
Introduction
Aging is a progressive decline in physiological integrity marked by homeostasis dysregulation, diminished physiological reserve, declining organ function, increased morbidity, and social vulnerability that ultimately results in a cascade of intertwined adverse health outcomes1. While aging research traditionally employs diverse approaches – biological, chronological, or socioeconomic – the World Health Organization (WHO) has proposed a novel framework focusing on phenotypic presentations across various functional domains2. The WHO defines healthy aging as lifelong optimization of functional ability through intrinsic capacity, a composite biological construct encompassing locomotion, cognition, vitality, psychological well-being, and sensory function. Cross-sectional3 and longitudinal studies4 revealed that individual intrinsic capacity domains do not independently influence health outcomes. Instead, these domains exhibit synergistic interactions, with clustering of impairments across domains potentiating the risk of adverse health consequences. Among intrinsic capacity domains, combined decline in cognition and mobility, termed physio-cognitive decline (PCD), shows the highest prevalence and is associated with the most severe health outcomes3,4.
Within the spectrum of dual cognitive and mobility decline5, physio-cognitive decline (PCD) is gaining research focus due to its association with severe health consequences like disability, dementia, and mortality6,7, as well as its potential reversibility with targeted intervention8,9. While large-scale epidemiological studies have solidified the significance of PCD as a distinct clinical entity, further investigations at the pathophysiological level have revealed specific findings, such as the involvement of miR-29b-3p10 and the cerebellar-limbic neurocircuit11. Despite the identification of novel biological findings, the underlying mechanisms responsible for PCD pathophysiology remain to be fully elucidated.
Metabolomics, a powerful systems-level approach, unveils distinct molecular signatures in biofluids like blood for complex syndrome like PCD. These signatures hold promise for elucidating biological pathways, novel diagnostics, and therapeutic targets12. While a few studies have explored metabolomic biomarkers for similar concepts like motoric cognitive risk syndrome13, the potential of metabolomics to uncover the biological underpinnings of PCD and shed light on muscle-brain crosstalk is largely unexplored. In addition to PCD, as defined by cross-sectional data, another form of concurrent cognitive and mobility decline has been characterized through long-term follow-up of cognitive and mobility changes over time. The dual decline group showed a significant association with future dementia risk, and both targeted and untargeted metabolomics approaches have been used to study the metabolomic profiles of dual decline compared to no decline groups14,15,16.
This study employs untargeted metabolomics to profile over 3800 metabolites in serum from well-characterized PCD and controls from Asian aging cohorts in Taiwan and Japan. By comparing metabolic profiles, we aim to identify biomarkers and pathways associated with PCD, hypothesizing that circulating metabolites reflect a muscle-brain crosstalk due to the established link between mobility frailty and cerebellar dysfunction in PCD11. Identifying these metabolic signatures could elucidate muscle-brain crosstalk, enabling effective PCD screening for targeted interventions and promoting health span.
Based on Limma results adjusting for clinical confounders across three Asian cohorts, this study demonstrates substantial perturbation of the serum metabolome in PCD. Specifically, both the glutathione metabolism and tryptophan metabolism pathways exhibit marked downregulation. Furthermore, we identify and validate eleven PCD-associated metabolites using reference standards, thereby establishing a potential biomarker panel that could enhance the diagnostic accuracy for PCD.
Methods
Study design and participants
This study leverages data from three existing population-based longitudinal aging cohorts: The Longitudinal Aging Study of Taipei (LAST) and the I-Lan Longitudinal Aging Study (ILAS) in Taiwan, and the National Institute for Longevity Sciences, Longitudinal Study of Aging (NILS-LSA) in Japan17. The physical function in ILAS and NILS-LSA was measured by handgrip strength or gait speed, and in the LAST cohort, handgrip strength and 5-time chair stand test were represent physical function in the LAST cohort to define mobility impairment of PCD. This study excluded participants who were younger than 65 years old or exhibited persons with disability and dementia. The definition of probable dementia was defined according to the following criteria: (1) total Mini-Mental State Examination (MMSE) score less than 24 for ILAS and NILS-LSA participants with six or more years of education or less than 14 for those with less than six years of education; (2) total Montreal Cognitive Assessment (MoCA) score less than 26 for LAST participants. To account for potential confounders like cultural background, socioeconomic status, and educational attainment inherent to each geographically distinct cohort, this study employed a case-control design within cohorts, followed by pooled analysis for enhanced statistical power. Specifically, we identified individuals exhibiting the lowest 20th percentile in physical and cognitive function as putative PCD cases and compared them to those in the top 20th percentile, designated as controls. Finally, we matched 197 pairs from 3 cohorts for analysis. All participants of this study provided written informed consent. The study protocol received approval from the Institutional Review Board (IRB) of National Yang-Ming University (No. YM103008 for ILAS; YM104121F-3 for LAST) and the Committee on Ethics of Human Research of the National Center for Geriatrics and Gerontology (No. 1453 for NILS-LSA). All datasets were accessed under appropriate ethical approvals and through direct collaboration with the principal investigators of each cohort, all of whom are listed as corresponding authors or co-authors in this manuscript. For the present study, data were utilized from the first wave of the LAST (2016 to 2019), third wave of ILAS (2018 to 2020), and seventh wave survey of NILS-LSA (2010 to 2012).
Demographic characteristics and functional assessment
Baseline demographic characteristics, lifestyle factors, and self-reported comorbid conditions (medical history) were collected through face-to-face interviews using validated questionnaires. Tobacco usage was categorized as either current smoking or non-smoking, while the status of alcohol consumption was divided into drinking or non-drinking habit. Cognitive function was assessed using MMSE for ILAS and NILS-LSA participants and MoCA for LAST participants. Handgrip strength was measured using a dynamometer (Smedley’s Dynamometer; TTM, Tokyo, Japan) of the dominant hand, while 6-meter walking speed was measured from a static start without deceleration. The five-time sit-to-stand test measured the time to complete five consecutive sit-to-stand movements. Within each cohort, a minimum of two of the following physical performance assessments were conducted: (1) 6-meter gait speed; (2) five-time sit-to-stand duration; (3) grip strength. Mobility impairment no disability (MIND) was determined if any one of the two tests falls below the sex-specific lowest quintile values. Cognitive impairment no dementia (CIND) was defined by the lowest quintile of MoCA or MMSE scores as cutoff values. Subjects exhibiting concurrent MIND and CIND were categorized as having PCD. For each PCD case, we manually selected one control participant whose physical and cognitive functions ranked in the highest quintile. Matching criteria included the sex and age (within 3 years). The flow chart illustrating participant selection in the NILS-LSA cohort is presented in Supplementary Fig. S1. Consequently, a total of 197 PCD cases with 197 controls, aged between 65 and 87 years, were included in the current analysis.
Serum metabolomics
Peripheral venous blood samples were obtained from all participants after 10 h overnight fast, and subsequently centrifuged at 4 °C and 3000 × g for 10 min to obtain serum samples, which were then stored at −80 °C until analysis. Each serum sample (60 µL) was spiked with internal standards and deproteinized by the addition of 240 µL methanol. Following centrifugation at 4 °C and 13,000 × g for 10 min, the resulting supernatant was lyophilized and stored at −80 °C for subsequent analysis.
Ultra-performance liquid chromatography-mass spectrometry (UPLC-MS) analysis was performed using a Waters Xevo G2-XS Q-Tof tandem mass spectrometer coupled with Acquity UPLC (Waters), employing a BEH C18 column (2.1 × 100 mm, 1.7 µm, Waters) maintained at 45 °C. The total run time was 9 minutes with a flow rate of 0.3 mL/min. Gradient conditions for sample analysis included 1% B from 0 to 0.5 minutes, 1–100% B from 0.5 to 4.5 minutes, and holding at 100% B for 0.5 minute, followed by a return to the initial composition over 1 minute; these final conditions were maintained for an additional 3 minutes. Mobile phase A consisted of an aqueous solution containing 0.1% formic acid, while mobile phase B comprised acetonitrile containing 0.1% formic acid. Leucine enkephalin was utilized for continuous mass calibration. MSE data in both negative and positive ion modes were acquired at both low and high collision energy, with a mass scan range of 50–1200 m/z. Furthermore, a pooled quality control (QC) reference was prepared, aliquoted, and analyzed during each LC-MS batch to facilitate normalization between different batches. The QC sample was injected at regular intervals, specifically after every six study samples, during the analytical run.
LC-MS data processing and analysis
Peak areas for metabolites were analyzed using Progenesis QI v3.0 (Nonlinear Dynamics, Waters) software. Molecular features were characterized by retention time, m/z, and ion intensity. Metabolites with a retention time exceeding 6 minutes were excluded from the analyses. Following default settings in ‘normalize to all compounds’ within Progenesis, the dataset underwent further processing using StatTarget18, employing the QC-based random forest signal correction (QC-RFSC) algorithm to normalize ion intensities, filter peaks with low quality (CV > 30%), and perform missing value imputation. Subsequently, ion intensities from duplicate measurements of each sample were averaged. Tentative identification of ionic features was achieved by matching monoisotopic mass (m/z) values with those in the Human Metabolome Database with a mass tolerance of 20 ppm. For in-house library matching, metabolites were identified by comparing retention times (<0.5 min) and m/z values (<10 ppm) of peaks with those of standards.
Multivariate analysis and pathway mapping
Principal component analysis (PCA) and orthogonal partial least-squares discriminant analysis (OPLS-DA) models scaling to unit variance were built using EZinfo (Umetrics, version 3.0). The overall quality of the OPLS-DA model was assessed by cumulative modeled variance R2Y (cum), seven-fold cross-validation predictive ability Q2 (cum) values, as well as a permutation test by MetaboAnalyst19. For the biological interpretations of untargeted metabolomics data, the Functional Analysis module of MetaboAnalyst 5.0 was utilized. Both Mummichog-based hypergeometric and gene set enrichment analysis (GSEA) algorithms were used. All differential metabolites with a P-value less than 0.05 were included in the Mummichog enrichment analysis. The analysis was conducted on the combined dataset for both positive and negative ion modes. A molecular weight tolerance of 10 ppm was employed, and features were ranked using t-scores. Matches were identified corresponding to [M + H]+, [M-H2O + H]+, [M + NH4]+, or [M+Na]+ adducts in ESI+ mode, and [M-H]–, [M-H2O-H]–, [M-2H]2-, or [M+Cl]– adducts in ESI- mode.
Statistics and reproducibility
Differential abundance analysis of log-transformed metabolite levels was performed using Limma (linear models for microarray data), which delivers powerful inference through variance modeling and moderated t-statistics20. The analysis adjusted for age, gender, education (>12 years [1], ≤12 years [0]), and comorbidities (hypertension, diabetes, osteoporosis, CKD). Each study participant constituted an independent biological replicate for statistical analysis. Sample size was determined based on feasibility considerations and comparable studies in the field. In the demographics table, continuous variables are reported as mean ± standard deviation, whereas categorical variables are presented as counts (percentages). Descriptive characteristics were compared using Student’s t-test or Chi-square analysis as appropriate. A significance threshold of 0.05 (P < 0.05) was adopted unless otherwise specified.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Results
Characteristics of the study population
We examined the serum metabolomic profile of individuals with PCD and controls in three aging cohorts: the Longitudinal Aging Study of Taipei (LAST, n = 202) and the I-Lan Longitudinal Aging Study (ILAS, n = 70) from Taiwan, and the National Institute for Longevity Sciences, Longitudinal Study of Aging (NILS-LSA, n = 122) from Japan. The mean age of all participants was 71.6 ± 4.7 years, with 63 % being female. Table 1 provides a summary of the baseline characteristics of all participants, including basic characteristics, functional assessments, anthropometric measurements, and laboratory results, and compares the two groups. PCD group exhibited a slightly older age, lower education level, and significantly worse physical and cognitive function compared with the control group.
Serum metabolomics profiling of PCD
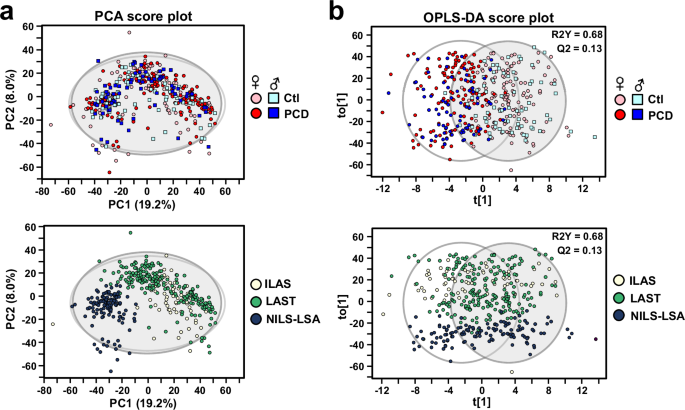
After signal correction and quality assurance procedures, 1885 and 1990 metabolic features were retained in positive and negative ion modes, respectively. We first evaluated stability and reproducibility of the LC-MS method by performing principal component analysis (PCA) on all samples, including the quality control (QC) samples. The QC samples clustered closely together in the PCA score plot (Supplementary Fig. S2a, b), indicating good analytical reproducibility. Additionally, the normalized abundance of dehydroepiandrosterone sulfate (DHEA-S) feature correlated with actual DHEA-S concentrations, with a correlation coefficient of 0.87 (Supplementary Fig. S2c), further confirming the analytical reliability. In applying statistical dimension reduction tools, while unsupervised PCA did not provide apparent separation between control and PCD samples (Fig. 1a), supervised Orthogonal partial least squares discriminate analysis (OPLS-DA) showed moderate differentiation between two groups (Fig. 1b). The OPLS-DA validation parameters were R2Y = 0.685 and Q2 = 0.131, both with permutation test P-value less than 0.05, indicating a modest but significant predicting performance (Supplementary Fig. S2d). We observed noticeable separation among Taiwan (ILAS & LAST) and Japan (NILS-LSA) cohorts (Fig. 1), indicating intrinsic differences in metabolite composition between different regions.

a PCA score plot for PCD and control samples based on 3875 metabolic features from positive and negative ion modes. Squares represent males, circles represent females; light colors (light blue, pink) indicate controls, dark colors (blue, red) indicate PCD. Lower panel: same PCA score plot stratified according to ILAS (light yellow), LAST (green) or NILS-LSA (dark blue) cohorts. b Score plots of OPLS-DA model built to discriminate between PCD and control subjects (R2Y = 0.68, Q2 = 0.13). Each dot represented an individual subject and color-coded based on grouping of PCD or cohort. Lower panel: same OPLS-DA score plot stratified by cohort. PCD physio-cognitive decline. Scaling: unit variance. ILAS I-Lan Longitudinal Aging Study, LAST Longitudinal Aging Study of Taipei, NILS-LSA National Institute for Longevity Sciences, Longitudinal Study of Aging.
Significant differential metabolites and pathways associated with PCD
Differential metabolite abundance was assessed using Limma based on linear regression models20. Age, sex, educational status, and four disease histories (hypertension, diabetes, osteoporosis, chronic kidney disease) were included as covariates (P < 0.1 in at least one of the three cohorts in Table 1). Of the detected 3875 metabolic features, 606 were found to be significantly different at a significance level of 0.05 (40 with FDR < 0.05 followed by Benjamini-Hochberg correction), as represented in volcano plot (Fig. 2a; Supplementary Table S1). To gain insight into these differential metabolic features, we used MetaboAnalyst to perform pathway analysis based on Mummichog and GSEA algorithms. Both Mummichog and GSEA results were summarized by Fisher’s combined probability test to calculate the combined P values. The top 17 pathways with Pcombined < 0.1 were highlighted in Fig. 2b. Among them, glutathione metabolism and tryptophan metabolism pathways were negatively regulated, while polyunsaturated fatty acid biosynthesis and bile acid biosynthesis pathways were active based on the enrichment score of GSEA (Supplementary Table S2).

a Volcano plot representing the differences in metabolite abundance in the serum samples of PCD (n = 197) and healthy controls (n = 197). The P values were obtained from Limma models including age, sex, educational status, and four disease history as covariates. The grey horizontal line on the plot shows the significant level (P value of 0.05). Up-regulated metabolic features indicated in red (n = 203) and down-regulated colored in blue (n = 403). b Significant enrichment of 17 pathways related by PCD using GSEA and mummichog algorithms. The size of the dot is proportional to its combined P value, which is calculated by Fisher’s method. PCD, physio-cognitive decline; GSEA, gene set enrichment analysis.
Association of PCD with serum metabolites in glutathione, tryptophan, urea cycle, and bile acid metabolism pathways
To ensure annotation accuracy, significant metabolites predicted by GSEA and mummichog algorithms were manually validated against authentic reference standards where available. The comprehensive list of validated metabolites is presented in Supplementary Fig. S3. Metabolites lacking sufficient MS/MS or retention time validation were excluded from subsequent analyses. Glutathione metabolism was the most significantly altered pathway associated with PCD, characterized by reduced levels of pyroglutamic acid (P = 0.035) and γ-glutamylcysteine (P = 0.004), both key components of the γ-glutamyl cycle (Fig. 3a). It might suggest dysregulation of glutathione homeostasis. Tryptophan metabolism was identified as the second most significantly altered pathway associated with PCD. Subjects with PCD exhibited markedly reduced levels of multiple tryptophan metabolites, including 3-hydroxykynurenine, anthranilic acid, tetrahydro-beta-carboline (THβC), tryptophan, 5-hydroxytryptophan, and melatonin (Fig. 3b). In the urea cycle/amino group metabolism pathway, we observed significant reductions in argininosuccinic acid, a critical intermediate in the urea cycle, and creatinine, a non-enzymatic breakdown product of creatine (Fig. 4a, b). Furthermore, the PCD group demonstrated significant alterations in bile acid metabolism compared to controls. Specifically, we detected significantly elevated levels of 7α-hydroxycholesterol, glycocholic acid, and taurodeoxycholic acid (P < 0.05) in PCD subjects. Additionally, we observed increasing trends in glycochenodeoxycholic acid levels and decreasing trends in lithocholic acid levels (P < 0.1). These findings collectively indicate dysregulation in both primary bile acid synthesis and secondary bile acid metabolism in PCD subjects (Fig. 4c, d).

a Serum levels of γ-glutamylcysteine and pyroglutamic acid involved in glutathione metabolism in PCD relative to control. Right panel: schematic of the γ-Glutamyl cycle. b Changes in metabolite levels involved in tryptophan metabolism in PCD relative to control. Right panel: a representation of molecules involved in tryptamine, kynurenine and serotonin pathway of tryptophan metabolism. PCD physio-cognitive decline. The annotated metabolites with direction of changes were indicated with colored arrows in the boxes. Metabolites shown in outlined box without arrows denote detectable but no marked change.

a Serum levels of argininosuccinic acid and creatinine involved in urea cycle/amino group metabolism in PCD relative to control. b A representation of molecules involved in urea cycle/amino group metabolism. c Serum levels of four bile acids (lithocholic acid, glycochenodeoxycholic acid, taurodeoxycholic acid, and glycocholic acid) and their precursor 7α-hydroxycholesterol in PCD versus control. d Schematic of the altered metabolites associated with bile acid metabolism. The annotated metabolites with direction of changes were indicated with colored arrows in the boxes. Metabolites shown in the outlined box without arrows denote detectable but no marked change.
Potential effects of gender and region on PCD metabolomic profiles
We further investigated potential gender-specific alterations in serum metabolomic profiles. As illustrated in Supplementary Fig. S4, notable sex differences were observed: the OPLS-DA model demonstrated a substantially higher R2Y value in females (0.86) compared to males (0.39) when comparing PCD and control groups. Limma modeling revealed 830 significant metabolic features (P < 0.05) in females versus 244 in males, with minimal overlap (59 features). We performed additional ANCOVA analyses on the complete dataset (Taiwan and Japan combined) with household income as an additional covariate. In these analyses, most 11 PCD markers retained significance after income adjustment, with exceptions only among borderline-significant metabolites (Supplementary Fig. S5a). Subsequently, we conducted stratified analyses by geographical region, performing separate differential expression analyses using Limma for Taiwanese and Japanese populations independently, without income adjustment. Several metabolites exhibited consistent regulation trends across regions, although statistical significance was limited in the NILS-LSA cohort (Supplementary Fig. S5b). Notably, pyroglutamic acid, anthranilic acid, and THβC displayed divergent regulation patterns between Taiwanese and Japanese cohorts.
Discussion
To the best of our knowledge, this is the first exploratory study to leverage data from multiple international, population-based aging cohorts to identify metabolomic signatures and potential biological mechanisms associated with PCD. Despite the OPLS-DA model’s inability to achieve complete separation between PCD and control groups (Fig. 1b), Limma analysis, adjusted for age, sex, education status, and comorbid conditions (hypertension, diabetes, osteoporosis, and chronic kidney disease), identified 606 significantly differentiated metabolic features. These features further implicated 17 distinct metabolic pathways. The metabolic changes pointed towards potential disruptions in energy homeostasis, antioxidant systems, lipid and amino acid metabolism processes underlying PCD. The metabolomics approach provided metabolic signatures and dysregulated pathways associated with dual declines, offering potential biomarkers and biological insights. A previous longitudinal study, utilizing plasma metabolomics data from both baseline and follow-up, analyzed with the MxP Quant 500 kit, identified the 10 most significant metabolites associated with longitudinal decline of both memory and gait14. Notably, we observed decreasing trends for two of these metabolites, lysoPC (C18:0) and histidine, in the PCD group (P < 0.1, Supplementary Table S3). A recent untargeted metabolomics study, employing four different LC-MS methods, demonstrated a total of 26 metabolites associated with cognitive decline and gait decline16. Among these, lysoPC (C20:5), which exhibited higher concentrations correlated with a reduced risk of dementia, also showed a significant reduction in the PCD group (P < 0.05, Supplementary Table S3). These findings suggest that specific metabolites, including lysoPC (C18:0), histidine, and lysoPC (C20:5), exhibit similar trends in both longitudinal and cross-sectional analyses of dual cognitive and mobility decline. This study identified dysregulation of glutathione metabolism, evidenced by decreased levels of pyroglutamic acid and γ-glutamylcysteine in PCD (Fig. 3), as a major pathway associated with the condition. Pyroglutamic acid, also known as 5-oxoproline, is an intermediate substrate involved in glutathione synthesis via the γ-glutamyl cycle. Reduced plasma pyroglutamic acid levels have been linked to step reduction21 and autism spectrum disorders22. The dual correlation of pyroglutamic acid levels may reflect a shared metabolic vulnerability affecting both muscle and brain function. Therapeutically, γ-glutamylcysteine has been shown to ameliorate oxidative injury in neurons23 and exhibit anti-inflammatory and anti-amyloid-β properties in mouse models24. While our findings indicated a reduction in systemic pyroglutamic acid and γ-glutamylcysteine levels in individuals with PCD, further studies are needed to explore whether restoring related metabolic pathways could mitigate age-related decline in cellular function. Several tryptophan metabolites exhibited significant alterations in the PCD group (Fig. 3b), including reduced levels of 3-hydroxykynurenine (3-HK), 5-hydroxytryptophan (5-HTP), melatonin, and tetrahydro-β-carboline (THβC). As tryptophan is a precursor to multiple metabolic pathways—namely serotonin, kynurenine, and indole pathways—these findings collectively suggest a potential disruption in systemic tryptophan metabolism in individuals with PCD. Tryptophan metabolism is crucial for immunity, neuronal function, and gut health, and its dysregulation in conditions such as cancer and neurodegeneration highlights its potential as a therapeutic target25. While 3-HK is considered as a neurotoxic metabolite, evidence from certain studies has demonstrated its ability to inhibit spontaneous lipid peroxidation and function as an antioxidant26. These findings indicate that 3-HK may act as a redox modulatory molecule rather than a solely neurotoxic metabolite27,28. While previous studies have reported elevated 3-HK levels in frail individuals29, our findings in the PCD cohort revealed a divergent pattern. This discrepancy may reflect differences in the underlying biological context, particularly given that PCD involves both cognitive and physical decline. Our data demonstrated altered levels of several bile acids, including glycocholic acid, lithocholic acid, and taurodeoxycholic acid, in individuals with PCD (Fig. 4c, d). Bile acids are synthesized from cholesterol in liver and are subsequently secreted following conjugation to either glycine or taurine, forming glycocholic acid and taurocholic acid, respectively. In the gastrointestinal tract, the gut microbiota further metabolize these primary bile acids into secondary bile acids through dehydroxylation or deconjugation processes. Recent studies have uncovered a potential link between gut microbiota and skeletal muscle, elucidating a new ‘gut–muscle axis’30, which is mediated by microbially derived bile acids. Alterations in bile acid composition have been observed in sarcopenia within liver cirrhosis patients31. In addition, cholic acid and deoxycholic acid have been shown to induce an atrophic effect on skeletal muscle fibers through the bile acid receptor, TGR5 (Takeda G protein-coupled receptor 5)32. Moreover, recent evidence indicates that circulating bile acids can cross the blood-brain barriers and interact with receptors in the brain33, potentially explaining the association between serum bile acid levels and cognitive decline.
Given that serum creatinine is primarily generated in skeletal muscle from phosphocreatine and creatine, the decreased amount of creatinine secreted into circulation might indicate a reduction in skeletal muscle mass. Similar reductions in serum creatinine have been reported in patients with Duchenne muscular dystrophy, where muscle wasting is a prominent feature34,35. While our findings align with the notion that lower creatinine may indicate compromised muscle status, further studies are needed to clarify the metabolic implications and whether creatinine levels could serve as a surrogate marker for muscle health in this context. Previous research has described PCD as a manifestation of accelerated aging36. To explore this further, we compared the metabolites associated with PCD in our study to known age-related metabolites reported in six prior studies37,38,39,40,41,42, as shown in Supplementary Table S4. While we observed a partial overlap, with several metabolites showing consistent age-related trends, others differed in direction. Such findings are consistent with the notion that PCD reflects an accelerated or divergent aging trajectory, though further validation is needed.
Our analysis revealed gender differences in both OPLS-DA and Limma models (Supplementary Fig. S4), with greater serum metabolomic separation observed in females. While sample size differences (males: n = 144; females: n = 250) may have influenced statistical power, the observed separation is consistent with our previous findings in the ILAS cohort, where female PCD subjects showed more distinct metabolomic profiles43. This pattern aligns with other studies reporting more pronounced age-related serum/plasma metabolomic changes in females37,42, underscoring the importance of sex as a key variable in aging-related metabolomics. While Li et al. identified metabolomic changes linked to concurrent cognitive and mobility decline, they did not perform sex-stratified analyses13. Although our current study addresses both sexes, further investigation with balanced sample sizes is needed to clarify how sex-specific factors influence the serum metabolome in PCD. In the subgroup analysis, ANCOVA was conducted to assess potential socioeconomic and regional influences. Including household income as a covariate did not alter our primary findings, suggesting minimal impact from socioeconomic factors. However, some PCD-related metabolites showed inconsistent trends between Taiwanese (ILAS & LAST) and Japanese (NILS-LSA) cohorts (Supplementary Fig. S5). Country-stratified analyses suggested potential cultural or environmental differences in biomarker profiles, though reduced statistical power limits firm conclusions. Notably, both Taiwanese and Japanese PCD participants showed decreased glutamylcysteine and 3-hydroxykynurenine levels (P ≤ 0.1), implicating glutathione and tryptophan metabolism as shared pathways in PCD across Asian populations.
Despite comprehensive research efforts, this study acknowledges some limitations. First, the inherent challenge of metabolite identification in untargeted metabolomics limited our characterization to a subset of observed features, despite the use of bioinformatic tools and authentic standards. The utility of these identified biomarkers may be constrained by the scarcity of knowledge about the affected biological pathways and function. Second, variables such as dietary intake, medical history, drugs, or supplements taken by the study participants could not be controlled in the present observational study design. Third, potential differences between the three cohorts could also be a concern, as revealed by the PCA and OPLS-DA models. Additional studies considering lifestyle factors in ethnically-diverse populations are warranted for better clarification. Fourth, although investigating the relationship between changes in circulatory metabolites with skeletal muscle or other organs is important, access to tissue specimens is critical to confirm the biological mechanisms. Thus, the mechanistic information provided through metabolite levels in circulation should be interpreted with caution. On the other hand, this study has several key strengths. First, it is the first large-scale, multi-cohort investigation of circulating metabolomic profiles related to PCD. Second, our results revealed novel associations between PCD and dysregulated pathways involving bile acid synthesis, glutathione metabolism, and tryptophan metabolism. Third, we verified 11 of the identified PCD markers using reference standards. These 11 metabolites could potentially form a biomarker panel to aid in the diagnosis of PCD and guide clinical decision-making. In conclusion, this study, analyzing data from the Taiwan (ILAS & LAST), and Japan (NILS-LSA) cohorts, identifies a distinct serum metabolomic signature for PCD in glutathione, bile acid, and tryptophan pathways. These findings provide a valuable foundation for future PCD metabolomic studies with the potential to inform interventions promoting healthy aging, but require further investigation with targeted assays to validate potential therapeutic targets or clinical outcome correlations.
Data availability
To protect the privacy of the participants, the data supporting the findings of this study are only available upon reasonable request from the corresponding author. Data access may be obtained through direct collaboration with the principal investigators of each cohort (L.-K.C. for ILAS, L.-N.P. for LAST, and H.A. for NILS-LSA participants) following appropriate ethical review and approval. The source data underlying all figures are available in the Supplementary Files: Supplementary Data 1 (Fig. 1), Supplementary Data 2 (Fig. 2a), and Supplementary Table S2 (Fig. 2b).
References
-
Folake Olubukola, A. & Olalekan Bukunmi, O. Deciphering the mechanisms, biochemistry, physiology, and social habits in the process of aging. Arch. Gerontol. Geriatr. 1, 100003 (2024).
-
Beard, J. R. et al. The World report on ageing and health: a policy framework for healthy ageing. Lancet 387, 2145–2154 (2016).
-
Meng, L. C. et al. Intrinsic capacity impairment patterns and their associations with unfavorable medication utilization: a nationwide population-based study of 37,993 community-dwelling older adults. J. Nutr. Health Aging 26, 918–925 (2022).
-
Meng, L. C. et al. Multi-trajectories of intrinsic capacity decline and their impact on age-related outcomes: a 20-year National Longitudinal Cohort Study. Aging Dis. 15, 2697–2709 (2023).
-
Tian, Q. et al. Dual cognitive and mobility impairments and future dementia – Setting a research agenda. Alzheimers Dement. 19, 1579–1586 (2023).
-
Chung, C. P. et al. Physio-cognitive decline syndrome as the phenotype and treatment target of unhealthy aging. J. Nutr. Health Aging 25, 1179–1189 (2021).
-
Lee, W. J. et al. Six-year transition of physio-cognitive decline syndrome: Results from I-Lan Longitudinal Aging Study. Arch. Gerontol. Geriatr. 102, 104743 (2022).
-
Liang, C. K. et al. Efficacy of Multidomain intervention against physio-cognitive decline syndrome: a cluster-randomized trial. Arch. Gerontol. Geriatr. 95, 104392 (2021).
-
Lee, W. J. et al. Enhancing intrinsic capacity and related biomarkers in community-dwelling multimorbid older adults through integrated multidomain interventions: ancillary findings from the Taiwan Integrated Geriatric (TIGER) Trial. J. Am. Med Dir. Assoc. 25, 757–763 e754 (2024).
-
Yang, C. P. et al. Muscle atrophy-related myotube-derived exosomal microRNA in neuronal dysfunction: Targeting both coding and long noncoding RNAs. Aging Cell 19, e13107 (2020).
-
Liu, L. K. et al. Cerebellar-limbic neurocircuit is the novel biosignature of physio-cognitive decline syndrome. Aging 12, 25319–25336 (2020).
-
Newgard, C. B. Metabolomics and metabolic diseases: where do we stand?. Cell Metab. 25, 43–56 (2017).
-
Li, W. et al. Plasma metabolomics and lipidomics signatures of motoric cognitive risk syndrome in community-dwelling older adults. Front. Aging Neurosci. 14, 977191 (2022).
-
Tian, Q. et al. Plasma metabolomic signatures of dual decline in memory and gait in older adults. Geroscience 45, 2659–2667 (2023).
-
Sultana, M. et al. A metabolomics analysis of a novel phenotype of older adults at higher risk of Dementia. J. Alzheimers Dis. 99, S317–s325 (2024).
-
Tian, Q. et al. Shared plasma metabolomic profiles of cognitive and mobility decline predict future dementia. Geroscience 46, 4883–4894 (2024).
-
Shimokata, H., Ando, F. & Niino, N. A new comprehensive study on aging-the National Institute for Longevity Sciences, Longitudinal Study of Aging (NILS-LSA). J. Epidemiol. 10, S1–S9 (2000).
-
Luan, H., Ji, F., Chen, Y. & Cai, Z. statTarget: A streamlined tool for signal drift correction and interpretations of quantitative mass spectrometry-based omics data. Anal. Chim. Acta 1036, 66–72 (2018).
-
Pang, Z. et al. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 17, 1735–1761 (2022).
-
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015).
-
Saoi, M. et al. Metabolic perturbations from step reduction in older persons at risk for Sarcopenia: Plasma biomarkers of abrupt changes in physical activity. Metabolites 9, 134 (2019).
-
Kuwabara, H. et al. Altered metabolites in the plasma of autism spectrum disorder: a capillary electrophoresis time-of-flight mass spectroscopy study. PLoS One 8, e73814 (2013).
-
Le, T. M. et al. gamma-Glutamylcysteine ameliorates oxidative injury in neurons and astrocytes in vitro and increases brain glutathione in vivo. Neurotoxicology 32, 518–525 (2011).
-
Liu, Y. et al. Supplementation with gamma-glutamylcysteine (gamma-GC) lessens oxidative stress, brain inflammation and amyloid pathology and improves spatial memory in a murine model of AD. Neurochem Int. 144, 104931 (2021).
-
Platten, M., Nollen, E. A. A., Rohrig, U. F., Fallarino, F. & Opitz, C. A. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 18, 379–401 (2019).
-
Leipnitz, G. et al. In vitro evidence for an antioxidant role of 3-hydroxykynurenine and 3-hydroxyanthranilic acid in the brain. Neurochem Int. 50, 83–94 (2007).
-
Colin-Gonzalez, A. L., Maldonado, P. D. & Santamaria, A. 3-Hydroxykynurenine: an intriguing molecule exerting dual actions in the central nervous system. Neurotoxicology 34, 189–204 (2013).
-
Colin-Gonzalez, A. L. et al. The Janus faces of 3-hydroxykynurenine: dual redox modulatory activity and lack of neurotoxicity in the rat striatum. Brain Res. 1589, 1–14 (2014).
-
Westbrook, R. et al. Kynurenines link chronic inflammation to functional decline and physical frailty. JCI Insight 5, e136091 (2020).
-
Mancin, L., Wu, G. D. & Paoli, A. Gut microbiota-bile acid-skeletal muscle axis. Trends Microbiol 31, 254–269 (2023).
-
Aliwa, B. et al. Altered gut microbiome, bile acid composition and metabolome in sarcopenia in liver cirrhosis. J. Cachexia Sarcopenia Muscle 14, 2676–2691 (2023).
-
Abrigo, J. et al. Cholic acid and deoxycholic acid induce skeletal muscle atrophy through a mechanism dependent on TGR5 receptor. J. Cell Physiol. 236, 260–272 (2021).
-
Xing, C. et al. Roles of bile acids signaling in neuromodulation under physiological and pathological conditions. Cell Biosci. 13, 106 (2023).
-
Boca, S. M. et al. Discovery of metabolic biomarkers for duchenne muscular dystrophy within a natural history study. PLoS One 11, e0153461 (2016).
-
Spitali, P. et al. Cross-sectional serum metabolomic study of multiple forms of muscular dystrophy. J. Cell Mol. Med. 22, 2442–2448 (2018).
-
Chen, L. K. & Arai, H. Physio-cognitive decline as the accelerated aging phenotype. Arch. Gerontol. Geriatr. 88, 104051 (2020).
-
Yu, Z. et al. Human serum metabolic profiles are age dependent. Aging Cell 11, 960–967 (2012).
-
Menni, C. et al. Metabolomic markers reveal novel pathways of ageing and early development in human populations. Int J. Epidemiol. 42, 1111–1119 (2013).
-
Auro, K. et al. A metabolic view on menopause and ageing. Nat. Commun. 5, 4708 (2014).
-
Chaleckis, R., Murakami, I., Takada, J., Kondoh, H. & Yanagida, M. Individual variability in human blood metabolites identifies age-related differences. Proc. Natl. Acad. Sci. USA 113, 4252–4259 (2016).
-
Lawton, K. A. et al. Analysis of the adult human plasma metabolome. Pharmacogenomics 9, 383–397 (2008).
-
Darst, B. F., Koscik, R. L., Hogan, K. J., Johnson, S. C. & Engelman, C. D. Longitudinal plasma metabolomics of aging and sex. Aging 11, 1262–1282 (2019).
-
Huang, Y. L. et al. Proteo-metabolomic insights for early dual physical and cognitive impairments: A search for biomarkers of healthy aging based on muscle-brain crosstalk. Aging Cell 24, e14407 (2025).
Acknowledgements
The authors acknowledge the technical services and support provided by the Mass Spectrometry Facility of National Yang Ming Chiao Tung University. The study was funded by the National Science and Technology Council (NSTC 112-2321-B-A49-006 and NSTC 113-2321-B-A49-012). This work is supported by the Interdisciplinary Research Center for Healthy Longevity of National Yang Ming Chiao Tung University from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education in Taiwan, National Health Research Institutes (NHRI-13A1-CG-CO-05-2426-3) and Intramural Research Program of the National Institute on Aging, Baltimore, MD.
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Medicine thanks the anonymous reviewers for their contribution to the peer review of this work. [A peer review file is available].
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Huang, YL., Chang, WJ., Lin, CH. et al. Data from multi-national aging cohorts show dysregulated metabolic pathways in people with physio-cognitive decline. Commun Med 5, 351 (2025). https://doi.org/10.1038/s43856-025-01073-5
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s43856-025-01073-5