Introduction
Since the commercialization of genetically modified (GM) crops in 1996, the area planted with such crops has been increasing year by year. In 2023, the global area under GM crop production increased 1.9% over the previous year to reach 206.3 million hectares, a new record area (http://gm.agbioinvestor.com, accessed on April 28, 2024). In China, no GM crops are currently planted yet, however, the Ministry of Agriculture and Rural Affairs of China has issued safety certificates for 20 transgenic corn hybrids, seven transgenic soybeans events, and two transgenic rice lines (http://www.moa.gov.cn/, accessed on May 10, 2024), GM crops will be commercially planted in China soon. Despite the well-documented benefits of GM crops, such as reduced pesticide use and improved farmer health, concerns persist regarding their potential environmental and food safety risks, including unintended effects on non-target organisms1.
Among GM crops, Bt rice has been a focal point of research due to its ecological implications. Rice fields are aquatic ecosystems where Bt proteins may enter the water through plant residues or pollen, exposing aquatic organisms like fish to these proteins. Previous studies have detected low concentrations of Bt proteins (up to 30 ng/L for Cry1Ab/Ac) in rice field waters2,3raising questions about their long-term effects on aquatic life. While extensive regulatory testing is conducted before GM crop approval, critical gaps remain in understanding the chronic and sublethal impacts of Bt proteins on non-target species, particularly under realistic exposure scenarios. Additionally, the random insertion of exogenous genes may affect the expression of endogenous genes in GM crops, leading to unexpected effects, such as changes in the expression levels of large amounts of elements, trace elements, anti-nutritional factors, or intrinsic plant toxins4,5. Studies have found that there is a certain amount of residual exogenous protein in GM crops after they are processed into feed, which may induce immune stress reactions in animals, affecting their growth, development, reproduction, etc67,. Therefore, whether GM crops have the same nutritional components as traditional crops and whether the exogenous genes and their expressed proteins pose potential risks to the growth and health of fish have gradually become a focus of attention for environmentalists and the fish farming industry.
Fish are highly sensitive to physical, chemical, and biological changes in their environment and have thus been widely used to monitor changes in water environment quality8. At present, ecotoxicological research on fish as a model animal focuses mainly on the toxic effects of pesticides, antibiotics, heavy metals, and other chemicals in water9,10. So far, there are some research on the safety of GM crops on fish. The experimental animals commonly used in this area are Atlantic salmon (Salmo salar), rainbow trout (Oncorhynchus mykiss), and zebrafish (Danio rerio)11. The common method is prepare a feed mixture that contains a certain amount of the GM crop and feed the fish for a period of time to evaluate their health status. Common toxicity parameters for fish include their growth rate, mortality rate, health index, organ function and development, histopathological and hematological indicators, antioxidant enzyme activity in organ tissues, and mRNA expression levels of sensitive proteins11. In terms of the safety evaluation of Bt crops on fish, the results of multiple studies are inconsistent. Some have shown that a diet containing Bt crops has no significant negative impact on the growth and development of fish when compared with a diet with the same crop from a non-transformed line12,13,14,15,16whereas other studies have shown that Bt crops can promote fish growth17,18. At least one study, however, has shown an opposite effect. Hemre et al. found that after 82 days of feeding, fish fed the GM maize showed slight but significant lower feed intake, which was followed by slight but significant lower specific growth rate and final body weights, compared with fish fed nGM maize19. These discrepancies highlight the need for systematic, multi-parameter evaluations to elucidate the ecological safety of Bt crops, especially for rice, which occupies a unique niche in aquatic-agricultural systems. While, the majority of existing research has concentrated on Bt maize or soybeans, with limited data on Bt rice—a staple crop in Asia. Furthermore, current studies frequently omit critical endpoints like gut microbiota analysis, now widely acknowledged as a vital biomarker for organismal health and environmental adaptation20.
The zebrafish (Danio rerio) is a model animal that is widely used in developmental biology, molecular genetics, and toxicological research because of its rapid development, high fecundity, high sensitivity to environmental contaminants, easy maintenance, and relatively low husbandry costs21,22. The International Organization for Standardization (ISO) and the Organisation for Economic Co-operation and Development (OECD) have officially listed zebrafish as a standard test species for chemical assessments and water quality evaluations. The genetically modified rice line T1C-19, which expresses the Cry1C protein, may be used as a commercial rice line in China because of its good resistance to lepidopteran rice pests including the stem borers Chilo suppressalis, Scirpophaga incertulas, and Cnaphalocrocis medinalis23,24.
This study addresses these gaps by evaluating the effects of transgenic cry1C rice (T1C-19) on zebrafish through a 90-day dietary exposure experiment. We examined a multiple test endpoints including growth parameters, digestive and antioxidant enzyme activities, and intestinal microbial diversity. Our findings aim to: (1) Provide empirical data on the chronic exposure effects of Cry1C protein in a realistic aquatic model. (2) Clarify inconsistencies in the literature by incorporating multi-dimensional health indicators. (3) Support regulatory decisions as China moves toward GM crop commercialization. By bridging these research gaps, results from this study contributes to a more nuanced understanding of Bt rice’s environmental and food safety, ensuring that policy and practice are grounded in rigorous science.
Materials and methods
Ethics statement
This study was approved by the Committee of Laboratory Animal Experimentation at Institute of Plant Protection, Chinese Academy of Agricultural Sciences (approval number: Ipp-201805F003). All procedures involving experimental animals were performed in accordance with the Guide for the Care and Use of Laboratory Animals25. Briefly, all the animals were humanely treated during this study, the anesthetic procedure with 0.1% MS-222 (Sigma-Aldrich) was adopted, if necessary, to reduce the suffering of the experimental animals. This study is reported in accordance with ARRIVE guidelines 2.0 (Animal Research: Reporting of In Vivo Experiment).
Transgenic rice
The transgenic rice line T1C-19 and its corresponding non-transgenic near-isoline Minghui 63 (MH63) were used for all experiments. The expression of the synthetic cry1C gene in T1C-19 rice is under the control of the corn ubiquitin promoter26. The MH63 line is commonly grown in China and is an elite indica line with cytoplasmic male sterility. Both rice lines were obtained from and preserved at Huazhong Agricultural University (Wuhan, China). It’s identification was confirmed by PCR analysis and ELISA detection as previously described26. The two rice lines were planted at the same time in adjacent plots in the experimental field station of the Institute of Plant Protection, Chinese Academy of Agricultural Sciences (39.50°N, 116.60°E). Rice plants were cultivated according to commonly used local agricultural practices but without application of chemical insecticides. Rice was harvested, and rice grains were collected, pooled, and stored at 4 °C until used. The conventional nutritional components of each rice line were analyzed by the Beijing Research Institute for Nutritional Resources, People’s Republic of China.
Test animals
Adult zebrafish (AB strain) were purchased from the Center of National Zebrafish Resources of China (Wuhan, Hubei, China). They were cultured in a recirculating aquarium system at ambient temperature (28 ± 1 °C) under 14-h light/10-h dark cycles according to standard procedures27. The fish were fed twice a day with freshly hatched brine shrimp (Artemia salina). To generate embryos, male and female fish were placed in a breeding tank, with a sex ratio of 2:1 males/females overnight. Spawning was triggered after the lights were turned on the next morning and was completed within 30 min. Embryos from the same pair of zebrafish were collected, and they were cultured to stage 45 days post-fertilization (dpf) with a diet of Paramecium or freshly hatched brine shrimp.
Diet formulation
Synthetic zebrafish diets were produced based on the nutrient composition of a commercial zebrafish feed. Two test diets contained 20% rice flour, whereas the control (CK) diet was formulated using wheat bran, corn flour, and stone powder without rice. The detailed compositions of the diets are shown in Table 1. All ingredients except rice grain were purchased from Beijing Ruiheng Junan Technology Co., Ltd (Beijing, China). The conventional nutrient composition of the diets was measured by the Beijing Research Institute for Nutritional Sources (Beijing, China).
Experimental design
Zebrafish (45 dpf) with uniform body length (~ 14.7 mm) and weight (~ 21.4 mg) were divided into three experimental groups, i.e., T1C-19, MH63, and CK groups; their sexes were not considered. Each group was randomly divided into four glass jars (20 × 34 × 24 cm) to generate four replicates, and each replicate group contained 8 or 10 zebrafish. The zebrafish were fed daily (~ 3% weight/body weight) for 90 days, and the water was exchanged 50% in every two days. The dissolved oxygen in the water was greater than 9.0 mg/L, the pH ranged from 7.0 to 7.8, the nitrite concentration was less than 0.05 mg/L, the ammonia nitrogen concentration was less than 0.02 mg/L, and the water temperature was maintained at ~ 28 °C.
During the feeding of zebrafish, the filtering and oxygenation equipment was turned off to avoid affecting the food intake of the fish and then was turned on after feeding. Animals were observed twice a day, dead fish were recorded and removed without delay during daily monitoring. The food intake and number of fish death were record, and their body length and weight were measured every 15 days. Animals were fasted overnight before being sacrificed. At the end of the experiment, the zebrafish were anesthetized by immersion in 1% MS-222 (Sigma Chemical Company, St. Louis, MO, USA). Before dissection, body weight and length were measured, and the gender was determined on the basis of gross gonadal morphology. After dissection, the obtained tissue samples were immediately frozen in liquid nitrogen and preserved at − 80 °C for subsequent bioassays.
Detection of enzymatic activities
The activities of trypsin and amylase in the intestine as well as super oxide dismutase (SOD) and catalasein (CAT) the liver were measured at the end of the experiment. The intestinal and liver tissues of three or four zebrafish in each glass jar (a total of 12–16 samples per treatment) were dissected under the stereoscope and then individually transferred into a 1.5-mL centrifuge tube and immediately stored at − 80 ℃ for the detection of the above mentioned four parameters. All parameters were detected using commercial kits provided by the JianCheng Bioengineering Institute (Nanjing, Jiangsu Province, China), and the specific product numbers of the above four kits were A080-2-2, C016-1-1, A001-1-2, and A007-1-1, respectively. All samples were homogenized in 0.8% physiological saline at a ratio of 1:9 (w/v) and centrifuged at 2500 g at 4 °C for 10 min. The supernatants were collected and immediately used for analysis.
Determination of Cry1C content
The Cry1C protein content in grains of rice, zebrafish diets, and feces was determined using a Bt Cry1C protein kit (EnviroLogix Inc., Portland, OR, USA; cat. No. AP007) with a detection limit of 0.1 ng/g total protein. Solid feces were collected immediately after the renewal of rearing water at the later stage of the present study. Before analysis, all samples were washed in phosphate-buffered saline/Tween-20 (PBS/Tween-20) to remove Bt toxins from their outer surface. For Cry1C extraction, samples of rice grains, diets or feces were weighed, lyophilized, and then homogenized in 1 mL PBS/Tween-20 using a micropestle and mortar on ice. After centrifugation and appropriate dilution of the supernatants, an enzyme-linked immunosorbent assay (ELISA) was performed with the Bt Cry1C protein kit. Optical density values were read using a microplate spectrophotometer (PowerWave XS2, BioTek, Winooski, VT, USA). A standard curve derived from a Cry1C standard provided with the kit was used to calculate Cry1C levels.
Determination of intestinal microbial diversity
The intestines were collected from five randomly selected male and female zebrafish from each treatment group, i.e. 10 total samples per treatment, and were quick-frozen in liquid nitrogen and kept at − 70 °C until analysis. Amplicon sequencing of gut bacteria was performed according to previous workflows28. Briefly, whole genomic DNA was extracted using the QIAamp PowerFecal Pro DNA Kits (Qiagen, Hilden, Germany; cat. No. 51804) to amplify the V3–V4 region of 16 S rDNA using the primer pair 338 F and 806R. DNA libraries were sequenced on an Illumina NovaSeq platform (Illumina, San Diego, CA, USA). After data filtration and chimera removal, the resulting effective tags were used to generate operational taxonomic units for species annotation using QIIME2 and R packages (v3.2.0). Alpha-diversity indices, such as the Chao1 richness estimator, Shannon diversity index, abundance-based coverage estimator (ACE), and the Simpson index were calculated using the amplicon sequence variants table in QIIME2 and are shown as the mean ± standard deviation (SD). A diversity analysis was performed to investigate the structural variation of microbial communities across samples and visualized via principal coordinate analysis, non-metric multidimensional scaling (NMDS), and the unweighted pair-group method with arithmetic means hierarchical clustering. Principal component analysis was conducted based on the genus-level compositional profiles, and the significance of differentiation of microbiota structure among groups was also assessed.
Data analyses
To evaluate the growth status of zebrafish, we calculated the condition factor (CF, g/cm3) and specific growth rate (SGR, %/day) at the end of the study period. We used the following equation: CF = W/L3 × 100, Where W means body weight, L means body length. SGR = (LnWf – LnWi) × 100/t, Where Wf and Wi mean final body weight and initial body weight respectively, and t means feeding days.
All data are presented as the mean ± SD unless indicated otherwise. The survival rates of zebrafish for each treatment relative to the control were compared using the χ2 test. A one-way ANOVA followed by Tukey’s honest significant differences (Tukey’s HSD) multiple comparison test was used to analyze differences in body length, body weight, and enzymatic activities, as well as the intestinal microbial diversity index, among the three treatment groups. Differences were considered significant at P < 0.05. All statistical analyses were conducted using the SPSS software package (version 17.0 for Windows, 2008).
Results
Nutrient composition of the diet
The conventional nutrient composition of the diets was measure, and the measured values of the nutrients in the T1C-19 diet were comparable to those in the MH63 and control diets (Table 2).
General health
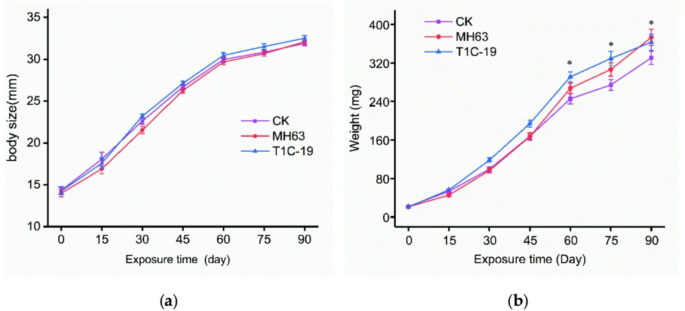
Animals were observed twice daily to confirm their well-being, and body weight and length were measured every 15 days. The mean body weights and body lengths of animals over 90 days for each treatment group are shown in Fig. 1. There was no significant difference in the body length of zebrafish among the three treatment groups during the whole experiment (Fig. 1a) and in body weight from day 0 to day 45 (Fig. 1b). There were significant differences in the body weight of the zebrafish on day 60, 75, and 90 (P < 0.05; Fig. 1b). When we analyzed male and female zebrafish separately for body length and weight differences at the end of the feeding trial, however, no statistically significant differences among the T1C-19, MH63, and control groups were found in either the male or female zebrafish (Table 3).

Body length and weight over time in zebrafish fed with the test or control diets. (a) Body length; (b) Body weight. *An asterisk indicates a significant difference relative to the other groups (one-way ANOVA followed by Tukey’s HSD test; P < 0.05).
The survival rate of zebrafish on day 90 in the MH63 treatment group was slightly lower than that of the CK and T1C-19 treatment groups, but there were no significant differences when the MH63 and T1C-19 treatment groups were compared with the CK group (MH63: χ2 = 0.57, P = 0.45; T1C-19: χ2 = 0.01, P = 0.94, respectively). As for CF and SGR, there was also no significant difference among the three treatments (P = 0.17 for CF and p = 0.21 for SGR; Table 4).
Enzymatic activities
The results showed that there were no significant differences in the activities of the two digestive enzymes among the three treatments (Table 5). The activities of SOD and CAT in zebrafish from the T1C-19 group were higher than those from the MH63 and CK groups; however, there were no significant differences among the three treatment groups (all P > 0.05).
Cry1C protein content
Cry1C was present in T1C-19 rice grains at an average level of 591.76 ± 31.72 ng/g and, as expected, was not detected (ND) in its not genetically modified strain, MH63 (Table 6). The amount of Cry1C present in the T1C-19 diet (137.28 ± 17.65 ng/g; Table 6) was similar to the predicted value (118.36 ng/g). To confirm whether zebrafish ingest Cry1C protein, feces were collected and the Cry1C protein content was determined by ELISA. Cry1C protein was detected in the feces of zebrafish fed the T1C-19 diet (91.01 ± 12.69 ng/g) but not in the feces of those fed the MH63 or control diet (Table 6).
Intestinal microbial diversity
To measure the diversity of zebrafish gut microbiota in each treatment group, we used intestinal samples from the fish to determine the alpha-diversity indices, including the Chao1 richness estimator and ACE, which reflect community richness, as well as the Shannon index and Simpson index, which are a measure of community evenness. The alpha-diversity indices calculated for each treatment group are shown in Table 7. The diet that included T1C-19 rice was associated with no significant differences in the diversity indices as compared with the MH63 and CK diets (all P > 0.05; Table 7).
A principal component analysis of the community structure at the genus level showed that the T1C-19 diet could be distinguished from the CK diet, but there was overlap between the T1C-19 diet and the diet with its parental strain, MH63, which resulted in samples from the two groups showing substantial clustering (Fig. 2a). The NMDS analysis results (Fig. 2b) were consistent with the above results and showed similar clustering between the Bt rice and non-Bt rice diets. A statistical comparison of the mean distances of samples from different groups showed that there were no significant differences among treatments (Fig. 2c).

A beta-diversity analysis of zebrafish intestinal microbes. (a) Principal component analysis, PCA. (b) Analysis of unweighted UniFrac NMDS. (c) Boxplot of unweighted UniFrac multiple comparison distances.
Raw sequencing reads of the zebrafish gut microbiota were submitted to the NCBI SRA (Submission ID: SUB15245224) and are accessible under BioProject ID PRJNA1248985.
Discussion
Food and environmental safety assessment is an important part of the risk assessment of GM crops. The principle of substantial equivalents is one of the four principles followed in the evaluation of GM crops, and thus it is necessary to determine the nutritional composition of GM crops before testing. Our results showed that the measured values of conventional nutritional components in the T1C-19 diet were comparable to those in the MH63 and control diets. This is confirmed by the many studies that have shown that there are no significant differences in conventional nutrient composition between Bt rice and non-Bt rice29,30,31,32.
In the natural environment, fish can be exposed to Bt protein through several routes. Bt protein can be passed along through the food chain, and Bt protein entering water via Bt crop residues or pollen33. However, the dose at which fish would be exposed to Bt protein in rice fields should be very low. The reported maximum Bt protein concentration in a natural aquatic environment is less than 130 ng/L for Bt corn34,35. For Bt rice, the maximum reported level is even lower, at 30 ng/L2–3. In our experiment, we collected and detected the Cry1C protein in grains of Bt rice, in the diet that included this rice, and in the feces of zebrafish that consumed the diet. Thus in our study the Cry1C protein was present and could pass down throughout the food chain, which is consistent with a previous report36.
Although there were no significant differences in the survival, growth, or development of zebrafish that were fed a diet containing Bt rice relative to those that were fed a diet with non-Bt rice, there was a significant difference in the weight of zebrafish between the two treatments after 60 days. This difference was absent, however, when male and female fish were analyzed separately. This is likely due to unequal ratios of males to females across the treatment groups (15:12 for T1C-19, 17:8 for MH63, and 16:18 for CK). It is well known that adult female fish generally weigh more than males because of reproductive needs. Female zebrafish exhibit a rounder body shape and greater body mass compared to males, primarily due to the development of ovaries and egg production. This sexual dimorphism in body shape and weight becomes particularly evident after sexual maturation (typically 3 months of age), and adult females can weigh 20–30% more than age-matched males9,37,38. In our study, the weight of female zebrafish at day 90 was ~ 30% heavier than that of male which is consistent with the above findings. Thus, the weight difference observed at day 60 is most likely attributable to sex ratio imbalance rather than Cry1C protein exposure.
Based on the reported literature, there is a debate regarding the effects of Bt crops on the development of non-target organisms (e.g., fish). Hemre et al. found a significant lower feed intake, specific growth rate and final body weights, compared with fish fed nGM maize19. While some studies have shown that a diet containing a Bt crop has no significant negative impacts on the growth and development of fish when compared with its non-transformed line12,13,14,15,16. Notably, some other studies have even shown that Bt maize can promote fish growth17,18. Sanden et al. found a significantly higher body mass increase, SGR and feed utilization than zebrafish fed the non-Bt diet, they postulated that these differences might be associated with reduced mycotoxin contamination in Bt maize18. Our results demonstrated no statistically significant differences in all measured growth and developmental endpoints between the Bt rice, non-Bt and control diet. These findings align with and substantiate previous reports on the nutritional equivalence of Bt and non-Bt crops in fish models.
A diet containing GM crops can cause biological changes at the molecular level and lead to oxidative stress in animals. For example, compared with fish fed non-GM crops, those fed GM crops showed significantly lower CAT activity and significantly higher levels of heat shock protein 70 in the liver39 decreased SOD and CAT expression and reduced activity of leucine aminopeptidase and maltase in the intestine39 and lower transcription of sod1 in the liver17. Sanden et al. fed zebrafish Bt maize diets for two generations and found no significant effects in the parental generation. In contrast, the liver and mid-intestinal gene transcript levels of CuZn SOD were significantly higher, and liver gene transcript levels of caspase 6 were significantly lower in the offspring of the fish fed the non-Bt diet relative to those fed the Bt diet18. In our experiments, T1C-19 rice did not cause significant changes in the activity of similar zebrafish enzymes, which is consistent with some other studies12,15. Although there have been statistically significant differences in the physiological indicators of fish that were fed Bt crops in the above studies, it is not clear whether these differences were caused by mycotoxin contamination or whether these differences may fall within the normal range of fluctuation.
Our study demonstrated that dietary exposure to Cry1C protein at 137.28 ± 17.65 ng/g in zebrafish feed—a concentration orders of magnitude higher than reported environmental levels (~ 30 ng/L in Bt rice fields)—elicited no adverse effects. While these findings support the safety of T1C-19 rice, we acknowledge that real-world exposure dynamics in aquatic ecosystems are more complex. Field studies report rapid dilution and degradation of Bt proteins in water, with concentrations declining due to seasonal rainfall, microbial activity, and adsorption to sediments. For instance, Liu et al. observed that Cry1Ab/1Ac protein from Bt rice persisted in water for < 7 days, with degradation accelerated by microbial proteases and organic matter2. Similarly, Strain & Lydy noted that Cry1Ab adhered to sediments within 48 h, reducing bioavailability to aquatic organisms35. These processes likely further minimize exposure risks in natural settings compared to controlled laboratory conditions. Our study’s high-dose paradigm provides a conservative safety threshold, yet future research should incorporate environmental variables (e.g., UV exposure, temperature shifts, and sediment interactions) to refine risk models. Investigating such degradation pathways would strengthen ecological relevance. While our data robustly indicate negligible risks under sustained high exposure, complementary field studies tracking Cry1C persistence and bio-accumulation would contextualize these findings for natural ecosystems.
Our ELISA results also confirmed the presence of Cry1C in zebrafish feces (91.01 ± 12.69 ng/g), demonstrating ingestion, the absence of tissue-specific data limits conclusions about absorption or bio-transformation. Prior studies on Bt proteins in fish (e.g., Gu et al.) detected no systemic accumulation in Atlantic salmon fed Bt maize40 suggesting minimal absorption. However, to fully exclude bio-accumulation risks for Cry1C, future work should quantify Cry1C in intestinal, liver, and plasma using sensitive techniques like immunohistochemistry or LC-MS/MS, which can distinguish intact proteins from degradation products. The lack of physiological effects (e.g., unchanged liver antioxidant enzymes, growth parameters) indirectly supports limited absorption. Nonetheless, direct tissue analyses would strengthen conclusions, as seen in Mao et al. where Cry1Ab/1Ac in macaque tissues confirmed low bioavailability41. We recommend such methods for future chronic exposure studies.
Some forms of inflammatory bowel disease are related to intestinal health disorders42 suggesting that the intestinal microecology is important not only to an animal’s digestive system but also to its immune status and overall health. In fact, changes in intestinal microorganisms are related to many factors such as age, dietary intake, drugs, or physical conditions43 among which dietary intake is one of the most important factors44. The Bt protein influences the intestinal microbial diversity and community composition of its target insect45. Nowadays, there is growing concern over whether the planting of GM crops will have negative effects on the intestinal microecology of non-target organisms, and thus it is necessary to evaluate the effects of GM crops on the intestinal microecology of animals within the ecosystem. Our results showed that, as compared with a diet containing non-Bt rice, a diet containing Bt rice had no significant effects on the alpha-diversity indices (Simpson, Chao1, ACE and Shannon) and beta-diversity indices of the intestinal microbial community of zebrafish. Using PCR-DGGE analysis, Cao et al. demonstrated that the fecal bacterial profiles of rats fed T1C-19 transgenic rice were comparable to those consuming non-Bt rice controls30. These findings align with our current results obtained from the same T1C-19 rice line, further supporting the conclusion that Cry1C protein expression does not significantly alter gut microbiota composition.
Through systematic experimental design and multi-parameter analysis, this study provides empirical evidence to confirm that Cry1C protein at tested doses causes no significant adverse effects on zebrafish. In fact, the evaluation of the effects of a diet with Bt rice has been carried out in a variety of animals, such as rats30,46 chickens47 cynomolgus macaques41 pigs48,49 carp16 frogs32 and other organisms. All of these studies have confirmed the safety of Bt rice from a dietary standpoint. This study provides scientific evidence to support the commercialization of T1C-19 rice as an aquafeed ingredient, facilitating the application of GM crops in the feed industry. As a model organism, zebrafish serve as an ecological indicator, our findings indirectly reflect the potential risks of GM rice to aquatic ecosystems, offering critical data for environmental safety assessments. Furthermore, as China progressively advances the commercial cultivation of GM crops, results from this study serve as valuable references for science-driven policymaking and regulatory frameworks, facilitating informed decision-making and public understanding of GM crop safety.
Conclusion
In the present study, zebrafish were exposed to Cry1C proteins by feeding them a diet containing T1C-19 rice grains for 90 days. No significant differences were found between Bt and non-Bt rice treatment in terms of survival rate, body length and body weight, condition factor (CF), specific growth rate (SGR), intestinal digestive activity (Trypsin and Maltase), liver antioxidant enzyme activity (SOD and CAT), as well as the intestinal microbial diversity. Based on the results, we conclude that transgenic rice T1C-19 exhibits the same safety profile as non-transgenic rice.
Data availability
The raw sequence data of the zebrafish gut microbiota have been added to the NCBI SRA with accession number PRJNA1248985.
References
-
Li, Y., Hallerman, E. M., Liu, Q., Wu, K. & Peng, Y. The development and status of Bt rice in China. Plant. Biotechnol. J. 14, 839–848 (2016).
-
Liu, Y. B., Li, J. S., Luo, Z. L., Wang, H. R. & Liu, F. The fate of fusion Cry1Ab/1Ac proteins from Bt-transgenic rice in soil and water. Ecotoxicol. Environ. Saf. 124, 455–459 (2016).
-
Wang, Y. M. et al. Determination of the movement and persistence of Cry1Ab/1Ac protein released from Bt Transgenic rice under field and hydroponic conditions. Soil. Biol. Biochem. 58, 107–114 (2013).
-
Cellini, F. et al. Unintended effects and their detection in genetically modified crops. Food Chem. Toxicol. 42, 1089–1125 (2004).
-
Somers, D. A. & Makarevitch, I. Transgene integration in plants: poking or patching holes in promiscuous genomes? Curr. Opin. Biotech. 15, 126–131 (2004).
-
Prescott, V. E. et al. Transgenic expression of bean α-amylase inhibitor in peas results in altered structure and immunogenicity. J. Agri Food Chem. 53, 9022–9030 (2005).
-
Frøystad-Saugen, M. K. et al. Distal intestinal gene expression in Atlantic salmon (Salmo salar, L.) fed genetically modified maize. Aquaacult Nutr. 15, 104–115 (2009).
-
Sanchez, W. et al. Fish health status as a freshwater quality indicator. Chem. Geol. 398, 32–45 (2012).
-
Hu, C., Bai, Y., Sun, B., Tang, L. & Chen, L. Significant impairment of intestinal health in zebrafish after subchronic exposure to Methylparaben. Sci. Total Environ. 838, 156389 (2022).
-
Kim, M., An, G., Park, J., Song, G. & Lim, W. Bensulide-induced oxidative stress causes developmental defects of cardiovascular system and liver in zebrafish (Danio rerio). J. Hazard. Mater. 455, 131577 (2023).
-
Sissener, N. H. et al. Genetically modified plants as fish feed ingredients. Can. J. Fish. Aquat. Sci. 68, 563–574 (2011).
-
Dong, S. S. et al. Ecotoxicological effects of Transgenic mCry1Ac maize (BT799) on zebrafish. J. Appl. Ecol. 30, 2845–2853 (2019).
-
Pham, M. et al. Preliminary studies on the effects of dietary genetically modified Soya and corn on growth performance and body composition of juvenile Olive flounder (Paralichthys olivaceus) and rockfish (Sebastes schlegeli). Fish. Aquat. Sci. 13, 63–70 (2010).
-
Sanden, M., Krogdahl, Å., Bakke-Mckellep, A. M., Buddington, R. K. & Hemre, G. I. Growth performance and organ development in Atlantic salmon, Salmo salar L. parr fed genetically modified (GM) soybean and maize. Aquacult. Nutr. 12, 1–14 (2006).
-
Xiang, D. et al. Safety assessment of subchronic feeding of insect-resistant and herbicide-resistant Transgenic soybeans to juvenile channel catfish (Ictalurus punctatus). Sci. Rep. 13, 5445–5445 (2023).
-
Xu, Z., Liu, Y., Xing, F. & Yang, X. Nutritional safety assessment of the brown rice genetically modified with CrylAc and SCK as feed on carp. Cereal Feed Ind. 7, 57–60 (2011).
-
Sissener, N. H. et al. Zebrafsh (Danio rerio) as a model for investigating the safety of GM feed ingredients (soya and maize); performance, stress response and uptake of dietary DNA sequences. Br. J. Nutr. 103, 3–15 (2010).
-
Sanden, M. et al. Cross-generational feeding of Bt (Bacillus thuringiensis)-maize to Zebrafsh (Danio rerio) showed no adverse effects on the parental or offspring generations. Br. J. Nutr. 110, 2222–2233 (2013).
-
Hemre, G. I. et al. Nutritional, physiological, and histological responses in Atlantic salmon, Salmo salar L, fed diets with genetically modified maize. Aquacult. Nutr. 13, l86–l99 (2007).
-
Nayak, S. K. Role of Gastrointestinal microbiota in fish. Aquac Res. 41, 1553–1573 (2010).
-
Hill, A. J., Teraoka, H., Heideman, W. & Peterson, R. E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 86, 6–19 (2005).
-
Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 269, 1–20 (2007).
-
Zheng, X. S. et al. Resistance performances of Transgenic Bt rice lines T2A-1 and T1c-19 against Cnaphalocrocis Medinalis (Lepidoptera: Pyralidae). J. Ecol. Entomol. 104, 1730–1735 (2011).
-
Wang, Y. N. et al. Comparison of three Transgenic Bt rice lines for insecticidal protein expression and resistance against a target pest, Chilo suppressalis (Lepidoptera: Crambidae). Insect Sci. 23, 78–87 (2016).
-
National Research Council (US). Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animals 8th edn (National Academies Press (US), 2011).
-
Tang, W. et al. Development of insect-resistant Transgenic indica rice with a synthetic cry1C* gene. Mol. Breed. 18, 1–10 (2006).
-
Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio) 5th edn (University of Oregon, 2000).
-
Chen, L. et al. Perfluorobutanesulfonate exposure causes durable and transgenerational dysbiosis of gut microbiota in marine Medaka. Environ. Sci. Technol. Lett. 5, 731–738 (2018).
-
Wang, Z. H. et al. Toxicological evaluation of Transgenic rice flour with a synthetic cry1Ab gene from Bacillus thuringiensis. J. Sci. Food Agric. 82, 738–744 (2002).
-
Cao, S. et al. Safety assessment of Transgenic Bacillus thuringiensis rice T1c-19 in Sprague-Dawley rats from metabonomics and bacterial profile perspectives. IUBMB Life. 64, 242–250 (2012).
-
Li, X. et al. Comparison of nutritional quality between Chinese indica rice with sck and cry1Ac genes and its nontransgenic counterpart. J. Food Sci. 72, S420 – 424 (2007).
-
Zhu, H. J. et al. A 90-day safety assessment of genetically modified rice expressing Cry1Ab/1Ac protein using an aquatic animal model. J. Agri Food Chem. 63, 3627–3633 (2015).
-
Carstens, K. et al. Genetically modified crops and aquatic ecosystems: considerations for environmental risk assessment and nontarget organism testing. Transgenic Res. 21, 813–842 (2012).
-
Tank, J. et al. Occurrence of maize detritus and a Transgenic insecticidal protein (Cry1Ab) within the stream network of an agricultural landscape. Proc. Natl. Acad. Sci. U S A. 107, 17645–17650 (2010).
-
Strain, K. E. & Lydy, M. J. The fate and transport of the Cry1Ab protein in an agricultural field and laboratory aquatic microcosms. Chemosphere 132, 94–100 (2015).
-
García, M., Ortego, F., Castañera, P. & Farinós, G. P. Effects of exposure to the toxin Cry1Ab through Bt maize fed-prey on the performance and digestive physiology of the predatory Rove beetle Atheta coriaria. Biol. Control. 55, 225–233 (2010).
-
Robison, B. D. et al. Sexual dimorphism in hepatic gene expression and the response to dietary carbohydrate manipulation in the zebrafish (Danio rerio). Comp. Biochem. Physiol. Part. D Genomics Proteom. 3, 141–154 (2008).
-
Green, G. B. H. et al. Body metrics and the gut Microbiome in response to macronutrient limitation in the zebrafish Danio rerio. Curr. Dev. Nutr. 7, 100065 (2023).
-
Sagstad, A. et al. Evaluation of stress- and immune-response biomarkers in Atlantic salmon, Salmo salar L., fed different levels of genetically modified maize (Bt maize), compared with its near-isogenic parental line and a commercial Suprex maize. J. Fish. Dis. 30, 201–212 (2007).
-
Gu, J., Bakke, A. M., Valen, E. C., Lein, I. & Krogdahl, Å. Bt-maize (MON810) and non-GM soybean meal in diets for Atlantic salmon (Salmo Salar L.) juveniles–Impact on survival, growth performance, development, digestive function, and transcriptional expression of intestinal immune and stress responses. PloS One. 9, e99932 (2014).
-
Mao, J. et al. A 52-week safety study in cynomolgus macaques for genetically modified rice expressing Cry1Ab/1Ac protein. Food Chem. Toxicol. 95, 1–11 (2016).
-
Nissen, L., Walter, C., Sgorbati, B., Biavati, B. & Cencic, A. Gut health promoting activity of new putative probiotic/protective Lactobacillus spp. Strains: A functional study in the small intestinal cell model. Int. J. Food Microbiol. 135, 288–294 (2009).
-
Nadal, A. L. et al. Feed, microbiota, and gut immunity: using the zebrafish model to understand fish health. Front. Immuno. 11, 114 (2020).
-
Claesson, M. J. et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 488, 178–184 (2010).
-
Sun, Z. B., Hu, Y. F., Song, H. J., Cong, S. B. & Wang, L. Cry1Ac mixed with gentamicin influences the intestinal microbial diversity and community composition of Pink bollworms. Life (Basel). 14, 58 (2023).
-
Schrøder, M. et al. A 90-day safety study of genetically modified rice expressing Cry1Ab protein (Bacillus Thuringiensis toxin) in Wistar rats. Food Chem. Toxicol. 45, 339–349 (2007).
-
Qin, H. F., Liu, Y. & Xing, F. G. Effects of Transgenic brown rice on nutritional of broiler chicken. Acta Agri Boreali-Sinica 26, 135–140 (2011).
-
Cao, Z., Gu, X. & Wang, Z. Effects of Transgenic Bt brown rice on growth, performance and some routine blood parameters in growing pigs. Acta Nutrimenta Sinica. 4, 377–382 (2014).
-
Liu, Q. et al. Effects of Transgenic Bt rice containing the Cry1Ab protein on the Gastrointestinal health of highly inbred Wuzhishan pigs after two generations of feeding. J. Agri Food Chem. 66, 10575–10587 (2018).
Acknowledgements
We thank Professor Yongjun Lin (Huazhong Agricultural University) for kindly providing transgenic rice seeds.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Gao, Y., Huang, C., Han, L. et al. Effects of genetically modified rice expressing Cry1C protein on zebrafish after a 90-day dietary intake. Sci Rep 15, 26533 (2025). https://doi.org/10.1038/s41598-025-12706-z
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12706-z