- Research
- Open access
- Published:
Biotechnology for Biofuels and Bioproducts volume 18, Article number: 77 (2025) Cite this article
Abstract
Background
Zeaxanthin is a yellow xanthophyll naturally found in plants and algae, where it plays a crucial role in light absorption and photoprotection. In mammals, ingestion of zeaxanthin through the diet is essential as it accumulates in the retina where it absorbs excessive blue light to protect photoreceptors from photooxidative stress. Chlamydomonas reinhardtii is an established model organism for pigment biosynthesis and bioengineering. Previous studies developed double knockout mutants (dzl) using CRISPR-Cas9 to eliminate ZEP and LCYE genes, achieving zeaxanthin production up to 6.84 mg/L with medium optimization. However, these approaches have not explored additional enzyme overexpression strategies combined with advanced cultivation techniques, leaving significant potential for enhanced zeaxanthin biosynthesis unexplored.
Results
In this study, we strategically enhanced zeaxanthin biosynthesis in C. reinhardtii by genome editing to knockout competing pathways coupled with overexpression of rate limiting enzymes and optimization of cultivation for efficient biomass accumulation. We employed the knockout of lycopene epsilon cyclase (LCYE; dL mutant), which resulted in a 2.83-fold increase in zeaxanthin levels. Additionally, knocking out zeaxanthin epoxidase (ZEP, dLZ mutant) redirected metabolic flux towards zeaxanthin biosynthesis, further enhancing its accumulation by 14.07-fold. Overexpression of β-carotene hydroxylase (CHYB, dLZ_C strains) enabled efficient hydroxylation of β-carotene and increasing zeaxanthin concentration further by1.80-fold without compromising growth. In addition, elevated acetate concentrations supported mixotrophic growth and resulted in a zeaxanthin yield of 21.68 ± 0.90 mg/L, threefold higher compared to previously reported values and a culminated 190-fold increase compared to the parental strain (UVM4) grown in standard medium.
Conclusion
Our study developed a zeaxanthin-producing mutant strain by integrating gene modification, gene overexpression, and culture optimization. Among various green microalgae, the engineered strain dLZ_C demonstrated notable zeaxanthin productivity, reaching 6.70 mg/L/day over a period of 3 days, suggesting its potential as a candidate for industrial production. Its improved efficiency may offer advantages for large-scale applications in microalgal-based zeaxanthin production. Additionally, these findings indicate that Chlamydomonas reinhardtii could serve as a viable and sustainable platform for biotechnological applications in the health, nutrition, and biotechnology sectors.
Background
Chlamydomonas reinhardtii is a model organism in basic research for photosynthesis [1], cilia biology [2], molecular genetics [3], cell cycle regulation [4], and stress-response mechanisms [5]. Recently, Chlamydomonas has gained recognition as a host for producing high-value chemicals, such as pigments [6,7,8]. Despite advancements in Cas9-mediated genome editing in both fundamental research and applied biotechnology [9,10,11], the genetic recalcitrance and ability for transgene silencing [12] limit stable and robust expression and remain a challenge for genetic engineering.
Previously generated UV-mutants (UVM) and advanced strategies for transgene design have been developed to enhance transgene expression [12,13,14]. These UVM strains, derived through random mutagenesis and selection, carry loss-of-function mutations in the gene encoding the NAD( +)-dependent sirtuin-type histone deacetylase (SRTA) in Chlamydomonas [12,13,14].
SRTA’s primary role is mediating the silencing of foreign DNA, such as transgenes, upon entry into the cell. Specifically, SRTA selectively represses extrachromosomal or unintegrated foreign DNA by promoting a repressive chromatin state through histone deacetylation, thereby preventing transcription of these sequences. Loss of SRTA function in UVM strains results in increased histone H3 and H4 acetylation levels and decreased H3K9me1 marks, which impedes the formation of repressive chromatin [12]. Importantly, SRTA operates without significantly affecting endogenous gene expression or genome stability, indicating its role is primarily transgene-specific rather than a broad regulator of nuclear gene expression [12]. These findings suggest that UVM strains serve as powerful platform strains for the production of high-value compounds or recombinant proteins for industrial applications [7, 13].
Carotenoid biosynthesis in green algae starts with the condensation of two molecules of geranylgeranyl pyrophosphate (GGPP; C20) by phytoene synthase activity in the chloroplast (Fig. 1a) [14]. The resulting phytoene is converted to lycopene through the activities of the two desaturases: phytoene desaturase (PDS) and ζ-carotene desaturase (ZDS), and two isomerases: 15-cis-ζ-carotene isomerase (Z-ISO) and lycopene isomerase (CrtISO). Lycopene is then cyclized by lycopene β-cyclase (LCYB) and/or lycopene ε-cyclase (LCYE) to form α-carotene (ε-ring and β-ring) or β-carotene (two β-rings) [6]. α-carotene is hydroxylated into lutein, whereas β-carotene is hydroxylated into zeaxanthin (β,β-carotene-3,3’-diol) by β-carotene hydroxylase (CHYB) or CYP97A. Zeaxanthin is epoxidized by zeaxanthin epoxidase (ZEP) to violaxanthin via antheraxanthin, and further converted to neoxanthin by neoxanthin synthase (NSY) [6].

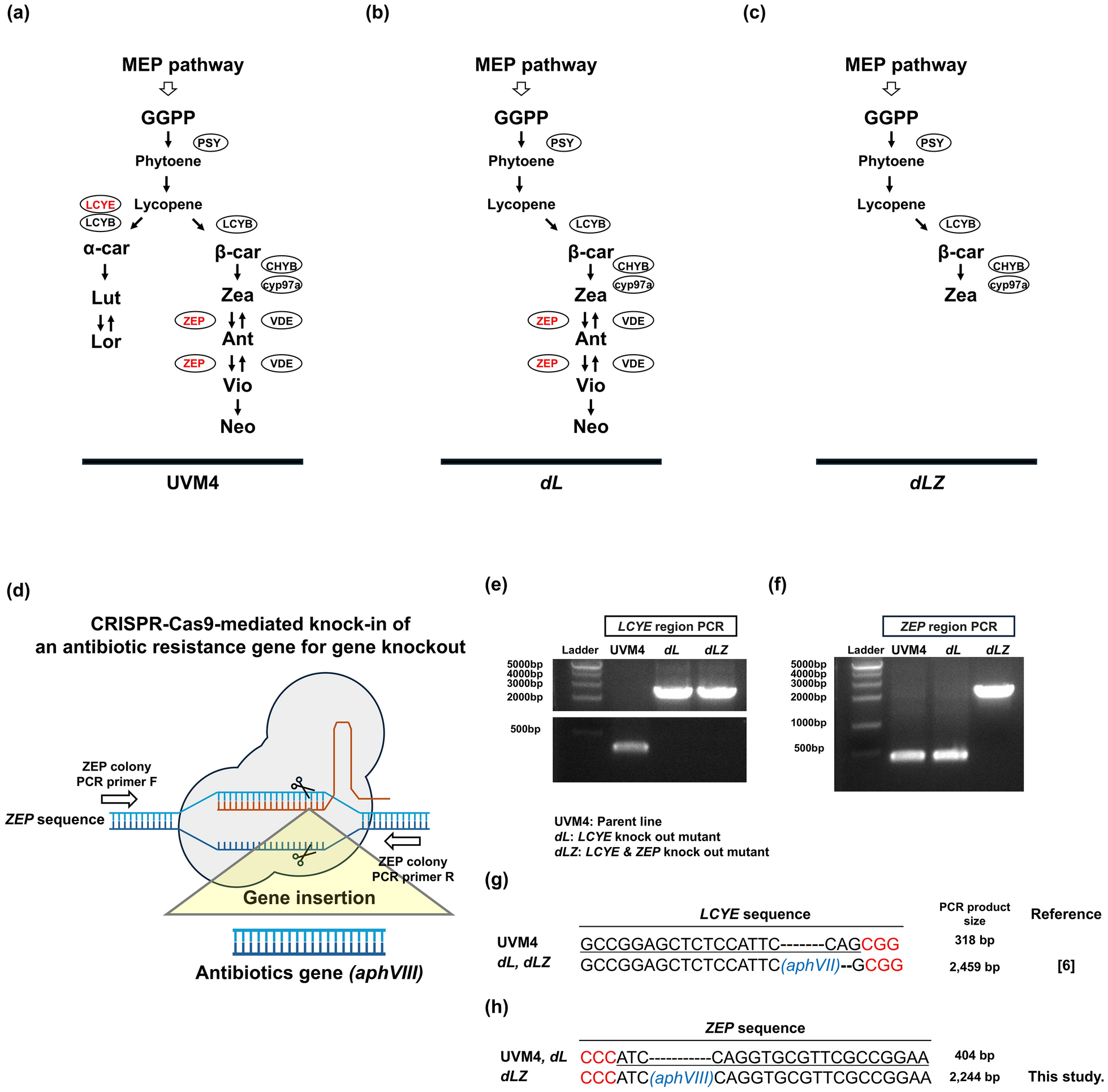
Generation of double knock out mutants of LCYE and ZEP using CRISPR-Cas9. a Carotenoid biosynthesis pathway in Chlamydomonas reinhardtii UVM4 (parent line), b dL mutant with mutation in LCYE, and c dLZ mutant with mutations in both LCYE and ZEP. d Schematic representation of the CRISPR-Cas9-mediated knock-in strategy for disrupting ZEP in the LCYE knockout mutant (dL). e Genomic PCR analysis of the LCYE and f ZEP regions. Increased PCR product sizes indicate the insertion of antibiotic resistance genes for gene knockout. g DNA sequence alignment of the LCYE and h ZEP regions, showing sgRNA target sequences (underlined), PAM sequences (highlighted in red), and inserted antibiotic resistance genes aphVII and aphVIII (marked in blue). The dL strain was derived from reference [6]. aphVII hygromycin resistance gene, aphVIII paromomycin resistance gene, Ant antheraxanthin, α-car α-carotene, β-car β-carotene, CHYB β-carotene hydroxylase, Cyp97a cytochrome p450 (carotene hydroxylases), GGPP geranylgeranyl pyrophosphate, LCYE lycopene ε-cyclase, LCYB lycopene β-cyclase, Lut lutein, Lor loroxanthin, MEP pathway 2-C-methyl-D-erythritol 4-phosphate pathway, Neo neoxanthin, PSY phytoene synthase, VDE violaxanthin de-epoxidase, Vio violaxanthin, Zea zeaxanthin, ZEP zeaxanthin epoxidase
Zeaxanthin is not synthesized in mammals [15] but plays a crucial role in the central fovea by filtering blue light and protecting photoreceptors from photooxidative stress [16, 17]. Along with lutein, zeaxanthin is an essential macular pigment for maintaining eye health, and has been extensively utilized in clinical practice [15]. Numerous studies have demonstrated that zeaxanthin improves visual performance [18, 19] and provides protection against age-related macular degeneration (AMD) [20, 21]. Intake from diets is important for human health and food supplementation has been shown to support this effect [15, 22]. In addition, zeaxanthin supports liver health by reducing oxidative stress and inflammation, and protection against nonalcoholic fatty liver disease (NAFLD) while mitigating liver fibrosis and lipid peroxidation [23]. In cardiovascular health, zeaxanthin prevents the oxidation of low-density lipoprotein, improves arterial stiffness, and supports anti-inflammatory mechanisms, thereby reducing the risk of atherosclerosis and related diseases [24, 25]. Zeaxanthin helps reduce UV-induced damage and inflammation, improves skin hydration and elasticity, and decreases lipid peroxidation [26, 27].
The main natural source of commercial zeaxanthin remains flower petals from marigold (Tagetes species) [28]. However, extraction from plant sources faces several limitations including low extraction rates, significant waste production, and environmental pollution issues [29, 30]. These challenges, combined with seasonal availability constraints and agricultural land requirements, limit the scalability of plant-based zeaxanthin production.
The increasing prevalence of age-related macular degeneration (AMD) [31, 32] and growing consumer preference for natural products have created an urgent need for sustainable, cost-effective, and environmentally friendly zeaxanthin production methods. Market data corroborates this increasing demand, with the synthetic zeaxanthin market expected to experience substantial growth—from USD 0.15 billion in 2024 to USD 0.30 billion by 2033—at a compound annual growth rate of 8.5% [33]. However, the limitations of current production methods highlight the need for alternative biotechnological approaches that can provide high-quality, bioactive zeaxanthin while addressing sustainability concerns.
Zeaxanthin production in green microalgae varies depending on species and cultivation conditions. Chromochloris zofingiensis bkt1, which has mutation in β-carotene ketolase (BKT1), accumulates up to 36.79 mg/L (7.00 mg/g DW) under high light and nitrogen starvation [34]. BKT1 normally converts β-carotene and zeaxanthin to keto-carotenoids, creating metabolic competition for zeaxanthin accumulation [6, 35]. Similarly, D. salina mp3, which has a mutation in ZEP, produces 8 mg/L (5.9 mg/g DW) under continuous low light [36]. Previously, Cas9-mediated genome editing was used to knock out LCYE in Chlamydomonas reinhardtii, producing the ΔLCYE#3 (dL) mutant, which lacks α-carotene and lutein but maintains cellular fitness and exhibits enhanced astaxanthin production capacity [6]. Similarly, functional knockout of LCYE and ZEP genes in C. reinhardtii CC-4349 (Δzl) resulted in increased zeaxanthin accumulation (5.24 mg/L, 7.28 mg/g DW) when grown in TAP medium, as shown in [8].
Despite advances in knockout-based strategies for zeaxanthin production, a critical gap remains in integrating gene knockouts with targeted enzyme overexpression and systematic cultivation optimization. Previous studies have primarily focused on individual gene knockouts, while the synergistic potential of combining these with CHYB overexpression has not been thoroughly investigated. Although zeaxanthin productivities of up to 6.84 mg/L have been reported, further enhancement is necessary to meet the rising market demand and to fully exploit the cost advantages of biotechnological production over traditional plant-based extraction. To address this, the present study establishes a comprehensive metabolic engineering framework that integrates targeted gene disruptions in the UVM4 strain with endogenous CHYB overexpression and cultivation medium optimization. This integrated approach aims to significantly enhance zeaxanthin yield and advance microalgal biotechnology toward industrial-scale, commercially viable production.
Results
Development of mutants with high zeaxanthin accumulation using CRISPR-Cas9
To achieve high levels of zeaxanthin in UVM4, a previously described LCYE knockout was used [6] and the respective mutant cell line designated as dL. This work resulted in an increased accumulation of β-branch carotenoids (β-carotene, zeaxanthin, antheraxanthin, and violaxanthin) (Fig. 1b). To further enhance zeaxanthin production, we used the CRISPR-Cas9-mediated knock-in method to knock out ZEP in the dL background, resulting in a double-knockout mutant of LCYE and ZEP, named dLZ (Fig. 1c, d). PCR amplification and subsequent sequencing analysis confirmed successful knockout of LCYE in the dL mutant and disruption of both LCYE and ZEP in the dLZ mutant (Fig. 1e–h).
The carotenoid biosynthesis pathway of C. reinhardtii is shown as Fig. 1a. The dL exhibits a modified carotenoid profile, with a deficiency in α-branch carotenoid due to the absence of LCYE (Fig. 1b). The dLZ on the other hand displays a further modified carotenoid profile, showing a deficiency in both α-branch carotenoids and downstream carotenoids from the zeaxanthin, due to the simultaneous knock out of LCYE and ZEP (Fig. 1c).
The UVM4, dL, and dLZ strains were cultured in TAP medium under fluorescent light (60–80 μmol photons m⁻2 s⁻1) for 3 days in 100 mL Erlenmeyer flasks with a 20 mL working volume, and the carotenoid profiles were analyzed (Fig. 2a). This flask setup was chosen to facilitate high-throughput screening while ensuring adequate aeration through optimal headspace-to-liquid ratio, thereby enabling efficient condition optimization with minimal reagent usage. The dL mutant, generated by knocking out LCYE, exhibited a complete absence of α-branch carotenoid (α-carotene, lutein and loroxanthin) (Fig. 1b, 2a). In contrast, the concentrations of zeaxanthin (from 0.11 ± 0.01 to 0.32 ± 0.03 mg/L), β-carotene (from 1.16 ± 0.13 to 1.54 ± 0.05 mg/L), antheraxanthin (from 0.15 ± 0.02 to 0.61 ± 0.02 mg/L) and violaxanthin (from 1.63 ± 0.21 to 3.16 ± 0.28 mg/L), were elevated in the dL mutant compared to the UVM4 (Fig. 2b, c). These findings are similar to recently described values [6]. The co-knockout of ZEP resulted in a deficiency in antheraxanthin, violaxanthin and neoxanthin synthesis in the dLZ mutant (Fig. 1c, 2a, c). Consequently, concentration of zeaxanthin (from 0.32 ± 0.03 to 4.33 ± 0.37 mg/L) increased significantly, while the concentration of β-carotene (from 1.54 ± 0.05 to 1.19 ± 0.13 mg/L) decreased compared to the dL mutants (Fig. 2b, c). Compared to the parental UVM4 strain (0.11 ± 0.01 mg/L), zeaxanthin levels increased to 2.83-fold in the dL and 38.0-fold in the dLZ. The cell numbers of UVM4 (16.25 ± 1.31 × 106 cells/ml), dL (17.38 ± 0.62 × 106 cells/ml), and dLZ (16.93 ± 0.04 × 106 cells/ml) strains were comparable under the tested culture conditions, with no significant differences observed (Fig. 2d).

Comparison of carotenoid profile and composition between UVM4, dL, and dLZ. a Carotenoid profile measured at 445 nm absorbance. b Zeaxanthin (Zea) concentration, c Concentration of α-carotene (α-car), lutein (Lut), β-carotene (β-car), antheraxanthin (Ant), and violaxanthin (Vio). d Cell numbers of UVM4, dL, and dLZ strains after 72 h of cultivation under 60–80 µmol photons m⁻2 s⁻1 fluorescent light. Experiments were performed in biological triplicates (n = 3). Values are presented as means ± standard deviation (SD). Statistical analysis was conducted using Student’s t-test. Significance levels were defined as follows: **: 0.001 < p < 0.01. Means in a row without a common superscript letter differ significantly (p < 0.05), as analyzed by one-way ANOVA followed by Tukey’s test. Chl a chlorophyll a, Chl b chlorophyll b, Car carotenoid
Overexpression of CHYB in the dLZ mutant to increase the carbon flux towards zeaxanthin
The dLZ strain generated from this study represents an ideal candidate for further improvement, as the carotenoid biosynthesis pathway is well conserved across plants and microalgae [37]. Overexpressing genes that encode key enzymes, such as β-carotene hydroxylase (CHYB), can enhance carbon flux into the carotenoid pathway, leading to increased zeaxanthin production (Fig. 3a). To achieve this, a previously described construct [35], which contains a strong psaD promoter and a synthetically optimized CHYB fused to the red fluorescent protein mRuby (Fig. 3b), was used for transformation of mutant dLZ (Fig. 3a). Three independent transformants were selected from a pool of candidates (dLZ_C1, dLZ_C2, and dLZ_C3) based on their relative fluorescence signals.

Overexpression of CHYB in dLZ to enhance carbon flux toward zeaxanthin production. a Modification of the carotenoid biosynthesis pathway to enhance zeaxanthin production through CHYB overexpression. b Schematic representation of the CHYB overexpression vector and mutant screening based on the mRuby fluorescence to OD₇₅₀ ratio. Arrows indicate selected mutant lines. c Cell numbers, d biomass, e total carotenoid concentration, f total chlorophyll concentration, g zeaxanthin concentration, and (h) β-carotene concentration were analyzed after 72 h of cultivation under 60–80 µmol photons m⁻2 s⁻1 fluorescent light. Experiments were performed in biological triplicates (n = 3). Values are presented as means ± standard deviation (SD). Statistical analysis was conducted using Student’s t-test. Significance levels were defined as follows: ***: p < 0.001, **: 0.001 < p < 0.01, *: 0.01 < p < 0.05
Among these transformants, both cell density (cells/mL) and biomass accumulation (g/L) were comparable after 3 days were comparable under the same cultivation setup (100 mL Erlenmeyer flasks with 20 mL working volume). Cell density for dLZ reached 16.24 ± 0.59 × 106 cells/mL, while dLZ_C variants ranged from 15.58 ± 1.06 × 106 (-4%) to 16.17 ± 0.68 × 106 (+ 0%) cells/mL, with a slight increase in cell density for candidate of dLZ_C2 (18.90 ± 0.84 × 106, + 16%). Similarly, biomass accumulation for dLZ was 0.65 ± 0.01 g/L, and for dLZ_C variants ranged from 0.60 ± 0.02 g/L (-7%) to 0.66 ± 0.03 g/L (+ 5%) (Fig. 3c, d).
Overexpression of CHYB led to a significant increase in total carotenoid, chlorophyll, and zeaxanthin levels, as well as an improvement in the Fv/Fm. For the total carotenoids, the dLZ reached 6.39 ± 0.85 mg/L, while the mutants exhibited levels ranging from 7.2 ± 0.75 mg/L (+ 13%) to 9.25 ± 0.68 mg/L (+ 45%) (Fig. 3e). Similarly, total chlorophyll levels increased in all mutants except for dLZ_C3, with the dLZ strain at 32.74 ± 1.01 mg/L, while mutants ranged from 34.94 ± 0.30 mg/L (+ 6%) to 38.38 ± 1.30 mg/L (+ 17%) (Fig. 3f). The zeaxanthin concentration in the dLZ strain was 4.54 ± 0.32 mg/L, whereas the mutant strains displayed significantly higher levels, ranging from 6.07 ± 0.17 mg/L (+ 34%) to 8.20 ± 0.40 mg/L (+ 80%) (Fig. 3g). Among the mutants, dLZ_C1 achieved the highest concentration of zeaxanthin (8.20 ± 0.40 mg/L), representing a 72-fold increase compared to UVM4 (0.11 ± 0.01 mg/L). β-carotene levels in most dLZ_C mutants (1.34 ± 0.12 to 1.35 ± 0.14 mg/L) remained comparable to the parent strain dLZ (1.27 ± 0.14 mg/L), with the exception of dLZ_C3 (1.06 ± 0.10 mg/L) (Fig. 3h). Among these, dLZ_C1 was selected to represent dLZ_C for further analysis in this study. The Fv/Fm ratio was measured in dLZ_C along with its parental cell lines (UVM4, dL, and dLZ), as shown in Fig. S1. The Fv/Fm ratio was lower than that of UVM4 and dL but higher than that of dLZ.
Optimization of the culture medium of the dLZ_C mutant for zeaxanthin production
Typically, C. reinhardtii is cultured in TAP medium containing 17.5 mM acetate [8]. A previous study showed that culturing Δzl strain in TAP medium with 35 mM acetate significantly enhanced both biomass and zeaxanthin production compared to standard TAP medium [8]. To optimize zeaxanthin production in the dLZ_C strain of C. reinhardtii, the effects of varying acetate concentrations (17.5–175.0 mM) in TAP medium were analyzed (Fig. 4).

Optimization of acetate concentration in TAP medium to enhance zeaxanthin production in dLZ_C. a Cell numbers from 74 to 146 h in TAP medium with acetate concentrations of 17.5–87.5 mM and b 105.0–175.0 mM. c Biomass and d zeaxanthin concentrations at 146 h in TAP medium with acetate concentrations of 17.5–175 mM. The experiment was conducted under 80–100 µmol photons m⁻2 s⁻1 LED light. Experiments were performed in biological triplicates (n = 3). Values are presented as means ± standard deviation (SD). Means in a row without a common superscript letter differ significantly (p < 0.05), as analyzed by one-way ANOVA followed by Tukey’s test
Cell densities increased with increased acetate concentration until 87.5 mM, but started to decrease from 105.0 to 175.0 mM (Fig. 4a, b). At higher acetate concentrations (70.0–175.0 mM), a substantial rise in cell numbers occurred from 74 to 98 h. The highest growth (57.8 × 106 cells/ml) of dLZ_C was observed at 87.5 mM acetate after 98 h, indicating that this concentration is optimal for maximizing cell proliferation. Biomass accumulation increased up to 105–140 mM acetate, after which it declined at 146 h (Fig. 4c). Additionally, biomass per cell and cell size exhibited an increase at high acetate concentrations (122.5–175 mM) (Fig. S2a, b).
To provide a clearer comparison of zeaxanthin production, final concentrations were analyzed at the stationary phase (146 h), where the highest yield of 21.68 mg/L was achieved at 105 mM acetate (Fig. 4d). Zeaxanthin per cell exhibited a pattern similar to biomass per cell (Fig. S2a, S2c). However, zeaxanthin content per biomass peaked at moderate acetate levels but declined significantly at 157.5–175 mM (Fig. S2d). Detailed time-course data on zeaxanthin accumulation, cell size, and biomass under varying acetate concentrations are presented in the supplementary materials (Fig. S2a, b, e, f), providing a comprehensive overview of growth and pigment production dynamics.
At higher acetate concentrations (105.0–175.0 mM), both biomass and zeaxanthin production plateaued or slightly declined (Fig. 4c, d). To better understand these reductions, we examined acetate consumption and monitored associated changes in pH and salinity (Fig. S3). Residual acetate was measured at 98 h to assess acetate uptake (Fig. S3a). The results showed that acetate was almost completely consumed up to 140.0 mM. However, at higher concentrations of 157.5 and 175.0 mM, residual acetate levels increased substantially to approximately 7.58 mM and 72.10 mM, respectively, indicating that acetate uptake capacity becomes limited at these elevated concentrations.
In conjunction with acetate consumption, pH changes were also monitored throughout the cultivation period. A progressive increase in pH was observed, with a maximum ΔpH of + 3.9 at 70.0 mM acetate, However, at elevated acetate levels (87.5–175.0 mM), the extent of pH elevation diminished, with only a ΔpH of + 2.3 recorded at 175.0 mM (Fig. S3b). Additionally, salinity measurements using a refractometer showed corresponding increases under high acetate conditions (Fig. S3c). This elevated salinity may have imposed additional osmotic stress on the cells, further inhibiting cellular activity and contributing to the observed declines in biomass and zeaxanthin production.
To assess the broader applicability of optimized conditions, UVM4, dLZ, and dLZ_C strains were cultivated in TAP medium with 105 mM acetate and analyzed after 98 h. In terms of cell numbers, all three strains exhibited comparable values: UVM4 (42.31 ± 4.19 × 106 cells/mL), dLZ (40.39 ± 1.82 × 106 cells/mL), and dLZ_C (46.72 ± 4.34 × 106 cells/mL) (Fig. S4a). Pigment concentration analysis revealed that zeaxanthin production in UVM4 was minimal reaching only 0.25 ± 0.07 mg/L, whereas dLZ exhibited a substantial increase, producing 13.27 ± 1.13 mg/L, which corresponds to a 52-fold increase compared to UVM4. Among mutants, dLZ_C demonstrated the highest zeaxanthin yield, reaching 21.23 ± 0.70 mg/L, representing an 83-fold increase relative to UVM4 (Fig. S4b).
Discussion
C. reinhardtii synthesizes various xanthophylls, among which zeaxanthin is well known for its eye health benefits and other physiological advantages [17]. However, the zeaxanthin biosynthesis in plants or algae is highly dependent on light intensity and low-light conditions typically induce rapid conversion into violaxanthin by ZEP via the intermediate antheraxanthin [38]. The disruption of ZEP enables continuous zeaxanthin accumulation, regardless of light exposure, leading to a mutant strain in which zeaxanthin serves as the terminal xanthophyll in the β-branch of the xanthophyll biosynthetic pathway [9, 39].
Additionally, to eliminate competition from the α-branch of the xanthophyll biosynthetic pathway, LCYE was knocked out, effectively redirecting metabolic flux toward β-carotene synthesis [8]. Our results showing increased xanthophyll cycle carotenoids (2.9-fold for zeaxanthin, 4.1-fold for antheraxanthin, and 1.9-fold for violaxanthin) are consistent with the original characterization by Kneip et al. [6], who observed similar increases of 1.9-fold, 2.25-fold, and 1.8-fold for the same carotenoids, respectively, confirming the reproducibility of this metabolic redirection without compromising cellular fitness.
The newly generated double knockout mutant dLZ strain, with both LCYE and ZEP knockouts in the UVM4 background, exhibited high levels of zeaxanthin production (4.33 mg/L, 6.73 mg/g) after 3 days under mixotrophic conditions (Fig. 2a and b). This performance is comparable to the previously reported dzl strain with identical genetic modifications in the CC-4349 background, which achieved 5.24 mg/L and 7.28 mg/g zeaxanthin production [8]. In contrast, zeaxanthin production in UVM4 was barely detectable under the same conditions (Fig. 2b). The cell densities of UVM4, dL, and dLZ strains were comparable, with no significant differences observed (Fig. 2d). These results align with findings from a previous study [8], further validating our strategy for zeaxanthin enhancement in C. reinhardtii. These results also confirm the effectiveness of metabolic engineering and the precision of CRISPR-RNP-based gene editing in C. reinhardtii.
The dLZ strain derived in this study originated from the UVM4 strain. This makes it an ideal candidate for further enhancement by increasing carbon flux into the xanthophyll pathway through the overexpression of a key enzyme like CHYB, thereby further improving zeaxanthin production (Fig. 3a). Recent studies have demonstrated that overexpressing the CHYB gene significantly enhances hydroxylation activity and redirects carbon flux toward the xanthophyll pathway in engineered microbial strains. When CHYB is co-overexpressed with BKT, these enzymes work synergistically to efficiently convert carotenes (β-carotene and canthaxanthin) into their corresponding xanthophylls (zeaxanthin and astaxanthin, respectively). This enzymatic cooperation ultimately leads to substantially increased astaxanthin production in the engineered strains [6, 35]. However, this study provides evidence that CHYB overexpression demonstrates hydroxylation activity through the increased zeaxanthin concentration in double knockout mutants, where zeaxanthin serves as the terminal carotenoid.
Interestingly, the CHYB overexpression not only increased carotenoid levels (Fig. 3e) but also significantly elevated total chlorophyll concentrations in dLZ_C (Fig. 3f), resulting in a substantial improvement in photosynthetic efficiency, as indicated by Fv/Fm (Fig. S1) compared to dLZ. In contrast, the photosynthetic capacity of dLZ cultured in HS medium was significantly lower than that of UVM4, retaining only 67% of its efficiency (Fig. S1). This result is consistent with a previous study [40], which reported that Δzl strains—sharing a similar genetic background—exhibit reduced photosynthetic efficiency due to lower chlorophyll pigment levels. The elevated zeaxanthin concentration of dLZ_C shown in Fig. 3g could be a direct result of CHYB activity, and the increase in total carotenoid levels appears to be primarily attributed to zeaxanthin accumulation. Generally, β-carotene, the substrate of CHYB, is expected to decrease as zeaxanthin levels increase. In this study, β-carotene concentration remained constant between dLZ and dLZ_C in these conditions (Fig. 3h). This suggests that, despite β-carotene serving as a substrate in the β-branch of xanthophyll biosynthesis, its unchanged levels indicate possible alterations of upstream carotenoid biosynthesis in response to CHYB activity at this culture condition. It is likely that in the CHYB-overexpressing strain (dLZ_C), the increase in total carotenoids contributes to thylakoid membrane stabilization [40, 41] or prevents chlorophyll degradation through its intrinsic antioxidant properties [42], thereby maintaining higher chlorophyll concentrations than dLZ line. As a result, dLZ_C may sustain higher photosynthetic efficiency compared to the dLZ strain.
C. reinhardtii can grow photoautotrophically under light conditions or photomixotrophically by utilizing acetate as an additional carbon source in the presence of light. As shown in Fig. 4, our results indicate that acetate concentration plays a crucial role in modulating growth patterns and zeaxanthin production in this strain. At low to moderate acetate concentrations (17.5–105.0 mM), dLZ_C exhibited enhanced biomass production, suggesting that elevated acetate levels effectively support cell growth. These findings align with previous studies on C. reinhardtii, Chlorella sorokiniana, and Scenedesmus obliquus, which reported similar acetate-induced growth stimulation [43,44,45].
Notably, at higher acetate concentrations (105.0–175.0 mM), biomass production of dLZ_C plateaued or slightly declined (Fig. 4c), potentially due to acetate-induced metabolic stress [46] and/or salinity-associated toxicity [47]. Although absolute zeaxanthin production increased up to 105 mM acetate, the conversion efficiency, defined as zeaxanthin yield per gram of acetate supplied, declined significantly (Fig. 4d). A similar trend was observed for biomass yield (Fig. 4c), suggesting that beyond a certain threshold, increasing acetate concentration does not proportionally improve productivity. This decline coincided with a marked increase in residual acetate levels above 140.0 mM (Fig. S3a), implying that acetate uptake becomes inefficient, likely due to transporter saturation or feedback inhibition in central metabolic pathways. The reduced pH elevation at high acetate levels (Fig. S3b) further supports this interpretation, since alkalization of the medium is generally associated with active acetate assimilation. The minimal ΔpH observed at 175.0 mM likely reflects reduced metabolic activity or increased cellular stress. In parallel, salinity also increased with acetate concentration (Fig. S3c), likely due to sodium accumulation from acetate supplementation combined with salts inherently present in TAP medium. This rise in osmotic pressure may have further compromised cell physiology and pigment biosynthesis [47]. Moreover, previous studies have shown that high acetate concentrations can directly suppress photosynthetic CO₂ assimilation in C. reinhardtii, even under saturating light conditions [48], suggesting that acetate imposes multiple modes of metabolic inhibition. Collectively, these findings indicate that while moderate acetate supplementation (70–105 mM) supports optimal biomass growth and zeaxanthin production, excessive acetate levels trigger metabolic and osmotic stress responses that impair carbon utilization efficiency and pigment biosynthesis.
This stress response is further reflected in the pattern of zeaxanthin accumulation. Zeaxanthin accumulation peaked at 70–105 mM acetate, yet at higher concentrations (> 105 mM), total zeaxanthin production declined while biomass remained relatively stable, resulting in a decreased zeaxanthin-to-biomass ratio (Fig. 4c, d, S2d). This shift indicates metabolic redirection toward intracellular storage compounds rather than zeaxanthin synthesis. The reduced cell number combined with increased cell size suggests stress-induced metabolic adaptation, where fewer but larger cells are produced (Fig. S2a-c). Consequently, zeaxanthin content per cell increased significantly under high acetate conditions (Fig. S2c), as the reduced zeaxanthin production was distributed among fewer cells. This suggests that excess acetate triggers stress responses, enhancing storage molecule accumulation as protective mechanisms while compromising zeaxanthin biosynthesis. Therefore, for applications aiming to maximize biomass production while maintaining a moderate zeaxanthin yield, acetate concentrations should be maintained within the 70–105 mM range. In contrast, higher acetate levels (> 105 mM) can enhance zeaxanthin accumulation per cell, but this comes at the expense of reduced biomass yield, requiring careful evaluation.
The dLZ_C strain developed in this study has several advantages over other green algae. A recent study reported a newly developed C. zofingiensis bkt1 that achieved a zeaxanthin yield of 36.79 mg/L within 12 days of cultivation, with a productivity of 3.07 mg/L/day [34]. In contrast, although the maximum zeaxanthin yield of dLZ_C (21.68 mg/L) was lower than that of the C. zofingiensis bkt1, its productivity (6.70 mg/L/day) was more than twice as high, indicating a faster production rate (Fig. S2e). One major limitation of Chlorella in pigment extraction is its thick cell wall, which often results in incomplete and labor-intensive pigment extraction [49, 50]. In contrast, dLZ_C, as a cell wall-less strain, facilitates a more efficient and less labor-intensive extraction process. Furthermore, the absence of lutein in dLZ_C eliminates the need for additional purification steps, thereby enhancing overall process efficiency [8]. Therefore, dLZ_C can be a promising alternative candidate to conventional sources for industrial-scale zeaxanthin production and is expected to be advantageous, particularly in applications requiring efficient and cost-effective downstream processing.
Conclusion
Our study successfully demonstrates the potential of Chlamydomonas reinhardtii as a robust microbial chassis for zeaxanthin biosynthesis through rational metabolic engineering. By disrupting competing pathways (LCYE and ZEP mutations) and enhancing metabolic flux via CHYB overexpression, we successfully developed the dLZ_C strain, which demonstrated an unprecedented 190-fold increase in zeaxanthin yield compared to the non-engineered parental strain (UVM4). With optimized acetate supplementation, dLZ_C achieved a productivity of 6.70 mg/L/day within 74 h (Fig. S2e) and zeaxanthin production of 21.68 ± 0.90 mg/L at 98 h (Table 1). This represents a 3.2-fold improvement over previously reported C. reinhardtii dzl mutants (6.84 mg/L) and significantly exceeds production levels from other eukaryotic green microalgal systems, positioning our strain among the most productive zeaxanthin-producing eukaryotic green microalgae documented to date. Overall, our findings confirm that stepwise metabolic engineering is an effective approach to enhance zeaxanthin production, and dLZ_C is a promising candidate for the commercial-scale production of high-purity zeaxanthin. Moving forward, our effort will focus on scaling up the cultivation system to ensure sustained zeaxanthin production and developing a cost-effective biorefinery process for efficient zeaxanthin extraction.
Material and methods
Strains and growth conditions
The C. reinhardtii strain UVM4 [51] and a previously generated dL line (Δ LCYE#3) [6] were routinely maintained on solid TRIS–acetate-phosphate (TAP) medium with pH 7.2 modified from [52] (Table S1) at 40–60 μmol photons m−2 s−1 fluorescent white light.
All cultivation experiments were conducted in 20 mL of medium within 100 mL Erlenmeyer flasks at 25 °C, with constant shaking at 120–130 rpm under continuous light. Fluorescent white light at 40–60 μmol photons m⁻2 s⁻1 was used for mutant analysis, while LED light at 80–100 μmol photons m⁻2 s⁻1 was used for acetate optimization. A starting cell density of 1 × 10⁶ cells/mL was used.
TAP medium supplemented with acetate was prepared by adding acetic acid and adjusting the pH to 7.2 with sodium hydroxide. Cell number and size were monitored using an automatic cell counter (EVE TM plus) without trypan blue dye staining. Biomass (dry cell weight) was measured gravimetrically by harvesting 5 mL cell culture and lyophilization in a pre-weighed 1.5 mL reaction tube [40]. Salinity was measured using a salinity refractometer (ATAGO, MASTER-S/MillM, Cat. 2493), and acetic acid concentration was determined using an Acetic Acid Assay Kit (Megazyme, Reference code: K-ACET, SKU: 700004254).
CRISPR-mediated genome editing and selection of mutants
Gene editing was based on the delivery of pre-assembled Cas9-sgRNA ribonucleoproteins (RNPs) and double stranded donor-DNA via electroporation [53]. The sgRNA binding site of ZEP of C. reinhardtii was previously identified [9]. The GeneArt™ Precision sgRNA Synthesis Kit (Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA) was used according to the manufacture’s protocol for sgRNA synthesis followed by purification using RNA Clean up. Donor DNA was amplified from the aphVIII cassette in a previously designed vector (pPEARL GenBank: KU531882.1) using target specific oligonucleotides (forward 5’-gtactagtggtacccgcttcaaatacgc-3’; reversed 5’-cggagatccacacacctgc-3’) followed by extraction from agarose gel.
C. reinhardtii transformation was performed by electroporation as previously described [8, 54, 55]. Briefly, a total of 7.0 × 107 cells were prepared for transformation. After washing with TAP-sucrose (40 mM) medium, 250 µL of cell suspension was incubated at 40 °C for 30 min. During this period, functional RNPs were assembled by mixing 28 μg sgRNA with 40 μg spCas9, followed by incubation at room temperature for 15 min. The cells were then incubated with pre-assembled CRISPR complexes and DNA for 5 min. Electroporation (600 V, 50 µF, 200 Ω) was performed in 4 mm electrode gap cuvettes. After 15 min at room temperature, the cells were transferred to 6-well microtiter plates containing 5 mL of TAP for overnight incubation for recovery. Subsequently, the cells were plated on TAP agar plates with appropriate antibiotics (30 μg/mL paromomycin). After an incubation period of 5 to 10 days, the colonies were transferred to 96-well plates for screening. Colony PCR was performed using a sufficient number of cells with the Chelex method [56]. Cells (50 µL) were harvested and resuspended in 50 µL of 5% Chelex-100. The mixture was incubated at 95 °C for 30 min, then centrifuged at 13,000 rpm for 10 min. The supernatant was used as a template. KOD One™ PCR Master Mix and the specified primers (Table S2) were used to amplify the target site.
Nuclear transformation of C. reinhardtii and fluorescence screening
The CHYB overexpression vector was previously designed [6, 35]. C. reinhardtii transformation was performed by electroporation with some modification from previously described [8]. A total of 1 µg of linearized plasmid DNA (XbaI, KpnI; Takara) without purification was used for transformation of 1 × 10⁷ cells. After washing the cells with TAP-sucrose (40 mM) medium, 250 µL of cell suspension was incubated with the linearized DNA for 5 min in a 4 mm cuvette. Following electroporation (600 V, 50 µF, 200 Ω) and a 15-min recovery period, the transformed cells were transferred to a 6-well plate containing 5 mL of TAP-sucrose (40 mM) medium for overnight incubation before plating onto TAP agar containing 10 µg/mL nourseothricin. After an incubation period of 5 to 10 days, the colonies were subsequently transferred to 96-well plates for screening. Cultured cells at sufficient cell density in 96-well plates were transferred to clear bottom 96-well plate (for optical density [OD] at 750 nm) and 96-well black plates (for fluorescence) [57, 58]. The OD750 nm and fluorescence of mRuby2 (excitation, 560 nm; emission, 600 nm) were measured using a Varioskan Flash microplate reader (Thermo Fisher Scientific). The fluorescence intensity was normalized to the OD750 nm of the sample [57]. Cell lines with a high fluorescence-to- OD750 nm ratio were selected.
Measurement of chlorophyll and total carotenoid
Total chlorophyll and carotenoids were extracted using 80% (v/v) acetone. The extracts were centrifuged (13,000 rpm, 5 min) to remove the cell debris, and the absorbance was measured at wavelengths of 663, 647, and 470 nm. The total chlorophyll and carotenoid contents were calculated using Lichtenthaler’s formula [59], with the specific calculations performed using Eq. (1), (2), (3), and (4).
$${text{Chlorophyll a }}left( {{text{mg}}/{text{L}}} right) , = { 12}.{25 } times A_{663 , nm} {-}{ 2}.{79 } times A_{647 , nm}$$
(1)
$${text{Chlorophyll b }}left( {{text{mg}}/{text{L}}} right) , = { 21}.{5}0 , times A_{647 , nm} {-}{ 5}.{1}0 , times A_{663 , nm}$$
(2)
$${text{Total chlorophyll}}left( {{text{mg}}/{text{L}}} right) = {text{Chlorophylla}}left( {{text{mg}}/{text{L}}} right) + {text{Chlorophyll b }}left( {{text{mg}}/{text{L}}} right)$$
(3)
$${text{Total carotenoid }}left( {{text{mg}}/{text{L}}} right) , = , left( {left( {{1}000 , times A_{470 , nm} } right) , {-} , left( {{1}.{82 } times Chl , a} right) , {-} , left( {{85}.0{2 } times Chl , b} right)} right)/{198}$$
(4)
Pigment analysis by HPLC
The pigments were analyzed as previously described [15]. Pigments were extracted using 90% (v/v) acetone and analyzed using a Shimadzu Prominence HPLC system (Model LC-20AD, Shimadzu, Kyoto, Japan) fitted with a Spherisorb 5.0 μm ODS1 column (4.6 × 250 mm, Waters, Milford, USA). The absorbance was measured at 445 nm using a photodiode array detector.
Photosynthetic activity measurements
Cells were cultured in TAP medium for 2 days and then transferred to HS medium for an additional 2 days. A total of 2 × 10⁷ cells were harvested (2,500 rpm, 5 min), suspended in 0.2 mL of HS medium, and transferred to a 96-well black-bottom plate. The cells were incubated in the dark for 20 min. Fv/Fm was measured using an Imaging-PAM M-series fluorometer (Heinz Walz, Germany) equipped with an IMAG-K4 CCD camera and an IMAG-MAX/L LED array (blue light at 450 nm).
Statistical analysis
Data were statistically analyzed using Student’s t-test or one-way ANOVA, as described by Assad [60]. Tukey’s multiple comparison test was used to assess the significance of the differences among the means. The data are expressed as mean ± standard deviation (SD).
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its Supplementary material 1.
Abbreviations
- β-car:
-
β-Carotene
- CHYB:
-
β-Carotene hydroxylase
- dL :
-
LCYE Knock out mutant in UVM4
- dLZ :
-
LCYE And ZEP knock out mutant in UVM4
- dLZ_C :
-
LCYE And ZEP knock out mutant with overexpression of CHYB in UVM4
- LCYB:
-
Lycopene beta cyclase
- LCYE:
-
Lycopene epsilon cyclase
- Zea:
-
Zeaxanthin
- ZEP:
-
Zeaxanthin epoxidase
References
-
Dupuis S, Merchant SS. Chlamydomonas reinhardtii: a model for photosynthesis and so much more. Nat Methods. 2023;20(10):1441–2. https://doi.org/10.1038/s41592-023-02023-6.
-
Silflow CD, Lefebvre PA. Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii. Plant Physiol. 2001;127(4):1500–7. https://doi.org/10.1104/pp.010807.
-
Crozet P, Navarro FJ, Willmund F, Mehrshahi P, Bakowski K, Lauersen KJ, Pérez-Pérez M-E, Auroy P, Gorchs Rovira A, Sauret-Gueto S. Birth of a photosynthetic chassis: a MoClo toolkit enabling synthetic biology in the microalga Chlamydomonas reinhardtii. ACS Synth Biol. 2018;7(9):2074–86. https://doi.org/10.1021/acssynbio.8b00251.
-
Spudich JL, Sager R. Regulation of the Chlamydomonas cell cycle by light and dark. J Cell Biol. 1980;85(1):136–45. https://doi.org/10.1083/jcb.85.1.136.
-
de Carpentier F, Lemaire SD, Danon A. When unity is strength: the strategies used by Chlamydomonas to survive environmental stresses. Cells. 2019;8(11):1307. https://doi.org/10.3390/cells8111307.
-
Kneip JS, Kniepkamp N, Jang J, Mortaro MG, Jin E, Kruse O, Baier T. CRISPR/Cas9-mediated knockout of the lycopene ε-cyclase for efficient astaxanthin production in the green microalga Chlamydomonas reinhardtii. Plants. 2024;13(10):1393. https://doi.org/10.3390/plants13101393.
-
Perozeni F, Cazzaniga S, Baier T, Zanoni F, Zoccatelli G, Lauersen KJ, Wobbe L, Ballottari M. Turning a green alga red: engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. Plant Biotechnol J. 2020;18(10):2053–67. https://doi.org/10.1111/pbi.13364.
-
Song I, Kim J, Baek K, Choi Y, Shin B, Jin E. The generation of metabolic changes for the production of high-purity zeaxanthin mediated by CRISPR-Cas9 in Chlamydomonas reinhardtii. Microb Cell Fact. 2020;19:1–9. https://doi.org/10.1186/s12934-020-01480-4.
-
Baek K, Kim DH, Jeong J, Sim SJ, Melis A, Kim J-S, Jin E, Bae S. DNA-free two-gene knockout in Chlamydomonas reinhardtii via CRISPR-Cas9 ribonucleoproteins. Sci Rep. 2016;6(1):30620. https://doi.org/10.1038/srep30620.
-
Ferenczi A, Chew YP, Kroll E, von Koppenfels C, Hudson A, Molnar A. Mechanistic and genetic basis of single-strand templated repair at Cas12a-induced DNA breaks in Chlamydomonas reinhardtii. Nat Commun. 2021;12(1):6751. https://doi.org/10.1038/s41467-021-27004-1.
-
Jeong B-R, Jang J, Jin E. Genome engineering via gene editing technologies in microalgae. Bioresour Technol. 2023;373:128701. https://doi.org/10.1016/j.biortech.2023.128701.
-
Neupert J, Gallaher SD, Lu Y, Strenkert D, Segal NA, Barahimipour R, Fitz-Gibbon ST, Schroda M, Merchant SS, Bock R. An epigenetic gene silencing pathway selectively acting on transgenic DNA in the green alga Chlamydomonas. Nat Commun. 2020;11(1):6269. https://doi.org/10.1038/s41467-020-19983-4.
-
Lauersen KJ, Berger H, Mussgnug JH, Kruse O. Efficient recombinant protein production and secretion from nuclear transgenes in Chlamydomonas reinhardtii. J Biotechnol. 2013;167(2):101–10. https://doi.org/10.1016/j.jbiotec.2012.10.010.
-
Razz SA. Comprehensive overview of microalgae-derived carotenoids and their applications in diverse industries. Algal Res. 2024. https://doi.org/10.1016/j.algal.2024.103422.
-
Song I, Kim S, Kim J, Oh H, Jang J, Jeong SJ, Baek K, Shin W-S, Sim SJ, Jin E. Macular pigment-enriched oil production from genome-edited microalgae. Microb Cell Fact. 2022;21(1):27. https://doi.org/10.1186/s12934-021-01736-7.
-
Barker FM, Snodderly DM, Johnson EJ, Schalch W, Koepcke W, Gerss J, Neuringer M. Nutritional manipulation of primate retinas, V: effects of lutein, zeaxanthin, and n–3 fatty acids on retinal sensitivity to blue-light–induced damage. Invest Ophthalmol Vis Sci. 2011;52(7):3934–42. https://doi.org/10.1167/iovs.10-5898.
-
Mares J. Lutein and zeaxanthin isomers in eye health and disease. Annu Rev Nutr. 2016;36(1):571–602. https://doi.org/10.1146/annurev-nutr-071715-051110.
-
Renzi LM, Hammond BR. The effect of macular pigment on heterochromatic luminance contrast. Exp Eye Res. 2010;91(6):896–900. https://doi.org/10.1016/j.exer.2010.09.015.
-
Hammond BR, Fletcher LM, Elliott JG. Glare disability, photostress recovery, and chromatic contrast: relation to macular pigment and serum lutein and zeaxanthin. Invest Ophthalmol Vis Sci. 2013;54(1):476–81. https://doi.org/10.1167/iovs.12-10411.
-
Wu J, Cho E, Willett WC, Sastry SM, Schaumberg DA. Intakes of lutein, zeaxanthin, and other carotenoids and age-related macular degeneration during 2 decades of prospective follow-up. JAMA Ophthalmol. 2015;133(12):1415–24. https://doi.org/10.1001/jamaophthalmol.2015.3590.
-
Bernstein PS, Li B, Vachali PP, Gorusupudi A, Shyam R, Henriksen BS, Nolan JM. Lutein, zeaxanthin, and meso-zeaxanthin: the basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog Retin Eye Res. 2016;50:34–66. https://doi.org/10.1016/j.preteyeres.2015.10.003.
-
Mrowicka M, Mrowicki J, Kucharska E, Majsterek I. Lutein and zeaxanthin and their roles in age-related macular degeneration—neurodegenerative disease. Nutrients. 2022;14(4):827. https://doi.org/10.3390/nu14040827.
-
Chamberlain SM, Hall JD, Patel J, Lee JR, Marcus DM, Sridhar S, Romero MJ, Labazi M, Caldwell RW, Bartoli M. Protective effects of the carotenoid zeaxanthin in experimental nonalcoholic steatohepatitis. Dig Dis Sci. 2009;54:1460–4. https://doi.org/10.1007/s10620-009-0824-2.
-
Mosca L, Rubenfire M, Mandel C, Rock C, Tarshis T, Tsai A, Pearson T. Antioxidant nutrient supplementation reduces the susceptibility of low density lipoprotein to oxidation in patients with coronary artery disease. J Am Coll Cardiol. 1997;30(2):392–9. https://doi.org/10.1016/s0735-1097(97)00188-5.
-
Wang W, Connor SL, Johnson EJ, Klein ML, Hughes S, Connor WE. Effect of dietary lutein and zeaxanthin on plasma carotenoids and their transport in lipoproteins in age-related macular degeneration. Am J Clin Nutr. 2007;85(3):762–9. https://doi.org/10.1093/ajcn/85.3.762.
-
González S, Astner S, An W, Pathak MA, Goukassian D. Dietary lutein/zeaxanthin decreases ultraviolet B-induced epidermal hyperproliferation and acute inflammation in hairless mice. J Invest Dermatol. 2003;121(2):399–405. https://doi.org/10.1046/j.1523-1747.2003.12355.x.
-
Palombo P, Fabrizi G, Ruocco V, Ruocco E, Fluhr J, Roberts R, Morganti P. Beneficial long-term effects of combined oral/topical antioxidant treatment with the carotenoids lutein and zeaxanthin on human skin: a double-blind, placebo-controlled study. Skin Pharmacol Physiol. 2007;20(4):199–210. https://doi.org/10.1159/000101807.
-
Bourdon L, Jensen AA, Kavanagh JM, McClure DD. Microalgal production of zeaxanthin. Algal Res. 2021;55: 102266. https://doi.org/10.1016/j.algal.2021.102266.
-
Song J, Yang Q, Huang W, Xiao Y, Li D, Liu C. Optimization of trans lutein from pumpkin (Cucurbita moschata) peel by ultrasound-assisted extraction. Food Bioprod Process. 2018;107:104–12. https://doi.org/10.1016/j.fbp.2017.10.008.
-
Manikandan R, Thiagarajan R, Goutham G, Arumugam M, Beulaja M, Rastrelli L, Skalicka-Woźniak K, Habtemariam S, Orhan IE, Nabavi SF. Zeaxanthin and ocular health, from bench to bedside. Fitoterapia. 2016;109:58–66. https://doi.org/10.1016/j.fitote.2015.12.009.
-
Deng Y, Qiao L, Du M, Qu C, Wan L, Li J, Huang L. Age-related macular degeneration: epidemiology, genetics, pathophysiology, diagnosis, and targeted therapy. Genes Dis. 2022;9(1):62–79. https://doi.org/10.1016/j.gendis.2021.02.009.
-
Korva-Gurung I, Kubin A-M, Ohtonen P, Hautala N. Incidence and prevalence of neovascular age-related macular degeneration: 15-year epidemiological study in a population-based cohort in Finland. Ann Med. 2023;55(1):2222545. https://doi.org/10.1080/07853890.2023.2222545.
-
Synthetic zeaxanthin market size and projections [Internet]. 2024 [cited 9 Jun 2025]. https://www.marketresearchintellect.com/product/global-synthetic-zeaxanthin-market-size-and-forecast/.
-
Huang W, Lin Y, He M, Gong Y, Huang J. Induced high-yield production of zeaxanthin, lutein, and β-carotene by a mutant of Chlorella zofingiensis. J Agric Food Chem. 2018;66(4):891–7. https://doi.org/10.1021/acs.jafc.7b05400.
-
Amendola S, Kneip JS, Meyer F, Perozeni F, Cazzaniga S, Lauersen KJ, Ballottari M, Baier T. Metabolic engineering for efficient ketocarotenoid accumulation in the green microalga Chlamydomonas reinhardtii. ACS Synth Biol. 2023;12(3):820–31. https://doi.org/10.1021/acssynbio.2c00616.
-
Kim M, Ahn J, Jeon H, Jin E. Development of a Dunaliella tertiolecta strain with increased zeaxanthin content using random mutagenesis. Mar Drugs. 2017;15(6):189. https://doi.org/10.3390/md15060189.
-
Chen H-H, Liang M-H, Ye Z-W, Zhu Y-H, Jiang J-G. Engineering the β-carotene metabolic pathway of microalgae Dunaliella to confirm its carotenoid synthesis pattern in comparison to bacteria and plants. Microbiol Spectr. 2023;11(2):e04361-e14322. https://doi.org/10.1128/spectrum.04361-22.
-
Bethmann S, Melzer M, Schwarz N, Jahns P. The zeaxanthin epoxidase is degraded along with the D1 protein during photoinhibition of photosystem II. Plant Direct. 2019;3(11): e00185. https://doi.org/10.1002/pld3.185.
-
Baek K, Yu J, Jeong J, Sim SJ, Bae S, Jin E. Photoautotrophic production of macular pigment in a Chlamydomonas reinhardtii strain generated by using DNA-free CRISPR-Cas9 RNP-mediated mutagenesis. Biotechnol Bioeng. 2018;115(3):719–28. https://doi.org/10.1002/bit.26499.
-
Kim M, Cazzaniga S, Jang J, Pivato M, Kim G, Ballottari M, Jin E. Photoautotrophic cultivation of a Chlamydomonas reinhardtii mutant with zeaxanthin as the sole xanthophyll. Biotechnol Biofuels Bioprod. 2024;17(1):41. https://doi.org/10.1186/s13068-024-02483-8.
-
Park S, Fischer AL, Steen CJ, Iwai M, Morris JM, Walla PJ, Niyogi KK, Fleming GR. Chlorophyll-carotenoid excitation energy transfer in high-light-exposed thylakoid membranes investigated by snapshot transient absorption spectroscopy. J Am Chem Soc. 2018;140(38):11965–73. https://doi.org/10.1021/jacs.8b04844.
-
Paulsen H. Pigment ligation to proteins of the photosynthetic apparatus in higher plants. Physiol Plant. 1997;100(4):760–8. https://doi.org/10.1111/j.1399-3054.1997.tb00003.x.
-
Chen C-Y, Ho S-H, Liu C-C, Chang J-S. Enhancing lutein production with Chlorella sorokiniana Mb-1 by optimizing acetate and nitrate concentrations under mixotrophic growth. J Taiwan Inst Chem Eng. 2017;79:88–96. https://doi.org/10.1016/j.jtice.2017.04.020.
-
Zhang Z, Tan Y, Wang W, Bai W, Fan J, Huang J, Wan M, Li Y. Efficient heterotrophic cultivation of Chlamydomonas reinhardtii. J Appl Phycol. 2019;31:1545–54. https://doi.org/10.1007/s10811-018-1666-0.
-
Liu J, Yin J, Ge Y, Han H, Liu M, Gao F. Improved lipid productivity of Scenedesmus obliquus with high nutrient removal efficiency by mixotrophic cultivation in actual municipal wastewater. Chemosphere. 2021;285: 131475. https://doi.org/10.1016/j.chemosphere.2021.131475.
-
Qiao H, Wang G, Liu K, Gu W. Short-term effects of acetate and microaerobic conditions on photosynthesis and respiration in chlorella sorokiniana gxnn 01 (chlorophyta) 1. J Phycol. 2012;48(4):992–1001. https://doi.org/10.1111/j.1529-8817.2012.01189.x.
-
Fields FJ, Ostrand JT, Mayfield SP. Fed-batch mixotrophic cultivation of Chlamydomonas reinhardtii for high-density cultures. Algal Res. 2018;33:109–17. https://doi.org/10.1016/j.algal.2018.05.006.
-
Heifetz PB, Forster B, Osmond CB, Giles LJ, Boynton JE. Effects of acetate on facultative autotrophy in Chlamydomonas reinhardtii assessed by photosynthetic measurements and stable isotope analyses. Plant Physiol. 2000;122(4):1439–46. https://doi.org/10.1104/pp.122.4.1439.
-
Cha KH, Lee HJ, Koo SY, Song D-G, Lee D-U, Pan C-H. Optimization of pressurized liquid extraction of carotenoids and chlorophylls from Chlorella vulgaris. J Agric Food Chem. 2010;58(2):793–7. https://doi.org/10.1021/jf902628j.
-
Luengo E, Condón-Abanto S, Álvarez I, Raso J. Effect of pulsed electric field treatments on permeabilization and extraction of pigments from Chlorella vulgaris. J Membr Biol. 2014;247:1269–77. https://doi.org/10.1007/s00232-014-9688-2.
-
Neupert J, Karcher D, Bock R. Generation of Chlamydomonas strains that efficiently express nuclear transgenes. Plant J. 2009;57(6):1140–50. https://doi.org/10.1111/j.1365-313X.2008.03746.x.
-
Gorman DS, Levine R. Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardi. PNAS. 1965;54(6):1665–9. https://doi.org/10.1073/pnas.54.6.1665.
-
Kim J, Lee S, Baek K, Jin E. Site-specific gene knock-out and on-site heterologous gene overexpression in Chlamydomonas reinhardtii via a CRISPR-Cas9-mediated knock-in method. Front Plant Sci. 2020;11: 509810. https://doi.org/10.3389/fpls.2020.00306.
-
Picariello T, Hou Y, Kubo T, McNeill NA, Yanagisawa H-A, Oda T, Witman GB. TIM, a targeted insertional mutagenesis method utilizing CRISPR/Cas9 in Chlamydomonas reinhardtii. PLoS ONE. 2020;15(5):e0232594. https://doi.org/10.1371/journal.pone.0232594.
-
Freudenberg RA, Wittemeier L, Einhaus A, Baier T, Kruse O. The spermidine synthase gene SPD1: A novel auxotrophic marker for Chlamydomonas reinhardtii designed by enhanced CRISPR/Cas9 gene editing. Cells. 2022;11(5):837. https://doi.org/10.3390/cells11050837.
-
Cao M, Fu Y, Guo Y, Pan J. Chlamydomonas (chlorophyceae) colony PCR. Protoplasma. 2009;235:107–10. https://doi.org/10.1007/s00709-009-0036-9.
-
Rasala BA, Barrera DJ, Ng J, Plucinak TM, Rosenberg JN, Weeks DP, Oyler GA, Peterson TC, Haerizadeh F, Mayfield SP. Expanding the spectral palette of fluorescent proteins for the green microalga Chlamydomonas reinhardtii. Plant J. 2013;74(4):545–56. https://doi.org/10.1111/tpj.12165.
-
Lauersen KJ, Baier T, Wichmann J, Wördenweber R, Mussgnug JH, Hübner W, Huser T, Kruse O. Efficient phototrophic production of a high-value sesquiterpenoid from the eukaryotic microalga Chlamydomonas reinhardtii. Metab Eng. 2016;38:331–43. https://doi.org/10.1016/j.ymben.2016.07.013.
-
Wellburn A, Lichtenthaler H, editors. Formulae and program to determine total carotenoids and chlorophylls a and b of leaf extracts in different solvents. In: SpringerPlus Photosynthesis, Brussels, Belgium, August 1–6, Springer; 1983, vol 2. https://doi.org/10.1007/978-94-017-6368-4_3.
-
Assaad HI, Zhou L, Carroll RJ, Wu G. Rapid publication-ready MS-Word tables for one-way ANOVA. Springerplus. 2014;3:474. https://doi.org/10.1186/2193-1801-3-474.
Acknowledgements
The authors would like to express thanks to Prof. Ralph Bock for providing the UVM4 strain and Yoongil Lee for helping to screen the double-knockout mutant of LCYE and ZEP (dLZ). This research was supported by the Basic Science Research Program (NRF-2020R1A2C2011998) and the National Research Foundation (NRF) funded by the Korean government (MSIT) (No. RS-2024-00458958) to E.J.
Funding
Basic Science Research Program (NRF-2020R1A2C2011998) and the National Research Foundation (NRF) funded by the Korean government (MSIT) (No. RS-2024-00458958).
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Jang, J., Baier, T., Kneip, J.S. et al. High-yield zeaxanthin production in Chlamydomonas reinhardtii via advanced metabolic pathway engineering. Biotechnol. Biofuels Bioprod. 18, 77 (2025). https://doi.org/10.1186/s13068-025-02676-9
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1186/s13068-025-02676-9