly increase microbial composition and diversity associated with nutrient acquisition in plant and soil resilience to stressors, potentially improving soil fertility, plant health, and crop productivity15,20,52,68.
Lastly, to address question 3, the relationship between soil properties in Table 1 and microbial community (at genus level) structure was determined with a Redundancy Analysis (RDA) (Fig. 7). Estimated soil properties showed uniformity in the pH, potassium, magnesium, sodium, nitrogen sources (total N, N-NO3, N-NH4), clay, and sand values across all sampling points. However, the moisture and calcium contents of soybean-precedent carrot (SCR) rhizosphere were significantly higher compared to those of monoculture and distant bulk soils (MCR and BS) (at p < 0.05). These differences underline the rhizosphere of carrots grown after soybean cultivation (SCR) as a more conducive environment for plant and microbial proliferation due to water and nutrient retention properties101. Although not significantly different, the shortfall in organic carbon and nitrogen (N-NO3 and total nitrogen) content of the soybean-precedent carrot rhizosphere compared to the monoculture rhizosphere could be attributed to their use for metabolic activities by diverse microbial communities during intricate interaction with plants and other organisms in the niche102. Considering their interactions with the moisture content and soil texture in soybean-precedent carrot rhizosphere, the distribution pattern of other nutrients, such as organic matter, nitrogen, and phosphorus, underscores their significance in metabolisms of entire plant vigor, plant growth, and root development via nutrient acquisition, and those of microbial communities68,103. Furthermore, the influence of the soil parameters on the microbial community structure was established with the trends on the RDA plot (Fig. 7). The study showed ample interaction between the test parameters and the composition of microorganisms across all sampling points (SCR, BS, and MCR). Generally, the soil factors including phosphorus, potassium, and sodium availability, moisture content, and soil texture contributed to the structure and diversity of microbial community in this study. This aligns with the results of Li, Li23 who reported these soil properties to greatly influence the bacterial community structure under different land use patterns. This study found silt, organic carbon, nitrogen contents, and sand to be the key drivers of microbial community structure and diversity. The silt and sandy texture of the soil separately explains variance in microbial community structure indicated by the clustering of different genera around them, suggesting their key role in the community composition and distribution of soil microorganisms104. According to Xia, Zheng105, the finer-textured part of the soil, sand, fosters a higher microbial community, most likely due to increased nutrients and water films occupying the larger pores in the sand, serving a protective role to the community105. Additionally, the majority of the bacterial genera sparsely concentrate around different nutritional components of the soils, which underlines the impact of nutrient availability on microbial diversity in soil ecosystems106.
Conclusion
Using a shotgun sequencing metagenomics approach, this study has provided substantial insight into the overall community composition and diversity of the carrot rhizo-microbiome under two cropping systems—monoculture and crop rotation. It established a remarkably microbial composition and diversity in the rhizosphere from carrots grown after soybean cultivation (under crop rotation) compared to the monoculture carrot rhizosphere. Also, the study unveiled bacteria as the dominating group comprising the microbial community found in the carrot rhizosphere under the two cropping systems. Going by the significant diversity in microbial composition in the soybean-precedent carrot rhizosphere, the study underscores the dynamic interactions among the microbiome at the soil-root interface during crop rotation practice. Several microorganisms that have been previously related to soil health-benefiting and plant growth-promoting abilities, ranging from organic carbon degradation to nutrient cycling and nitrogen fixation, were embodied by soybean-precedent carrot rhizosphere. The study showed that the crop rotation system particularly with soybeans significantly promoted the microbial community diversity, potentially improving soil fertility, plant health, and crop productivity. The insights relating to the soybean-precedent carrot rhizosphere uncovered in this study could be exploited for biotechnological machinery to attain sustainable agricultural practice at this time of need. Notably, a significant number of unclassified groups of microorganisms were recorded in the study, presenting the chance to unravel novel species of agricultural and biotechnological applications. It is also imperative to deepen research on how these cropping systems drive the functionality of the carrot rhizosphere core microbiome (including the habitat itself) to further elucidate the hidden treasure of the microhabitat for potential benefit to mankind. Ultimately, this study could inform decision-making on cropping systems and soil management practices toward sustainable agriculture and food safety.
Data availability
The standard sequence data obtained in this work have been deposited in the NCBI Sequence Read Archive (SRA) under accession numbers PRJNA1170280, PRJNA1170282, and PRJNA1174177.
References
-
Bagci, I. et al. The application of organic matter temporarily shifts carrot prokaryotic communities in the endosphere but not in the rhizosphere. Microorganisms. 11 (10), 2377 (2023).
-
Qasim, M. Beneficial microbes in plant health: from biocontrol agent to plant growth promoting rhizobacteria. Int. J. Res. Adv. Agric. Sci. 2 (3), 39–49 (2023).
-
Trivedi, P. et al. Enabling sustainable agriculture through Understanding and enhancement of microbiomes. New Phytol. 230 (6), 2129–2147 (2021).
-
Tosi, M. et al. It takes three to tango: the importance of microbes, host plant, and soil management to elucidate manipulation strategies for the plant Microbiome. Can. J. Microbiol. 66 (7), 413–433 (2020).
-
Dlamini, S. P., Akanmu, A. O. & Babalola, O. O. Rhizospheric microorganisms: the gateway to a sustainable plant health. Front. Sustain. Food Syst. 6, 925802 (2022).
-
Yang, Y. et al. Effects of grazing intensity on diversity and composition of rhizosphere and non-rhizosphere microbial communities in a desert grassland. Ecol. Evol. 13 (7), e10300 (2023).
-
Kumar, A. et al. Phytobiomes: Harnessing the power of microbial communities for plant health. Microbiol. Res. J. Int. 34 (10), 1–19 (2024).
-
Zhao, M. et al. Rhizosphere regulation with cerium oxide nanomaterials promoted Carrot taproot thickening. Environ. Sci. Nano. 11 (8), 3359–3373 (2024).
-
Dang, K. et al. Root exudates of ginger induced by ralstonia solanacearum infection could inhibit bacterial wilt. J. Agric. Food Chem. 71 (4), 1957–1969 (2023).
-
Rachwał, K. et al. Is soil management system really important? Comparison of microbial community diversity and structure in soils managed under organic and conventional regimes with some view on soil properties. PLoS One. 16 (9), e0256969 (2021).
-
Anjani, G., Ayustaningwarno, F. & Eviana, R. Critical review on the immunomodulatory activities of carrot’s β-carotene and other bioactive compounds. J. Funct. Foods. 99, 105303 (2022).
-
Triviño, N. J. et al. Carrot genotypes differentially alter soil bacterial communities and decomposition of plant residue in soil. Plant. Soil. 486 (1), 587–606 (2023).
-
Paparella, A. et al. Challenges and opportunities in the sustainable improvement of carrot production. Plants. 13(15), 2092 (2024).
-
Shah, K. K. et al. Diversified crop rotation: an approach for sustainable agriculture production. Adv. Agric. 2021 (1), 8924087 (2021).
-
Fadiji, A. E., Kanu, J. O. & Babalola, O. O. Impact of cropping systems on the functional diversity of rhizosphere microbial communities associated with maize plant: a shotgun approach. Arch. Microbiol. 203 (6), 3605–3613 (2021).
-
Song, Q. et al. Overfertilization reduces tomato yield under long-term continuous cropping system via regulation of soil microbial community composition. Front. Microbiol. 13, 952021 (2022).
-
Yang, X. et al. Diversifying crop rotation increases food production, reduces net greenhouse gas emissions and improves soil health. Nat. Commun. 15 (1), 198 (2024).
-
Shi, X. et al. Insight into plant-microbe interactions in rhizosphere for sustainable agriculture in the new crops era. New. Crops. 1, 100004 (2024).
-
Kumar, A. & Dubey, A. Rhizosphere microbiome: engineering bacterial competitiveness for enhancing crop production. J. Adv. Res. 24, 337–352 (2020).
-
Zhou, Y. et al. Crop rotation and native microbiome inoculation restore soil capacity to suppress a root disease. Nat. Commun. 14 (1), 8126 (2023).
-
Agyekum, D. V. et al. Diversity and function of soybean rhizosphere Microbiome under nature farming. Front. Microbiol. 14, 1130969 (2023).
-
Kuerban, M. et al. Microbial soil legacies of crops under different water and nitrogen levels determine succeeding crop performance. Plant. Soil. 485 (1), 167–180 (2023).
-
Li, N. et al. Impacts of rotation-fallow practices on bacterial community structure in paddy fields. Microbiol. Spectr. 10 (4), e00227–e00222 (2022).
-
Dyson, L. L. A heavy rainfall sounding climatology over gauteng, South Africa, using self-organising maps. Clim. Dyn. 45 (11–12), 3051–3066 (2015).
-
Song, H. et al. Study on the mechanism of carotenoid production and accumulation in orange red carrot (Daucus carota L). Sci. Hort. 327, 112825 (2024).
-
Das, S. et al. Soil microbial biomass carbon stock and its relation with Climatic and other environmental factors in forest ecosystems: A review. Acta Ecol. Sin. 43 (6), 933–945 (2023).
-
Enagbonma, B. J. & Babalola, O. O. Metagenomics reveals the Microbiome multifunctionalities of environmental importance from termite mound soils. Bioinform. Biol. Insights. 17, 11779322231184025 (2023).
-
Wakung’oli, M. et al. Termite societies promote the taxonomic and functional diversity of archaeal communities in mound soils. Biology. 9 (6), 136 (2020).
-
Enagbonma, B. J., Ajilogba, C. F. & Babalola, O. O. Metagenomic profiling of bacterial diversity and community structure in termite mounds and surrounding soils. Arch. Microbiol. 202 (10), 2697–2709 (2020).
-
Enagbonma, B. J., Aremu, B. R. & Babalola, O. O. Profiling the functional diversity of termite mound soil bacteria as revealed by shotgun sequencing. Genes 10 (9), 637 (2019).
-
Handayani, E. P. & Isnaini, S. Analysis of chemical soil properties and social economic study of swampland rice productivity. Malays. J. Soil. Sci. 27, 186–195 (2023).
-
Hasana, H., Beyene, S. & Kifilu, A. Influence of phosphogypsum amendments on the performance of wheat (Triticum aestivum L.) and chemical properties of sodic soil of alage, central rift Valley of Ethiopia. J. Soil. Sci. Plant. Nutr. 23 (3), 3888–3903 (2023).
-
Zhu, B. et al. α-Pyrone mediates quorum sensing through the conservon system in nocardiopsis Sp. Microbiol. Res. 285, 127767 (2024).
-
Babalola, O. O., Adebayo, A. A. & Enagbonma, B. J. Shotgun metagenomics dataset of the core rhizo-microbiome of monoculture and soybean-precedent Carrot. BMC Genomic Data. 26, 26 (2025).
-
Song, J. et al. Interpretation of vaginal metagenomic characteristics in different types of vaginitis. Msystems. 9 (3), e01377–e01323 (2024).
-
Chen, S. Ultrafast one-pass FASTQ data preprocessing, quality control, and deduplication using Fastp. Imeta. 2 (2), e107 (2023).
-
Cheng, W. Y. et al. High sensitivity of shotgun metagenomic sequencing in colon tissue biopsy by host DNA depletion. Genom. Proteom. Bioinform. 21 (6), 1195–1205 (2023).
-
Deng, C. et al. Genome-centric metagenomics provides new insights into the microbial community and metabolic potential of landfill leachate microbiota. Sci. Total Environ. 816, 151635 (2022).
-
Li, C. et al. Expanded catalogue of metagenome-assembled genomes reveals resistome characteristics and athletic performance-associated microbes in horse. Microbiome 11 (1), 7 (2023).
-
Lian, Q. et al. A pan-genome of 69 Arabidopsis Thaliana accessions reveals a conserved genome structure throughout the global species range. Nat. Genet. 1–10 (2024).
-
Hu, B. et al. Gut microbiota facilitates adaptation of the plateau Zokor (Myospalax baileyi) to the plateau living environment. Front. Microbiol. 14, 1136845 (2023).
-
Hanson, N. W., Konwar, K. M. & Hallam, S. J. LCA*: an entropy-based measure for taxonomic assignment within assembled metagenomes. Bioinformatics. 32 (23), 3535–3542 (2016).
-
Adebayo, A. A. et al. Plant growth promoting potentials of novel phosphate-solubilizing bacteria isolated from rumen content of white Fulani cattle, Indigenous to Nigeria. Biologia. 78 (1), 201–215 (2023).
-
Joseph, B. & Babu, S. Effect of organic and chemical fertilizer on the diversity of rhizosphere and leaf microbial composition in sunflower plant. Curr. Microbiol. 81 (10), 331 (2024).
-
Enagbonma, B. J. & Babalola, O. O. Metagenomics shows that termite activities influence the diversity and composition of soil invertebrates in termite mound soils. Appl. Environ. Soil. Sci. 2022 (1), 7111775 (2022).
-
Fadiji, A. E., Kanu, J. O. & Babalola, O. O. Metagenomic profiling of rhizosphere microbial community structure and diversity associated with maize plant as affected by cropping systems. Int. Microbiol. 24 (3), 325–335 (2021).
-
Lee, U. J. et al. Ca. Nitrosocosmicus members are the dominant archaea associated with plant rhizospheres. mSphere. e00821-24 (2024).
-
Zhu, J. et al. Non-negligible role of soil archaeal communities in improving microbial stability of tomato fields under aeration drip irrigation. Sci. Hort. 319, 112179 (2023).
-
Wei, Y. et al. Effects of oil pollution on the growth and rhizosphere microbial community of Calamagrostis Epigejos. Sci. Rep. 15 (1), 1278 (2025).
-
Xun, W. et al. Specialized metabolic functions of keystone taxa sustain soil Microbiome stability. Microbiome. 9, 1–15 (2021).
-
Jena, J. et al. Role of Legumes in Cropping System for Soil Ecosystem Improvement. Ecosystem Services: Types, Management and Benefits, vol. 415 (Nova Science Publishers, Inc, 2022).
-
Bak, G. et al. Crop rotation affects biological properties of rhizosphere soil and productivity of Kimchi cabbage (Brassica Rapa ssp. pekinensis) compared to monoculture. Hortic. Environ. Biotechnol. 63 (5), 613–625 (2022).
-
Babalola, O. O. & Enagbonma, B. J. Dataset of shotgun metagenomic evaluation of sorghum bicolor rhizosphere Microbiome in soils preceded by Glycine max. Data Brief. 111270 (2025).
-
Zhong, Y. et al. Cooperative interactions between nitrogen fixation and phosphorus nutrition in legumes. New Phytol. 237 (3), 734–745 (2023).
-
Zhao, Y. et al. Variation in bacterial community structure in rhizosphere and bulk soils of different halophytes in the yellow river delta. Front. Ecol. Evol. 9, 816918 (2022).
-
Wang, P. et al. Effect of soil management systems on the rhizosphere bacterial community structure of tobacco: continuous cropping vs. paddy-upland rotation. Front. Plant Sci. 13, 996858 (2022).
-
Adedeji, A. A. & Babalola, O. O. Rhizosphere: A complex determinant of soil microbial community. Ann. Univ. Oradea Fasc Biol. 27 (1), 71–81 (2020).
-
Chen, J. et al. Bulk soil microbial reservoir or plant recruitment dominates rhizosphere microbial community assembly: evidence from the rare, endangered Lauraceae species cinmaomum Migao. Ecol. Ind. 148, 110071 (2023).
-
Ayangbenro, A. S. et al. Harnessing the rhizosphere soil microbiome of organically amended soil for plant productivity. Agronomy 12 (12), 3179 (2022).
-
Olanrewaju, O. S. & Babalola, O. O. The rhizosphere microbial complex in plant health: A review of interaction dynamics. J. Integr. Agric. 21 (8), 2168–2182 (2022).
-
Akinola, S. A. & Babalola, O. O. The fungal and archaeal community within plant rhizosphere: a review on their contribution to crop safety. J. Plant Nutr. 44 (4), 600–618 (2021).
-
Molefe, R. R., Amoo, A. E. & Babalola, O. O. Communication between plant roots and the soil microbiome; involvement in plant growth and development. Symbiosis. 90 (3), 231–239 (2023).
-
Van Cauwenberghe, J. & Simms, E. L. How might bacteriophages shape biological invasions? Mbio. 14 (5), e01886–e01823 (2023).
-
Moreira, Z. P. M. et al. Engineering plant microbiomes by integrating eco-evolutionary principles into current strategies. Curr. Opin. Plant. Biol. 71, 102316 (2023).
-
Enagbonma, B. J., Fadiji, A. E. & Babalola, O. O. Anthropogenic fertilization influences a shift in barley rhizosphere microbial communities. PeerJ. 12, e17303 (2024).
-
Molefe, R. R., Amoo, A. E. & Babalola, O. O. Metagenomic insights into the bacterial community structure and functional potentials in the rhizosphere soil of maize plants. J. Plant Interact. 16 (1), 258–269 (2021).
-
Liu, Q. et al. Changes in soil microbial biomass, diversity, and activity with crop rotation in cropping systems: A global synthesis. Appl. Soil. Ecol. 186, 104815 (2023).
-
Sun, L. et al. The shift of soil microbial community induced by cropping sequence affect soil properties and crop yield. Front. Microbiol. 14, 1095688 (2023).
-
Du, S. et al. Rhodococcus Qingshengii facilitates the phytoextraction of zn, cd, ni, and Pb from soils by sedum Alfredii hance. J. Hazard. Mater. 424, 127638 (2022).
-
Yadav, P., Chandra, R. & Pareek, N. Plant growth promoting mesorhizobia as a potential inoculant for Chickpea (Cicer arietinum L.): A review. Bhartiya Krishi Anusandhan Patrika. 37 (4), 328–333 (2022).
-
Rafique, M. et al. Tripartite microbial augmentation of Bradyrhizobium diazoefficiens, Bacillus sp. MN54, and Piriformospora indica on growth, yield, and nutrient profiling of soybean (Glycine max L). Front. Microbiol. 15, 1437489 (2025).
-
da Neto, C. Bacterial consortia among Bradyrhizobium species, azospirillum baldaniorum and Bacillus pumilus promote plant growth and efficient symbiotic nitrogen fixation in mung bean. Symbiosis. 93 (3), 255–267 (2024).
-
Kumar, M. et al. Biodiversity of Methylotrophic microbial communities and their potential role in mitigation of abiotic stresses in plants. Biologia. 74, 287–308 (2019).
-
Wang, F. et al. Differences in the soil prokaryotic Microbiome during continuous and single cultivation of three varieties of Chinese cabbage. Eurasian Soil. Sci. 57 (10), 1674–1687 (2024).
-
Wu, X. T. et al. Adaptive changes in bacterial and archaeal community structures and functions occur under primordial conditions. ACS Earth Space Chem. 8 (7), 1470–1479 (2024).
-
Li, H., Hill, N. & Wallace, J. A perennial living mulch system fosters a more diverse and balanced soil bacterial community. PLoS One. 18 (8), e0290608 (2023).
-
Mitra, D. et al. Actinobacteria-enhanced plant growth, nutrient acquisition, and crop protection: advances in soil, plant, and microbial multifactorial interactions. Pedosphere. 32 (1), 149–170 (2022).
-
Singh, P. et al. Unlocking the strength of plant growth promoting Pseudomonas in improving crop productivity in normal and challenging environments: a review. J. Plant Interact. 17 (1), 220–238 (2022).
-
Bovio-Winkler, P., Cabezas, A. & Etchebehere, C. Unveiling the hidden diversity and functional role of Chloroflexota in full-scale wastewater treatment plants through genome-centric analyses. ISME Commun. 4 (1), ycae050 (2024).
-
Zhang, Y. et al. Metagenomic insights into microbial variation and carbon cycling function in crop rotation systems. Sci. Total Environ. 947, 174529 (2024).
-
Ling, N., Wang, T. & Kuzyakov, Y. Rhizosphere bacteriome structure and functions. Nat. Commun. 13 (1), 836 (2022).
-
Chepsergon, J. & Moleleki, L. N. Rhizosphere bacterial interactions and impact on plant health. Curr. Opin. Microbiol. 73, 102297 (2023).
-
Benaissa, A. Rhizosphere: role of bacteria to manage plant diseases and sustainable agriculture—A review. J. Basic Microbiol. 64 (3), 2300361 (2024).
-
Zhang, M. et al. Molecular analysis of microbial nitrogen transformation and removal potential in the plant rhizosphere of artificial tidal wetlands across salinity gradients. Environ. Res. 215, 114235 (2022).
-
Abid, A. A. et al. Unraveling nitrogen loss in paddy soils: A study of anaerobic nitrogen transformation in response to various irrigation practice. Environ. Res. 252, 118693 (2024).
-
Zhu, H. et al. The accumulation of polysaccharides in dendrobium officinale is closely related to rhizosphere bacteria. Int. Microbiol. 1–12 (2024).
-
Hussain, B. et al. Niche-specific modulation of long-chain n-alkanes degrading bacterial community and their functionality in forest habitats across the leaf litter-soil compartments. Appl. Soil. Ecol. 195, 105248 (2024).
-
Cao, T. et al. Synergy of saprotrophs with mycorrhiza for litter decomposition and hotspot formation depends on nutrient availability in the rhizosphere. Geoderma. 410, 115662 (2022).
-
Hua, L. et al. Root exudate-driven rhizospheric recruitment of plant growth-promoting rhizobacteria. Pedosphere. 35 (1), 216–228 (2024).
-
Ou, Y. et al. Low dose phosphorus supplementation is conducive to remediation of heavily petroleum-contaminated soil—From the perspective of hydrocarbon removal and ecotoxicity risk control. Sci. Total Environ. 929, 172478 (2024).
-
Ma, J. et al. Update on new trend and progress of the mechanism of polycyclic aromatic hydrocarbon biodegradation by rhodococcus, based on the new understanding of relevant theories: A review. Environ. Sci. Pollut. Res. 30 (41), 93345–93362 (2023).
-
Grevesse, T. et al. Degradation pathways for organic matter of terrestrial origin are widespread and expressed in Arctic ocean microbiomes. Microbiome. 10 (1), 237 (2022).
-
Eichlerová, I., Šnajdr, J. & Baldrian, P. Laccase activity in soils: considerations for the measurement of enzyme activity. Chemosphere. 88 (10), 1154–1160 (2012).
-
Shobade, S. O., Zabotina, O. A. & Nilsen-Hamilton, M. Plant root associated chitinases: structures and functions. Front. Plant Sci. 15, 1344142 (2024).
-
Zhou, Z., Zhang, Y. & Zhang, F. Abundant and rare bacteria possess different diversity and function in crop monoculture and rotation systems across regional farmland. Soil Biol. Biochem. 171, 108742 (2022).
-
Wangthaisong, P. et al. The type IV secretion system (T4SS) mediates symbiosis between Bradyrhizobium sp. SUTN9-2 and legumes. Appl. Environ. Microbiol. 89 (6), e00040–e00023 (2023).
-
Yang, Y. et al. Deciphering microbial community and nitrogen fixation in the legume rhizosphere. J. Agric. Food Chem. 72 (11), 5659–5670 (2024).
-
Park, Y. et al. Strategies of organic phosphorus recycling by soil bacteria: acquisition, metabolism, and regulation. Environ. Microbiol. Rep. 14 (1), 3–24 (2022).
-
Liu, A. et al. Interaction and regulation of carbon, nitrogen, and phosphorus metabolisms in root nodules of legumes. Front. Plant Sci. 9, 1860 (2018).
-
Jin, L. et al. Effects of different vegetable rotations on the rhizosphere bacterial community and tomato growth in a continuous tomato cropping substrate. PLoS One. 16 (9), e0257432 (2021).
-
Philippot, L. et al. The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 22 (4), 226–239 (2024).
-
Schmidt, J. E. et al. Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling. Microbiome. 7, 1–18 (2019).
-
Carkner, M. K., Gao, X. & Entz, M. H. Ideotype breeding for crop adaptation to low phosphorus availability on extensive organic farms. Front. Plant Sci. 14, 1225174 (2023).
-
Xue, P. et al. Distinctive role of soil type and land use in driving bacterial communities and carbon cycling functions down soil profiles. Catena. 223, 106903 (2023).
-
Xia, Q. et al. Soil pore size distribution shaped not only compositions but also networks of the soil microbial community. Appl. Soil. Ecol. 170, 104273 (2022).
-
Shao, L. et al. Soil organic matter and water content affect the community characteristics of arbuscular mycorrhizal fungi in Helan mountain, an arid desert grassland area in China. Front. Microbiol. 15, 1377763 (2024).
Acknowledgements
The authors acknowledge the support of the management of Rosaly Farms Tarlton, Gauteng, South Africa, for granting permission to access and sample their carrot farmland soil. AAA thanks the North-West University for the Doctoral bursary. Also, OOB acknowledges the grant from the International Centre for Genetic Engineering and Biotechnology (ICGEB) (Grants number: CRP/ZAF22-93).
Funding
The International Centre for Genetic Engineering and Biotechnology (ICGEB) (Grants number: CRP/ZAF22-93) awarded to OOB supported this work.
Ethics declarations
Consent for publication
All authors approved the manuscript for publication.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Adebayo, A.A., Enagbonma, B.J. & Babalola, O.O. Comparative metagenomics on community structure and diversity of rhizomicrobiome associated with monoculture and soybean precedent carrot. Sci Rep 15, 28161 (2025). https://doi.org/10.1038/s41598-025-13605-z
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-13605-z