
Introduction
Mechanobiology is a research field that elucidates the effects and mechanisms of mechanical stress on cells, tissues, and organism. Insights gained from basic research in mechanobiology are expected to contribute to creating groundbreaking therapeutic techniques in a wide range of medical fields, including regenerative medicine1,2. In particular, in 2006, Discher et al. reported that when stem cells were cultured two-dimensionally on hydrogels of different stiffness, the stem cells sensed the difference in stiffness, controlling the direction of differentiation depending on the stiffness3. Regarding this phenomenon, cells receive external mechanical stress using various mechanosensors. The mechanical stress input to cells from the mechanosensors is then converted into biochemical reactions within the cells, which ultimately induce nuclear translocation of transcription factors and transcription coupling factors, such as YAP, through complex cascade reactions occurring in the cytoplasm. The mechanism by which the differentiation response is changed via gene expression level adjustments, known as mechanotransduction, has been uncovered in recent years4,5,6,7,8,9,10,11,12,13. These studies have shown that appropriate mechanical stress are powerful and robust differentiation inducers essential for development of regenerative medicine technologies. These studies have also demonstrated that hydrogels can exhibit unique mechanical properties similar to those of various soft tissues based on the type of monomer and crosslinking method selection, making them a suitable three-dimensional scaffolding material for mechanobiology14,15,16. Meanwhile, research focusing in mechanosensors that lead to the occurrence of mechanotransduction has been actively conducted, identifying various types of mechanosensors and their functions. Among them, integrins are the most common and vital mechanosensors17,18. Integrins are membrane proteins present on the cell surface, and as receptors for the extracellular matrix, they play an biochemically instrumental role in adhesive responses between cells and the extracellular matrix. Integrins also play a role in physically connecting between extracellular matrix and the intercellular framework, transmitting extracellular mechanical stress into the cell. Usually, integrins have been reported to alter cellular responses by receiving signal transducers, stiffness, shear stress, tensile and compressive stress, and gravity3,19,20,21,22. In particular, shear stress and tensile stress are applied to integrins in muscle tissue in vivo21, and gravity and compressive forces are applied to integrins in bone and cartilage tissue to promote cell activation22. They thereby control cell shape, motility, differentiation, and other properties through mechanotransduction23.
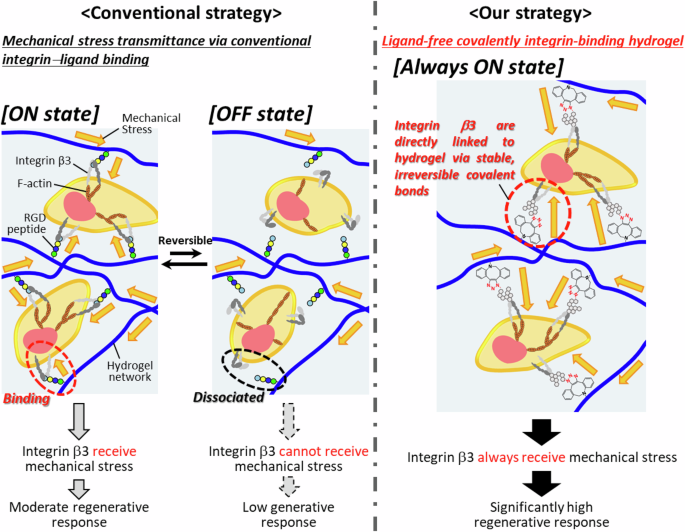
In tissue engineering field, a method that administers stem cell encapsulated inside the scaffolding hydrogels into damaged tissues is understood to be promising to promote tissue regeneration. By combing this conventional Method to the insights from mechanobiology, it is reasonably conceived that if the transplanted cells inside the hydrogel can be made to effectively receive the shear stress, tensile stress, compressive stress, and gravity generated in the living body, it may be possible to effectively elicit a regenerative response of the transplanted cells24. Although successful cases are still few, in recent years, several research groups have initiated the challenge of developing technology that utilizes mechanical stress in the living body to promote the regenerative responses of transplanted cells within scaffolding hydrogel. However, hydrogels essentially have the ability to buffer mechanical stress by stress relaxation, making it difficult to efficiently transmit mechanical stress generated outside the hydrogel in vivo to transplanted cells present within the hydrogel. To address this difficulty, in these studies, researchers have, without exception, focused on integrins as the essential mechanosensors and adopted strategies to enable the integrins of transplanted cells within the hydrogel to sense the mechanical stress of the body effectively. Following this strategy, researchers have constructed hydrogels modified with ligands for integrins. For example, in 2020, Garcia et al. prepared PEG hydrogels modified with the GFOGER peptide, a specific ligand for integrin α2β1, and with the GAOGER peptide, which does not bind to integrin α2β1. They examined the bone differentiation of mesenchymal stem cells induced by the reception of mechanical stress via integrin–ligand binding25. Consequently, the efficiency of bone differentiation was low in the GAOGER peptide-modified hydrogel. Meanwhile, the GFOGER peptide-modified hydrogel promoted bone differentiation, resulting in the regeneration of bone tissue with approximately twice the bone mass. Additionally, in 2020, Wang et al. prepared collagen hydrogels modified with LXW7 peptide, a specific ligand for integrin αvβ3, and collagen hydrogels not modified with LXW7 peptide. They examined the formation of vascular networks by vascular endothelial cells induced by the reception of mechanical stress via integrin–ligand binding. Consequently, no vascular network structure was formed in LXW7-unmodified collagen hydrogels, whereas a dense vascular network structure was formed in LXW7-modified collagen hydrogels26. These pioneering research suggest that the cells within the hydrogel indeed seems to be received mechanical stress generated in the body through the integrin−ligand binding. However, an important concern is that the regeneration efficiency was not high. In other words, this implies that this conventional method, which utilizes integrin–ligand binding, a mechanism that cells naturally possess, is ineffective in fully harnessing the mechanical stress of the living body. The cause of this is attributed to the fact that the integrin−ligand binding is broken when an external mechanical stress of about greater than 50 pN is applied27,28,29. And cells in biological tissues are usually subjected to external mechanical stress. For example, it is known that muscle satellite cells within skeletal muscle are subjected to tensile and shear stress of approximately 50 pN for each contraction movement of the muscle tissue23. In other words, the integrin–ligand binding is mediated by weak interactions such as hydrogen bonding and electrostatic interaction, causing the binding to break and dissociate (OFF state) from the application of intense mechanical stress (Fig. 1, left)30. Additionally, it is understood that when cells migrate or divide, the binding between the integrins of transplanted cells and the ligands modified on the hydrogel is severed. They prompt the transplanted cells to enter an OFF state so that the mechanical stress of the living body is not received31. Moreover, since integrins are calcium ion-dependent receptors, a concentration of calcium ions above a certain threshold is required to receive mechanical stress effectively. In other words, the fundamental problem with the conventional method, which utilizes integrin–ligand binding within hydrogel, is the low efficiency of receiving mechanical stress on the grounds of hydrogelʹs ability to buffer mechanical stress as well as the weak binding between the integrin of the transplanted cells and the ligand modified on the hydrogel, its reversibility, plasticity, and its dependence on the cell cycle and the extracellular environment.

This system focuses on integrin as cellular elements that receive mechanical stress from the alginates, and actin filaments connecting the integrin to the nuclear lamina allow mechanical stress to be transmitted from the integrin through the cytoplasm to the nucleus.
As a rational idea to solve this problem, we have devised a concept that does not use ligands for integrins (Fig. 1, right). Specifically, this is a method to directly link the integrin of transplanted cells to the hydrogel via a covalent bond. In our method, fundamentally different from conventional ones, azide groups were introduced into the integrinʹs sugar chain by glycometabolic engineering32,33. Glycometabolic engineering is a technology to artificially change the structure of sugar chains on the cell surface. By applying glycometabolic engineering, it is possible to target specific sugar metabolism pathways and introduce reactive functional groups into sugar chains on the cell surface34. In general, the glycans with functional groups introduced by the glycometabolic pathway can form bonds through selective chemical reactions while maintaining biocompatibility35,36,37. As an alternative to ligands, the hydrogel-forming polymers were modified with cyclooctyne groups (DBCO), which selectively form covalent bonds with the azide groups38,39. A bioorthogonal click reaction between the azide and DBCO groups forms a direct covalent bond between the cellular integrin and the hydrogel. This technique is essentially different from conventional methods in that the integrins of the transplanted cells are directly bound to the hydrogel via stable and irreversible covalent bonds, so that the integrin–hydrogel bond is always in an ON state. This linkage structure can highly efficiently receive mechanical stress generated in the body regardless of the intensity of the stress, the cell cycle, or the extracellular environment. Therefore, the technique is expected to yield high cellular responses which promote tissue regeneration. In this article, our developed method is called “ligand-free covalently integrin-linking hydrogel”.
In this article, we chose skeletal muscle as a representative tissue model in which intense mechanical stress occurs. We evaluated the physicochemical properties of the ligand-free covalently integrin-binding hydrogel, its mechanical stress transmitting properties, cell responses, and skeletal muscle tissue regenerative capability in an effort to verify the usefulness of our concept. As the results, we demonstrate that the ligand-free approach that directly links a hydrogel with the integrin of transplanted cells via covalent bond can enable transplanted cells to efficiently receive mechanical stress in the body and elicit a regenerative response. Further, we found that when cells were exposed to mechanical stress in ligand-free covalently integrin-linking hydrogel, the mechanical stress was rapidly and reliably transmitted to the nucleus through a mechanism that was essentially different from previously reported mechanotransduction. We discovered that this resulted in a regenerative response in the transplanted cells with high efficiency.
Results
Design, preparation, and characterization of the ligand-free covalently integrin-linking hydrogels
For designing our ligand-free covalently integrin-linking hydrogels, we targeted integrin, a membrane protein rich in sialic acid40, as the key element for covalent linking to a scaffolding hydrogel. Azide groups can be introduced into sialic acids via metabolic glycoengineering, enabling these sites on integrin to serve as artificial linking points. As an alternative to ligands for the azide-modified integrin, the hydrogel-forming polymers were modified with DBCO groups, which selectively form covalent bonds with the azide groups. A bioorthogonal click reaction between the azide and DBCO groups forms a direct covalent bond between the cellular integrin and the hydrogel (Fig. 2a). In this study, we chose skeletal muscle as a representative tissue model in which intense mechanical stress occurs, and constructed ligand-free covalently integrin-linking hydrogels using muscle satellite cells (HskMSCs). We hypothesized that the efficiency of mechanical stress transmission depends on the number of linking points between the integrin of HskMSCs and the hydrogel, and thus attempted to control the number of azide groups introduced into integrin of HskMSCs. To analyze the azide groups introduced into HskMSCs, we used clickable rhodamine, which selectively binds to azide groups. We controlled the number of azide groups introduced by varying the incubation time in a medium containing Ac4ManNAz (Supplementary Fig. 1). Azide-modified HskMSCs reacted with clickable rhodamine were cultured and grown in medium that did not contain Ac4ManNAz for several days, after which the fluorescence intensity of rhodamine was quantified. As a result, the fluorescence intensity decreased over time, suggesting that in this system, the amount of azide groups is halved due to cell proliferation (division), and therefore as cell proliferation progresses, the decrease in the number of azide groups due to proliferation exceeds the increase in the amount of azide groups introduced, and after 48 h (Supplementary Fig. 1), the apparent amount of azide groups per cell (rhodamine fluorescence intensity) appears to have decreased.

a Schematic illustration of the construction of a human skeletal muscle satellite cells (HskMSCs) gel. b CLSM image of DiO-labeled N3(+)HskMSCs treated with Cy3-labeled Alg- DBCO10 (n = 2, independent samples). Green: DiO and red: Cy3. Scale bar indicates 50 μm. c Gelation times of the gels prepared by Alg-DBCO10 and N3(+)HskMSCs with different numbers of introduced azide groups per cell (n = 3). Data are presented as mean ± SD. d Saturated G’ values of the gels prepared by Alg-DBCO10 and N3(+)HskMSCs with different numbers of introduced azide groups per cell (n = 3). Data are presented as mean ± SD. e CLSM images of the gels prepared by Alg-DBCO10 and N3(+)HskMSCs with different numbers of introduced azide groups per cell after the treatments with live/dead assay. Green: calcein-AM and red: PI. Scale bars indicate 100 μm. f Viability of N3(+)HskMSCs in the gels, quantified from the corresponding CLSM images of the live/dead assay (n = 3). Data are presented as mean ± SD. g Immunoblot analysis of control N3(+)HskMSCs (left) and shRNA ITGB3(+)N3(+)HskMSCs treated with antibodies against Integrin-β3 and α-tubulin as a loading control (n = 2, independent samples).
When clickable rhodamine was administered to azide-modified HskMSCs (N3(+)HskMSCs) at different concentrations, the amount of unreacted clickable rhodamine remaining in the supernatant was quantified (Supplementary Figs. 2, 3, and 4), allowing the calculation of the number of azide groups introduced. The estimated number of azide groups introduced per cell was 2.1 × 1010 after 12 h of treatment, 2.6 × 1010 after 24 h of treatment, and 3.1 × 1010 after 48 h of treatment (Supplementary Table 1).
Alginate, which is a biocompatible material, was used as the mechanical stress conductive rigid polymer to form ligand-free covalently integrin-linking hydrogels. The monomers of alginate, mannuronic acid, and guluronic acid possess a cyclic structure with high planarity, with hydrogen bonds formed between the monomers to suppress free rotation, thereby making alginate mechanically rigid. DBCO-modified alginate was synthesized via a one-step coupling reaction (Supplementary Fig. 5), and 1H-NMR measurements confirmed the production of Alg-DBCO10, in which each alginate molecule was chemically modified with 10 DBCO groups (Supplementary Fig. 6). Dynamic light scattering (DLS) measurements estimated the hydrodynamic radius of Alg-DBCO10 in an aqueous solution to be 1480 nm (Supplementary Fig. 7).
When Cy3-labeled Alg-DBCO10 was added to the culture medium of N3(+)HskMSCs, whose cell membranes were labeled green with DiO, yellow fluorescence was observed on the cell membrane (Fig. 2b). This confirmed that the azide groups on HskMSCs and Alg-DBCO10 were linked via a click reaction. We then attempted to form ligand-free covalently integrin-linking hydrogels with varying connectivity by reacting HskMSCs (2.0×106 cells) with different numbers of azide groups (2.1 × 1010, 2.6 × 1010, and 3.1 × 1010) with a 2% Alg-DBCO10 aqueous solution (100 µL). The covalent linkages between numerous N3(+)HskMSCs and Alg-DBCO10 would result in a hydrogel state, we performed rheological analysis. The reactants exhibited a hydrogel state, regardless of the number of azide groups introduced, confirming the formation of ligand-free covalently integrin-linking hydrogels (Supplementary Fig. 8). The gelation time after the start of the click reaction (Fig. 2c) and the storage modulus of the hydrogels G’ (Fig. 2d) varied depending on the number of azide groups, indicating that the number of azide groups introduced correlates with connectivity. In this system, in order to form gels, a certain number of azide groups introduced into the integrin and DBCO groups introduced into the alginate must react. However, since alginate has a negative charge due to its carboxyl group, electrostatic repulsion occurs between the alginate and the negatively charged cell membrane, which makes it difficult for the azide group and the DBCO group to approach each other. This could make the click reaction appear slower, and gelation takes a relatively long time. Additionally, we confirmed that the cells in the hydrogels had a high survival rate of approximately 95% (Fig. 2e, 2f).
Next, we examined the introduction of azide groups into integrin. The gene ITGB3, which encodes integrin β3, was knocked down using shRNA. The expression level of integrin β3 in HskMSCs treated with shRNA [shRNA ITGB3(+)HskMSC] was approximately 50% of that in normal HskMSCs (Fig. 2g), confirming the successful generation of ITGB3 knockdown HskMSCs. After treating shRNA ITGB3(+)HskMSCs with Ac4ManNAz, the number of azide groups introduced was analyzed through a reaction with clickable rhodamine. The fluorescence intensity of shRNA ITGB3(+)N3(+)HskMSCs was reduced to approximately 58% of that observed in normal N3(+)HskMSCs treated with Ac4ManNAz under identical conditions (Supplementary Fig. 9). Quantification of the unreacted clickable rhodamine (Supplementary Fig. 10) revealed that the number of azide groups introduced into ITGB3(+)HskMSCs was 1.8×1010 (Supplementary Table 1), which was significantly lower than that introduced into normal N3(+)HskMSCs (3.1 × 1010). These findings indicate the successful introduction of azide groups into integrin β3 for normal N3(+)HskMSCs, with subsequent covalent linkage to DBCO-modified alginate. Thus, we constructed three types of ligand-free covalently integrin-linking hydrogels with varying covalent linking points between the integrin β3 involved in HskMSCs and the hydrogel networks. This allowed us to evaluate the effects of the ligand-free direct linking the integrin β3 of HskMSCs to hydrogel networks on mechanical stress conductive properties, eliciting cellular responses, and tissue regeneration response. In this study, we prepared a control hydrogels without integrin β3−hydrogel linking using alginate-lacking DBCO groups (Supplementary Table 1). A comparison of these hydrogels will facilitate the elucidation of the significance of the covalently direct linking.
Rapid and stable transmittance of mechanical stress to the nucleus in the ligand-free covalently integrin-linking hydrogels
First, we examined the effect of the ligand-free direct linking the integrin β3 of HskMSCs to hydrogel networks on mechanical stress conductive properties. Since it is reported that when cells are received certain mechanical stress the nucleus becomes flattened41, we quantified the nuclear flattening using the Nuclear Shape Index (NSI) in this study42. Mechanical compression was applied to the ligand-free covalently integrin-linking hydrogels for only 10 min. The morphology of the nuclei in HskMSCs located at the middle plane of the hydrogels was then analyzed (Supplementary Fig. 11). In the control hydrogel where there were no covalent linking between the integrin β3 and hydrogel networks (linking points: 0), the nucleus remained round regardless of the applying mechanical compression (Fig. 3a, b), suggesting that mechanical stress did not reach the nucleus. In contrast, within our ligand-free covalently integrin-linking hydrogels, the nuclei were obviously flattened (Fig. 3a, b), indicating that the mechanical stress applied from outside the hydrogels was transmitted to the cells located inside the hydrogel, eventually reaching the nucleus. Significantly, the degree of nuclear flattening increased with higher numbers of azide groups, indicating enhanced connectivity. This suggests that a more covalently linked structure efficiently transmits mechanical stress to cells, thereby increasing the mechanical stress reaching the nucleus.

a CLSM images of the cell nucleus in the ligand-free covalently integrin-linking hydrogels prepared by Alg-DBCO10 and N3(+)HskMSCs with different numbers of introduced azide groups per cell. Blue: DAPI. Scale bars indicate 10 µm. b NSI of the N3(+)HskMSCs in the gels with/without mechanical stress-loading (n = 5, independent samples). Black: without mechanical stress, Red: with mechanical stress. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05, **P < 0.01. Source data is provided as a Source Data file. c CLSM images of the cell nucleus in the gels prepared by Alg-DBCO10 and Alg-RGD50, N3(+)HskMSCs with/without mechanical stress-loading. Blue: DAPI. Scale bars: 10 µm. d NSI of the N3(+)HskMSCs in the gels with/without mechanical stress-loading (n = 5, independent samples). Black: without mechanical stress, Red: with mechanical stress. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and ***P < 0.001. Source data is provided as a Source Data file. e CLSM images of the cell nucleus in the gels prepared by Alg-DBCO10 and N3(+)HskMSCs and shRNA ITGB3(+)N3(+)HskMSCs with/without mechanical stress-loading. Blue: DAPI. Scale bars: 10 µm. f NSI of the N3(+)HskMSCs and shRNA ITGB3(+)N3(+)HskMSCs in the gels with/without mechanical stress-loading (n = 5, independent samples). Black: without mechanical stress, Red: with mechanical stress. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and ***P < 0.001. Source data is provided as a Source Data file. g CLSM images of the cell nucleus in the gels prepared by Alg-DBCO10 and cytochalasin D(+)N3(+)HskMSCs with/without mechanical stress-loading. Blue: DAPI. Scale bars: 10 µm. h NSI of the cytochalasin D( + )N3(+)HskMSCs in the gels with/without mechanical stress-loading (n = 5, independent samples). Black: the number of azide groups introduced is 0, Red: the number of azide groups introduced is 3.1 × 1010. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and **P < 0.01. Source data is provided as a Source Data file. i Immunoblot analysis of normal N3(+)HskMSCs (left) and shRNA LMNA(+)N3(+)HskMSCs treated with antibodies against lamin A/C and α-tubulin as a loading control. j CLSM images of the cell nucleus in the gels prepared by Alg-DBCO10 and N3(+)HskMSCs and shRNA LMNA( + )N3(+)HskMSCs with/without mechanical stress-loading. Blue: DAPI. Scale bars: 10 µm. k NSI of the N3(+)HskMSCs and shRNA LMNA(+)N3(+)HskMSCs in the gels with/without mechanical stress-loading (n = 5, independent samples). Black: without mechanical stress, Red: with mechanical stress. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and ***P < 0.001. Source data is provided as a Source Data file.
To explore in detail the need for this covalent integrin β3−hydrogel networks linking, a conventional hydrogel in which integrin of cells are non-covalently coupled to hydrogels via interactions with specific ligands for integrin, RGDs, was used as control hydrogels. For the purpose, alginate was chemically modified with RGD instead of DBCO (Supplementary Fig. 12), and the number of RGD molecules introduced was quantified using 1H-NMR (Supplementary Fig. 13), resulting in Alg-RGD10 and Alg-RGD50, where 10 and 50 RGD molecules were modified per alginate molecule, respectively. Physically cross-linked hydrogels of Alg-RGD10 and Alg-RGD50 were prepared by adding calcium ions, and the adhesion morphology of HskMSCs seeded on these hydrogels was evaluated to assess integrin β3−RGD connectivity. While cells displayed a round morphology on the Alg-RGD10 hydrogel similar to that on plain alginate, they adhered and exhibited a spread morphology akin to cells on glass on the Alg-RGD50 hydrogel (Supplementary Fig. 14), indicating effective binding between the RGD peptide and integrin β3 of HskMSCs. The number of binding points to integrin β3 present in 100 µL of Alg-RGD50 (2%) was estimated to be 2.2 × 1017 (Supplementary Table 2). Gelation of Alg-RGD50 and HskMSC mixture solutions (Supplementary Fig. 15) confirmed the formation of hydrogel networks via conventional integrin β3−RGD linkages. Note that, within the control hydrogels, the nuclei remained round even when mechanical compression was applied (Fig. 3c, d), suggesting that mechanical stress did not reach the nuclei. In other words, the binding of integrin β3 to RGD, a mechanism that cells naturally possess, was not sufficient to transmit 10 min of mechanical stress to the nucleus of cells located inside the hydrogel. Thus, it is demonstrated that the low efficiency of receiving mechanical stress for the conventional method owing to the weak binding between the ligand modified on the hydrogel and the integrin of the cells.
Next, we investigated the relationship between mechanical stress-related elements on the cell side (integrins, actin filaments, and nuclear lamina) and mechanical stress conductive properties. Initially, we examined integrins that are covalently linked to hydrogel and receive mechanical stress. In the control hydrogels prepared using shRNA ITGB3(+)N3(+)HskMSCs and Alg-DBCO10, the nucleus maintained a round shape even when mechanical stress was applied (Fig. 3e, f). These findings demonstrate that when integrin β3 is knocked down, mechanical stress applied from outside the hydrogel fails to reach the nucleus of cells located inside the hydrogel. Thus, it is evident that, in our ligand-free covalently integrin-linking hydrogels, integrin β3 serves as the recipient of mechanical stress from the covalently linked hydrogel networks, acting as an input element into the cell.
Next, we focused on actin filaments as an element that transmits mechanical stress input from integrin β3 into the nucleus. When the ligand-free covalently integrin-linking hydrogel was treated with the actin polymerization inhibitor cytochalasin D and compressed, the nuclei of the cells within the hydrogel remained round and did not flatten (Fig. 3g, h). This outcome demonstrated that mechanical stress was nullified when actin filament polymerization was inhibited, preventing its transmission to the nucleus, despite its initial input into the cells. Thus, in the ligand-free covalently integrin-linking hydrogels, actin filaments function as crucial elements transmitting mechanical stress from integrin β3 to the nucleus.
To elucidate the specific location within the nucleus where the mechanical stress acts, we focused on the nuclear lamina (lamina A/C), which mechanically supports the nuclear membrane and upholds the nuclear shape. We generated HskMSCs in which the lamin A/C gene LMNA was knocked down using shRNA [shRNA LMNA(+)HskMSCs]. The expression level of lamin A/C in shRNA LMNA(+)HskMSCs was approximately 30% of that observed in normal HskMSCs (Fig. 3i). Additionally, we confirmed a significant reduction in the fluorescence derived from lamin A/C through immunofluorescence staining at the cellular level (Supplementary Fig. 16A, 16B). Moreover, in shRNA LMNA(+)HskMSCs, the nuclear area expanded approximately two-fold owing to structural support defects (Supplementary Fig. 16C), confirming the generation of LMNA-knockdown HskMSCs. Subsequently, we analyzed the azide groups introduction into shRNA LMNA(+)HskMSCs using clickable rhodamine. The fluorescence intensity of azide-modified shRNA LMNA(+)HskMSCs was comparable to that of normal HskMSCs under the same conditions (dotted line in Supplementary Fig. 17). The number of azide groups introduced into shRNA LMNA(+)HskMSCs was estimated to be 3.1 × 1010 (Supplementary Table 3) by quantifying unreacted clickable-rhodamine in the supernatant (Supplementary Fig. 18). It was confirmed that the number of azide groups introduced into the cells did not change, even when lamin A/C was knocked down. In the ligand-free covalently integrin-linking hydrogels constructed using shRNA LMNA(+)N3(+)HskMSCs and Alg-DBCO10, the nuclei remained round and did not flatten, even when mechanical stress was applied (Fig. 3j, k). These results suggest that lamin A/C knockdown prevents the nucleus from flattening despite mechanical stress reaching the nucleus, suggesting that the mechanical stress that reaches the nucleus acts on the nuclear lamina to change the morphology of the nucleus.
By integrating these results, we demonstrated that in our ligand-free covalently integrin-linking hydrogels, the mechanical stress applied from outside the hydrogel was first channeled into the cell via integrin β3, which is covalently linked to alginate hydrogel networks. Mechanical stress is subsequently transmitted to lamin A/C within the nucleus through the cytoplasm via the actin network. This sequential process induces significant deformation of the nuclear morphology.
Mechanical transmittance to the nucleus elicits cellular regenerative responses
We investigated whether mechanical stress reaching the nucleus triggers cellular regenerative responses. It is well known that, in conventional method, the extracellular mechanical stress received via integrin–ligand binding is converted into a biochemical cascade in the cytoplasm over several hours. For instance, nuclear translocation of the transcription coactivator YAP, which is involved in the Hippo pathway, has a mechanism to control muscle fiber size, but is known to take a long time to affect within living tissues43. This process ultimately leads to the nuclear translocation of YAP and subsequent changes in cellular responses via the modulation of its target gene expression44,45,46,47,48. By contrast, as mentioned above, the extracellular mechanical stress directly reached to the nucleus of cells and altered its morphology in our ligand-free covalently integrin-linking hydrogel. Considering this, we hypothesized that YAP translocation occurs rapidly without typical slow conversion into a biochemical cascade pathway. To test this hypothesis, we subjected the ligand-free covalently integrin-linking hydrogels to mechanical stress for only 10 min and analyzed the intracellular dynamics of YAP (Supplementary Fig. 19). In the control hydrogel lacking integrin−hydrogel covalent linkages (the number of linking point: 0), YAP predominantly resided in the cytoplasm, regardless of applying mechanical stress, with a low nuclear localization rate of approximately 40% (Fig. 4a, b). Similarly, in the ligand-free covalently integrin-linking hydrogels with 2.1 × 1010 and 2.6 × 1010 azide groups, although there was a slight change in nuclear morphology under mechanical stress (Fig. 3a), the nuclear localization rate of YAP remained around 40% (Fig. 4a, b). In contrast, in the ligand-free covalently integrin-linking hydrogel with 3.1 × 1010 azide groups, the nuclear morphology was most deformed under the mechanical cues, and YAP translocation into the nucleus was significantly promoted, with the nuclear localization rate being high at approximately 70% (Fig. 4a, b). We also examined the nuclear localization rate of YAP in conventional hydrogels which mechanical stress is solely transmitted via integrin β3–ligand binding using Alg-RGD50 hydrogels. In this control hydrogel, YAP predominantly resided in the cytoplasm, regardless of applying mechanical stress, with a low nuclear localization rate of approximately 40% (Fig. 4c, d). These results demonstrate that in the ligand-free covalently integrin-linking hydrogel with enough high integrin β3−hydrogel connectivity (3.1 × 1010 azide groups), short-term transient mechanical stress load alone can promote rapid and reliable nuclear translocation of YAP, akin to the direct application of mechanical stress to the nucleus using AFM49. This suggests that the degree of change in nuclear morphology due to the direct transmission of mechanical stress to the nuclear lamina is strongly related to the ability of YAP to translocate to the nucleus.

a CLSM images of N3(+)HskMSCs in the gels with varying covalent connectivity treated with anti-YAP antibody with/without mechanical stress-loading. Green: YAP and blue: DAPI. Scale bars indicate 10 μm. b Nuclear/cytoplasm localization ratio of YAP in N3(+)HskMSCs in the gels with varying covalent connectivity (n = 5, independent samples). Black: the number of azide groups introduced is 0, Yellow: the number of azide groups introduced is 2.1 × 1010, Green: the number of azide groups introduced is 2.6 × 1010, Red: the number of azide groups introduced is 3.1 × 1010. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and **P < 0.01. Source data is provided as a Source Data file. C CLSM images of N3(+)HskMSCs/Alg-RGD50 in the gels treated with anti-YAP antibody and mechanical stress-loading. Green: YAP and Blue: DAPI. Scale bars indicate 10 μm. d Nuclear/cytoplasm localization ratio of YAP in N3(+)HskMSCs/Alg-RGD50 in the gels (n = 5, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and **P < 0.01. Source data is provided as a Source Data file. e CLSM images of shRNA LMNA(+)N3(+)HskMSCs in the gels treated with anti-YAP antibody and mechanical stress-loading. Green: YAP and Blue: DAPI. Scale bars indicate 10 μm. f Nuclear/cytoplasm localization ratio of YAP in shRNA LMNA(+)N3(+)HskMSCs in the gels (n = 5, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05. Source data is provided as a Source Data file. g CLSM images of importazole(+)N3(+)HskMSCs in the gels treated with anti-YAP antibody and mechanical stress-loading. Green: YAP and Blue: DAPI. Scale bars indicate 10 μm. h Nuclear/cytoplasm localization ratio of YAP in importazole(+)N3(+)HskMSCs in the gels (n = 5, independent samples). Data are presented as mean ± SD. i CLSM images of FITC-dextran70k-loading N3(+)HskMSCs in the gels treated with/without mechanical stress-loading. Green: FITC-dextran70k and blue: DAPI. Scale bars indicate 10 μm. j Nuclear/cytoplasm localization ratio of FITC-dextran70k in N3(+)HskMSCs in the 3D network structures (n = 5, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and **P < 0.01. Source data is provided as a Source Data file.
For further investigation, we constructed a ligand-free covalently integrin-linking hydrogel using shRNA LMNA(+)N3(+)HskMSCs with 3.1 × 1010 azide groups, lamin A/C knockdown, and examined the nuclear translocation of YAP under mechanical stress. In this hydrogel, the nuclear localization rate of YAP remained constant at approximately 40% regardless of the mechanical cue (Supplementary Fig. 20). This rate was significantly lower than the nuclear localization rate of approximately 70% observed in the ligand-free covalently integrin-linking hydrogel using normal N3(+)HskMSCs under mechanical stress (Fig. 4e, f). This suggests that effective nuclear translocation of YAP requires mechanical stress to be transmitted to the nuclear lamina, significantly deforming nuclear morphology. Because importin is known to drive the nuclear translocation of YAP50, we examined the relationship between YAP nuclear translocation in our hydrogels and importin. We treated a ligand-free covalently integrin-linking hydrogel with 3.1 × 1010 azide groups with the importin inhibitor, importazole, compressed the structure for 10 min to apply mechanical stress, and evaluated YAP nuclear translocation. The nuclear localization rate of YAP remained unchanged, even when importin was inhibited (Fig. 4g, h). This suggests that the observed promotion of the nuclear translocation of YAP in our hydrogels was not mediated by importin. Instead, it appeared to be due to the deformation of nuclear pores caused by mechanical stress directly transmitted to the nuclear lamina, leading to significant changes in nuclear morphology. To test this hypothesis, we introduced dextran (MW: 40 kDa)-FITC, a molecule that does not spontaneously translocate into the nucleus, into HskMSCs located inside our hydrogel with 3.1 × 1010 azide groups. We examined the effects of mechanical stress on the translocation of the dextran into the nucleus. When mechanical stress was applied for 10 min, dextran-FITC translocation into the nucleus was significantly enhanced compared to that in the absence of mechanical stress (Supplementary Fig. 21). Additionally, when dextran (MW: 70 kDa)-FITC, which has a molecular weight similar to that of YAP, was introduced, the nuclear translocation significantly increased under mechanical stress (Fig. 4i, j). This suggests that the observed promotion of YAP’s nuclear translocation in our developed hydrogels results from the deformation and expansion of nuclear pores caused by mechanical stress directly transmitted to the nuclear lamina.
Next, we evaluated whether nuclear translocation of YAP in the ligand-free covalently integrin-linking hydrogel induced changes in gene expression levels and cellular responses. Specifically, we assessed muscle differentiation of HskMSCs by examining the expression level of MyoD1, a master myogenic transcription factor, play an integral role in muscle differentiation. Three types of HskMSCs-loading hydrogels with different covalent connectivity were cultured in a medium devoid of biochemical muscle differentiation inducers and subjected to mechanical stress similar to that generated in skeletal muscle tissue for 5 days. Subsequently, MYOD1 and its encoding protein, MyoD1, were quantitatively analyzed. In the control hydrogel without integrin β3−hydrogel covalent connections (the number of linking point: 0), MyoD1 protein expression was low regardless of the presence or absence of mechanical stress, indicating a lack of muscle differentiation (Fig. 5a, b). Similarly, in our developed hydrogels with 2.1 × 1010 and 2.6 × 1010 azide groups, MyoD1 protein (Fig. 5a, b) and MYOD1 gene (Fig. 5c) expression levels remained low irrespective of mechanical stress, suggesting that muscle differentiation did not occur in these conditions. In contrast, within our developed hydrogel with 3.1 × 10¹⁰ azide groups, the expression level of the protein MyoD1 significantly increased only in the presence of mechanical stress (Fig. 5a, b), and the expression level of the gene MYOD1 also showed a significant increase (Fig. 5c, d). These results align well with the degree of nuclear morphological deformation and the nuclear localization rate of YAP. In other words, in ligand-free covalently integrin-linking hydrogel with enough high integrin β3−hydrogel linking points, mechanical stress alone was sufficient to significantly increase the expression levels of differentiation-related genes and proteins, leading to muscle differentiation even in the absence of biochemical differentiation-inducing factors. This demonstrated the potential of our developed hydrogel to induce cellular responses through mechanical stress, highlighting its promise for advancing tissue engineering.

a CLSM images of N3(+)HskMSCs in the gels with varying covalent connectivity treated with Alexa Fluor 488-labeled anti-MyoD1 antibody and with/without mechanical stress-loading. Green: MyoD1 and blue: DAPI. Scale bars indicate 50 μm. b Fluorescence intensities of Alexa Fluor 488-labeled anti-MyoD1 antibody in N3(+)HskMSCs in the gels (n = 10, independent samples). Black: without mechanical stress, Red: with mechanical stress. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and ***P < 0.001. Source data is provided as a Source Data file. c Relative expression level of MYOD1 in N3(+)HskMSCs in the gels with varying covalent connectivity and with mechanical stress-loading (n = 3, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05 and **P < 0.01. Source data is provided as a Source Data file. d Relative expression level of MYOD1 in N3(+)HskMSCs in the gels with/without mechanical stress-loading (n = 3, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and **P < 0.01. Source data is provided as a Source Data file. e CLSM images of shRNA ITGB3(+)N3(+)HskMSCs in the gels treated with Alexa Fluor 488-labeled anti-MyoD1 antibody with mechanical stress-loading. Green: MyoD1 and blue: DAPI. Scale bars indicate 50 μm. f Fluorescence intensities of Alexa Fluor 488-labeled anti-MyoD1 antibody in shRNA ITGB3(+)N3(+)HskMSCs in the gels (n = 10, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and ***P < 0.001. Source data is provided as a Source Data file. g CLSM images of shRNA LMNA(+)N3(+)HskMSCs in the gels treated with Alexa Fluor 488-labeled anti-MyoD1 antibody with mechanical stress-loading. Green: MyoD1 and blue: DAPI. Scale bars indicate 50 μm. h Fluorescence intensities of Alexa Fluor 488-labeled anti-MyoD1 antibody in shRNA LMNA(+)N3(+)HskMSCs in the gels (n = 10, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and ***P < 0.001. Source data is provided as a Source Data file.
For further investigation, we examined the effects of knocking down integrin β3 and lamin A/C, on these responses. In the ligand-free covalently integrin-linking hydrogels constructed with shRNA ITGB3(+)N3(+)HskMSCs, where integrin β3 was knocked down, mechanical stress failed to induce nuclear translocation of YAP (Supplementary Fig. 22). Furthermore, the expression level of MyoD1 was significantly lower compared to hydrogels using normal N3(+)HskMSCs (Fig. 5e, f). This indicates that mechanical stress transmittance via integrin β3−hydrogel covalent linkages is necessary for initiating the downstream cellular responses required for muscle differentiation. Similarly, in the hydrogel constructed with shRNA LMNA(+)N3(+)HskMSCs, where lamin A/C was knocked down, the expression level of MyoD1 was notably reduced compared to hydrogels using normal N3(+)HskMSCs (Fig. 5g, h). These findings indicate that effective muscle differentiation of HskMSCs in our hydrogels hinges upon the efficient transmission of mechanical stress to the nuclear lamina. Based on the results mentioned above, it is revealed that rapid and reliable transmittance of mechanical stress to the nucleus of HskMSCs, achieved by ligand-free covalently linking between the integrin β3 and scaffolding hydrogel with high connectivity, is crucial for eliciting robust cellular regenerative responses.
Regenerative capabilities of ligand-free covalently integrin-linking hydrogels
We employed a striated muscle defect model using female nude mice to assess tissue regenerative capability of ligand-free covalently integrin-linking hydrogels. Part of the skeletal muscle tissue (100 mg) from the hind limb of mice was resected, and in this model, approximately 58% of the skeletal muscle in the hind limb is removed, which is a relatively severe injury. We injected ligand-free covalently integrin-linking hydrogels composed of DiI-labeled N3(+)HskMSCs (2.0 × 106 cells) with varying numbers of azide groups (2.1 × 1010, 2.6 × 1010, 3.1 × 1010) and a 2% Alg-DBCO10 aqueous solution (100 µL) into the defect site (Supplementary Fig. 23). Our developed hydrogels allowed reactants to remain in a liquid state until a click reaction proceeded, facilitating the construction of an injectable hydrogel at the injection site within the body. First, the function of the reconstructed muscle tissue was evaluated from muscle force measurements to ensure that the function of the reconstructed muscle tissue was comparable to that of normal skeletal muscles, because several studies have reported that grip strength measurements were used to evaluate the functional recovery of muscle tissue with muscle dysfunction51,52. Muscle strength analysis after 14 days revealed that the hydrogel with enough high integrin β3−hydrogel covalent connectivity (azide group number: 3.1 × 1010) exhibited significantly higher muscle strength recovery compared to hydrogels lacking integrin β3−hydrogel covalent linking (number of linking points: 0) or those with lower connectivity (azide group numbers: 2.1 × 1010, 2.6 × 1010) (Fig. 6A). Muscle strength, treated with the hydrogel with enough high integrin β3−hydrogel covalent connectivity (azide group number: 3.1 × 1010), recovered to approximately 90% of normal muscle strength prior to injury. To investigate the effect of gender differences, the regeneration efficiency of the gel was evaluated using male nude mice, and the regeneration efficiency of skeletal muscle was found to be equivalent to that of females (Supplementary Fig. 24).

a Muscle force recovery ratio of femoral skeletal muscle tissue-injured mice after the treatments with the gels prepared with Alg-DBCO10 and N3(+)HskMSCs with varying covalent connectivity (n = 3, independent samples). Black: the number of azide groups introduced is 0, Yellow: the number of azide groups introduced is 2.1 × 1010, Green: the number of azide groups introduced is 2.6 × 1010, Red: the number of azide groups introduced is 3.1 × 1010. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05. Source data is provided as a Source Data file. b CLSM images for sections of the reconstructed muscle tissues at day 14 after the treatments with the gels with varying covalent connectivity. HskMSCs were labeled red color with DiI. Tissue sections were stained with Hematoxylin & Eosin (HE) and Masson’s-Trichrome techniques. Scale bars indicate 500 µm. c Area (%) of muscle fibers in the reconstructed tissues at day 14 after the treatments with the gels with varying covalent connectivity. Dotted line indicates area (%) of muscle fibers for healthy muscle tissue (n = 3). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05 and **P < 0.01. Source data is provided as a Source Data file. d Cross-sectional area of muscle fibers in the reconstructed tissues at day 14 after the treatments with the gels with varying covalent connectivity. Dotted line indicates cross-sectional area of muscle fibers for healthy muscle tissue (n = 3). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P 0.05 and **P < 0.01. Source data is provided as a Source Data file. e Area (%) of collagen fibers deposited in the reconstructed tissues at day 14 after the treatments with the gels with varying covalent connectivity. Dotted line indicates area (%) of collagen fibers deposited in healthy muscle tissue (n = 3). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05 and **P < 0.01. Source data is provided as a Source Data file.
Hereafter, experimental results obtained using female nude mice are presented. After 14 days using female mice, the reconstructed tissues were excised (Supplementary Fig. 25), and weighted. The weight of the muscle tissue reconstructed by the hydrogel with enough high integrin β3−hydrogel covalent connectivity was approximately 96 mg, which means that the reconstruction rate was approximately 96%. It was found that most of the injured skeletal muscles were reconstructed without overgrowth or degeneration. Moreover, the reconstructed tissues were sectioned, and analyzed using Hematoxylin and Eosin (H&E) and Masson’s trichrome staining (Fig. 6b). The control hydrogel lacking integrin β3−hydrogel covalent linking and our hydrogels with lower connectivity showed significantly lower muscle tissue content (<20% of tissue), smaller myofibers cross-sectional area, and abnormal collagen fiber deposition (Fig. 6c–e). In contrast, skeletal muscles reconstructed by our hydrogels with high covalent connectivity demonstrated a significantly higher proportion of myofibers (Fig. 6c) and a muscle cross-sectional area similar in size (Fig. 6d) to those found in normal skeletal muscles. Furthermore, there were no signs of abnormal collagen fiber deposition (Fig. 6e).
Notably, the reconstructed myofibers by our developed hydrogel with enough high covalent connectivity exhibited red fluorescence from DiI, and the percentage for the DiI-positive myofibers is more than 94%. This indicates that the transplanted HskMSCs differentiated into muscle cells within the hydrogel and organized into muscle fibers. Immunofluorescence staining with phalloidin and myosin heavy chain (MYH3) further confirmed the alignment of actin filaments and MYH3 in the regenerated muscle fibers, resembling normal skeletal muscle structure (Fig. 7a, b). Tensile testing of the regenerated muscle tissue revealed a maximum tensile strength at tissue breakage similar to normal muscle tissue (Supplementary Fig. 26), suggesting comparable mechanical strength. Namely, the ligand-free covalently integrin-linking hydrogels demonstrated high capability in regenerating muscle tissue in vivo. The high capability would be attributed to the hydrogel’s ability to transmit mechanical stress generated in the muscle tissue efficiently to the nuclear lamina of the transplanted HskMSCs, facilitated by its high integrin β3−hydrogel covalent connectivity. We hypothesized that muscle regeneration would be impaired if the stretching movement of skeletal muscles, which is the source of mechanical stress, was suppressed. To test this hypothesis, we constructed a mouse model with injured sciatic nerve and hindlimb skeletal muscles and (Supplementary Fig. 27) and injected a hydrogel with the highest integrin β3−hydrogel covalent connectivity into the defect site and assessed muscle regeneration efficiency. The results revealed low muscle tissue content in the reconstructed tissue, abnormal collagen fiber deposition, and no recovery of muscle strength even after 30 days (Supplementary Fig. 28). This aligns with our hypothesis and revealed that the ligand-free covalently integrin-linking hydrogels can effectively utilize tissue-generated mechanical stress to induce a regenerative response of transplanted cells.

a Sections of reconstructed muscle tissues along the longitudinal axis of the muscle fiber at day 14 after the treatments with the gels with/without covalent connection (n = 3, independent samples). HskMSCs were labeled green color with DiO. These sections were immunostained with Alexa Fluor 594-labeled phalloidin. Green: DiO, Blue: DAPI, and Red: phalloidin. Scale bars: 50 µm. b Sections of reconstructed muscle tissues along the longitudinal axis of the muscle fiber at day 14 after the treatments with the gels with/without covalent connection (n = 3, independent samples). HskMSCs were labeled green color with DiO. The sections were immunostained with Alexa Fluor 546-labeled anti-MYH3. Green: DiO, Blue: DAPI, Red: MYH3. Scale bars: 50 µm.
To compare the tissue regenerative capability of our developed hydrogels with conventional hydrogel which mechanical stress is solely transmitted via usual integrin β3–ligand binding, we exmained tissue regenerative capability of the hydrogel constructed using Alg-RGD50 and normal HskMSCs. The conventional hydrogel exhibited a significantly lower muscle force recovery rate (Fig. 8a) by comparing that of our developed hydrogels. The reconstructed tissue (Supplementary Fig. 29) was histologically analyzed using H&E and Masson’s trichrome staining after 14 days (Fig. 8b). As expected, the reconstructed tissue by the conventional hydrogel showed significantly lower myofiber content (Fig. 8c) and higher collagen fiber deposition (Fig. 8d). These findings clearly demonstrate the essential role of stable covalent linking between the integrin of transplanted cells and scaffolding hydrogel in effectively transmitting mechanical stress generated by tissue dynamics.

a Muscle force recovery ratio of femoral skeletal muscle tissues injured mice after the treatments with the gels prepared with Alg-DBCO10 solution and N3(+)HskMSCs treated with Ac4ManNAz for 48 h and control HskMSCs-loaded Alg-RGD50 gel (n = 3). Yellow: Alg-RGD50, Red: Alg-DBCO10. Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05 and **P < 0.01. Source data is provided as a Source Data file. b CLSM images of sections of the reconstructed muscle tissues at day 14 after the treatments with the gels prepared with Alg-DBCO10 and N3(+)HskMSCs and control HskMSCs-loading Alg-RGD50 gels. HskMSCs were labeled red color with DiI. Scale bars indicate 500 µm. c Area (%) of muscle fibers in the reconstructed tissues at day 14 after the treatments with the gels prepared with Alg-DBCO10 and N3(+)HskMSCs and control HskMSCs-loading Alg-RGD50 gels. Dotted line indicates area (%) of muscle fibers for healthy muscle tissue (n = 3). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05. Source data is provided as a Source Data file. d Area of collagen fibers deposited in the reconstructed tissues at day 14 after the treatments with the gels prepared with Alg-DBCO10 and N3(+)HskMSCs and control HskMSCs-loading Alg-RGD50 gels. Dotted line indicates area (%) of collagen fibers deposited in healthy muscle tissue (n = 3). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and **P < 0.01. Source data is provided as a Source Data file.
Validity of ligand-free covalently integrin-linking hydrogels as promising tissue engineering technology
Finally, we assessed the potential utility of our developed hydrogels as an advanced in vivo tissue engineering technology. Matrigel, a representative cell transplantation hydrogel containing various extracellular matrices and growth factors known to promote tissue regeneration through biochemical stimuli53,54,55, was used as a positive control. We injected ligand-free covalently integrin-linking hydrogel composed of DiI-labeled N3(+)HskMSCs (2.0 × 106 cells) with highly introduced azide groups (3.1 × 1010) and a 2% Alg-DBCO10 aqueous solution (100 µL) or Matrigel encapsulating the same number of HskMSCs into the defect site of skeletal muscle injured mice. Tissue reconstruction (Supplementary Fig. 30) and muscle force recovery (Supplementary Fig. 31) were analyzed after 14 days. Compared to Matrigel, our developed hydrogel exhibited a significantly higher muscle force recovery. Moreover, the reconstructed tissue showed significantly greater myofiber content (Supplementary Fig. 32A, 32B) and reduced collagen fiber deposition (Supplementary Fig. 32A, 32C). MYH3 expression was not detected in the tissue reconstructed with HskMSC-encapsulated Matrigel (Supplementary Fig. 33). These results demonstrate that the ligand-free covalently integrin-linking hydrogel possesses significantly superior tissue regeneration capability compared to Matrigel. Matrigel’s tissue regenerative capability solely relies on biochemical stimulation provided by biomolecules involved. In contrast, the ligand-free covalently integrin-linking hydrogel employs alginate as hydrogel-forming biopolymers, which does not provide any biochemical cues to the cells. Considering this, its high tissue regenerative capability can be mainly caused by the rapid and reliable mechanical stress transmittance capability.
To assess the systemic impact of the ligand-free covalently integrin-linking hydrogel in vivo, we performed blood tests on a mouse model of muscle injury treated with the hydrogels. The results showed values for all test items within the reference ranges, comparable to those of untreated mice (Fig. 9), indicating biocompatibility of our hydrogels56.

Yellow bands indicate the reference value for healthy mice (n = 3, independent samples). Data are presented as mean ± SD. Two-sided Student’s t test was used to analyze the significance, and *P < 0.05. Source data is provided as a Source Data file.
Discussion
Elosegui-Artola et al. reported that mechanical stress applied directly to the nucleus from outside the cell using AFM deforms the nuclear membrane pore, causing YAP to migrate into the nucleus, resulting in rapid cellular responses49. However, this approach is impractical for in vivo applications due to the need for individual cell manipulation and the limitations of AFM in living organisms. In response to these challenges, we attempted to create a scaffolding hydrogel that can rapidly, efficiently and reliably transmit mechanical stress generated in the body to the nuclei of transplanted cells located inside the hydrogel, eliciting a regenerative response from the cells (Fig. 10). In this hydrogel, mechanically rigid alginate was selected as hydrogel networks to effectively receive mechanical stress, facilitating transmittance it to integrin β3 covalently linked to alginate hydrogel networks, and subsequently channeled into the cell. Mechanical stress propagates through the cytoplasm via the actin network, reaching lamin A/C in the nucleus and causing significant nuclear deformation. Within only 10 min of applying mechanical stress, this hydrogel stimulates YAP nuclear translocation in HskMSCs within the hydrogel, promoting differentiation towards muscle cells. Moreover, a portion of the mechanical stress is redistributed among neighboring cells via three-dimensional alginate networks, enhancing overall tissue regeneration efficiency. Thus, we found that when cells were exposed to mechanical stress in the ligand-free covalently integrin-linking hydrogel, the mechanical stress was rapidly and reliably transmitted to the nucleus through a mechanism that was essentially different from previously reported mechanotransduction. Moreover, we discovered that this resulted in a regenerative response in the transplanted cells with high efficiency.

Schematic illustration of the features of the ligand-free covalently integrin-linking hydrogels revealed from the data shown in this article.
By contrast, previously reported conventional hydrogel utilize the non-covalent, weak, reversible, cell cycle and extracellular environment-dependent integrin−ligand binding, a mechanism that cells naturally possess. Our developed hydrogel is fundamentally different from conventional hydrogel in that the integrins of the transplanted cells are directly bound to the hydrogel via stable and irreversible covalent bonds so that the integrin–hydrogel bond is always in an ON state. This linkage structure can highly efficiently receive mechanical stress generated in the body regardless of the intensity of the stress, the cell cycle, or the extracellular environment. Thus, the ligand-free covalently integrin-linking hydrogel represents a significant advancement over conventional scaffolding hydrogels reliant on biochemical stimuli. Alginate, the polymer used, is highly biocompatible57,58, as confirmed by safety testing. Our developed hydrogel is notable for its versatility and scalability. It can be implemented in any cell type capable of incorporating azide groups through metabolic glycoengineering. Azide groups can be introduced into diverse cell types, including mesenchymal stem cells and iPS cells59,60,61,62,63,64, making our developed hydrogel highly adaptable and scalable. Thus, our developed hydrogel stands as a next-generation tissue engineering technology applicable to living organisms, especially for the tissue constantly generating mechanical stress including skeletal muscle, heart, tendon, cartilage, and skin.
Methods
Ethical statement
All animal experiments and all experimental protocols were approved by Konan University Animal Experiment Committee (Approval number: K-23-13) and conformed to the Guidelines for the Care and Use of Laboratory Animals published by the National Institutes of Health.
Materials
D-Mannosamine hydrochloride, 2-azidoacetic acid, acetic anhydride, and pyridine were purchased from Sigma-Aldrich. Sodium alginate (Mw: 250,000) were kindly supplied from Mochida Pharmaceutical Co., Ltd. 4-(4,6-Dimethoxy-1,3,5-triazin-2-yl)−4-methylmorpholinium chloride n-hydrate (DMT-MM) was purchased from Wako Pure Chemical. DBCO-carboxyrhodamine 110 and dibenzylcyclooctyne-PEG4-amine (DBCO-PEG4-amine) were purchased from Click Chemistry Tools. Poly-L-lysine hydrochloride (Poly-L) was purchased from Peptide Institute, Inc. Other reagents and solvents available in extra-pure grade were obtained commercially and used without further purification.
Cell culture
Human skeletal muscle satellite cells (HskMSCs) were purchased from ScienCell Research Laboratories. Poly-L solution was added to sterile water to a predetermined concentration (2 µg/cm2) and coated at 37 °C in a humidified atmosphere containing 5.0% CO2 for one day. Poly-L solution was removed and washed twice with phosphate buffered saline (PBS). HskMSCs were cultured in Poly-L-coated dishes in Skeletal Muscle Cell Medium (SkMCM) (ScienCell Research Laboratories) supplemented with 5% FBS and antibiotic solution containing penicillin (100 units/mL), skeletal muscle cell growth supplement (100 units/mL) at 37 °C in a humidified atmosphere containing 5.0% CO2.
Preparation and quantification of azide-modified HskMSC
HskMSC (2.0 × 104) were seeded on 30 mm glass bottom dish, and then 2 mL of SkMCM containing Ac4ManNAz (100 µM) was added to the dish and incubated at 37 °C for 12, 24, 48, and 72 h. The supernatant was removed and then 2 mL of SkMCM was freshly added to the dish. DBCO-carboxyrhodamine 110 (final concentration: 5 µM) was added to the dish and incubated at 37 °C for 1 h. HskMSC attached on the dish were washed twice with PBS and 1 mL of Live Cell Imaging Solution (Life Technologies) was added, and then the cells were observed by confocal laser scattering microscopy (CLSM, Nikon A1R). To prevent saturation of the intensity, we firstly set the gain to adjust the detector signal using HskMSC incubated with Ac4ManNAz for 48 h at 37 °C providing maximum fluorescence intensity. After that, CLSM observations of the HskMSC treated with different culture days of Ac4ManNAz were performed using the same gain value. Also, CLSM observations of HskMSC (2.0 × 104) treated with Ac4ManNAz for different culture days (12, 24, and 48 h) with different concentrations of DBCO-carboxyrhodamine 110 (1, 3, 5, 7, 10, and 15 μM) were then performed with the same gain values. The fluorescence intensity is represented as the average of nine cells analyzed, and these are expressed as mean ± SD.
Synthesis of alkyne-modified alginate polymer
DBCO-PEG4-amine (2.1 mg, 4.01 μmol) and DMT-MM (1.3 mg, 4.70 μmol) dissolved in 1 mL of pure water was added to 30 mL of Alg (100 mg, 0.40 μmol) solution, and stirred at room temperature for 1 week. The reaction mixture was dialyzed (MWCO: 14,000) against pure water for 2 days, then the resultant solution was freeze-dried to give white powder of DBCO-modified sodium alginate (Alg-DBCO). Molecular structure of Alg-DBCO was determined by 1H-NMR analysis (D2O, 90 °C). The hydrodynamic radius of Alg-DBCO10 solutions (0.05%) in pure water were measured by dynamic light scattering (DLS, ZETASIZER NanoSeries ZEN-3600, Malvern).
Synthesis of RGD-modified alginate polymer
Gly-Arg-Gly-Asp-Ser-Pro (GRGDSP) (11.7 mg, 20 μmol), EDC (4.6 mg, 24.0 μmol) and Sulfo-NHS (10.4 mg, 48.0 μmol) dissolved in 1 mL of pure water was added to 30 mL of Alg (100 mg, 0.40 μmol) solution, and stirred at room temperature for 1 week. The reaction mixture was dialyzed (MWCO: 14,000) against pure water for 2 days, then the resultant solution was freeze-dried to give white powder of RGD-modified sodium alginate (Alg-RGD). Molecular structure of Alg-RGD was determined by 1H-NMR analysis (D2O, 90 °C).
Morphology of cells cultured on RGD-modified alginate hydrogels
Solutions of Alg-RGD10 and Alg-RGD50 (total volume: 50 μL) were placed on glass-bottomed petri dishes and incubated at 37 °C for 3 h. Two mL of SkMCM solution in the presence of CaCl2 solution (0.5%) was added on the alginate solutions gently and incubated for 3 h at 37 °C to give Alg-RGD10 and Alg-RGD50 physical cross-linked hydrogels. After removing SkMCM solution (2 mL), HskMSCs (1.0 × 105/100 µL) were seeded on the Alg-RGD10 and Alg-RGD50 gels and cultured for 4 h at 37 °C in a humidified atmosphere containing 5.0% CO2. After a predetermined time, the cultured cells were observed by confocal microscope (CLSM, Nikon A1R).
Bioorthogonal reaction between azide-modified cells and Alg-DBCO10
Pellet of azide-modified HskMSCs (2.0 × 106) treated with Ac4ManNAz for 48 h was suspended with 100 μL of Cy3-labaled Alg-DBCO10 solution (2%, w/v) in HEPES buffer and cells were completely dispersed by gentle pipetting at 37 °C and incubated at 37 °C for 3 h. The cell dispersion was washed with PBS to remove unreacted Cy3-labaled Alg-DBCO10. SkMCM was added, and cell dispersion were seeded to Poly-L-lysine coated glass bottomed dishes, incubated in an incubator for one day, and observed by confocal microscope (CLSM, Nikon A1R).
Retroviral vectors and retroviral infection
To generate retroviruses encoding shRNAs against ITGB3, LMNA, human ITGB3 target sequence, 5’-CCTGCACCTTTAAGAAAG-3’, human LMNA target sequence, 5’-GAGAUCGAUAACGGGAAGC-3’, were cloned into a pSuper retro puro (Oligoengine, Seattle, WA) vector. The retrovirus vector encoding shRNA was co-transfected with pAmpho (TAKARA) helper plasmid into HEK293T cells in 10 cm dish using the PEI Max transfection regent (Polyscience Inc.). Each plasmid was mixed by 2 µg and then mixed into 240 µL of Optimem (gibco). It was then mixed with 16 µL of PEI max, subsequently incubate for 15 min, and then used at the transfection. At 24 h after transfection, the medium was changed the 10 mL of fresh medium, and 24 h later, supernatants containing virus were collected. The supernatants containing virus were filtered using the 0.45 µm of Millex-HP (Merck Millpore). The medium in the presence of 8 µg/mL polybrene were added to 6 well dish cultured HskMSCs. Subsequently, 6 well dish was centrifuged at 150 × g for 1 h in AX-521 centrifuge (TOMY). Infected cells were selected using puromycin (1 µg/mL) for 3 days.
Immunoblot analysis
Cells were solubilized with protein extraction buffer (50 mM Tris pH 7.4, 150 mM NaCl, 1% Triton X-100, 1% SDS, 10 mM EDTA, 1 mM Na3VO4, 10 mM NaF), and protease inhibitor cocktail (Nacalai Tesque, Inc., Kyoto, Japan), sonicated, and centrifuged at 20,000 × g for 15 min. Supernatants were used as whole cell extracts and subjected to SDS-polyacrylamide gel electrophoresis (SDS-PAGE). Integrin β3/ITGB3/CD61 Antibody (D-11: sc-365679, this antibody is recommended for detection of Integrin β3 of mouse, rat, human and origin by WB, IP, IF, IHC(P), and ELISA, Santa Cruz Biotechnology, Inc.). Lamin A/C Monoclonal Antibody (mab636: MA3-1000, this antibody is recommended for detection of Lamin A/C of human, mouse, pig and origin by WB, IF, IHC(P), and Flow, Thermo Fisher Scientific Inc.)
Rheological analyses of ligand-free covalently integrin-linking hydrogel
Pellet of azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for different culture times (12, 24, and 48 h) were suspended with 100 μL of Alg-DBCO10 solution (2%, w/v) in HEPES buffer (100 mM, pH 7.4) and cells were completely dispersed by gentle pipetting. Rheological analyses of the reactants were performed on MCR 302 rheometer (Anton Paar) using a standard steel parallel-plate geometry of 25 mm in diameter. Oscillatory time and frequency were performed at 37 °C, and the storage modulus (G’) and loss modulus (G”) were recorded. The reactants were cast between the lower plate and upper plate. To prevent evaporation of water and better temperature control during testing, the plates were enclosed in a chamber. Time zero was set as the moment at which the reactants were cast on the plate. The time sweep data collection was started from time zero to 18,000 sec. The strain was maintained at 5% and operated at 10 rad/s. Values are average of three independent experiments and are expressed as mean ± SD.
In vitro cell viability in ligand-free covalently integrin-linking hydrogel
Pellet of azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for different culture times (12, 24, and 48 h) was suspended with 100 μL of Alg-DBCO10 solution (2%, w/v) in HEPES buffer and cells were completely dispersed by gentle pipetting. The cell dispersion was put on cell culture inserts and incubated at 37 °C for 30 min, allowing the formation of ligand-free covalently integrin-linking hydrogel. Two mL of SkMCM solution was added on the gels gently and cultured for 4 days at 37 °C in a humidified atmosphere containing 5.0% CO2. After predetermined period of time, calcein-AM (DOJINDO) and propidium iodide (PI, DOJINDO) were added in the medium and reacted for 1 h at 37 °C, then the gels were washed with PBS twice and CLSM observation was carried out in 1 mL of Live Cell Imaging Solution. Values are average of three separate experiments and are expressed as mean ± SD. As control sample, azide-modified HskMSCs encapsulating alginate gel, azide-modified HskMSCs (2.0 × 106) treated with Ac4ManNAz for 48 h were well suspended with 100 μL of Alg solution (2%, w/v) in HEPES buffer in the presence of CaCl2 solution (0.5%), was prepared. The cell viability of HskMSCs in the control gel was also examined with the same assay. Values are average of three independent experiments and are expressed as mean ± SD.
Nuclear morphology and nuclear localization of YAP in ligand-free covalently integrin-linking hydrogel
Pellet of azide-modified HskMSCs (2.0 × 106) treated with Ac4ManNAz for different culture times (12, 24, and 48 h) was suspended with 100 μL of Alg-DBCO10 solution (2%, w/v) in HEPES buffer and cells were completely dispersed by gentle pipetting. The cell dispersion was put on cell culture inserts and incubated at 37 °C for 30 min, allowing the formation of ligand-free covalently integrin-linking hydrogel. Two mL of SkMCM solution was added on the gels gently and cultured at 37 °C in a humidified atmosphere containing 5.0% CO2. As control sample, azide-modified HskMSCs encapsulating alginate gel, azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for 48 h were well suspended with 100 µL of Alg solution (2%, w/v) in HEPES buffer in the presence of CaCl2 solution (0.5%), was prepared. After 1 day of culture, the gels were removed from the cell culture insert and constant pressure (stress: 10%) was applied from the top of the gel for 10 min only using MCR 302 rheometer (Anton Paar). The treated gels were washed with PBS twice, then treated with 10% formalin solution at room temperature for 30 min. The gels were rinsed with PBS three times, and polyoxyethylene(10) octylphenyl ether (0.1%) was added and incubated at room temperature for 5 min. The gels were rinsed with PBS three times, and blocking buffer was added and incubated at room temperature for 30 min. The gels were reacted with primary antibody (Anti-YAP antibody (D8H1X) XP® Rabbit mAb #14074, Species Reactivity: human, mouse, rat, hamster, and monkey, Application: WB, IP, IHC, IF/ICC, ChIP, Cell Signaling Technology, Inc.) for overnight at 4 °C, then washed with PBS, followed by reacted with secondary antibody with Alexa Fluor® 488 for 1 h. The gels were washed with PBS, counterstained with DAPI, and CLSM observation was performed. The obtained images were evaluated for changes in the morphology of cell nuclei, Nuclear shape index (NSI), and the nuclear localization ratio of YAP. The evaluation methods previously reported for NSI were used. The nuclear localization rate of YAP is evaluated from the ratio of the fluorescence intensity of YAP in the cytoplasm to the fluorescence intensity of YAP in the cell nucleus. In addition, the same analyses were performed for the ligand-free covalently integrin-linking hydrogel prepared using HskMSCs treated with shRNA (LMNA and ITGB3).
The gels prepared using azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for 48 h were treated with cytochalasin D (0.5 µM) for 30 min, then the 3D networks were subjected to compressive stimulation using the same procedure described above and evaluated for changes in cell nuclei morphology. The gels prepared using azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for 48 h were treated with importazole (40 µM) for 1 h, then the gels were subjected to compressive stimulation using the same procedure described above and evaluated nuclear localization of YAP.
Pellet of azide-modified HskMSCs (2.0 × 106) treated with Ac4ManNAz for 48 h was suspended with 100 μL of Alg-RGD50 solution (2%, w/v) in HEPES buffer and cells were completely dispersed by gentle pipetting. The cell dispersion was put on cell culture inserts and incubated at 37 °C for 30 min, allowing the formation of ligand-free covalently integrin-linking hydrogel. Two mL of SkMCM solution was added on the gels gently and cultured at 37 °C in a humidified atmosphere containing 5.0% CO2. The gels were subjected to compressive stimulation using the same procedure described above and evaluated for changes in cell nuclei morphology and nuclear localization of YAP. Values are average of five independent experiments and are expressed as mean ± SD.
Nuclear localization of Dextran-FITC in ligand-free covalently integrin-linking hydrogel
HskMSC were seeded on 10 cm dish, and then 10 mL of SkMCM containing Ac4ManNAz (100 µM) was added to the dish and incubated at 37 °C for 48 h. The supernatant was removed and then 5 mL of serum-free SkMCM containing dextran (40 kDa)-FITC (2 mg/mL) was freshly added to the dish and incubated at 37 °C for 1 h. Pellet of azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for 48 h and dextran-FITC introduced was suspended with 100 µL of Alg-DBCO10 solution (2%, w/v) in HEPES buffer and cells were completely dispersed by gentle pipetting. The cell dispersion was put on cell culture inserts and incubated at 37 °C for 30 min, allowing the formation of ligand-free covalently integrin-linking hydrogel. Two mL of SkMCM solution was added on the gels gently and cultured at 37 °C in a humidified atmosphere containing 5.0% CO2. As control sample, azide-modified HskMSCs encapsulating alginate gel, azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for 48 h and dextran-FITC introduced were well suspended with 100 μL of Alg solution (2%, w/v) in HEPES buffer in the presence of CaCl2 solution (0.5%). After 30 min, the gel was removed from the cell culture insert and constant pressure (stress: 10%) was applied from the top of the gel for 10 min only using MCR 302 rheometer (Anton Paar). The treated gel was washed with PBS twice, then treated with 10% formalin solution at room temperature for 30 min. The gel was rinsed with PBS three times, and polyoxyethylene(10) octylphenyl ether (0.1%) was added and incubated at room temperature for 5 min. The gels were rinsed with PBS three times, and blocking buffer was added and incubated at room temperature for 30 min. The gels were washed with PBS, counterstained with DAPI at room temperature for 30 min, and CLSM observation was performed. The nuclear localization rate of dextran-FITC is evaluated from the ratio of the fluorescence intensity of FITC in the cytoplasm to the fluorescence intensity of dextran-FITC in the nucleus. In addition, the same analysis was performed for the gels prepared using azide-modified HskMSC (2.0 × 106) treated with Ac4ManNAz for 48 h and dextran (70 kDa)-FITC.
Muscle differentiation in ligand-free covalently integrin-linking hydrogel
Pellet of azide-modified HskMSCs (2.0 × 106) treated with Ac4ManNAz for different culture days (12, 24, and 48 h) was suspended with 100 μL of Alg-DBCO10 solution (2%, w/v) in HEPES buffer and cells were completely dispersed by gentle pipetting and incubated at 37 °C for 3 h. The cell dispersion was washed with PBS to remove unreacted Alg-DBCO10. SkMCM was added, and the gels were seeded into fibronectin-coated silicone chambers (4 cm2, NST-CH-10, Japan), incubated for 24 h to allow cell adhesion, and then the silicone chambers were set in a mechanical extension model (NST-1400, Japan) and subjected to mechanically stressed by 8% extension at a frequency of 10 cycles/min (Stretch group). This stretch stimulus mimicked the stretching dynamic movement of skeletal muscle tissue in vivo (10 cycles/min, 8% fractional shortening). As a control, the gels seeded on fibronectin-coated silicon chamber were cultured without stretching. After 5 days of continuous stretch stimulation, the gels was washed with PBS twice, then treated with 10% formalin solution at room temperature for 30 min. The gels were rinsed with PBS three times, and polyoxyethylene(10) octylphenyl ether (0.1%) was added and incubated at room temperature for 5 min. The gels were rinsed with PBS three times, and blocking buffer was added and incubated at room temperature for 30 min. The gels were reacted with primary antibody (Anti MYOD1 antibody 18943-1-AP, this antibody targets MYOD1 in WB, IHC, IF/ICC, ChIP, ELISA applications and shows reactivity with human, mouse, rat samples, Proteintech Group, Inc.) for overnight at 4 °C, then washed with PBS, followed by reacted with secondary antibody with Alexa Fluor® 488 for 1 h. The gels were washed with PBS, counterstained with DAPI, and CLSM observation was performed. In addition, the same analyses were performed for the gels prepared using HskMSCs treated with shRNA (LMNA and ITGB3). Values are average of ten independent experiments and are expressed as mean ± SD.
Quantitative real-time PCR
After 5 days of continuous stretch stimulation of the gels and the control samples, total RNA was isolated and purified using PureLink RNA Mini Kit (Invitrogen). RNA was quantified micro-spectrophotometer NanoDrop and 1 μg of RNA per treatment condition was added to 20 μL of reverse transcription reactions using the SuperScrip IV VIL Master Mix (Invitrogen). Thermal cycler conditions for reverse transcription were 25 °C for 10 min, 50 °C for 10 min, and 85 °C for 10 min and cDNA was stored at –20 °C prior to use in real time PCR. Two microliters of each reverse transcription reaction was used for 20 μL of real time PCR using the 96 well plate. Pre-validated primers and probes specific for GAPDH (assay ID Hs02786624_m1) and MYOD1 (assay ID Hs00159528_m1) were purchased from Applied Biosystems. Real time PCR were run in an Applied Biosystems StepOnePlus at thermal cycler conditions of 95 °C for 20 s, followed by 40 cycles at 95 °C for 1 s and 60 °C for 20 s. The 2-ΔΔCt method was employed in the qRT-PCR analysis. Values are average of three independent experiments and are expressed as mean ± SD.
Tissue regeneration capability of ligand-free covalently integrin-linking hydrogel
All mice used in the experiment were housed under the following conditions: light period: 8:00 A.M. to 8:00 P.M., dark period: 8:00 P.M. to 8:00 A.M., temperature: 25 °C, humidity: 70%. Nude mice (ICR-nu/nu, 5 weeks old, female and male) were anesthetized with isoflurane. The skin of the hind limb was opened, and the femoral muscle was gently exposed. In host femoral muscle, a local injury was created by removing 100 mg of skeletal muscle fibers. The skin was then carefully closed and the incision was sutured (4/0 silk). One day after the surgery, 200 μL of ligand-free covalently integrin-linking hydrogel precursor solution, pellet of azide-modified HskMSC cells (4.0 × 106) treated with Ac4ManNAz for different culture days (12, 24, and 48 h) was suspended with Alg-DBCO10 solution, was injected into the injury site using a syringe with a 26 G needle. In addition, control samples, azide-modified HskMSCs (4.0 × 106) treated with Ac4ManNAz for 48 h was suspended with 200 μL of Alg, Alg-RGD50 and Matrigel, were injected to the defect sites by the same procedures. After predetermined period of time, the muscle strength of the injured hind limbs was quantitatively measured using a grip-strength meter (MELQUEST, GMP-100B). Values are average of three independent experiments and are expressed as mean ± SD. At days 14 after the transplantation, the mice were sacrificed and the reconstructed skeletal muscle tissues were carefully excised, cryosections of the tissues were prepared (Thermo Scientific, CryoStar NX70). The cryosections were analyzed by CLSM. Moreover, the cryosections were stained with H&E and Masson’s trichrome staining, then analyzed by microscopy. The area (%) of muscle fibers, and collagen fibers in the reconstructed muscle tissues was quantified by Image J analysis using the corresponding microscopy images. Values are average of three independent experiments and are expressed as mean ± SD. Additionally, the cryosections were immunostained with Alexa Fluor® 546-labeled anti-MYH3 antibody (F1.652, Reactivity: human, mouse, rat, and rabbit, Application: WB, IHC, IF, IP, Santa Cruz Biotechnology, Inc.), then analyzed by CLSM.
Tensile test
Nude mice (ICR-nu/nu, 5 weeks old, female) were anesthetized with isoflurane. The skin of the hind limb was opened, and the femoral muscle was gently exposed. In host femoral muscle, a local injury was created by removing 100 mg of skeletal muscle fibers. The skin was then carefully closed and the incision was sutured (4/0 silk). One day after the surgery, 200 μL of ligand-free covalently integrin-linking hydrogel precursor solution, pellet of azide-modified HskMSC cells (4.0 × 106) treated with Ac4ManNAz for 48 h was suspended with Alg-DBCO10 solution, was injected into the injury site using a syringe with a 26 G needle. In addition, control sample, azide-modified HskMSCs (4.0 × 106) treated with Ac4ManNAz for 48 h was suspended with 200 μL of Alg, were injected to the defect sites by the same procedures. At days 14 after the transplantation, the mice were sacrificed, and the reconstructed skeletal muscle tissues were carefully excised. The reconstructed tissue was subjected to tensile test with samples clamped in the direction of the muscle fibers. Tensile tests were carried out on an EZ-LX (Simazu) with a load cell of 50 N using the strain rates of 5 mm/min.
Tissue regenerative capabilities of ligand-free covalently integrin-linking hydrogel for sciatic nerve injury mouse
Nude mice (ICR-nu/nu, 5 weeks old, female) were anesthetized with isoflurane. The skin of the hind limb was opened, and the femoral muscle was gently exposed. In the host thigh muscle, the sciatic nerve was torn 1 mm after creating a local injury by removing 100 mg of skeletal muscle fibers. The skin was then carefully closed and the incision was sutured (4/0 silk). One day after the surgery, 200 µL of the gel precursor solution, pellet of azide-modified HskMSC cells (4.0 × 106) treated with Ac4ManNAz for 48 h was suspended with Alg-DBCO10 solution, was injected into the injury site using a syringe with a 26 G needle. After predetermined period of time, the muscle strength of the injured hind limbs was quantitatively measured using a grip-strength meter (MELQUEST, GMP-100B). Values are average of three independent experiments and are expressed as mean ± SD. At days 14 after the transplantation, the mice were sacrificed and the reconstructed skeletal muscle tissues were carefully excised, cryosections of the tissues were prepared (Thermo Scientific, CryoStar NX70). The cryosections were analyzed by Masson’s trichrome staining, then analyzed by microscopy.
Biochemical test of ligand-free covalently integrin-linking hydrogel
Nude mice (ICR-nu/nu, 5 weeks old, female) were anesthetized with isoflurane. The skin of the hind limb was opened, and the femoral muscle was gently exposed. In host femoral muscle, a local injury was created by removing 100 mg of skeletal muscle fibers. The skin was then carefully closed and the incision was sutured (4/0 silk). One day after the surgery, 200 μL of gel precursor solution, pellet of azide-modified HskMSCs (4.0 × 106) treated with Ac4ManNAz for 48 h was suspended with Alg-DBCO10 solution, was injected into the injury site using a syringe with a 26 G needle. At days 14 after the transplantation, a simultaneous skin and abdominal incision was made from the top of the external urethral opening to the xiphoid process. The skin and abdominal muscles were then cut along the auxiliary bone on both sides. A 24 G syringe was inserted parallel to the abdominal vena cava, and the inner tube was pulled to collect blood. The blood was mixed by gently inverting 5 times, after confirming coagulation, the blood was centrifuged at 1700 × g for 15 min at room temperature. Only the supernatant was collected and biochemical tests were commissioned (Oriental Yeast India Pvt. Ltd.).
Statistical analysis
All statistical data are presented as mean ± standard deviation (SD). Two-sided Student’s t test was used to analyze the significance, and *P < 0.05, **P < 0.01, ***P < 0.001. Source data is provided as a Source Data file.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.