Introduction
Cauliflower (Brassica oleracea var. botrytis L.) belongs to the family of Brassicaceae. Also, many studies revealed that there are more than 350 genera and about 4000 species1 and their varieties share a common C genome (n = 9)2 often known by the name “cole vegetables”3.
The edible part of a cauliflower is curd, and botanically known as the pre-floral fleshy apical meristem or immature inflorescence4. Cauliflower is a vegetable with good nutritional value, as its dry matter content is 11.6%5, and it is considered a source of fiber, proteins, vitamin B complex and many minerals, especially potassium and calcium. Regular consumption of cruciferous vegetables (cauliflower, cabbage, broccoli) helps reduce the risk of chronic heart diseases and cancer6.
Worldwide, the cultivation area of cauliflower has reached about 1357.19 thousand hectares with a production of more than 25 million tons, and its production in the Arab countries reaches 0.683 million tons with an area of 26.53 thousand hectares7.
However, the demand for hybrids of cauliflower has increased in the last years, due to maturity earliness, high yield, and resistance to biotic and abiotic stresses. An ideal hybrid is produced by crossing two diverse inbred lines which are produced either through self-pollination or doubled haploid (DH) technology. But in cauliflower, self-pollination is restricted due to self-incompatibility (SI) and cytoplasmic male sterility (CMS)8 so needs a large number of generations to attain homozygosity. Thus, the viable option to produce inbred lines is through haploid production. The development of an inbred line through the DH technique will require less time and have complete homozygosity as compared to an inbred line produced through self-pollination8,9. Haploid plants can either be produced from the male floral part i.e., androgenesis or the female floral part i.e., gynogenesis. In Brassica, androgenesis is preferred for haploid production10. Anther culture technique is the most viable and efficient method of producing homozygous doubled haploid plants within a short period11,12. The production of haploid plants by anthers culture in cruciferous vegetables was first reported for common cabbage Brassica oleracea var. capitate by Kameya and Hinata in 197010, while Ockendo succeeded in producing DH cauliflower lines from anthers culture in 198813.
The successful production of androgenic plantlets largely depends on the type of species, genotypes, physiology of the donor plants, stage of development of microspores/pollen, composition of culture medium, carbon source, sucrose levels, plant growth hormones and pretreatment temperatures14,15.
Cultural media composition influences the androgenic process in a great way. A complete nutrient media composition containing all the mineral salts, sucrose and vitamins is required for androgenesis. The most commonly used basal media compositions in androgenesis are MS medium16 B5 medium17 Nitsch and Nitsch18 and N619.
In a study of Fuller and Turton (1989)20 to investigate the response of winter cauliflower cultivars grown in Britain to anthers culture revealed that 13 of the 15 cultivars were responded. As well, they also explained that successfulness of anthers culture varied and depended on the cultivar and medium culture, and the percentage of embryos obtained by this method ranged from zero to 16%.
So, Moreover, Yang et al. (1992)21in another study to determine the factors affecting the success of in vitro cauliflower anther culture, they were found that adding cytokine BAP at different concentrations (0.02, 0.1, 1) mg/L to the culture medium led to a reduction in embryo production, and the negative effect increased with increasing hormone concentration in the medium.
Furthermore, Cilingir et al. (2017)22 studied three cultivars of red cabbage to investigate the effectiveness of in vitro anther culture technology, the flower buds to be used for anther culture were taken with a length of 3–5 mm and then cultured on two nutrient media, MS and B5, supplemented with 2.4D, NAA, glutamine, serine, silver nitrate, IAA, BAP and different concentrations of sucrose to stimulate the formation of callus, embryos and haploid plants, their results showed that the culture medium, sucrose concentration and genotypes have an important role in the formation of haploid embryos.
Also, Sood et al. (2019)23 were carried out their experiment to study the callus induction from anthers culture and they used three Brassica campestris cultivars (HPBS-1, KBS-3, HPKM-04), MS and N6 mediums, two levels of sucrose (3% and 4%), and three different hormonal combinations HM1 (0.5 mg/L 2,4-D) HM2 (0.5 mg/L 2,4-D + 1.0 mg/L NAA) HM3 (1.0 mg/L NAA), the results showed that HPBS-1 gave the best induction when cultivated on N6 medium with a hormonal combination of 0.5 mg/L 2,4-D + 1.0 mg/L NAA and 3% sucrose.
The in vitro regeneration of cauliflower through anther culture technique isn’t used in our country yet, so, there is an indispensable need for studying the anther culture technique for improvement of local types of cauliflower. Therefore, the present study was undertaken to observe the androgenic responses of five Syrian local types of cauliflower and the control, Casper F1 to anther culture, and determine the ideal levels of plant growth regulators of MS medium.
Results and discussion
Callus induction
Analysis of variance for callus induction performances of the types, medium composition and their interactions are presented in Tables 1, 2 and 3. In this study, callus induction frequency among the types and days taken for callus induction showed a statistically significant effect between types (Table 1). Our results showed that the maximum value of callus induction frequency was obtained in Gtyp1 (54.44%), and significantly was higher than all other types, followed by Casper F1 and Gtyp3 with the value of callus induction 47.04 and 38.15%, respectively. As well, the minimum value was observed in Gtyp2 (16.67%). Also, the results presented in Table 1 showed that the number of days required for the callus induction was 21.83 days in Gtyp2, followed by Casper F1 (27.17 days). However, the maximum number of days for the callus induction was recorded in the Gtyp5 (31.49 days). The differential response of different genotypes for days to callus induction was also reported by Alam et al. (2009)24 Khan et al. (2009)25 Sayem et al. (2010)26 and Singh et al. (2022)15.
Our results revealed that there are significant differences between the medium composition used to grow the anthers, the callus induction frequency was the highest in MS1 and MS2 mediums (56.8 and 54.94%, respectively), as compared to the others. While the treatment MS0 was failed (Table 2). Among the five treatment compositions of the medium containing different concentrations of the growth regulators, MS3: MS + 0.3 mg/L 2,4-D + 1 mg/L BAP required minimum time (24.9 days) for callus induction with significant differences compared to the other medium, while the MS4: MS + 0.7 mg/L 2,4-D + 1 mg/L BAP took maximum time (32.33 days) for callus induction. On the other hand, the higher response of mediums to the percent of callus induction was perceived in MS1 (56.8%), followed by MS2 (54. 94%). While the minimum percent was recorded in MS4 (31.57%). From these results, it was observed that the performance of the medium with these concentrations of 2,4-D were significantly different suggesting that both days to callus induction and percent of callus induction were greatly influenced by the concentrations of 2,4-D. Also, the results of Narasimhulu and Chopra (1987)27 Ockenden and Mc Cleriaghan (1993)28 Sayem et al. (2010)26 and Reetisana et al. (2018)29 were similar to our consequences. Also, a study by Islam et al. (2022)30 showed that Brassica genotype (CM800) anthers cultured on medium supplemented with 4 mg/L 2, 4-D and 1 mg/L BAP exhibited the significantly highest response for callus induction in minimum days (18 days) and the maximum percentage of callus (56.2%).
The type × medium interaction was found to be significant for days to callus induction (Table 3). So, the required days for callus induction in Gtyp2 were found in MS1 (22.33 days). On the other side, there was also a significant difference in the callus induction frequency, the callus induction frequency was the highest significantly in Gtyp1 with MS1 (88.9%), then with MS2 medium (75.9%) and Casper F1 with MS1 (75.9%), while the lowest callus induction frequency was significantly lower for Gtyp4 with MS3 (16.67%).
These results confirm that the callus induction frequency is mainly determined by the presence of auxin 2,4-D in the induction medium. The results also indicate the role of genetic variation in the callus induction frequency in the studied types of cauliflower. However, the study of Rashid et al. (2009)31 indicated that the increase in callus induction in the presence of auxin 2,4-D results from its role in encouraging mitotic division, as auxin increases the rate of RNA synthesis. Also, Centeno et al. (1996)32 mentioned the importance of growth regulators in the growth and differentiation of callus tissues through the direct role of auxins in expanding the plant cell and increasing its permeability and flexibility, and the role of cytokinins in cell division, and the manufacture of some important proteins and RNA formation. As for the variation in the response of varieties to callus induction, many studies have shown that callus induction varies among species and even among varieties of the same species33. Similar results were reported by Reetisana et al. (2018)29 in which a significant effect of genotype and media interaction was observed for days taken to initiate callus.
Shoot regeneration
Our results in the present study indicated significant variations among the types and treatments for days to shoot regeneration and for the shoot regeneration frequency (Table 4). Among the types, Casper F1 started shoot regeneration early (31.25 days) as compared to other types, while Gtyp1 took more days for shoot regeneration (52.08 days). Also, the highest percentage of shoot regeneration was noticed in Casper F1 (39.31%) as compared to the other types. However, the minimum percentage of shoot regeneration was detected in Gtyp2 (28.43%). As well as the results of this study revealed significant differences in the number of plantlets among the types. Otherwise, the plantlets number of Casper F1was 5.17, while it was the minimum in Gtyp5 (2.17 plantlets). The outcomes presented in Table (5) indicated that the medium MS1 revealed a lowest time for shoot initiation (38.72 days). The period for shoot initiation by using MS4 was 46.28 days and 46 days in MS3. The shoot regeneration frequency was found to be the highest significantly in MS1 (38.8%), which was significantly superior to the other media. MS4 medium recorded the lowest shoot regeneration frequency (23.29%). MS1 recorded the highest value for the number of plantlets (4.61 plantlets) with significant differences with the other media, while MS4 medium recorded the lowest value for the number of plantlets (2.83 plantlets). Similar to our findings, the results of Khan et al. (2009)26 and Sayem et al. (2010)27 revealed that the different concentrations of plant growth regulators in the medium culture led to significant variations in shoot regeneration frequency.
On the other hand, the type × medium interaction was found to be significant for days to shoot initiation. Casper F1× MS1 took the lowest value for days taken to initiate shoot regeneration (28.33 days), whereas Gtyp1 × MS2 took the highest value for days taken to initiate shoot regeneration (56.33 days). (Table 6). Also, the highest value of number of plantlets was obtained in Casper F1 (8.33 plantlets) when grown in the medium MS1, while the lowest value of number of plantlets was noticed in local Gtyp4 (1.33 plantlets) when grown in MS4. These results indicate that the important factor determining the regeneration of the shoot from callus is the presence of cytokine in the shoot regeneration medium because it plays a role in forming buds from undifferentiated tissues (callus), and cell division, Also, it activates the synthesis of proteins necessary for division, and the balance between auxin and cytokine in the medium culture was more effective for shoots regeneration34. While Ravanfar et al.35 concluded that BAP was the most effective plant growth regulator used for shoot regeneration and multiplication for broccoli. Also, Zhao et al. (2010)36 indicated that the number of plantlets is affected by different hormonal compatibility, growth regulator concentrations, and genetic material.
Ploidy level of regenerated plantlets
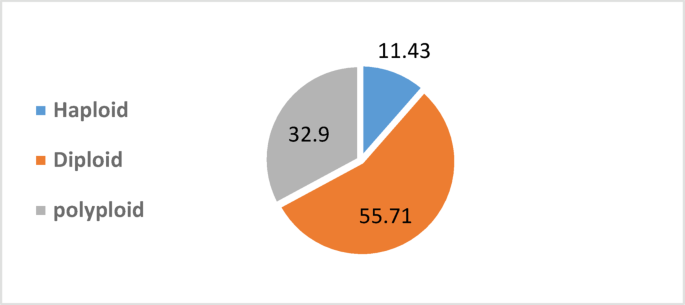
In the current study, the slides were prepared from the root tips of regenerated plantlets to count the mitotic chromosome number. Since the diploid chromosome number of cauliflower is 2n = 18, the haploid plantlet root tips contained 9 chromosomes, and the polyploid plantlet root tips contained more than 18 chromosomes. The results presented in Table 7 indicate that there are many variations in the regeneration plantlets number and the percentages of ploidy levels among the cauliflower types. The total number of regeneration plantlets from anthers culture in all types was 70 plantlets. The classification of these plants revealed that 11.43% were haploid, 55.71% diploid, and 32.9% polyploidy plants (Fig. 1). The diploid and polyploid plantlets were spontaneously produced during anthers culture. Several studies show that in vitro plant tissue culture can lead to polyploidy as a result of exposing plant cells to abnormal conditions that affect cell division., D’Amato (1985)37 indicated that polyploid plants can arise during anther culture due to mitotic spindle dysfunction, where environmental stressors or hormonal imbalances disrupt chromosome segregation, resulting in endoreduplication. Also, Phillips et al. (1994)38. reported that nuclear fusion or restitution nuclei, where incomplete cell division or nuclear fusion leads to polyploid cells. Barow and Meister (2003)39 revealed that the endoreduplication, caused by asynchronous cell cycles or failed cytokinesis in meristematic cells, is often induced by in vitro stress. As well as high concentrations or imbalanced ratios of auxins and cytokinins can disturb normal cell cycle progression and induce chromosomal duplication16. The maximum number of regenerated plantlets was observed in Casper F1 (20 plantlets), while the minimum number was shown in Gtyp4 (7 plantlets). As well, haploid plantlets were ranged from 0% in Gtyp1 to 16.67% in Gtyp3, diploid plants were ranged from 45% in Casper F1 to 75% in Gtyp1, and polyploidy plants were ranged from 25% in Gtyp1 to 40% in Casper F1. However, the results of Keller and Armstrong (1983)40 Prabhudesai and Bhaskaran (1993)41 and Wang et al. (1999)42 revealed that there are many haploid and ploidy plants among different genotypes of cauliflower produced by using the anther culture technique. Also, Wang et al. (1999)42 indicated that ploidy levels were genotype and species specific. Sunderland et al. (1974)43 reported that another factor affecting the ploidy level is the stage of pollen grain development when anthers are cultured. Singh et al. (2022)15 also reported that genotypic differences were recorded in obtaining the percent ploidy among different genotypes, and it is still unknown why some genotypes/species produce more haploids than others. Our results are consistent with the results of Stipic and Campion (1997)44 on anther cultures of three cauliflower genotypes, the chromosome number analysis of 79 plants showed that 15% were haploid, 78% were diploid, and 7% were polyploid.

Ploidy level of regenerated plants.
Conclusion
Traditionally, the development of homozygous inbred lines takes years, while double haploid technology is a single- step route. However, it is dependent on many factors like genotype and media compositions. Therefore, it has to be standardized for each genotype to be used in the breeding program. It was found that Gtyp1 gave the highest response to callus induction when cultured on MS + 0.3 mg/L 2,4-D + 1 mg/L BAP medium and the maximum plantlets regeneration frequency was observed in Casper F1 when grown on MS + 0.5 mg/L NAA + 2 mg/L 2ip medium, indicating that cytokinins played an important role in shoot regeneration. Also, our study revealed that ploidy analysis confirmed the presence of 11.43% haploids 55.71% diploids and 32.9% polyploids. The protocol thus developed in this way will help researchers to produce homogeneous lines in a relatively shorter time.

Different stages of haploid development in cauliflower “Casper F1”; a– Donor plant, b– Unopened flower buds, c– Microspore examine by acetocarmine, d– Anthers culture in MS1, e – f: Callus induction, g– Embryogenic callus formation, h– Shoot regeneration from embryogenic callus, i– Plantlet development, j– Hardening of plantlets, k-i: Haploid plant.
Materials and methods
Donor plant material and growing conditions
This research was done during the period 2022–2023 at al-Ghouta research station/General Commission for Scientific Agricultural Research, located in Damascus, Syria (33º 24.64ʹ N, 36º 30.87ʹ E and 616 m above the mean sea level), and the in vitro anthers culture was carried out at the laboratory of plant biotechnology/National Commission for Biotechnology in Damascus. In this study, five local types (Gtyp1, Gtyp2, Gtyp3, Gtyp4, and Gtyp5) in addition to the control Casper F1, were cultivated in the field, with dimensions (80 and 70 cm) between plants and lines respectively. The results of the soil analysis showed that the soil is sandy clay, (pH = 8.11), its organic matter content is good, the percentage of calcium carbonate is high, and its phosphorus and potash content is good (Table 8).
Anther culture technique
The unopened flower buds (4–4.5 mm in size) of all types were collected before 10 a.m. Then, the collected flower buds were wrapped in aluminum foil and kept in the refrigerator at 4 °C for 24 h. After the cold treatment, they were surface sterilized under aseptic conditions in a laminar airflow chamber. They were rinsed in 70% ethanol for 30 s, then immersed in a 4% sodium hypochlorite solution for 15 min, followed by three washings with sterile distilled water45 then placed on sterile filter paper to extract the anthers. The anthers were cultured horizontally on the surface of the solid medium in a petri dish containing 20 ml of media, 18 anthers were cultured in each dish. In this study, we repeated the culture 7 times for every treatment of each type. After that, the samples were incubated under conditions (25 ± 2 °C and 70% humidity) for callus induction46. Callus formation was observed after 3–4 weeks of anthers culture. Then, a callus mass with a diameter of more than 1 cm was transferred to test tubes which contained 20 ml shoot initiation media, and the culture was repeated 9 times for every treatment of each type, and incubated under conditions (25 ± 1 °C and16 hours of photoperiod). Then when the sub- cultures were proliferated and differentiated into shoots and plantlets (2–3 cm in length) they aseptically and separated from each other to culture again in test tubes have mediums of root induction to induce roots, also, the culture was repeated 9 times for every treatment of each type, The plantlets (one plantlet in each test tube) are placed strictly on the surface of the solid medium without going deep into it. The optimal levels of growth regulators were determined by studying callus induction frequency, days to callus induction47shoot regeneration frequency, days took to initiate shoot regeneration, and a number of plantlets. Figures (2) shows the different stages of haploid development in cauliflower “Casper F1” through anther culture.
Media used for anther culture
Callus induction media
MS medium of Murashing and Skoog (1962)16 with pH 5.8, has been supplemented with 7 g/L agar and 30 g/L sucrose, in addition to the plant growth regulators (6-benzylaminopurine BAP and 2,4-dichlorophenoxyacetic acid 2,4-D) which were presented in Table 9. Else, the medium culture was sterilized by using the autoclave at 120 °C for 22 min.
Shoot regeneration media
In this stage, the MS medium was prepared as like as the medium of callus induction.
But, in these medium, plant growth regulators (Table 9) consist of naphthalene acetic acid (NAA), 6-benzyl adenine (BA), 2-isopentenyl adenine (2iP), and 6-benzyl amino purine (BAP).
Ploidy analysis
The direct method described by Darlington and Lacour (1976)48 was adopted to investigate the ploidy level of the plant regenerated from anther culture by using the standard acetocarmine root tip squashing technique. The root tips with a length of 0.5 cm were fixed in the rate 1:3 of glacial and ethanol, respectively for 24 h. Then the fixed root tips were washed with running water and preserved in 70% ethanol. The root tips were taken in a clean slide and the tissue was allowed to stain for 20 min in 2% acetocarmine stain. Then, the stained tissue was softened with 0.1 N HCl for 50 s, also, warmed over a spirit lamp and squashed with a needle and a cover slip was placed over it. Then a filter paper was placed over the coverslip and calm pressure was applied for further flattening and spreading of the chromosomes and observed under a microscope (100X).
Observations recorded
-
Number of days to callus induction.
The number of days to callus induction was recorded from the starting anthers culture to the beginning of callus induction.
-
Callus induction frequency.
This index was calculated according to the following formula used by Singh et al. (2022)15:
Callus induction frequency % = (::frac{mathbf{N}mathbf{u}mathbf{m}mathbf{b}mathbf{e}mathbf{r}:mathbf{o}mathbf{f}:mathbf{a}mathbf{n}mathbf{t}mathbf{h}mathbf{e}mathbf{r}mathbf{s}:mathbf{s}mathbf{h}mathbf{o}mathbf{w}mathbf{i}mathbf{n}mathbf{g}:mathbf{c}mathbf{a}mathbf{l}mathbf{l}mathbf{u}mathbf{s}:mathbf{i}mathbf{n}mathbf{d}mathbf{u}mathbf{c}mathbf{t}mathbf{i}mathbf{o}mathbf{n}}{mathbf{T}mathbf{o}mathbf{t}mathbf{a}mathbf{l}:mathbf{n}mathbf{u}mathbf{m}mathbf{b}mathbf{e}mathbf{r}mathbf{s}:mathbf{o}mathbf{f}:mathbf{a}mathbf{n}mathbf{t}mathbf{h}mathbf{e}mathbf{r}mathbf{s}:mathbf{c}mathbf{u}mathbf{l}mathbf{t}mathbf{u}mathbf{r}mathbf{e}mathbf{d}}times::100)
-
Shoot Regeneration Frequency
Also, this index was calculated according to the following formula used by Singh et al. (2022)15:
Shoot Regeneration Frequency % = (:frac{mathbf{N}mathbf{u}mathbf{m}mathbf{b}mathbf{e}mathbf{r}:mathbf{o}mathbf{f}:mathbf{c}mathbf{a}mathbf{l}mathbf{l}mathbf{i}:mathbf{s}mathbf{h}mathbf{o}mathbf{w}mathbf{i}mathbf{n}mathbf{g}:mathbf{r}mathbf{e}mathbf{g}mathbf{e}mathbf{n}mathbf{e}mathbf{r}mathbf{a}mathbf{t}mathbf{i}mathbf{o}mathbf{n}:}{mathbf{N}mathbf{u}mathbf{m}mathbf{b}mathbf{e}mathbf{r}:mathbf{o}mathbf{f}:mathbf{c}mathbf{a}mathbf{l}mathbf{l}mathbf{i}:mathbf{c}mathbf{u}mathbf{l}mathbf{t}mathbf{u}mathbf{r}mathbf{e}mathbf{d}}times::100:)
-
Number of regenerated plantlets.
-
Ploidy level of regenerated plantlets.
Statistical analysis
The data were tested and subjected to analysis of variance (ANOVA) by using GenState software version 12, to obtain the effect of various nutrient media, types and their interactions based on a factorial randomized block design. A comparison of means was performed by using the LSD test (p < 0.01).
Data availability
The datasets used and analysed during the current study available from the corresponding author on reasonable request.(Sumaya Jabal, PhD student. email; somijabsll@gmail.com).
Abbreviations
- BA:
-
6-benzyladenine
- BAP:
-
6-benzylaminopurine
- 2,4-D:
-
2,4-dichlorophenoxyacetic acid
- 2iP:
-
N6-2-isopentenyl adenine
- NAA:
-
α-naphthalene acetic acid
- MS:
-
Murashige and Skoog medium
References
-
Heywood, V. H., Brummitt, R. K., Culham, A. & Seberg, O. Flowering plant families of theworld. Ontario: Firefly Books, 88, (2007).
-
Thamburaj, S. & Singh, N. Textbook of vegetables, tubercrops and spices. Indian Council Agricul. Res. (2001).
-
Maggioni, L., von Bothmer, R., Poulsen, G. & Lipman, E. Domestication, diversity and use of Brassica oleracea L., based on ancient Greek and Latin texts. Genet. Resour. Crop Evol. 65 (1), 137–159 (2018).
-
Chittora, A. & Singh, D. K. Genetic variability studies in early cauliflower (Brassica Oleracea var. botrytis L). Electro J. Plant. Breed. 6 (3), 842–847 (2015).
-
Holland, B., McCance, R. A., Widdowson, E. M., Unwin, I. D. & Buss, D. H. Vegetables, herbs and spices: Fifth supplement to McCance and Widdowson’s The Composition of Foods. Royal Soci Chemis 5, (1991).
-
Mandrich, L. & Caputo, E. Brassicaceae-derived anticancer agents: towards a green approach to beat cancer. Nutrients 12 (3), 868 (2020).
-
FAOSTAT, Food and agriculture organization of the United Nations. (2020). http://faostat.fao.org/. Rome, Italy.
-
Dogru, S. M., Cilingir, A., Kurtar, E. S. & Balkaya, A. Usage of haploid and doubled-haploid techniques in the cultivar breeding of cabbage group vegetables. Turk. Seed Union Maga. 5 (20), 30–33 (2016).
-
Tadesse, W., Inagaki, M., Tawkaz, S., Baum, M. & Van Ginkel, M. Recent advances and application of doubled haploids in wheat breeding. Afri J. Biotech. 11 (89), 15484–15492 (2012).
-
Kameya, T. & Hinata, K. Induction of haploid plants from pollen grains of Brassica. Japan J. Breed. 20 (2), 82–87 (1970).
-
Ali, J. et al. Improved anther culture media for enhanced callus formation and plant regeneration in rice (Oryza sativa L). Plants 10 (5), 839 (2021).
-
Gosal, S. S., Pathak, D., Wani, S. H., Vij, S. & Pathak, M. Accelerated breeding of plants: methods and applications. Accelerated Plant Breeding, Volume 1: Cereal Crops, 1–29. (2020).
-
Ockendon, D. J. The ploidy of plants obtained from anther culture of cauliflowers (Brassica Oleracea var. botrytis). Ann. Appl. Boil. 113 (2), 319–325 (1988).
-
Chaudhary, S., Katoch, V., Kapila, R. K., Sharma, A. & Sharma, S. Response of sucrose, PGRs and pretreatments on another culture efficiency in broccoli (Brassica Oleracea L. Var. Italica Plenck). J. Pharmacog Phytochem. 8, 566–573 (2019).
-
Singh, R., Devi, R. & Sarao, N. K. Generation of doubled haploids in cauliflower. Heliyon, 8 (12), (2022).
-
Murashige, T. & Skoog, F. A. Revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Planta. 15, 473–495 (1962).
-
Gamborg, O. L., Miller, R. & Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exper. Cell Resear. 50 (1), 151–158 (1968).
-
Nitsch, C. & Nitsch, J. P. The induction of flowering in vitro in stem segments of Plumbago indica L. I. The production of vegetative buds. Planta. 72 (4), 355–370 (1967).
-
Chu, C. C. The N6 Medium and its Applications To Anther Culture of Cereal Crops pp. 43–50 (Proc Symp Plant Tissue Cult Peking, Science, 1981).
-
Fuller, M., Turton, P. & Grout, S. Anther culture of winter-heading cauliflower. Int. Symp. Vitro Cult. Hortic. Breed. 280, 329–332 (1989).
-
Yang, Q., Chauvin, J. E. & Hervé, Y. A study of factors affecting anther culture of cauliflower (Brassica Oleracea var. botrytis). Plant. Cell. Tissue Organ. Cult. 28, 289–296 (1992).
-
Cilingir, A., Dogru, S. M., Kurtar, E. S. & Balkaya, A. Anther culture in red cabbage (Brassica oleraceae L. Var. capitata subvar. rubra): embryogenesis and plantlet initiation. Ekin J. 3 (2), 82–87 (2017).
-
Sood, S. & Kumari, V. Efficient callus induction through anther culture in cultivars of Brassica campestris var. Brown Sarson. Int. J. Curr. Microbiol. App. Sci. 8(5), 1003–1012 (2019).
-
Alam, M. A., Haque, M. A., Hossain, M. R., Sarker, S. C. & Afroz, R. Haploid plantlet regeneration through anther culture in oilseed Brassica species. Bangl J. Agricultural Res. 34 (4), 693–703 (2009).
-
Khan, M. M. A., Hassan, L., Ahmad, S. D., Shah, A. H. & Batool, F. In vitro regeneration potentiality of oil seed Brassica genotypes with differential BAP concentration. Pak J. Bot. 41 (3), 1233–1239 (2009).
-
Sayem, M. A., Maniruzzaman, M., Siddique, S. S. & Al-Amin, M. In vitro shoot regeneration through anther culture of Brassica spp. Bangla J. Agricul. Res. 35 (2), 331–341 (2010).
-
Narasimhulu, S. B. & Chopra, V. L. Tissue culture studies in Brassica carinat A. Braun. Cruciferae Newsl. 12 (1), 70 (1987).
-
Ockendon, D. J. & McClenaghan, R. Effect of silver nitrate and 2, 4-D on anther culture of Brussels sprouts (Brassica Oleracea var. gemmifera). Plant Cell, Tissue Organ Cult. 32, 41–46 (1993).
-
Reetisana, N., Devi, T. R., Devi, H. N., Laishram, J. M. & Pyngrope, A. H. In vitro haploid plantlet regeneration through anther culture in locally adapted cultivar of Indian mustard (Brassica juncea L. Czern and Coss). Int. J. Curr. Microbiol. App Sci. 7 (4), 8–19 (2018).
-
Islam, M. M., Bhuiyan, M. S. H. & Nadim, M. K. A. In vitro callus regeneration of brassica ssp through anther culture. Bangladesh J. Nuclear Agric. 36 (1), 115–119 (2022).
-
Rashid, U., Ali, S., Ali, G. M., Ayub, N. & Masood, M. S. Establishment of an efficient callus induction and plant regeneration system in Pakistani wheat (Triticum aestivum) cultivars. Electro J. Biotech. 12 (3), 4–5 (2009).
-
Centeno, M. L., Rodríguez, A., Feito, I. & Fernández, B. Relationship between endogenous auxin and cytokinin levels and morphogenic responses in Actinidia deliciosa tissue cultures. Plant Cell Rep. 16, 58–62 (1996).
-
Shyam, C. et al. Plant regeneration in Indian mustard [Brassica juncea (Linn.) Czern & Coss]: Experimental investigation. book: current Topics in Agricultural Sciences. Publisher. BP Global Inter. 3 (9), 120–135 (2021).
-
Morel, G. C., Matin & Muller, C. Laguuerison des pommes De terre atteintes irus. de maladies a virus. Ann. Physiol. Veg. 10 (2), 113–139 (1968).
-
Ravanfar, S. A., Aziz, M. A., Kadir, M. A., Rashid, A. A. & Sirchi, M. H. T. Plant regeneration of Brassica oleracea subsp. Italica (Broccoli) CV green Marvel as affected by plant growth regulators. African J. Biotechnology, 8 (11) (2009).
-
Zhao, L., Liu, S. & Song, S. Optimization of callus induction and plant regeneration from germinating seeds of sweet sorghum (Sorghum bicolor Moench). Afri J. Biotechn. 9 (16), 2367–2374 (2010).
-
D’Amato, F. & Bayliss, M. W. Cytogenetics of plant cell and tissue cultures and their regenerates. CRC. Crit. Rev. Plant Sci. 3 (1), 73–112 (1985).
-
Phillips, R. L., Kaeppler, S. M. & Olhoft, P. Genetic instability of plant tissue cultures: breakdown of normal controls. Proceedings of the National Academy of Sciences, 91 (12), 5222–5226 (1994).
-
Barow, M. & Meister, A. Endopolyploidy in seed plants is differently correlated to systematics, organ, life strategy and genome size. Plant. Cell. Environ. 26 (4), 571–584 (2003).
-
Keller, W. A. & Armstrong, K. C. Production of haploids via anther culture in Brassica oleracea var. italica. Euphytica 32, 151–159 (1983).
-
Prabhudesai, V. & Bhaskaran, S. A continuous culture system of direct somatic embryogenesis in microspore-derived embryos of Brassica juncea. Plant. Cell. Repor. 12 (5), 289–292 (1993).
-
Wang, M., Farnham, M. W. & Nannes, J. S. P. Ploidy of broccoli regenerated from microspore culture versus anther culture. Plant. Breedi. 118 (3), 249–252 (1999).
-
Sunderland, N., Collin, G. B. & Dunwell, J. M. The role of nuclear fusion in pollen embryogenesis of Datura innoxia. Planta 117, 227–241 (1974).
-
Stipic, M. & Campion, B. An improved protocol for androgenesis in cauliflowers (Brassica Oleracea var. botrytis). Plant. Breed. 116 (2), 153–157 (1997).
-
Germana, M. A. Gametic embryogenesis and haploid technology as valuable support to plant breeding. Plant. Cell. Repor. 30, 839–857 (2011).
-
Dias, J. S. & Martins, M. G. Effect of silver nitrate on anther culture embryo production of different Brassica oleracea morphotypes. Scientia Horti. 82 (3–4), 299–307 (1999).
-
Nasircilar, A. G., Turgut, K. & Fiskin, K. Callus induction and plant regeneration from mature embryos of different wheat genotypes. Pakistan J. Bota. 38 (3), 637 (2006).
-
Darlington, C. D. & Cour, L. L. The handling of chromosomes. (6thedn.) Allen and Unwin, London (1976).
Acknowledgements
This work was supported by Damascus University, faculty of agriculture and department of horticulture science and the general commission for scientific agricultural research, Damascus Syria. As well, a lot of thanks to the Dr. Fahed Albiski, Dr. Mouhamad Alhoshan and Mr. Youssef Al Saadi for their help during the period of study and preparing my articl.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Jabal, S., Murshed, R. & Alabdalla, O. In vitro haploid plantlet regeneration through anther culture in some local types of Syrian cauliflower (Brassica Oleracea var. botrytis L.). Sci Rep 15, 27581 (2025). https://doi.org/10.1038/s41598-025-12802-0
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12802-0