
Introduction
Milk fat globule-EGF-factor 8 (MFGE8) also known as Lactadherin is a secreted extracellular glycoprotein originally found to be expressed abundantly in the mammary gland epithelium. Its transcripts level in the mammary gland increases from pregnancy to lactation, suggesting its role in the transitional development of the mammary gland1. Besides the mammary gland, it has also been discovered to be widely expressed in a variety of organs such as lungs, heart, kidney, spleen, intestine, and brain by a variety of cells including splenocytes, dendritic cells, macrophages, glial cells, intestinal epithelial cells and astrocytes2,3,4. Structurally, this protein has two domains: an N–terminal domain consisting of two repeated EGF-like domains and a C-terminal domain with two anionic phospholipid binding Discoidin/F5/8 C domains5. The RGD motif of the second EGF- like domain facilitates cell-cell adhesion by recognizing the integrin family of receptors viz. αvβ3 and αvβ5 6,7. The C terminal domain is involved in the immune function of phagocytes7,8. The MFGE8 has a more prominent role as an anti-inflammatory molecule, it has been shown to decrease the LPS-mediated production of proinflammatory cytokines from macrophages9, in the pathogenesis of SLE10,11,12, sepsis11,13,14, atherosclerosis15, rheumatoid arthritis, and osteoarthritis16,17. However in atherosclerosis, it has an inflammatory role as compared to other pathologies where it serve as an anti-inflammatory molecule. In addition to its crucial involvement in providing the architecture to the mammary gland and as an anti-inflammatory molecule, MFGE8 has a wide range of physiological functions viz. clearance of apoptotic cells7, promotion of colorectal cancer18, tumorigenesis19,20, fertilization21,22.
Knockout studies are widely used as standard protocol for the association of functions with the gene. The MFGE8 knockout studies has established its diverse roles in several pathophysiologies such as protective role in pancreatic inflammation, pulmonary fibrosis23, Endothelial-mesenchymal transition3. We hypothesize that MFGE8 gene knock out in BuMEC (Buffalo Mammary Epithelial Cell line) by CRISPR/Cas9 system in association with quantitative proteomics approach will reveal its role in milk synthesis and mammary gland development. Till date, we know that it plays a crucial role in mammary gland remodeling by facilitating the clearance of apoptotic cells during involution24. Earlier, our group identified MFGE8 as a marker protein for high milk yield in Sahiwal cows. The shRNA based knockdown of MFGE8 gene negatively impacted cytoskeletal remodeling and cell proliferation in comparison to the normal wild type cells26. In this study, a complete knockout of MFGE8 has been achieved using CRISPR cas-9 approach. The resulting changes in the total proteome has been investigated to understand the functional role MFGE8 in MECs.
Results
Successful generation of CRISPR/cas9 mediated MFGE8 knockout BuMEC
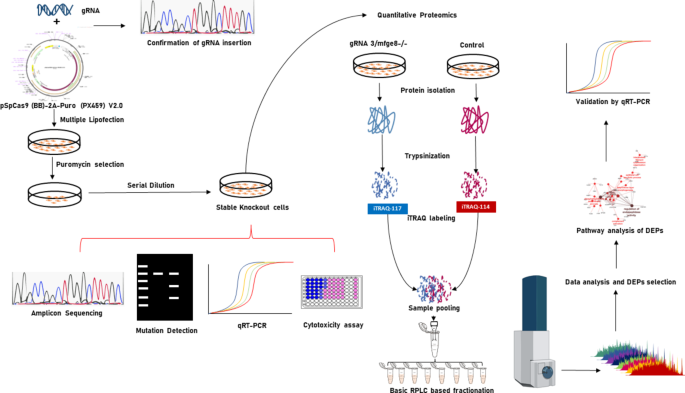
In this study, we generated MFGE8 gene knockout BuMEC using the CRISPR/Cas9-system, followed by the generation of a comprehensive proteome profile using quantitative proteomics and Fig. 1 provides a comprehensive depiction of the strategy utilized. To generate MFGE8 knockout BuMEC, three gRNAs viz. gRNA2, gRNA3 and gRNA4 (Fig. 2A) were synthesized against second and third exon of MFGE8 gene. gRNA2 and gRNA3 targeted the positive and negative strand of second exon respectively while gRNA4 targeted the positive strand of the third exon. The pSpCas9 (BB)-2A-puro expression vector constructs containing these gRNAs were sequenced (Fig. 2B) and transfected into BuMEC (Fig. 2C). Continuous puromycin selection and low density seeding were done to enrich the transfected cells. PCR based Guide-it Mutation Detection kit assay (Takara Bio USA, Inc.) showed single distinct band in control group while three bands were observed from MFGE8 knockout BuMEC in all three gRNAs (gRNA2- 742 bp, 527 bp, 215 bp; gRNA3- 716 bp, 506 bp, 210 bp; gRNA4- 657 bp, 378, 279 bp) (Fig. 2D) which revealed successful mutations at MFGE8 gene by the selected gRNAs. We also compared the expression level of cas9 and MFGE8 genes among control and knock-out cells obtained from the three gRNAs. The results showed that cas9 expressed abundantly in the transfected cells by all gRNAs but maximum expression of cas9 was observed in gRNA3 transfected cells (Fig. 3C). Next, MFGE8 gene expression was found to be significantly reduced in all three gRNA transfected cells as compared to control cells but gRNA3 much more efficiently reduced MFGE8 gene expression (Fig. 3A). Western blotting results exhibited loss of detectable MFGE8 in cells transfected by gRNA3 and thus confirmed the knock-out of MFGE8 gene (Fig. 3B). Thereby, gRNA3 transfected cells were used to validate mutations by NGS amplicon sequencing (Fig. 4). Approximately 54% of total reads showed indels, 3–4 bp upstream to PAM site in 2nd exon and the observed indels at the target site of MFGE8 occurred over a range of mutation sizes i.e. from 1 to 33 bp and are shown in Fig. 4. These cells were further cultured for several passages under puromycin selection and stable knock-out cells thus obtained were named as mfge8/-/gRNA3 and used for further experiments.
Post transfection, BuMEC appeared enlarged and distorted and also showed a slower growth rate as compared to control cells. To further study the cell growth and proliferation rates, MTT and BrDU assays were performed in control BuMEC and MFGE8 cells at time intervals for12h, 24 h, 36 h, 48 h, 60 h and 72 h. The results indicated that cell growth and proliferation rates declined significantly in mfge8-/-/gRNA3 cells as compared to control (Fig. 3D & E).
Proteome analysis by mass spectrometry
To explain how the loss of MFGE8 gene expression alters biological pathways, a global quantitative proteome analysis was performed using the three cell lines (mfge8-/-/gRNA2, mfge8-/-/gRNA3 and mfge8-/-/gRNA4 cells) resulting from three different guide RNAs used for genome editing. All three groups of MFGE8 knock-out cells and control BuMEC were grown to isolate proteins, equal amount of proteins from each group were taken for tryptic digestion, peptide iTRAQ labeling, sample fractionation and LC-MS/MS based analysis were performed. RAW files generated were analyzed with Proteome Discoverer (v2.2) against the Uniprot Bovine reference proteome database and both peptide spectrum match and protein false discovery rate were set to 0.01 FDR. A total of 4276 proteins were obtained from control BuMEC while mfge8-/-/gRNA2, mfge8-/-/gRNA3 and mfge8-/-/gRNA4 cells yielded 4278, 4282 and 4275 proteins respectively, amidst these, 4267, 4272 and 4270 proteins were shared by knock-out cells with control cells respectively (Supplementary Material 2 S1) (Fig. 5E). 99.3% proteins were found common among mfge8-/-/gRNA2, mfge8-/-/gRNA3 and mfge8-/-/gRNA4 cells and 0.07%, 0.05% and 0.2% proteins were found exclusively in mfge8-/-/gRNA2, mfge8-/-/gRNA3 and mfge8-/-/gRNA4 cells. Protein abundance ratios were log 2 transformed and the statistical analysis revealed 423, 154 and 178 differentially expressed genes (DEGs) among mfge8-/-/gRNA2v/s control, mfge8-/-/gRNA3 v/s control and mfge8-/-/gRNA4 v/s control cells respectively (Supplementary Material 3 S2). Quantitative proteomics data results also coincided with the transcriptomic expression of MFGE8 in cells created through three different gRNAs i.e. maximum reduction in MFGE8 at RNA and proteome level was observed in mfge8-/-/gRNA3 cells.
Although a significant decrease in fold change of lactadherin was observed in case of all three gRNAs as compared to control, the maximum decrease (0.36 fold) was observed in cell line edited with gRNA3 labelled with iTRAQ isobaric tag 117 (Fig. 5). Hence, further analysis was limited to mfge8-/-/gRNA3 group. The maximum sequence coverage of proteins identified in our data were in the range of 1–98% with maximum sequence coverage falling in the range of 1–50% (Fig. 5A). The overall abundance intensity profile of proteins were homogeneous throughout the different groups (Fig. 5B). In- depth proteome analysis of mfge8-/-/gRNA3 and control BuMEC, helped in prediction of localization of identified proteins in cellular compartments of BuMEC. We found that 15.95% proteins of whole proteome were from the nucleus, while 18.48% were associated with the cytoplasm. A major fraction of proteins i.e. around 21.41% belonged to membrane proteins and remaining proteins were dispersed in other cell compartments including cytoskeleton 3.81%, cytosol 12.30%, endoplasmic reticulum 3.90%, Golgi 3.24%, mitochondrial 7.38% and extracellular or secreted proteins 1.73% (Fig. 5F). Among 178 DEGs in mfge8-/-/gRNA3, total 124 were found to be differentially upregulated (cutoff ≥ 1.5 fold) compared to control BuMEC (Supplementary Material 3 S2) and these included SPC25, AFAP1L1, FABP5, S100A9, INHBA, APOA2, PBK, OSBPL1A, FABP4, TRAF5, KNG1, PALMD, STK3, NDUFA12, GJB2, RAB27B, VIM, ACTA2, APOA1 and S100A4 which are involved in pathways. 54 genes were found to be differentially downregulated (cut off ≤ 0.6 fold), the highly downregulated genes included SERPINH1, FABP3, KRT72, MFGE8, RBX1, KRT4, UBE4B, KRT74, CYR61, LY6G6C, MT2A and SERPINB4. The most interesting observation was that MFGE8 (lactadherin) was the fourth most downregulated gene in group mfge8-/-/gRNA3 labelled with iTRAQ label 117 (Fig. 5D). Majority of DEPs were found to be associated with late endosome membrane, regulation of hydrolase activity, endopeptidase activity, phosphatidyl biosynthetic process, labyrinthine layer morphogenesis and intermediate filament while some DEPs were involved in epithelial cell apoptotic process, cytoskeletal organization, complement activation, ubiquitin conjugating enzyme activity and calcium dependent protein binding pathways etc.
Top signifcantly upregulated or downregulated proteins from mfge8-/-/gRNA3 cells relative to control BuMEC are summarized in Supplementary Material 3 S2. Hierarchial clustering analysis was performed to obtain an overview of the expression profile of DEGs in control and mfge8-/-/gRNA3 BuMEC (Fig. 5C).
Functional classification annotation analysis of differentially expressed proteins
To further scrutinize, Gene Ontology classification clusters based on biological processes, cellular components, and molecular functioning were generated from DEPs by GO analysis of mfge8-/-/gRNA3 cells. Moreover, pathway pathway analyses were performed using ClueGo of Cytoscape to attain an in-depth understanding of DEGs in mfge8-/-/gRNA3 cells. GO terms for cellular component involved spindle microtubule (11.11%), neuron projection cytoplasm (11.11%), late endosome membrane (33.33%), intermediate filament (33.33%) and actin filament (11.11%) (Fig. 6B). In terms of biological process, intermediate filament cytoskeleton organization (6.25%), positive regulation of apoptotic signaling pathway (6.25%), regulation of endopeptidase activity (25.0%), labyrinthine layer morphogenesis (18.75%), phosphatidylcholine biosynthetic process (18.75%) and complement activation (18.75%) were the major GO terms that were enriched (Fig. 6A). Furthermore, molecular function involved negative regulation of hydrolase activity (47.62%), ubiquitin conjugating enzyme activity (9.52%), transferrase activity transferring alkyl or aryl (other than methyl) groups (4.76%), intermediate filament binding (4.76%), scaffold protein binding (4.76%), monocarboxylic acid binding (9.52%), calcium- dependent protein binding (4.76%), regulation of endopeptidase activity (14.29%) and negative regulation of hydrolase activity (47.62%) GO terms to be significantly enriched (Fig. 6C).
Validation of proteomics data with real-time PCR
The mRNA expression levels of some differentially expressed proteins from quantitative proteomics data viz. FABP5, MT2A, SERPINH1, APOA2, MFGE8, AFAP, FABP3, FABP4 and PBK were also validated by quantitative PCR. The mRNA expression levels of FABP5 and APOA2 genes were significantly (P < 0.01) upregulated in mfge8-/-/gRNA3 BuMEC as compared to control cells while mRNA abundance of MT2A, MFGE8 and SERPINH1 genes was significantly (P < 0.01) downregulated in mfge8-/-/gRNA3 BuMEC as compared to control cells (Fig. 7). mRNA expression profiles of these genes was in correspondence to their expression pattern at proteomic level as observed in quantitative proteomics dataset. However, AFAP, FABP4 and PBK were differentially upregulated at proteome level, their transcriptome profile didn’t correlate with the proteomics data. Similarly, FABP3 was differentially downregulated as per proteomics data, on the contrary, its mRNA abundance was found high in control cells as compared to mfge8-/-/gRNA3 BuMEC. This could be due to different regulatory mechanisms at mRNA and protein expression levels.

Strategy for generation of MFGE8 gene KO BuMEC based on CRISPR/Cas9-system and experimental design for quantitative proteomics.

CRISPR-cas9 mediated generation of MFGE8 KO BuMEC. (A) Sequence information of three gRNAs targeting early exonic regions of MFGE8 gene and plasmid pSpCas9(BB)-2 A-Puro (PX459) from Dr. Feng Zhang’s lab containing Cas9 from S. pyogenes with 2 A-Puro, and cloning backbone for gRNA. (B) Confirmation of ligation of gRNAs into vector backbone by sequencing. (C) Lipofection of vector construct containing gRNAs into BuMEC and subsequent puromycin selection. magnification 200X. (D) PCR based Guide-it Mutation Detection kit assay. Lane M: 100 bp ladder, lane 1: gRNA3- 716 bp, 506 bp, 210 bp; lane 2: gRNA2- 742 bp, 527 bp, 215 bp; lane 3: gRNA4- 657 bp, 378 bp, 279 bp; lane 4: Genomic PCR analysis from Control BuMEC.

Validation using different biological assays. (A) Relative mRNA expression of MFGE8 gene in MFGE8 KO BuMEC generated using three different gRNAs and control BuMEC. (B) Western blot analysis of MFGE8 protein in control and BuMEC. ß-actin (equivalent protein taken as control). (C) Relative mRNA expression of Cas9 genes in mfge8-/-/gRNA3 BuMEC generated using three different gRNAs and control BuMEC. (D) Comparison of relative cell viability of control and mfge8-/-/gRNA3 BuMEC at 12 h time interval from 12 h to 72 h. (E) Comparison of relative cell proliferation of control and KO BuMEC at 72 h. Values are presented as mean ± SEM. All experiments were performed in triplicate (n = 3), and significant differences were confirmed statistically (*p < 0.05, **p < 0.01, ***p < 0.001).

NGS amplicon sequencing of gRNA3 transfected cells. (A) Target fragment amplification of MFGE8 gene. Lane M: 100 bp ladder; Lane 1: Genomic PCR product from control BuMEC (314 bp); Lane2: Genomic PCR product from MFGE8 KO BuMEC (314 bp). (B) Reads from NGS showed indels 3–4 bp upstream to PAM site (AGG-107 bp) in 2nd exon. (C) A 54% of total reads showed indels. (D) The observed indels occurred over a range of mutation sizes of 1–33 bp, 3–4 bp upstream to PAM site.

(A) Protein sequence coverage of the detected proteins in mfge8-/-/gRNA3 (117), BuMEC. (B) Scatter plot of changes in protein abundance in mfge8-/-/gRNA2 (115), mfge8-/-/gRNA3 (117) and mfge8-/-/gRNA4 (116) and control BuMEC. (C) Heat Map presentation of hierarchically clustered differentially expressed proteins in control and mfge8-/-/gRNA3 BuMEC. Red colour depicts high while blue colour depicts low expression levels. (D) Relative expression of MFGE8 protein in mfge8 knock-out BuMEC generated using three different gRNAs and control BuMEC. (E) Venn diagram representing the distribution of proteins in control and mfge8 knock-out cells. Peptides were labelled with iTRAQ labels 114- control, 115- mfge8-/-/gRNA2, 116- mfge8-/-/gRNA4 and 117- mfge8-/-/gRNA3. Control BuMEC, mfge8-/-/gRNA2, mfge8-/-/gRNA3 and mfge8-/-/gRNA4 yielded a total of 4276, 4278, 4282 and 4275 proteins respectively, among these, 4267, 4272 and 4270 proteins were shared by knock-out cells with control cells respectively. F) In- depth proteome analysis of mfge8-/-/gRNA3 and control BuMEC, helped in prediction of localization of identified proteins in cellular compartments of BuMEC.

Gene Ontology (GO) annotation of differentially expressed proteins in Pathway–pathway interaction analysis. (A) Biological Function (B) Cellular Component (C) Molecular Functions. Node size depicts the number of genes mapped to the function and node colour denotes the statistical significance.

Validation of quantitative proteomics data by quantitative real time PCR. Relative mRNA expression levels of some differentially expressed proteins viz. FABP5, MT2A, SERPINH1, APOA2, MFGE8 examined by qPCR in mfge8-/-/gRNA3 and control BuMEC. Values are mean ± SEM. All experiments were performed in triplicate (n = 3) and significant differences were confirmed statistically (*p < 0.05, **p < 0.01, ***p < 0.001).
Discussion
This study explored the role of MFGE8 in the regulation of lactation in buffalo mammary epithelial cells (BuMECs) and also analyzed the expression dynamics of associated proteins to identify the complex milk signalling pathways affected by MFGE8 knock down. MFGE8 is a multifunctional protein and its role in mammary gland development and phagocytic clearance of apoptotic cells via αVβ3/5 integrins signalling are well known. Besides these, it is also associated with anti-inflammatory activities, tumorigenesis, vascular modeling and osteoclastogenesis5,19,27. A previous study has shown that MFGE8 is upregulated in mammary epithelial cells isolated from high milk producing Sahiwal cows and suggested that it is a potential marker for high milk yield25. However, one of the most fundamental methods for establishing the importance and function of genes is through loss-of-function studies28 and CRISPR/Cas9 based genome-editing is being widely used as a quintessential technique to generate knock-outs29,30. Thereby, MFGE8 knock-out cells were generated using CRISPR-cas9 technology in the present study to study the role of MFGE8 in lactation biology.
We used plasmid-based approach for generating MFGE8 gene knock-out BuMEC as this method offers several advantages over RNP based delivery method31. Firstly, a huge number of cheap and highly stable CRISPR plasmids with suitable drug-resistance and fluorescent markers are available that can be easily modified for desired gene knock-out and knock-in experiments32. Secondly, these provide robust efficiencies by allowing transient and safe delivery into various cell types30. In the present study, we used pSpCas9 (BB)-2 A-puro expression vector which was deliberated to stimulate nonhomologous end joining to repair double strand breaks and produce the insertions or deletions (indels) at the target site regions. We designed three guide RNAs for creating MFGE8 gene knock-out BuMEC by targeting the early exons and the functional domains of MFGE8 gene.
Multiple lipofections, continuous puromycin selection and low density seeding led to selection of transfected cells with indels in MFGE8 gene. As evident from mutation detection assay and q-PCR results, all three gRNAs were capable of efficiently editing the target gene. However, gRNA 3 was found to be more efficient than other two gRNAs and NGS amplicon sequencing results further showed that the gRNA 3 induced indels that led to frame-shifting and created functional knock-outs. In a previous study, authors have also suggested that designing and targeting gRNAs from exons that encode functional domains of protein more efficiently ablates gene function than targeting early exons28. Consistent with the mRNA expression analysis, the western blot analysis showed negligible expression levels of MFGE8 protein in MFGE8 knock-out BuMEC than in control cells. The transfected cells in the present study showed distorted morphology and a lower growth rate as compared to control cells as validated by MTT, BrDU assays, which may be attributed to dysfunction of MFGE8 gene, which was in line with previous studies26,33.
Comprehensive iTRAQ based quantitative proteomics analysis was performed between MFGE8 knock-out and wild type BuMEC to unravel the possible proteins involved and associated lactation signalling pathways. Our proteomics data revealed that MFGE8 dysfunction leads to proteome changes in several biological processes. We found 178 differentially expressed proteins (DEPs), out of these, 124 proteins were significantly upregulated (cutoff ≥ 1.5 fold) while 54 proteins were downregulated (cut off ≤ 0.6 fold). The identified DEPs are associated with phosphatidyl biosynthetic process, labyrinthine layer morphogenesis, epithelial cell apoptotic process, intermediate filament and cytoskeletal organization, regulation of endopeptidase activity, complement activation, negative regulation of hydrolase activity, ubiquitin conjugating enzyme activity and calcium dependent protein binding pathways etc. Interestingly, several DEPs belonged to genes that have been reported as markers associated with milk production trait including FABP3, FABP4, FABP5, KNG1, MT2A, CD82, SLC7A1 and SERPINH134,35,36. FABP3 and MT2A were downregulated and FABP4, FABP5, KNG1, CD82, SLC7A1and SERPINH1 were upregulated.
MFGE8 knock-out strongly affected cytoskeleton organization in buffalo mammary epithelial cells in our study which was in line with our previous study which reports that suppression of MFGE8 leads to reorganization of cytoskeleton26. Many proteins such as Actin-related protein 2/3 complex subunit 5-like protein (ARPC5L), Actin, aortic smooth muscle (ACTA2), Actin filament-associated protein 1-like 1 (AFAP1L1) and Vimentin (VIM) were found to be up regulated while Keratin, type II cytoskeletal 72 (KRT72), Keratin, type II cytoskeletal 74 (KRT74) and Keratin, type II cytoskeletal 79 (KRT79) were found to be down regulated. Cytoskeletal proteins are abundantly present in mammary epithelial cells and are also differentially expressed during lactation stages25. Reorganization of the intermediate filaments (vimentin, keratins) and microfilaments (actin) is highly required for the complex cell polarization process and milk fat droplet transportation37. Interestingly, our data shows that actin filament-associated protein 1 (AFAP1) is highly upregulated and a previous study reports that AFAP1 knock-out mice have a lactation deficiency marked by a concurrent decrease in lipid production and a decreased expression of several lipogenic genes38. This might indicate a compensatory role of AFAP1 in MFGE8 knock out cells, restoring lipogenesis. This claim is supported by increased expression of several lipogenic genes like FABP4, FABP5, ApoA1 and ApoA2, the only exception was the reduced expression of FABP3 gene. Even though we observed significant upregulation of most of the lipogenic genes, lipid droplet secretion could be affected due to the negative regulation of hydrolase function. The genes ApoA1 and ApoA2 associated with the function were found to be significantly upregulated. Some previous studies have reported that MFGE8 regulates the absorption of dietary fat by enterocytes through binding of the αvβ3 and αvβ5 integrins39. Authors have further reported that MFGE8 and the αvβ3 and αvβ5 integrins coordinate to activate a PI3 kinase/mTORC2/PKCζ–dependent pathway that leads to increased intracellular Triglyceride (TG) hydrolase activity. Thus, it promotes enterocyte hydrolysis of cytoplasmic lipid droplets and the fatty acids released may be utilized for chylomicron production40. Impaired TG hydrolase activity leads to accumulation of cytoplasmic lipid droplets which may inhibit release of milk fat globules (MFGs) into the lumina of mammary alveoli that may be associated with reduction of milk lipids41. From our in vitro study, we found that hydrolase activity is negatively regulated due to knock-out of MFGE8 gene, which in turn reduces the secretion of milk fat droplets. However, an in vivo study is further required to reinforce this claim.
In the present study, protein levels of FABP3 gene reduced drastically in MFGE8 knock-out cells while expression of proteins FABP4 and FABP5 have been found markedly higher in knock-out cells. Role of mammary gland epithelial cells in the biosynthesis of milk fat in buffalo has been well established and some previous studies have reported remarkably high expression of several FABPs in bovine mammary gland tissue during lactation42,43. These proteins affect the trafficking of long-chain fatty acids (LCFAs), regulates lipid homeostasis in different tissues and also play role in inflammatory responses and tumor cells progression44,45. In dairy cattle, polymorphism in FABP genes has been associated with milk traits and one SNP has been reported as a potential marker for evaluating milk performance46. FABP3 is the most abundant protein among FABP family proteins and regulates milk fat synthesis by altering the expression of sterol response element-binding protein1 (SREBP1) and nuclear receptor peroxisome proliferator-activated receptor receptor (PPARg) in bovine mammary epithelial cells36,47. FABP4 has been mapped on a location on chromosome 14 which is rich in quantitative trait loci for milk traits in cows and thus has been considered a key candidate that affects milk yield and milk protein content48. FABP5 functions by the transcriptional activation of the PPAR β/δ receptor and is involved in tumour development, cell growth and survival49. Differential expression of FABP5 protein in mammary tissues during the dry, puberty and lactating period suggests its role in mammary gland development and lactation biology50. Significant modulation of expression of FABP family proteins in MFGE8 knock-out cells in the present study suggests, key association of MFGE8 gene with mammary gland development and lactation signaling pathways. However, the possibility of involvement of several other genes cannot be denied, the contention that MFGE8 and FABP family genes are involved in biosynthesis of milk, is worthy of further investigation.
Moreover, key GO term showing significant enrichment in the biological process, phosphatidylcholine biosynthetic process, is also intricately linked to the biosynthesis of milk fat51. Particularly, the active involvement of phosphatidylcholine in this pathway has garnered attention, as evidenced by its higher ratios to phosphatidylethanolamine in milk from cows with smaller MFGs52. Variation in MFGs size underscores the importance of phosphatidylcholine in modulating the composition of milk lipids. Phosphatidyl choline biosynthetic pathway is thus essential for the coordinated control of cellular mechanisms involved in milk production53, highlighting its significance in the overall process of milk fat synthesis and secretion.
The S100 proteins are some of the prolific bioactive molecules involved in numerous physiological events such as calcium homeostasis, differentiation, apoptosis, antimicrobial activity, proliferation, inflammation54. Interestingly, we observed increased expression of S100 proteins (S100 A9, S100A2, S100A12 and S100A4) in our data in association with mfge8-/-/gRNA3 cell line. The increased expression of S100 proteins is intriguing as not single but four isoforms of S100 proteins were identified with increased fold change. Out of four S100 proteins, S100A9 has the highest value of fold change (3.32). Several studies have shown the pro-inflammatory function of S100A9 and S100A12 55,56,57,58. The S100 proteins are closely involved in immunomodulation; however, their involvement in milk production trait is least explored. We found only two reports associating the S100 proteins with the milk yielding traits of the dairy cow. The increased expression of S100 proteins in low milk yielding animals is associated with the inhibition of casein kinase, which in turn assist in cell survival, and continuance of milk synthesis and secretion59. Further, the previous work carried out in our lab showed that S100 proteins were upregulated in low yielding animals. The increased expression of S100 in knock out cells indicates that milk production potential has been severely compromised.
Gene SLC7A1 was found significantly upregulated (2.3 folds) in mfge8-/-/gRNA3 cells in our study. The gene encodes cationic amino acid transporter 1 (CAT-1) protein which is involved in the transport of amino acids across cell membranes. Authors have previously reported a significant upregulation in mammary amino acid transporters in heat stress-induced cows and SLC7A1 gene is also one among them60. This coordinated increase in the expression of genes involved in amino acid transporters, particularly during early and peak lactation, suggests a potential compensatory mechanism to enhance the availability of amino acids within mammary tissue, thereby promoting milk protein synthesis and cellular growth61. The orchestrated upregulation in SLC7A1 gene observed in our study suggests a potential new connection between MFGE8, SLC7A1 and milk production. However, additional research is necessary to uncover the complex molecular pathways underlying this relationship.
The CD82 gene, also known as the Tetraspanin 27 (TSPAN27) gene, encodes a protein called tetraspanin-28 or CD82 and exhibits multifaceted roles in various physiological and pathological processes, including cancer62. It serves as a suppressor of metastasis63. A previous study reported that CD82 knock out in human oral squamous cell carcinoma (OSCC) cell lines resulted in the significant downregulation of S100 proteins64. They further claimed that CD82 shows a potential correlation with and might govern the expression of various S100 family proteins, such as S100A7, S100A7A, S100A6, S100A8, and S100A9, in both OSCC cell lines and tissue. In our study, we have also observed significant upregulation of CD82 in mfge8-/-/gRNA3 cells and a concurrent increase in S100 expression, suggests a potential interplay between MFGE8 and CD82 in modulating S100 protein levels. Moreover, high expression of MFG-E8 has been reported in various types of cancer in humans including OSCC65,66. This further indicates that a connection might exist between MFGE8 and CD82, which carries implications for the pathogenesis of oral cancer and potential therapeutic approaches.
The cell viability assays showed the reduced viability in the MFGE8 knockout cells. However, the reduced viability trend observed even after 72 h of transfection when transfected cells has attained the normal morphology and adapted after knockout experiment. The gene ontology of differentially expressed proteins showed enrichment of positive regulation of apoptotic signaling pathway (6.25%) and the DEPs IRF3, VIM, XIAP, CTSH, CAPNS1, CDKN2A, APP, PTGS, TUBA4A, HDAC1, and ACTB from all the three MFGE8 knock out cell lines in our study are implicated in apoptotic signaling pathways associated with MFGE8. This suggests a strong correlation between the knockout of the MFGE8 gene and the induction of apoptosis. This finding aligns with previous studies that have reported a significant increase in apoptotic cells in MFGE8 knockout or knockdown cell lines. These cell lines were mostly carcinomas where knockout or silencing of MFGE8 gene effectively reduced the tumor progression65,67,68. Its role as an anti-apoptotic factor is established in other cell lines as well69,70,71,72. Besides influencing apoptosis, the MFGE8 has key role in apoptotic clearances10,15,73,74. The apoptotic clearances is necessary event in the remodeling of mammary gland during involution process. MFGE8 has significant contribution in the remodeling of mammary gland at the time of involution. The MFGE8 null mice mammary gland showed increased number of apoptotic cells during early involution, which led to the eventual development of inflammation of mammary gland24. A different study pointed out the same results in mice with additional observation of duct ectasia with periductal mastitis and the impaired redevelopment of mammary gland for the second litter of the mice75. The apoptotic clearances are essential during the changes in mammary gland landscapes. Considering the paramount role of MFGE8 in the apoptotic clearance, the complete absence of it can create hurdle during transition into different stages of lactation as this stage transitions requires remodeling of mammary gland.
GO term, neuron projection cytoplasm was also found to be significantly enriched in cellular component in mfge8-/-/gRNA3 cells suggesting some association between MFGE8 and neurogenesis. In alignment with our findings, a previous study has also reported MFGE8 as a neural-stem-cell-enriched factor4. Specifically, this study elucidates the role of MFGE8 in maintaining quiescence and preventing the developmental exhaustion of neural stem cells, thereby sustaining continuous neurogenesis. This finding suggests that MFGE8 may serve as a niche factor that supports the quiescent state of neural stem cells, potentially by providing cues or signals that promote quiescence and inhibit activation via mTOR1 signaling pathway. Furthermore, the study demonstrates that MFGE8 deletion leads to the depletion of neural stem cells and a decrease in adult neurogenesis, emphasizing the critical role of MFGE8 in maintaining the neural stem cell pool and promoting neurogenesis in the adult brain.
Some pathways like ubiquitin-conjugating enzyme activity and endopeptidase activity were also significantly changed. Genes encoding Ubiquitin-conjugating enzyme E2 G1 (UBE2G1), Ubiquitin-conjugating enzyme E2 (UBE2S) and Ubiquitin-conjugating enzyme E2 H (UBE2H) were all upregulated while some genes involved in endopeptidase activity like Pro-cathepsin (CTSH), FAM162A, Legumain (LGMN), KNG1 were upregulated and one gene Cysteine-rich, angiogenic inducer, 61 (CYR61) was downregulated. No direct co-relation was found among these genes with MFGE8 knock-out. The transcriptomic levels of some differentially expressed proteins were also validated by quantitative PCR. mRNA expression levels of FABP5, MT2A, SERPINH1 and APOA2 genes were similar to the expression profile at their protein level while transcriptome profile of some genes including AFAP, FABP3, FABP4 and PBK didn’t correlate with their proteomics expression profile. This could be due to various regulatory mechanisms at transcriptomic and proteomic levels.
Materials and methods
Designing of gRNAs and plasmid construction
Bicistronic expression vector (pSpCas9 (BB)-2 A-Puro) (PX459) V2.0 was a gift from Feng Zhang of Broad Institute of MIT and Harvard76. The plasmid contains a sgRNA scaffold, Cas9 gene and puromycin resistance gene. Three gRNAs and their deletion primers (Table 1) were designed from early exons (2nd and 3rd) of MFGE8 gene using CRISPOR (http://crispor.tefor.net/). For cloning gRNAs into the vector, complementary oligo pairs containing BbsI restriction enzyme site overhangs were synthesized, phosphorylated and annealed. These phosphorylated and annealed oligos were ligated into the oligo cloning site before the gRNA scaffold into BbsI (New England Biolabs, Beverly, MA, USA) digested pSpCas9 (BB)-2 A-puro vector. Recombinant clones were screened by PCR using U6 F (sequence from vector) as forward primer and gRNA reverse as reverse primer and validated by sanger sequencing.
Cell culture and transfection of MFGE8 specific CRISPR/ Cas9 vector into BuMEC
The BuMEC was previously established in our lab at Animal Biotechnology center of ICAR-NDRI, Karnal [ExPASy accession: Buffalo 2012(CVCL_M445)] and was used for the transfection purpose77. Briefly, cells were cultured in DMEM/F12 media (Gibco BRL, United States) supplemented with 10% FBS (Gibco BRL, United States), 5 mg/ml bovine insulin (Sigma, United States), 1 mg/ml hydrocortisone (Sigma, United States), 10 ng/ml EGF (Sigma, United States), 100 U/ml penicillin, and 5 mg/ml streptomycin (Sigma, United States) and were incubated with 5% CO2 at 37ºC and used at 70–80% confluency for transfection. The pSpCas9 (BB)-2 A-puro expression vectors containing gRNAs against MFGE8 gene (6ug) were transfected three times by lipofectamine 3000 (Invitrogen Thermo Fisher Scientific, Waltham, MA, United States) into BuMEC. After 48 h of transfection, cells were subjected to puromycin selection (2ug/ml) (Invitrogen Thermo Fisher Scientific, Waltham, MA, United States) and the selected cells were maintained in complete media containing 1 ug/ml puromycin for 2–3 weeks. The cells were then trypsinized and seeded at low density in 96-well cell culture plates in complete medium containing 1 ug/ml puromycin and further cultured for cryopreservation and DNA isolation for detection of mutations.
Transfected cells were screened for mutation detection by amplicon sequencing. For the same, a 314 bp fragment spanning the target region was amplified from DNA isolated from transfected and control (non-transfected) cells using specific deletion primers via PCR cycling conditions: initial denaturation at 95oC for 3 min, followed by 32 cycles of 95oC for 30 s, 60oC for 30 s, 72oC for 30 s, and a final extension at 72oC for 10 min. The PCR products were run on gel, excised and purified by the Gene JET PCR Purification kit (Thermo Fisher Scientific, Waltham, MA, United States) and sequenced by Next generation sequencing for assessing the targeted genome editing in MFGE8 gene in transfected cells.
Gene expression analysis by qPCR
RNA was isolated from transfected and nontransfected cells using TRIzol reagent (Sigma, United States). Subsequently, cDNA was synthesized using superscript reverse transcriptase III (Invitrogen, USA). Quantitative real-time PCR (qPCR) was done using CFX96 C1000 Thermal Cycler (Bio-Rad, Hercules, CA) for MFGE8 and cas9 genes. The reaction mixture (10 µL) contained 5 µL of SYBR Green Master Mix, 0.2 µL of 10µM of each primer (Table 2), and cDNA was obtained from both cell types. Thermal cycling conditions for the genes consisted of initial denaturation for 3 min at 95ºC, followed by 40 cycles of 10 s at 95ºC, 58 ºC annealing temperature for 15 s and 15 s at 72ºC, followed by final extension of 15 s at 72ºC. Relative gene expression was determined using 2–ΔΔCT method, where DCt = Ct (target gene)—Ct (internal reference) and DDCt = DCt sample—DCt calibrator. Control BuMEC served as calibrator and GAPDH was taken as an internal reference gene. All experiments were executed in triplicates.
MFGE8 expression analysis by Western blotting
BuMEC transfected by gRNA3 and control BuMEC grown at 75–80% confluence, were washed three times with PBS and trypsinized and the isolated cell pellet was suspended in 2% SDS in water. The obtained mixtures were sonicated (30% amplitude, 0.5 cycle for 10s; 4 times) for maximum protein output from cells. Sonicated samples were centrifuged at 12,000 g to remove undisrupted cells and supernatant was taken in fresh tube. Proteins were precipitated by adding 4 volumes of cold acetone followed by overnight incubation at 200 °C and the precipitated proteins were collected by centrifugation at 12,000 g for 20 min. The Protein estimation was performed using bradford reagent (Sigma, United States) as per manufacturer’s instructions. 10 µg protein from each group was loaded on the 12% SDS-PAGE and resolved using a Mini VE complete gel electrophoresis system (GE Healthcare, Sigma, USA). Proteins were transferred to the polyvinylidene difluoride (PVDF) membrane using a Trans-blot apparatus (GE, USA). After the protein transfer to PVDF membrane, the membranes were cut into different lanes (with the reference of marker and comb) prior to processing with antibodies. The membranes were subsequently blocked with 5% skimmed milk powder (Sigma, USA) overnight at 4 °C. They then were incubated with primary antibodies of MFGE8 (Santa Cruz Biotechnology, California) (1:1000) and beta-actin (Sigma, USA) (1:1000) for 2 h at RT with gentle agitation. The membranes were further washed thoroughly with TBST (3 × 15 min) and then incubated with peroxidase conjugated goat anti-rabbit (1:40000) and anti-mouse (1:40000) secondary antibodies, respectively for 1 h at RT. Each membrane was washed extensively with TBST (8 × 15 min) followed by their development using enhanced chemiluminescent substrate (Bio-Rad laboratories, US). The protein bands were detected with X-ray film.
Cell viability and proliferation assay for comparative analysis of MFGE8 Knock-out and control BuMEC
The relative cell viability of Knock-out and control BuMEC at 12 h time intervals from 12 to 72 h (12, 24, 32, 48, 60, and 72 h) posttransfection was determined by MTT [3-(4, 5-di-methylthiazol-2-yl) 2, 5- diphenyl tetrazolium bromide] assay. Briefly, 1 × 104 cells were seeded in 96-well plates in complete media for 24 h at 37 °C and 5% CO2 concentration. After the incubation period, 10 µl of the MTT labeling reagent (final concentration 0.5 mg/ml) was added to each well and incubated for 4 h. Next, 100 ul dimethyl sulfoxide was added and the plate was shaken on an orbital shaker for 15 min. to dissolve formazan crystals completely. The optical density of the formazan product was measured at 570 nm using NanoQuant spectrophotometer (TECAN, Infinite 200 PRO, Mannedorf, Switzerland). For measuring cell proliferation among MFGE8 knock-out and control BuMEC, BrdU (5-bromo-20-deoxyuridine) assay was performed using kit (Roche Mannheim, Germany) according to the manufacturer’s instructions. In brief, 1 × 104 cells were seeded in 96-well cell culture plates in 100ul complete media and incubated for 24 h in a humidified environment at 37 °C. After incubation, 10 µl/ well BrdU labeling solutions were added and cells were incubated for 24 h. After removing culture medium, cells were fixed and denatured and the integrated BrdU was measured by taking absorbance at 370 nm using NanoQuant spectrophotometer. The experiments were performed in three trials and each with three replicates.
Quantitative proteome profiling in mfge8-/-/gRNA2, mfge8-/-/gRNA3, mfge8-/-/gRNA4 and control BuMEC
Protein was isolated from mfge8-/-/gRNA2, mfge8-/-/gRNA3, mfge8-/-/gRNA4 and control BuMEC following the protocol mentioned above. After protein estimation using Bradford reagent (Sigma, United States), 100 µg protein were taken from each group and reduced by freshly prepared 45 mM DTT (dissolved in 50 mM Ammonium Bicarbonate) for 1 h at 50 °C. The protein samples were alkylated by incubating in 10 mM Iodoacteamide for 1 h. The alkylation reaction was quenched by adding 45 mM DTT. Protein samples were digested by incubating with sequencing grade modified Trypsin (1:20 enzyme to protein ratio, (Mass Spec Grade, Promega, Madison, WI, United States) for 18 h at 37 °C. Trypsin reaction was quenched by 10% Trifluoroacetic acid. SDS-PAGE was run to confirm the digestion of protein samples. The iTRAQ labeling was performed as per the manufacturer’s instructions (AB Sciex, Marsh Rd., Foster City, CA, United States). Briefly, the sample peptides were incubated with labels for 2 h at RT (114- control, 115- mfge8-/-/gRNA2, 116- mfge8-/-/gRNA4 and 117- mfge8-/-/gRNA3). The labelled samples were pooled and fractionated using high pH reverse phase fractionation column (Pierce™ High pH Reversed-Phase Peptide Fractionation Kit, catalog: 84868) (fraction 1: 5% ACN, fraction 2: 10% ACN, fraction 3: 15% ACN, fraction 4: 20% ACN, fraction 5: 25% ACN, fraction 6: 35% ACN fraction 7: 50% ACN, fraction 8: 70% ACN). Fractions were dried using SpeedVac (Thermo savant ISS110 SpeedVac concentrator ISS110-230, USA) and were dissolved in Buffer A (2% ACN + 0.1% FA). All the experiments were performed using EASY-nLC 1200 system (Thermo Fisher Scientific) coupled to Thermo Fisher-QExactive equipped with nanoelectrospray ion source. 1.0 µg of the peptide mixture was loaded on C18 column 50 cm, 3.0 μm Easy-spray column (Thermo Fisher Scientific). The peptides were loaded with buffer A and eluted with a 0–40% gradient of buffer B (80% acetonitrile, 0.1% formic acid) at a flow rate of 300 nl/min for 100 min. MS data was obtained by using a data-dependent top10 method dynamically choosing the most abundant precursor ions from the survey scan.
Data processing
All samples were processed and 8 RAW files generated were analyzed with Proteome Discoverer (v2.2) against the Uniprot Bos_taurus 9913 reference proteome database. For Sequest search, the precursor and fragment mass tolerances were set at 10 ppm and 0.02 Da, respectively. The protease used to generate peptides, i.e. The maximum number of missed cleavages was set to two, and the enzyme specificity was set for trypsin/P (cleavage at the C terminus of “K/R: unless followed by “P”). iTRAQ4plex / +144.102 Da at “K” and Carbamidomethyl on cysteine as fixed modification for database search. Both peptide spectrum match and protein false discovery rate were set to 0.01 FDR.
Differentially expressed proteins (DEPs) were selected for further functional analysis. Upregulated proteins were selected with fold change ≥ 1.5 and downregulated proteins were selected with a fold change ≤ 0.6. To further decipher the biological interpretation of this large lists of DEPs, we employed Cytoscape plugin ClueGO. It integrates Gene Ontology (GO) terms as well as pathways and generates a functionally organized GO/pathway network. Significant pathway-pathway interaction networks with p < 0.05 were considered for further analysis. The node size denotes the number of genes mapped to a particular GO term (larger the node size, larger the number of genes mapped to the function) and the color denotes the significance of the function (darker color nodes depict the higher significance).
Statistical analysis
Data are shown as mean ± SEM. Differences in means of cell viability, proliferation rates and expression of MFGE8, cas9 and other genes in MFGE8 knock-out and control BuMEC were compared using one-way ANOVA. Statistical analysis was performed using GraphPad prism (version 8) software. The differences were considered to be significant at p < 0.05. Mass spectrometry results are represented as relative log2 fold change.
Conclusions
MFGE8 is an important protein in mammary gland tissue which has been described to be involved in lactation physiology in multiple ways. However, the functional significance of MFGE8 has not been proven vis a vis directly interacting molecular partners. In the present work, MFGE8 knockout BuMEC were generated by targeting the early exonic regions using CRISPR-cas9 gRNA cargos. The quantitative proteomic study of the knockout cell line showed perturbation in different cellular and molecular pathways. Post knockout the cells showed a distorted morphology and slow proliferation rate which may be associated with increase in apoptosis. Previous literature has discussed role of MFGE8 in reshaping of mammary gland architecture by apoptotic clearance as it goes through cyclical changes. The disturbance in remodelling events as a result of dysfunction of MFGE8 can consequently result in reduced milk synthesis. In addition, milk fat synthesis and secretion was found to be severely affected as genes like FABP4, FABP5, ApoA1 and ApoA2 were found to be upregulated and TG hydrolase pathway was negatively regulated. Knocking out MFGE8 results in increased level of S100 proteins which is related to low milk yield. Thus, MFGE8 functions can be considered very important particularly from view point of maintenance of cellular architecture, lipogenesis and negative regulation of hydrolase activity in Buffalo mammary epithelial cells. Since, lactation biology is complex series of events involving repeated cycles of proliferation, differentiation and involution under endocrine regulation, such knock out studies should be complemented with co culture experiments under hormonal regulation to mimic real physiology of lactation.
Data availability
The datasets generated and/or analysed during the current study are available in the [PRIDE] repository, [Project accession: PXD048149; Reviewer account details: Username: reviewer_pxd048149@ebi.ac.uk, Password: M3dGBTTh].
References
-
Oshima, K. et al. Lactation-dependent expression of an mRNA splice variant with an exon for a multiply O-glycosylated domain of mouse milk fat globule glycoprotein MFG-E8. Biochem. Biophys. Res. Commun. 254, 522–528 (1999).
-
Kusunoki, R. et al. Roles of milk fat Globule-Epidermal growth factor 8 in intestinal inflammation. Digestion 85, 103–107 (2012).
-
Wang, B. et al. MFGE8 is down-regulated in cardiac fibrosis and attenuates endothelial-mesenchymal transition through Smad2/3-Snail signalling pathway. J. Cell. Mol. Med. 24, 12799–12812 (2020).
-
Zhou, Y. et al. Autocrine Mfge8 signaling prevents developmental exhaustion of the adult neural stem cell pool. Cell. Stem Cell. 23, 444–452e4 (2018).
-
Raymond, A., Ensslin, M. A. & Shur, B. D. SED1/MFG-E8: a bi-motif protein that orchestrates diverse cellular interactions. J. Cell. Biochem. 106, 957–966 (2009).
-
Couto, J. R., Taylor, M. R., Godwin, S. G., Ceriani, R. L. & Peterson, J. A. Cloning and sequence analysis of human breast epithelial antigen BA46 reveals an RGD cell adhesion sequence presented on an epidermal growth factor-like domain. DNA Cell. Biol. 15, 281–286 (1996).
-
Hanayama, R. et al. Identification of a factor that links apoptotic cells to phagocytes. Nature 417, 182–187 (2002).
-
Reddy Nanga, R. P., Vivekanandan, S. & Yoon, H. S. Expression, purification and characterization of C2 domain of milk fat globule-EGF-factor 8-L. Protein Expr Purif. 52, 329–333 (2007).
-
Miksa, M. et al. Maturation-induced down-regulation of MFG-E8 impairs apoptotic cell clearance and enhances endotoxin response. Int. J. Mol. Med. 22, 743–748 (2008).
-
Hanayama, R. et al. Autoimmune disease and impaired uptake of apoptotic cells in MFG-E8-deficient mice. Science 304, 1147–1150 (2004).
-
Miksa, M. et al. Dendritic cell-derived exosomes containing milk fat globule epidermal growth factor-factor VIII attenuate Proinflammatory responses in sepsis. Shock 25, 586–593 (2006).
-
Yamaguchi, H. et al. Milk fat globule EGF factor 8 in the serum of human patients of systemic lupus erythematosus. J. Leukoc. Biol. 83, 1300–1307 (2008).
-
Chung, C. S., Xu, Y. X., Chaudry, I. H. & Ayala, A. Sepsis induces increased apoptosis in Lamina propria mononuclear cells which is associated with altered cytokine gene expression. J. Surg. Res. 77, 63–70 (1998).
-
Miksa, M. et al. Immature dendritic cell-derived exosomes rescue septic animals via milk fat globule epidermal growth factor-factor VIII [corrected]. J. Immunol. 183, 5983–5990 (2009).
-
Ait-Oufella, H. et al. Lactadherin deficiency leads to apoptotic cell accumulation and accelerated atherosclerosis in mice. Circulation 115, 2168–2177 (2007).
-
Albus, E. et al. Milk fat Globule-Epidermal growth factor 8 (MFG-E8) is a novel Anti-inflammatory factor in rheumatoid arthritis in mice and humans. J. Bone Min. Res. 31, 596–605 (2016).
-
Sinningen, K. et al. Loss of milk fat globule-epidermal growth factor 8 (MFG-E8) in mice leads to low bone mass and accelerates ovariectomy-associated bone loss by increasing osteoclastogenesis. Bone 76, 107–114 (2015).
-
Zhao, Q. et al. MFG-E8 overexpression promotes colorectal cancer progression via AKT/MMPs signalling. Tumour Biol. 39, 1010428317707881 (2017).
-
Ko, D. S. et al. Milk fat Globule-EGF factor 8 contributes to progression of hepatocellular carcinoma. Cancers (Basel). 12, E403 (2020).
-
Neutzner, M. et al. MFG-E8/lactadherin promotes tumor growth in an angiogenesis-dependent Transgenic mouse model of multistage carcinogenesis. Cancer Res. 67, 6777–6785 (2007).
-
Ensslin, M. A. & Shur, B. D. Identification of mouse sperm SED1, a bimotif EGF repeat and Discoidin-Domain protein involved in sperm-Egg binding. Cell 114, 405–417 (2003).
-
Trigg, N. A. et al. A novel role for milk fat globule-EGF factor 8 protein (MFGE8) in the mediation of mouse sperm-extracellular vesicle interactions. Proteomics 21, e2000079 (2021).
-
Atabai, K. et al. Mfge8 diminishes the severity of tissue fibrosis in mice by binding and targeting collagen for uptake by macrophages. J. Clin. Invest. 119, 3713–3722 (2009).
-
Atabai, K. et al. Mfge8 is critical for mammary gland remodeling during Involution. Mol. Biol. Cell. 16, 5528–5537 (2005).
-
Janjanam, J. et al. Comparative 2D-DIGE proteomic analysis of bovine mammary epithelial cells during lactation reveals protein signatures for lactation persistency and milk yield. PLOS ONE. 9, e102515 (2014).
-
Verma, A. K., Ali, S. A., Singh, P., Kumar, S. & Mohanty, A. K. Transcriptional repression of MFG-E8 causes disturbance in the homeostasis of cell cycle through DOCK/ZP4/STAT signaling in Buffalo mammary epithelial cells. Front. Cell. Dev. Biology 9, (2021).
-
Hajishengallis, G. MFG-E8, a novel homeostatic regulator of osteoclastogenesis. Inflamm. Cell. Signal. 1, e285 (2014).
-
Shi, J. et al. Discovery of cancer drug targets by CRISPR-Cas9 screening of protein domains. Nat. Biotechnol. 33, 661–667 (2015).
-
Mandl, M. et al. Sprouty1 prevents cellular senescence maintaining proliferation and differentiation capacity of human adipose stem/progenitor cells. J. Gerontol. Biol. Sci. Med. Sci. 75, 2308–2319 (2020).
-
Mehrabian, M. et al. CRISPR-Cas9-based knockout of the prion protein and its effect on the proteome. PLoS One. 9, e114594 (2014).
-
Kamali, E., Rahbarizadeh, F., Hojati, Z. & Frödin, M. CRISPR/Cas9-mediated knockout of clinically relevant alloantigenes in human primary T cells. BMC Biotechnol. 21, 9 (2021).
-
Giuliano, C. J., Lin, A., Girish, V. & Sheltzer, J. M. Generating single Cell-Derived knockout clones in mammalian cells with CRISPR/Cas9. Curr. Protoc. Mol. Biol. 128, e100 (2019).
-
Carrascosa, C. et al. MFG-E8/lactadherin regulates cyclins D1/D3 expression and enhances the tumorigenic potential of mammary epithelial cells. Oncogene 31, 1521–1532 (2012).
-
Dado-Senn, B. et al. RNA-Seq reveals novel genes and pathways involved in bovine mammary Involution during the dry period and under environmental heat stress. Sci. Rep. 8, 11096 (2018).
-
Ma, Y. et al. Genetic markers associated with milk production traits in dairy cattle. Agriculture 11, 1018 (2021).
-
Mu, T. et al. Regulation of key genes for milk fat synthesis in ruminants. Front. Nutr. 8, 765147 (2021).
-
Lu, J. et al. The protein and lipid composition of the membrane of milk fat globules depends on their size. J. Dairy. Sci. 99, 4726–4738 (2016).
-
Cunnick, J. M. et al. Actin filament-associated protein 1 is required for cSrc activity and secretory activation in the lactating mammary gland. Oncogene 34, 2640–2649 (2015).
-
Khalifeh-Soltani, A. et al. Mfge8 promotes obesity by mediating the uptake of dietary fats and serum fatty acids. Nat. Med. 20, 175–183 (2014).
-
Khalifeh-Soltani, A. et al. Mfge8 regulates enterocyte lipid storage by promoting enterocyte triglyceride hydrolase activity. JCI Insight. 1, e87418 (2016).
-
Mather, I. & Keenan, T. Origin and secretion of milk lipids. J. Mammary Gland Biol. Neoplasia. 3, 259–273 (1998).
-
Bionaz, M. & Loor, J. J. ACSL1, AGPAT6, FABP3, LPIN1, and SLC27A6 are the most abundant isoforms in bovine mammary tissue and their expression is affected by stage of lactation. J. Nutr. 138, 1019–1024 (2008).
-
Nafikov, R. A. et al. Association of polymorphisms in solute carrier family 27, isoform A6 (SLC27A6) and fatty acid-binding protein-3 and fatty acid-binding protein-4 (FABP3 and FABP4) with fatty acid composition of bovine milk. J. Dairy. Sci. 96, 6007–6021 (2013).
-
Wang, W. et al. FABP5 correlates with poor prognosis and promotes tumor cell growth and metastasis in cervical cancer. Tumour Biol. 37, 14873–14883 (2016).
-
Zhang, Y. et al. Tracing the evolution of fatty acid-binding proteins (FABPs) in organisms with a heterogeneous fat distribution. FEBS Open. Bio. 10, 861–872 (2020).
-
Ye, T. et al. Evolutionary and association analysis of Buffalo FABP family genes reveal their potential role in milk performance. Genes 13, 600 (2022).
-
Liang, X. et al. Rapid and highly efficient mammalian cell engineering via Cas9 protein transfection. J. Biotechnol. 208, 44–53 (2015).
-
Zhou, H. et al. Variation in the bovine FABP4 gene affects milk yield and milk protein content in dairy cows. Sci. Rep. 5, 10023 (2015).
-
Kannan-Thulasiraman, P., Seachrist, D. D., Mahabeleshwar, G. H., Jain, M. K. & Noy, N. Fatty acid-binding protein 5 and PPARbeta/delta are critical mediators of epidermal growth factor receptor-induced carcinoma cell growth. J. Biol. Chem. 285, 19106–19115 (2010).
-
Li, P. et al. FABP5 is a critical regulator of methionine- and estrogen-induced SREBP-1c gene expression in bovine mammary epithelial cells. J. Cell. Physiol. 234, 537–549 (2018).
-
Kinsella, J. E. & McCarthy, R. D. Biosynthesis of secretory lipids from [2-14 C]acetate by bovine mammary cells in vitro. Biochim. Et Biophys. Acta (BBA) – Lipids Lipid Metabolism. 164, 518–529 (1968).
-
Walter, L. et al. Milk fat globule size development in the mammary epithelial cell: a potential role for ether phosphatidylethanolamine. Sci. Rep. 10, 12299 (2020).
-
Smoczyński, M. Role of phospholipid flux during milk secretion in the mammary gland. J. Mammary Gland Biol. Neoplasia. 22, 117–129 (2017).
-
Donato, R. et al. Functions of S100 proteins. Curr. Mol. Med. 13, 24–57 (2013).
-
Hessian, P. et al. NMRP-8 and MRP-14, two abundant Ca(2+)-binding proteins of neutrophils and monocytes. J Leukoc Biol 53:197–204. J. leukocyte Biol. 53, 197–204 (1993).
-
Lagasse, E. & Clerc, R. G. Cloning and expression of two human genes encoding calcium-binding proteins that are regulated during myeloid differentiation. Mol. Cell. Biol. 8, 2402–2410 (1988).
-
Marenholz, I., Heizmann, C. W. & Fritz, G. S100 proteins in mouse and man: from evolution to function and pathology (including an update of the nomenclature). Biochem. Biophys. Res. Commun. 322, 1111–1122 (2004).
-
Yang, Z. et al. Proinflammatory properties of the human S100 protein S100A12. J. Leukoc. Biol. 69, 986–994 (2001).
-
Burgoyne, R. D. & Geisow, M. J. The Annexin family of calcium-binding proteins. Review Article. Cell. Calcium. 10, 1–10 (1989).
-
Dado-Senn, B., Skibiel, A. L., Dahl, G. E., Apelo, A., Laporta, J. & S. I. & Dry period heat stress impacts mammary protein metabolism in the subsequent lactation. Anim. (Basel). 11, 2676 (2021).
-
Kaufman, J. D., Kassube, K. R., Almeida, R. A. & Ríus, A. G. Short communication: high incubation temperature in bovine mammary epithelial cells reduced the activity of the mTOR signaling pathway. J. Dairy. Sci. 101, 7480–7486 (2018).
-
Upheber, S. et al. Alternative splicing of KAI1 abrogates its tumor-suppressive effects on integrin αvβ3-mediated ovarian cancer biology. Cell. Signal. 27, 652–662 (2015).
-
Liu, W. M. & Zhang, X. A. KAI1/CD82, a tumor metastasis suppressor. Cancer Lett. 240, 183–194 (2006).
-
Reddi, K. K. et al. Tetraspanin CD82 correlates with and May regulate S100A7 expression in oral Cancer. Int. J. Mol. Sci. 25, 2659 (2024).
-
Yamazaki, M. et al. MFG-E8 expression for progression of oral squamous cell carcinoma and for self-clearance of apoptotic cells. Lab. Invest. 94, 1260–1272 (2014).
-
Okamoto, A. et al. Immunological and clinicopathological significance of MFG-E8 expression in patients with oral squamous cell carcinoma. Pathol. Oncol. Res. 26, 1263–1268 (2020).
-
Jinushi, M. et al. Milk fat globule EGF-8 promotes melanoma progression through coordinated Akt and twist signaling in the tumor microenvironment. Cancer Res. 68, 8889–8898 (2008).
-
Yang, Y. et al. Reduction in milk fat globule-EGF factor 8 inhibits triple-negative breast cancer cell viability and migration. Oncol. Lett. 17, 3457–3465 (2019).
-
Bu, H. F. et al. Milk fat globule-EGF factor 8/lactadherin plays a crucial role in maintenance and repair of murine intestinal epithelium. J. Clin. Invest. 117, 3673–3683 (2007).
-
Fang, Y. Y. & Zhang, J. H. MFG-E8 alleviates oxygen-glucose deprivation-induced neuronal cell apoptosis by STAT3 regulating the selective polarization of microglia. Int. J. Neurosci. 131, 15–24 (2021).
-
Gao, Y. Y. et al. Recombinant milk fat globule-EGF factor-8 reduces apoptosis via integrin β3/FAK/PI3K/AKT signaling pathway in rats after traumatic brain injury. Cell. Death Dis. 9, 845 (2018).
-
Zhao, Q., Yu, Y., Zuo, X., Dong, Y. & Li, Y. Milk fat globule-epidermal growth factor 8 is decreased in intestinal epithelium of ulcerative colitis patients and thereby causes increased apoptosis and impaired wound healing. Mol. Med. 18, 497–506 (2012).
-
Fuller, A. D. & Van Eldik, L. J. MFG-E8 regulates microglial phagocytosis of apoptotic neurons. J. Neuroimmune Pharmacol. 3, 246–256 (2008).
-
Hanayama, R., Miyasaka, K., Nakaya, M. & Nagata, S. MFG-E8-dependent clearance of apoptotic cells, and autoimmunity caused by its failure. Curr. Dir. Autoimmun. 9, 162–172 (2006).
-
Hanayama, R. & Nagata, S. Impaired Involution of mammary glands in the absence of milk fat globule EGF factor 8. Proc. Natl. Acad. Sci. U S A. 102, 16886–16891 (2005).
-
Ran, F. A. et al. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 8, 2281–2308 (2013).
-
Anand, V. et al. Establishment and characterization of a Buffalo (Bubalus bubalis) mammary epithelial cell line. PLoS One. 7, e40469 (2012).
Acknowledgements
The authors acknowledge DST-SERB for providing funds for conducting the research.
Funding
This research was funded by DST-SERB, grant number EMR/2017/000152.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Vats, P., Kumar, R., Kumar, R. et al. Deciphering the role of MFGE8 in lactation using CRISPR-CAS9 based gene editing in Buffalo mammary epithelial cells. Sci Rep 15, 30194 (2025). https://doi.org/10.1038/s41598-025-00155-7
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-00155-7