
Introduction
Ticks (order Ixodida) are vectors of arthropod pathogens of public health and veterinary importance in tropical and subtropical regions of the world1. Ticks are associated with direct and indirect limitations to livestock health and productivity2. Ticks transmit zoonotic pathogens belonging to Babesia spp., Borrelia spp., Rickettsia spp. Ehrlichia spp., Francisella tularensis, Coxiella burnetii, and viruses such as Nairovirus (Bunyaviridae) that causes Crimean-Congo hemorrhagic fever (CCHF) and tick-borne encephalitis virus (Flaviviridae)3,4,5,6. Therefore, controlling the transmission of ticks is necessary to prevent the prevalence of the disease7. To reduce the impact of ticks on livestock health as well as eliminate their negative effects on public health, acaricides, manual tick removal, animal movement control and vaccination are often applied to control ticks8. Until now, the use of acaricides has been the most effective measure of tick and tick-borne diseases (TBDs) for control9. However, the ever-increasing cost of acaricides in less developed countries, which are most affected by ticks and TBDs, as well as their environmental impact and contamination of milk and meat products with drug residues, suggests that acaricides cannot be routinely used for tick control10. In addition, excessive use and abuse of acaricides causes resistance in the tick population11. In light of this, future tick control strategies must depend on the integration of economically, environmentally and vaccination efficient applications12. Due to the lack of cooperation of some Asian and African countries that have high levels of livestock tick infestation, and in addition to challenges such as the desired effect of vaccines against different tick species based on the genetic diversity of ticks to date, there has been very limited development of anti-tick vaccines. For example, the only commercially available recombinant vaccine against Rhipicephalus microplus (R. microplus) under the brand name GAVAC and TickGARD were developed more than 2 decades ago13,14. These vaccines were designed based on the BM86 gene of R. microplus which has 1950 nucleic acids with 650 amino acids that are coding for a protein with 86 kDa. The Bm86 gene is a glycoprotein in mid gut membrane cells in ticks that have a critical role in digestive and blood feeding14,15.
Several studies have demonstrated the potential of immunological approaches to tick control based on physiological processes. Existing tick vaccines act by stimulating humoral and cellular responses against tick midgut cell membrane antigens. Despite the many advantages of tick control through vaccination, this strategy is currently ineffective due to antigenic sequence variations between tick populations and geographically isolated species that result in vaccine resistance in some areas [and impaired efficacy in others]. These limitations necessitate the search for alternative antigens for the development of next-generation vaccines16.
In the 1990s, antigen-based vaccines were developed that could provide immune protection against ticks. Vaccine development based on latent antigens is attractive because the host and parasite do not have an evolved immune interaction. These antigens are not normally exposed to the host and therefore do not play a role in naturally acquired immunity17,18. These analyses concern the identity and predicted binding sites of MHC-I and II, as well as comparing N-glycosylation in addition to the amount of cysteine in the epidermal growth factor-like domain. The results of this study provide a new potential target antigen for producing a vaccine against Iranian ticks in cattle19. In the early 1990s, vaccines based on concealed antigens were developed that could induce immunological protection against tick infestations in the wild and in humans and other animals that were infected with the virus20. Development of concealed antigens-based vaccines is attractive because the host and parasite would not have evolved immunological interactions. These antigens are not normally exposed to the host and therefore do not usually play a role in naturally acquired immune responses to the host antigen17,21. The aim of this study was to design in silico a recombinant protein vaccine targeting the HM86 protein of the tick Hyaloma marginatum (order Ixodididae) as a potential anti-tick vaccine for the control of tick-borne diseases with an emphasis on Crimean-Congo fever.
Materials and methods
Evolutionary analysis of HM 86 Hyaloma marginatum
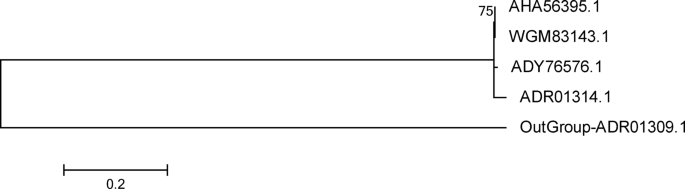
In the first step, the protein sequences of the HM 86 gene of Hyalomma marginatum available in the NCBI and UniProt databases were extracted, then FASTA sequences with accession numbers ADR01314.1, AHA56395.1, ADY76576.1, WGM83143.1 were selected, and ADR01309.1 was also saved as an outgroup, and finally aligned using MEGA version.6.0 software to draw a phylogenetic tree.
Antigenicity prediction
The VaxiJen server (version 2.0) was used to predict the antigenicity of the protein, using the bacterial model with a threshold of 0.4. Proteins with high antigenicity scores were selected for further analysis21,22.
Predict disulfide bonds and domains of BM86 protein
SSpro 4.0 server was used to predict the number of domains and the number of protein disulfide bonds based on the protein secondary structure sequence of Hm86.
Allergenicity prediction
AllergenFP version 1.1 was used to predict allergenicity. This online bioinformatics tool utilizes a novel descriptor-based fingerprint approach to identify potential allergens23.
Protein physico-chemical and functional analysis
Various physicochemical properties of the proteins were predicted using the ProtParam server23. Parameters estimated included molecular weight (MW), stability index, isoelectric point (IP), half-life in vitro and in vivo, aliphatic index, and the grand average of hydropathicity (GRAVY). Additionally, conserved domains and important functional sites within the proteins were identified using InterPro24.
Docking of synthetic TLR-2 and Hm86 protein
Hm86 is the most important surface protein of the stomach of Hyalomma marginatum. In the present study, it was considered as a ligand. The Hm86 sequence was extracted from NCBI and then, docking with TLR2 and Hm86 protein was performed using the HDOCK online server. Finally, their docking energy and RMSD were obtained.
Docking of modeled TLR-2 and Hm86 by HADDOCK 2.4 server
Molecular binding is a process that involves the interaction between a ligand molecule and a receptor molecule to form a stable adduct. Also, the calculated score was provided as a measure of the degree of binding interaction25. Therefore, the structure of TLR2 was used as the receptor (PDBID: 3A7B) and the vaccine (Hm86) protein as the ligand26. Finally, the binding affinity of the vaccine structure to TLR-2 (PDB ID: 3A7B) was assessed by molecular docking using the HADDOCK 2.4 server. Both the TLR-2 receptor PDB files and the vaccine structure were uploaded to this server. The server finds the model with the lowest electrostatic energy by applying the Fourier correlation algorithm to the models. The most efficient vaccine-TLR-2 complex was selected for further molecular dynamics simulation analysis based on its low binding energy27.
Binding free energy calculations by MM-GBSA
Binding free energy calculations were performed using the MM-GBSA to quantify the binding affinity beyond the raw docking for the HM86 protein with the TLR2 receptor. Protein-peptide binding was re-scored by measuring the overall binding free energy ΔGbind (MM-GBSA) to predict the binding affinity of HM86 peptides to the TLR2 receptor protein. Also, the per-residue free energy decomposition of their most important interacting residues was calculated. Then, more quantitative variables related to the energy contribution by ligands and receptor residues (with high negative energy values) were analyzed using principal component analysis with the help of the following R packages: (https://cran.r-project.org) including factoextra, surveyminer, ggpubr, ggcorrplot, fastqcr.
Prediction of the Bcell epitope
B-cell epitopes, recognized by the B-lymphocyte receptor, play a very important role in enhancing the immune response of organisms. In this study, B-cell epitopes were predicted using the ABCpred server28. The mentioned server can predict and display epitopes using an artificial neural network. In this server, epitopes from various organisms such as bacteria, viruses, and parasites are present with an accuracy of 65.9%.
MHC-I and MHC-II binding epitope prediction
The Rankpep online server was used to search and identify MHC-I and MHC-II binding regions. binding threshold Scores greater than 17.5 for MHC-I and 42.5 for MHC-II were considered as predicted binders for the selected MHC alleles.
Results
Selection Hm86 gene Hyalomma marginatum (Antigen) and evaluation of evolutionary analysis
The current study was focused on predicting the vaccine candidates for Hm86-Hyalomma marginatum anti-tick vaccine by a computational reverse vaccinology approach for which a total of 664 amino acids of Hm86 (ADR01314.1) were extracted from the UniProt database in FASTA format. These protein sequences were analyzed for their antigenic properties that potentially induce immunogenicity. The phylogeny tree shows a cladogram for Hm86 protein (Fig. 1). A phylogenetic tree was constructed using sequences from the Bm86 gene that had been deposited in GenBank by other researchers. The neighbor-joining algorithm was used to construct the cladogram. Gene Bm86 of hyalomma marginatum shows a relatively close relationship to gene Bm86 Rhipicephalus microplus.

Phylogenetic analysis of the selected Hm86 proteins. Evolutionary analyses were performed in MEGA6.0 software. The strain of Dermacentor variabilis homologous to Bm86 (ADR01309.1) was used as an out group.
Antigenicity prediction
By submitting the Hm86 protein sequence to the VaxiJen version 2.0 bioinformatics tool, the analysis yielded an overall protective antigen prediction score of 0.7402, indicating that the protein is a probable antigen.
Predict disulfide bonds and domains of BM86 protein
The Hm86 protein comprises three domains: domain 1 spans amino acids 1–178, domain 2 covers residues 179–557, and domain 3 includes residues 558–664. Additionally, the protein contains 15 disulfide bonds, as detailed in Table 1.
Evaluation of allergenicity
The AllergenFP server version 1.0 was used to assess the allergenicity of the vaccine. Based on its results, the vaccine is hypoallergenic and its Tanimoto coefficient is 0.5495867768595041, meaning that the vaccine is designed based on the most similar classified protein of the potential allergen.
Evaluation of physico-chemical characteristics
Based on the ProtParam server results, the Hm86 protein consists of 664 amino acids with a calculated molecular weight of 72.3 kDa. The predicted isoelectric point (IP) of 5.96 indicates that the protein is nearly neutral. The estimated half-life of the protein is approximately 30 h in mammalian reticulocytes, over 20 h in yeast, and more than 10 h in Escherichia coli. With an instability index of 42.00, the protein is classified as unstable. The aliphatic index and grand average of hydropathicity (GRAVY) were calculated to be 62.45 and − 0.424, respectively. A high aliphatic index suggests that the protein is stable across a wide temperature range, while the negative GRAVY value indicates hydrophilicity, implying good interaction with surrounding water molecules.
Docking of synthetic TLR-2 and Hm86 protein
Figure 2 shows the binding of TLR2 to Hm86 protein. Their binding energy and RMSD are also given in Table 2. Based on the findings, the binding energy and RMSD range of the selected modeled is from − 283.23 to -242.95 kcal/mL and 96.50 to 81.50 angstroms, respectively. The best result was for model number 1 because it has the most negative docking score.
Roughly speaking, the probability of two molecules binding is high when the confidence score is above 0.7; it is possible when the confidence score is between 0.5 and 0.7; and it is low when the confidence score is less than 0.5. However, due to the empirical nature of the confidence score, it is better to study it carefully.
Docking of modeled TLR-2 and Hm86 by HADDOCK 2.4 server
Ticks are blood-feeding arthropods that can parasitize vertebrates for short periods of time. Tick saliva contains biologically active molecules, and suppression of the host immune response via the TLR2 pathway may contribute to tick feeding and pathogen transmission29. TLR2 activation induces the recruitment of adaptor molecules such as TIRAP, MyD88, IRAKs, and TRAF6, and activates MAPK, IKK, and NF-B, as well as the regulation of genes such as inducible nitric oxide synthase (iNOS), tumor necrosis factor (TNF-α), and tumor necrosis factor-α (TNF-α)30. TLR2 agonists that have a higher binding affinity for TLR2 can therefore induce both cellular and humoral immune responses. Using the HADDOCK web server, an energy-optimized 3D model of the potential vaccine structure (Fig. 2) was docked to TLR2 to assess whether the designed vaccine and the innate immune receptor interacted with each other. The results of molecular docking between the Hm86 protein as a ligand and TRL4 as a receptor showed that the binding occurs well, and here the first 5 residues that have the highest binding are shown in Tables 3 and 4, and the position of each amino acid is also shown in Fig. 2. Overall, the statistics obtained from the HADDOCK servers indicate that the vaccine structure is completely intact with the TLR2 protein. Reveal high-ranking binding compounds for protein-protein complexes (Fig. 2).

Molecular docking of Hm86 -TLR2 constructed by HDOCK server. The ligand is shown in green and the receptor in blue.
Binding free energy calculations by MM-GBSA

MM-GBSA-Binding free energy calculations.
From the first 10 models of binding free energies obtained based on MM-GBSA, model number 1 of the TRL2 and Hm86 complex was selected with a binding free energy of -283.23 kcal/mol, which is obtained from the combination of ELE (electrostatic energy), VDW (van der Waals interaction energy), SA (surface area for estimating the free energy of nonpolar solvation) and GB (generalized Born model for estimating the free energy of polar solvation) which constitute the total binding free energy.
The binding free energy based on MM-GBSA for the model number one complex was 29.9 kcal/mol, where the highest energy contribution due to VDM was − 57.62 kcal/mol, the results are shown in Fig. 3. Based on the molecular docking performed as shown in Table 4, five residues, such as B-GLU-172, B-TRP-144, B-ASN-174, B-THR-189, B-THR-131, had strong affinity for the TRL2 receptor through electrostatic, hydrophobic interactions and two or more hydrogen bonds. Their binding energies were − 2.63, -2.21, -2.03, -1.77 and − 1.73 kcal/mol, respectively.
Identification of the Bcell epitope
Antigens in Hm86 induce antibody production and activate memory B cells through Linear B cell epitopes in the adaptive immune system. In the present study, predicted B-cell epitopes for Yi using the ABCPred server. A peptide score above the threshold (0.51) means that the sequence is more likely to be an epitope. All peptides shown in Table 5 were selected above the threshold value.(Table 5).
Major histocompatibility complex (MHC) MHC-I and MHC-II binding epitope prediction
124 MHC-I binding epitopes were predicted for the Hm86 protein sequence by the RANKPEP server. Of the predicted epitopes, 4 sequences, shown in Table 6 along with their features, had higher scores based on the defined binding threshold of 17.50 and the predicted binding threshold of 17.50. In addition, the RANKEPB server was again used to predict MHC-II binding epitopes from the Hm86 protein sequence. Finally, 215 epitopes were predicted in a wide range of scores, and the number of 5 initial sequences in which none of them met the connection threshold score (42.50) is shown in Table No. 7. (Table 7)
Discussion
Hyalomma marginatum is a hard tick that widely infests cattle, causing reduced livestock production and transmitting various pathogens. One of the effective ways to control ticks is to use recombinant vaccines. Due to the different varieties, there is a need to produce and develop specific vaccines to protect livestock in each geographical area. Bm86 antigens can effectively protect livestock from tick infestation. Based on immunological information approaches, a recombinant vaccine for ticks was analyzed in this study, which provides a promising path for the future development of an efficient antigenic vaccine for the control of Hyalomma marginatum tick, the major vector of Crime-Congo fever. Based on in silico antigenic analysis of the complete genome of Bm86, it does not cause any allergic reactions in humans. In addition, the hyalomma marginatum protein in this study design has both MHC-1 and MHC-2 epitopes, so it may have a greater effect than single-antigen-based vaccines. Several studies have reported the advantages and experimental validity of recombinant vaccines using these immunological tools31,32. Several vaccines have been developed using this approach against various infectious diseases, and have now been experimentally validated and approved for clinical trials33,34. Hyalomma marginatum proteins were identified and characterized using online B-cell servers. Epitopes are unique regions present on proteins that are important in stimulating immune responses at both the cellular and humoral levels35. The Bm86 gene sequence of the Hyalomma marginatum tick was evaluated in silico and was shown to have non-allergenic and antigenic properties. In silico results of HM protein showed that it is non-allergenic and antigenic. Also, the physicochemical parameters of this proposed vaccine have 644 amino acids that encode a protein with a molecular weight of 73.2 kDa. With a theoretical pI value of 5.96, it confirms the prediction that the protein is neutral in the environment. Younas, M. studies in 2024 reported similar results for the BM86 protein. This potential vaccine candidate (HM86) has hydrophilic properties, as the values of the aliphatic index and the average hydropathy (GRAVY) of the vaccine were evaluated as 62.45 and − 0.424, respectively. Similar results were reported in a study conducted in 2024, which showed a GRAVY value of -0.22835. The in-silico analysis also showed that the antigen is non-allergenic, heat-stable and hydrophobic. Figure 2 shows that the structure of the Bm86 vaccine from Hyalomma marginatum is located in the C-terminal region of TLR2, which exhibits several hydrophobic and vdW bond contacts. In the Bm86-TLR2 binding and overlap, amino acids Y376, E375 K383 play a key role in forming H-bonds. In current study, HDOCK server was used to perform docking. Other studies conducted by Shabani et al. in 2022 on calculating the docking of the monkeypox used the same servers36. In the present study, the binding affinity of the HM86 protein to TLR2 was performed using a web server and showed that the potential vaccine candidate (HM86) with high binding affinity is likely to activate the immune system response in the animal. A similar study was conducted for the BM86 protein and showed that the interactions of the BM86 vaccine structure have a binding affinity to TLR2 with higher affinity for TLR2, which may induce the acquired and innate immune responses of the immune system to activate35. finally, the putative Bm86 vaccine has the ability to stimulate both humoral and cellular immune responses based on the analyses performed. Bm86 is antigenic and non-allergenic. In this study, the structure of the Bm86 vaccine was obtained using immunological approaches. However, additional in vivo and in vitro evaluations are needed to confirm the vaccinability. The recombinant Bm86 protein from Hyalomma marginatum is currently being evaluated by researchers in the laboratory.
Conclusion
The use of vaccines for immunological control is one of the effective methods for controlling tick infestation in cattle. Accordingly, vaccines produced with recombinant Bm86 protein have good efficacy, if the recombinant vaccine is a protein from a specific tick species in a specific region, they are much more effective in combating ticks in that region. Because there is polymorphism in the Bm86 protein gene. Of course, numerous efforts have been made by researchers to search for new proteins, antigens based on other alleles of the Bm86 gene or orthologous genes that can stimulate an immune response in cattle and allow the creation of more effective recombinant vaccines. Countries such as Colombia and Mexico have designed vaccines from national or local tick strains that have shown promising results in those countries. The effectiveness of the antigen on local tick alleles shows that it is very important to improve the effectiveness of vaccines for the production of anti-tick vaccines. This research aimed to use bioinformatics-based information security technologies to develop a recombinant protein vaccine against hyalomma marginatum. A promising vaccine was validated by molecular docking, molecular dynamics simulation. As a result, studies on multi-antigenic vaccines have been underdeveloped to date and have been limited to controlled trials due to difficulties in antigen formulation. Hence, it is important for researchers to address these issues in future studies to effectively combat tick-borne diseases.
Data availability
The datasets used and/or analysed during the current study are available from the corresponding author upon reasonable request.
References
-
Jongejan, F. & UILENBERG, G. The global importance of ticks. Parasitology 129 (S1), S3–S14 (2004).
-
Wikel, S. K. Ticks and tick-borne infections: complex ecology, agents, and host interactions. Veterinary Sci. 5 (2), 60 (2018).
-
Kassiri, H., Dehghani, R., Kasiri, M. & Kasiri, M. D. R. A review on the reappearance of Crimean-Congo hemorrhagic fever, a tick-borne Nairovirus. Entomol. Appl. Sci. Lett. 7 (1-2020), 81–90 (2020).
-
Nasiri, Z. et al. Bionomics and ecological characteristics of hard ticks of Ixodidae in Fars province, Southwestern Iran. Exp. Appl. Acarol. 94 (3), 39 (2025).
-
Nasiri, Z. et al. Molecular identification of Coxiella burnetii and Anaplasma Ovise in ticks isolated from the livestock of Fars province, Iran. Egypt. J. Veterinary Sci. 1–9. (2025).
-
Mohammadi, J. et al. Moemenbellah-Fard, frequency of pyrethroid resistance in human head louse treatment: systematic review and meta-analysis. Parasite 28, 86 (2021).
-
de La Fuente, J., Kocan, K. & Contreras, M. Prevention and control strategies for ticks and pathogen transmission. Rev. Sci. Tech. 34 (1), 249–264 (2015).
-
Chizyuka, H. & Mulilo, J. Methods currently used for the control of multi-host ticks: their validity and proposals for future control strategies. Parassitologia 32 (1), 127–132 (1990).
-
Domingos, A., Antunes, S., Borges, L. & Rosario, V. E. Approaches towards tick and tick-borne diseases control. Rev. Soc. Bras. Med. Trop. 46, 265–269 (2013).
-
Makala, L., Mangani, P., Fujisaki, K. & Nagasawa, H. The current status of major tick borne diseases in Zambia. Vet. Res. 34 (1), 27–45 (2003).
-
Jabbar, A. et al. Tick-borne diseases of bovines in pakistan: major scope for future research and improved control. Parasites Vectors. 8, 1–13 (2015).
-
Ghosh, S., Azhahianambi, P. & Yadav, M. Upcoming and future strategies of tick control: a review. J. Vector Borne Dis. 44 (2), 79 (2007).
-
Rodriguez, M. et al. Control of boophilus Microplus populations in grazing cattle vaccinated with a Recombinant Bm86 antigen Preparation. Vet. Parasitol. 57 (4), 339–349 (1995).
-
Abood, A., Salman, A. M., El-Hakim, A. E., Abdel-Aty, A. M. & Hashem, A. M. Purification and characterization of a new thermophilic collagenase from nocardiopsis dassonvillei NRC2aza and its application in wound healing. Int. J. Biol. Macromol. 116, 801–810 (2018).
-
Kamau, L. M. Isolation, Recombinant Expression and Characterization of Polymorphism of BM86 Vaccine Antigen Homologues from Rhipicephalus Appendiculatus (Kenyatta University Nairobi, Kenya, 2005).
-
Kasaija, P. D. et al. Inspiring anti-tick vaccine research, development and deployment in tropical Africa for the control of cattle ticks: review and insights. Vaccines 11 (1), 99 (2022).
-
de la Fuente, J. et al. Prevalence of tick-borne pathogens in Ixodid ticks (Acari: Ixodidae) collected from European wild Boar (Sus scrofa) and Iberian red deer (Cervus Elaphus hispanicus) in central Spain. Eur. J. Wildl. Res. 50 (4), 187–196 (2004).
-
De La Fuente, J. & Kocan, K. Strategies for development of vaccines for control of Ixodid tick species. Parasite Immunol. 28 (7), 275–283 (2006).
-
Parthasarathi, B. Evaluation of Cross-protective Efficacy of Multi-antigen Co-Immunization against Rhipicephalus Microplus and Hyalomma Anatolicum (Indian Veterinary Research Institute, 2021).
-
Zinkernagel, R. M. On natural and artificial vaccinations. Annu. Rev. Immunol. 21 (1), 515–546 (2003).
-
de la Fuente, J. & Kocan, K. M. Advances in the identification and characterization of protective antigens for Recombinant vaccines against tick infestations. Expert Rev. Vaccines. 2 (4), 583–593 (2003).
-
Doytchinova, I. A. & Flower, D. R. VaxiJen: a server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 8, 1–7 (2007).
-
Gasteiger, E. et al. Protein identification and analysis tools on the ExPASy server, The proteomics protocols handbook 571–607. (2005).
-
Finn, R. D. et al. The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 44 (D1), D279–D285 (2016).
-
Lengauer, T. & Rarey, M. Computational methods for biomolecular Docking. Curr. Opin. Struct. Biol. 6 (3), 402–406 (1996).
-
Bishop, L. J., Stutzer, C. & Maritz-Olivier, C. More than three decades of Bm86: what we know and where to go. Pathogens 12 (9), 1071 (2023).
-
Rahman, M. M., Masum, M. H. U., Parvin, R., Das, S. C. & Talukder, A. Designing of an mRNA vaccine against high-risk human papillomavirus targeting the E6 and E7 oncoproteins exploiting immunoinformatics and dynamic simulation. PloS One. 20 (1), e0313559 (2025).
-
Malik, A. A., Ojha, S. C., Schaduangrat, N. & Nantasenamat, C. ABCpred: a webserver for the discovery of acetyl-and butyryl-cholinesterase inhibitors. Mol. Diversity 1–21. (2022).
-
Kotál, J. et al. Modulation of host immunity by tick saliva. J. Proteom. 128, 58–68 (2015).
-
Greyling, N. & Ricinus-Infested Comparative Immune-Profiling of Tick Vaccine Formulations and the Role of Co-Infestation With Babesia Microti in Ixodes BALB/c Mice, University of Pretoria (South Africa), (2019).
-
Zheng, B. et al. Towards an ensemble vaccine against the pegivirus using computational modelling approaches and its validation through in Silico cloning and immune simulation. Vaccines 9 (8), 818 (2021).
-
Kalita, P., Padhi, A. K., Zhang, K. Y. & Tripathi, T. Design of a peptide-based subunit vaccine against novel coronavirus SARS-CoV-2. Microb. Pathog. 145, 104236 (2020).
-
Caro-Gomez, E., Gazi, M., Goez, Y. & Valbuena, G. Discovery of novel cross-protective rickettsia prowazekii T-cell antigens using a combined reverse vaccinology and in vivo screening approach. Vaccine 32 (39), 4968–4976 (2014).
-
Mehla, K. & Ramana, J. Identification of epitope-based peptide vaccine candidates against enterotoxigenic Escherichia coli: a comparative genomics and immunoinformatics approach. Mol. Biosyst. 12 (3), 890–901 (2016).
-
Younas, M. et al. RESEARCH ARTICLE construction of multi-epitope vaccine against the rhipicephalus microplustick: an immunoinformatics approach. Trop. Biomed. 41 (1), 84–96 (2024).
-
Shabani, S., Rashidi, M., Radgoudarzi, S. & Jebali, A. The validation of artificial anti-monkeypox antibodies by in Silico and experimental approaches. Immun. Inflamm. Dis. 11 (4), e834 (2023).
Acknowledgements
The authors thank the Research and Technology of Shiraz University of Medical Sciences, Iran. This article is the result of Project No. 27148 with Code of Ethics IR.SUMS.SCHEANUT.REC.1401.139 approved by Shiraz University of Medical Sciences.
Funding
The research was funded by Shiraz University of Medical Sciences with Code 27148.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Shahriari-Namadi, M., Azizi, K. & Alipour, H. In-silico study: Hm86 protein of Hyalomma marginatum (Ixodida) as a candidate to provide anti-tick vaccine. Sci Rep 15, 27008 (2025). https://doi.org/10.1038/s41598-025-12871-1
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12871-1