Introduction
Clinopodium nepeta (L.) Kuntze is a robust aromatic plant belonging to the Lamiaceae (Labiatae) family. It is widely found in the Mediterranean region and has been commonly utilized for its stimulant, sweat-inducing, antiseptic, mucolytic, antispasmodic, diuretic, carminative, and tonic features1,2,3. Clinopodium species, which are abundant in bioactive substances, are reported to exhibit various biological activities4,5,6. Previously, phytochemicals such as terpenes, saponins, glycosides, flavonoids, alkaloids, sterols and tannins were identified in C. nepeta7,8. Numerous reports can be found in the literature investigating the essential oil content of C. nepeta7,8,9,10,11,12,13. These studies identified the primary constituents of C. nepeta essential oil as pulegone, menthone, isomenthone, piperitone, piperitenone, neo-menthol, trans-menthone, 1,8-cineole, pinocampene, cis-isopulegone, menthol, thymol, piperitenone oxide, limonene, and trans-piperitone, varying according to the plant source. Contrary to its essential oil, studies on the phenolic profile of its extracts are limited3,8,14,15. The research findings indicated that C. nepeta is abundant in various phenolic compounds, such as caffeine, gallic acid, and the phenolic acids rosmarinic, caffeic, and vanillic acids, along with syringic acid, p-coumaric acid, ferulic acid, and benzoic acid. In these studies, in which the subspecies of C. nepeta was not specified, gallic acid, rosmarinic acid, caffeine, caffeic acid, vanillic acid, syringic acid, p-coumaric acid, ferulic acid and benzoic acid were determined as the main phenolic components. No research was discovered concerning the phenolic content of the other subspecies (Clinopodium nepeta subsp. spruneri, Syn: C. nepeta subsp. glandulosum) that we assessed as part of this study.
Many medicinal plants are rich in phenolic compounds with distinct and powerful biological activities. These compounds are involved in defense mechanisms against UV, drought, heat, pathogens, predators, interspecies competition, and pollination in plants. In medical terms, these compounds perform a crucial function in the removal of free radicals that cause cell damage and thus show the ability to prevent the formation of cancer, cardiovascular and neurodegenerative diseases16,17. In particular, abiotic factors are known to significantly affect plant development and the synthesis of secondary metabolites. Based on this information, investigations have been executed to analyze the effects of stressors such as temperature, light, UV, drought and many other factors on the growth of the plant and the production of secondary metabolites18.
Lately, plant tissue culture processes have been evaluated as an effective tool for the production of medicinal plants and their bioactive phytochemicals19. In this way, the destruction of natural habitats can be prevented, plants that are superior in terms of content and yield can be created, and continuity can be ensured in the manufacturing of valuable plant secondary products regardless of time and seasonal conditions. Exposing the organs, tissues or cells of the plant to stressors is the leading method used to improve the manufacturing of valuable phytochemicals in plants. Research has demonstrated that secondary metabolites are produced as defensive responses of plants to such stressors20. Recently, studies on the use of nanoparticles as elicitors that affect diverse morphological, physiological and biochemical mechanisms in plant metabolism have become widespread, especially in vitro, and it has been reported that they play a considerable role in the biosynthesis of valuable phytochemicals21,22,23,24. Among these, silver nanoparticles (AgNPs) are one of the most widely used due to their potential to influence growth, biomass, and accumulation of phytochemicals21,22,25,26. For instance, it was reported that 10 mg/L AgNP application decreased shoot elongation in Linum usitatissimum and Hordeum vulgare27, while studies with Brassica juncea28 and Phaseolus vulgaris and Zea mays29 showed positive effects of AgNP application on plant growth. Tymoszuk and Miler30 found that a 10 mg/L concentration of AgNP applied to Streptocarpus × hybridus in vitro enhanced micropropagation efficiency. In P. vulgaris, Z. mays and Lycopersicon esculentum species subjected to AgNPs at different concentrations, considerable increases in biomass, reductions in oxidative stress, enhancements in flavonoid content, and notably high total phenolic contents were observed31. Although there are limited records on the micropropagation of C. nepeta32 and the production of its secondary metabolites, to the best of our knowledge, there is no available information on the potential influence of nanoparticles on the micropropagation of this species and the production of its bioactive phytochemicals with antioxidant and enzyme inhibitory activities.
Therefore, the present study was designed to clarify whether silver nanoparticle (AgNP) treatments can stimulate the enhanced accumulation of valuable secondary metabolites in C. nepeta subsp. spruneri under in vitro conditions. Specifically, we aimed to determine the concentration-dependent effects of AgNPs (1.0, 5.0, 10.0, 20.0, and 50.0 mg/L) on shoot growth, phenolic content, essential oil composition, antioxidant activity, and acetylcholinesterase (AChE) inhibition. The in vitro system was selected to provide a controlled environment, allowing precise evaluation of AgNP-mediated metabolic responses while also offering a sustainable platform for potential commercial exploitation of bioactive phytochemicals.
Results and discussion
Effects of AgNP applications on growth parameters
Table 1 summarizes the effects of silver nanoparticles (AgNPs) on the in vitro development of C. nepeta subsp. spruneri. The results clearly demonstrate that all measured growth parameters-including shoot elongation, node number, leaf number, and biomass-were progressively reduced with increasing AgNP concentration, while root formation was completely inhibited at all tested doses. Although the control group exhibited the highest values for shoot and root development, a concentration-dependent decline was evident across all AgNP-treated groups, with the strongest inhibition recorded at 50.0 mg/L. Interestingly, only the number of shoots per explant showed a relative increase under AgNP exposure, suggesting a stress-induced stimulation of shoot proliferation. Overall, these findings indicate the dose-dependent inhibitory effects of AgNPs on plant growth and development.
Our results are consistent with several previous studies reporting growth inhibition under high AgNP concentrations. For example, Jiang et al.33 observed that Spirodela polyrrhiza exposed to AgNPs exhibited reduced biomass, restricted shoot growth, and root abscission, while Kaveh et al.34 similarly noted biomass reduction in Arabidopsis at 5–20 mg/L AgNP. Dimkpa et al.35 also documented concentration-dependent inhibition of shoot and root elongation in wheat, and comparable negative effects on rice growth were described by other authors36. These reports parallel our findings, confirming that AgNPs often impair root formation and limit shoot development, particularly at higher concentrations.
On the other hand, some studies have indicated that low AgNP levels may stimulate growth. For instance, in Oryza sativa, root growth was enhanced at 30 mg/L but inhibited at 60 µg/mL37, while in Capsicum annuum, 1 mg/L AgNP reduced shoot elongation and biomass38. Similar stimulatory effects at low doses have been reported in maize, where 20–60 µg/L AgNP promoted growth, although the effective range varied depending on plant species29. Together with our results, these contrasting reports suggest that the physiological responses to AgNPs are highly dependent on plant type, nanoparticle concentration, size, and exposure duration.
Total phenolic content and total flavonoid content of extracts
In Table 2, the total phenolic content (TPC) and total flavonoid content (TFC) values of C. nepeta subsp. spruneri shoots grown at different AgNP concentrations are presented as mg GAE/g extract and mg QE/g extract, respectively. The corresponding values for the in vitro control and the wild plant are also provided. A remarkable increase was observed in TPC and TFC values in all AgNP applications compared to the control group. The wild plant exhibited higher TPC and TFC values than the control, reflecting the natural accumulation of these metabolites under diverse environmental stress conditions.
The TPC values for AgNP treatments increased significantly from 82.37 to 137.88 mg GAE/g extract with increasing AgNP concentration from 1.0 to 20.0 mg/L, while TFC values remained relatively stable. Notably, the TPC value at 20.0 mg/L was higher than that of the wild plant. At lower concentrations (e.g., 1.0 mg/L), AgNP appeared to enhance TPC while reducing TFC, whereas higher concentrations showed a more variable effect. Despite these fluctuations, the maximum TPC (137.88 mg GAE/g extract) and TFC (10.81 mg QE/g extract) values were both recorded at 20.0 mg/L AgNP.
Plants are known to increase secondary metabolite production in response to stress. Elicitors added to culture media can activate defense mechanisms and stimulate the accumulation of phenolics and flavonoids39,40. Nanomaterials have also been shown to act as abiotic elicitors, although their effects are strongly dependent on plant species, nanoparticle type, and applied concentration41,42,43,44. Our findings are consistent with previous studies reporting that AgNP treatments enhanced TPC and TFC in plant extracts22,45. However, unlike those reports, we observed that TFC did not increase at higher AgNP concentrations, indicating a dose- and species-dependent response.
The comparison with the wild plant group provides additional insight. The higher TPC and TFC values in wild plants compared to in vitro controls likely result from continuous exposure to environmental stresses such as temperature fluctuations, UV radiation, and microbial interactions, which stimulate phenolic metabolism. By contrast, the in vitro control plants grow under stable, stress-free conditions and therefore accumulate lower levels of these compounds. Interestingly, AgNP treatment was able to elevate phenolic and flavonoid levels in vitro seedlings to values comparable to, or even exceeding, those of the wild plant, particularly at 20.0 mg/L.
The increased accumulation of phenolics and flavonoids in AgNP-treated seedlings may thus reflect the activation of stress-related pathways, including the phenylpropanoid pathway, which plays a central role in the biosynthesis of these compounds42. This adaptive metabolic adjustment highlights the potential of AgNPs as abiotic elicitors to enhance phytochemical production under controlled in vitro conditions.
AgNP treatments enhanced rosmarinic acid levels
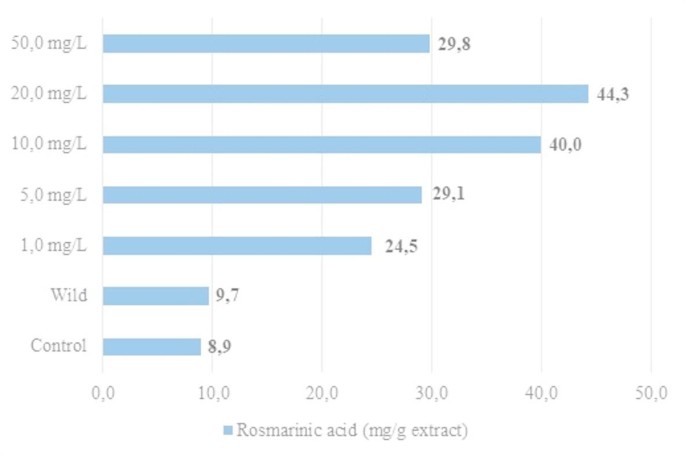
Rosmarinic acid (RA) is a valuable phenolic compound commonly found in the Lamiaceae family and has been reported as the predominant phenolic responsible for bioactivity in various Clinopodium species3,46,47,48. In this study, RA levels in methanolic extracts of C. nepeta subsp. spruneri seedlings treated with different concentrations of AgNP, the control group, and wild plants were determined and are presented in Fig. 1. RA levels were consistently higher in AgNP-treated seedlings compared to both the control group and the wild plants. The maximum RA content (44.3 mg/g extract) was observed in seedlings exposed to 20.0 mg/L AgNP, while the lowest RA concentrations were found in the control group (8.9 mg/g) and wild plants (9.7 mg/g). RA accumulation increased with AgNP concentration up to 20 mg/L, but declined at higher concentrations, with a marked decrease at 50 mg/L.

The content of rosmarinic acid in the AgNP-treated seedlings, the control group, and the wild plant of C. nepeta subsp. spruneri.
Interestingly, wild plants exhibited much higher TPC and TFC values than in vitro controls, but their RA levels remained low and comparable to controls. This discrepancy may be attributed to differences in metabolic regulation under natural versus in vitro conditions. Wild plants are continuously exposed to a range of abiotic stresses (UV radiation, temperature fluctuations, soil microorganisms, etc.), which can strongly stimulate the accumulation of general phenolic and flavonoid compounds as broad-spectrum defense metabolites17,49. However, RA biosynthesis is regulated through a more specific branch of the phenylpropanoid pathway, which may not be equally induced under natural conditions. In contrast, AgNP application appears to act as a direct elicitor of RA biosynthesis, particularly by stimulating key enzymes such as phenylalanine ammonia-lyase (PAL), tyrosine aminotransferase (TAT), and rosmarinic acid synthase (RAS)47,48,50. This could explain why AgNP-treated seedlings accumulated substantially higher RA levels compared to both controls and wild plants, despite the latter having higher TPC and TFC contents.
Beyond their quantitative increase, the biological significance of these metabolites should also be emphasized. Rosmarinic acid is widely recognized for its anti-inflammatory, antioxidant, antidiabetic, antiviral, antitumor, neuroprotective, and hepatoprotective properties49,50. Likewise, isoeugenol has been reported as a bioactive phenylpropanoid with strong antioxidant and antimicrobial activities, along with potential antitumor and neuroprotective effects51. Thus, the enhanced RA accumulation and the induction of isoeugenol under AgNP elicitation not only highlight the unique metabolic responses of C. nepeta subsp. spruneri but also underline the pharmaceutical relevance of these findings.
The decline in RA levels at 50 mg/L AgNP may reflect a threshold beyond which nanoparticles exert phytotoxic effects rather than elicitor activity. At such high concentrations, AgNPs can cause excessive oxidative stress, impair enzymatic activities, and disrupt metabolic balance28,29,43. Consequently, RA biosynthesis may be downregulated or inhibited, leading to reduced accumulation. Similar inhibitory effects of high nanoparticle doses on phenolic compounds have been reported in other medicinal plants, supporting the dose-dependent nature of this response.
Taken together, these findings indicate that AgNP treatments act as a strong elicitor of RA biosynthesis in C. nepeta subsp. spruneri seedlings, with the most pronounced effect at 20 mg/L, while excessive concentrations may cause metabolic inhibition rather than stimulation.
Antioxidant activity increased under AgNP treatments
The antioxidant activity of the methanolic extracts was assessed by DPPH and FRAP assays (Table 3). Both assays consistently demonstrated that the seedlings exposed to 20 mg/L AgNP exhibited the strongest antioxidant capacity, whereas the control group showed the weakest activity. In the DPPH assay, the SC50 value of 26.82 µg/mL in the 20 mg/L group reflected a markedly higher radical scavenging effect compared to the control (105.66 µg/mL). A similar trend was observed in FRAP analysis, where the reducing power of the 20 mg/L group clearly exceeded that of the control seedlings. Interestingly, extracts of wild plants displayed antioxidant activity comparable to or slightly higher than some AgNP treatments, but still lower than the peak values obtained at 20 mg/L. This finding suggests that natural environmental stressors may induce the synthesis of antioxidant metabolites in wild plants, but controlled elicitation with AgNPs at optimal concentrations can further amplify this response under in vitro conditions.
These results are consistent with previous reports showing that nanoparticles at appropriate concentrations can enhance antioxidant activity by stimulating the accumulation of phenolic and flavonoid compounds52. Several studies have further confirmed that nanoparticles act as abiotic elicitors influencing plant physiology and secondary metabolite accumulation53,54. For instance, ZnO-NPs applied to Stevia rebaudiana callus cultures enhanced antioxidant capacity and metabolite production55. However, at higher doses (50 mg/L in our study), a decline in activity was recorded, likely due to phytotoxic effects of excessive nanoparticle exposure, which impair normal metabolism rather than promote it. Taken together, these findings indicate that AgNPs can serve as effective elicitors of antioxidant responses in C. nepeta subsp. spruneri seedlings in a concentration-dependent manner, with 20 mg/L identified as the most effective treatment.
AChE inhibition in wild and AgNP-treated seedlings
Table 3 shows the IC50 values of methanolic extracts from control, AgNP-treated seedlings, and wild plants, compared with donepezil as a positive control. Among the tested groups, the lowest IC50 (3.37 mg/mL) was obtained from the wild plant, indicating the strongest acetylcholinesterase (AChE) inhibition. This was closely followed by the 20 mg/L AgNP treatment (3.46 mg/mL). By contrast, the control group exhibited the weakest inhibition (IC50: 13.26 mg/mL). Although all extracts displayed lower potency than donepezil (IC50: 1.25 mg/mL), both the 20 mg/L AgNP and wild plant extracts showed notable activity, highlighting the potential of AgNP treatment as a viable alternative to wild harvesting.
These observations are consistent with the trends seen for total phenolic content (TPC), rosmarinic acid (RA) accumulation, and antioxidant capacity, where higher phenolic levels generally coincided with stronger AChE inhibition. This supports previous findings that phenolics, in addition to their antioxidant roles, contribute to cholinesterase inhibition and may have relevance in Alzheimer’s therapy56. However, the superior inhibition recorded in wild plants, despite their lower RA levels compared to treated seedlings, suggests the involvement of other bioactive metabolites such as alkaloids, terpenoids, or essential oil constituents. This interpretation is supported by Costa et al.49, who reported that although Lavandula viridis micropropagated plants accumulated more phenolics and exhibited higher antioxidant activity, wild specimens displayed stronger anticholinesterase effects.
Similarly, our results show that while AgNP-treated seedlings accumulated higher phenolic contents and antioxidant activity than in vitro controls, their AChE inhibition remained slightly below that of wild plants. This indicates that AgNP elicitation enhances cholinesterase inhibitory activity, but not to the full extent achieved by natural environmental stressors acting on wild plants. Nonetheless, the substantial activity observed in the 20 mg/L AgNP group underscores the value of nanoparticle elicitation for generating bioactive plant material in vitro, offering a sustainable strategy that reduces dependence on wild populations.
Essential oil composition of seedlings
Figure 2 presents the percentages of the five main components identified by GC-MS analysis of seedlings treated with various AgNP concentrations. Other identified compounds are listed in the supplemental materials (SF1). The dominant essential oil components across different groups included isoeugenol, menthalactone, limonene, terpinen-4-ol, bornyl acetate, p-cymen-8-ol, benzyl alcohol, isopulegol, octadecane, β-thujone, safranal, α-himachalene, trans-sabinene hydrate, lauryl alcohol, and carvacrol. The compositions varied depending on AgNP concentration and growth conditions (in vitro vs. wild). Isoeugenol was predominant in AgNP-treated seedlings, whereas menthalactone was dominant in both control and wild samples. Isoeugenol was detected at 34.47%, 36.57%, 34.38%, and 28.25% in seedlings treated with 1.0, 5.0, 10.0, and 20.0 mg/L AgNP, respectively. Menthalactone was highest in the control group (64.44%) and also dominant in wild plants (48.29%). Increasing AgNP concentration led to a decrease in isoeugenol and a partial increase in menthalactone, with the highest menthalactone level observed at 50.0 mg/L. Additionally, limonene and terpinen-4-ol were found at notable levels in some treatments. In wild plants, isoeugenol was absent, and lauryl alcohol was the second most abundant component (15.40%).

Five main components of the essential oil in the AgNP-treated seedlings, the control group and the wild plant.
Previous studies on C. nepeta essential oils were generally performed with wild plants collected from natural habitats rather than in vitro-cultured material13,57,58,59. For example, Öztürk et al.13 reported piperitone oxide (47.8%), limonene (18.6%), and piperitone oxide II (13.6%) as the major compounds in C. nepeta subsp. glandulosum. Similarly, Debbabi et al.57 found piperitone oxide and piperitenone oxide as dominant constituents. Other reports also identified pulegone, piperitenone, isomenthone, and 1,8-cineole as major components58,59. In contrast, our study revealed menthalactone and isoeugenol as predominant constituents under in vitro conditions with AgNP treatments.
To the best of our knowledge, no previous study has reported the effects of AgNP treatments on the essential oil composition of Clinopodium species. Therefore, direct comparisons with similar works are not available. The observed shift from menthalactone to isoeugenol dominance in AgNP-treated seedlings may be interpreted as a stress-induced reorientation of secondary metabolism, possibly favoring the phenylpropanoid pathway over terpenoid biosynthesis. This explanation is provided as our own interpretation, based on established knowledge of plant secondary metabolism, rather than on specific prior studies.
The higher proportion of lauryl alcohol in wild plants compared to in vitro controls could be attributed to environmental stress factors such as temperature fluctuations, UV radiation, and biotic interactions, which are absent under controlled culture conditions. Similar shifts in essential oil composition between wild and in vitro plants have been reported in other Lamiaceae members, where wild plants accumulate higher levels of alcohols and terpenoids as adaptive responses60.
Overall, these findings highlight the strong influence of growth conditions and AgNP elicitation on essential oil profiles. Notably, menthalactone appears to be reported here for the first time as a major constituent in Clinopodium essential oils, while isoeugenol-previously detected only in low amounts in Satureja odora (syn: Clinopodium odorum)51 -is, to our knowledge, documented for the first time as a predominant compound in C. nepeta subsp. spruneri seedlings grown in vitro under AgNP treatments.
Conclusion
In conclusion, the present study demonstrated that silver nanoparticle (AgNP) treatments influenced growth, phenolic accumulation, antioxidant activity, enzyme inhibition, and essential oil composition of C. nepeta subsp. spruneri seedlings under in vitro conditions. The results suggest that moderate concentrations of AgNPs (particularly 20 mg/L) may stimulate the production of bioactive compounds such as rosmarinic acid and isoeugenol, whereas higher concentrations can exert phytotoxic effects. These findings highlight the potential of AgNP elicitation for enhancing secondary metabolite production in medicinal and aromatic plants. However, some limitations should be acknowledged: silver accumulation in plant tissues was not directly quantified, and all experiments were conducted under in vitro culture conditions, which may not fully represent natural or field environments. Future studies incorporating elemental analyses and alternative cultivation systems will be essential to validate and extend these insights.
Materials and methods
Collection and germination of seeds
For germination experiments, mature seeds of Clinopodium nepeta subsp. spruneri were obtained from adult plants, population at 40°59’35.8″N 39°41’35.2″E in Trabzon, Turkey. The seeds were allowed to dry at ambient temperature for a week and theerafter stored at + 4 °C. Afterwards, the surface sterilization was carried out in accordance with the study submitted in the literature61. In short, mature seeds were kept in sucrose solution (5%, with 1–2 drops of commercial bleach) for 12 h and then the seeds taken from this solution were treated with H2O2 for 30 m. Seeds were washed with sterile dH2O (3 times, 5 min each) and transferred to Murashige&Skoog (MS) including vitamins medium62 containing 20 g/L sucrose and 7 g/L plant agar. Incubation conditions for all cultures were designed at 25 °C, 16 h of light under cold white fluorescent light (50 µmol m−2 s−1) and 8 h of darkness. Murashige&Skoog including vitamins (M0222), sucrose (S0809) and phyto agar (P1003) were purchased from Duchefa Biochemie (Netherlands).
Wild plants (before flowering stage) were obtained from the abovementioned population (coordinates were given above) for comparison with shoots obtained from different in vitro conditions. The plant material, Clinopodium nepeta subsp. spruneri, was identified by Prof. Dr. Mutlu GULTEPE (Giresun University). A voucher specimen (ESP55) has been deposited in the Herbarium of Espiye Vocational School, Giresun University, Turkey. According to national regulations, no specific permissions or licenses are required for the collection of Clinopodium nepeta subsp. spruneri. All activities complied with the general principles of institutional, national, and international guidelines for plant research. Collected plant materials were shade-dried, then mechanically grounded into the powder and stored in closed containers at + 4 °C to be used in related studies.
AgNP elicitation
After germination, the seedlings were incubated for a total of 60 days under the same culture conditions (MS medium supplemented with 30 g/L sucrose and 7 g/L plant agar, pH 5.8, sterilized at 121 °C for 15 min), with subculturing every 30 days (2 times in total). Nodal explants (3–4 explants from each seedling), approximately 1 cm in length and containing a single node, were then excised. These explants were further subcultured by transferring them onto the same MS medium to obtain a sufficient number of explants. Cultures were maintained at 25 °C under a 16/8 h light/dark photoperiod.
To establish the impact of AgNPs, concentrations of 1.0, 5.0, 10.0, 20.0, and 50.0 mg/L were selected based on the study by Jadczak et al.63 and added to the MS medium. The silver nanoparticles (AgNPs) used in this study were purchased from Nanografi Nanotechnology (Ankara, Türkiye; Catalog No: NG04EO0102). According to the manufacturer’s specifications, the AgNPs exhibited a high purity of 99.995% and consisted of spherical particles with an average size ranging from 45 to 75 nm, stabilized with approximately 0.25% polyvinylpyrroline (PVP). These particles were black in color, had a cubic crystal structure, and a true density of 10.6 g/cm³. Additional properties include a melting point of 961.9 °C and a boiling point of 2212 °C. Trace elemental analysis indicated that the levels of Fe, Bi, Pb, and Sb were all below 0.003%.
A stock solution of AgNPs (10 mg/mL), prepared in deionized water, was sterilized by autoclaving and added to the autoclaved nutrient media in appropriate amounts before solidification. The AgNP-free medium served as the control group. All media were supplemented with 30 g/L sucrose and 7 g/L plant agar and were sterilized in an autoclave at 121 °C for 15 m. Nodal segments obtained from previously subcultured seedlings were inoculated onto the AgNP-containing media and incubated at 25 °C under a 16-h light (cool white fluorescent lamps, 50 µmol m⁻² s⁻¹)/8-h dark photoperiod for 30 d. For each treatment, 25 magenta boxes, each containing 5 explants, were used.
Extraction of plant materials
After harvesting, the in vitro-grown seedlings were dried in an incubator at 40 °C and ground into fine powder. The extraction of the powdered plant samples was performed using 99% methanol with a maceration procedure by shaking on an orbital shaker for 24 h at 25 °C. The extracts obtained after this process were first pre-filtered using filter paper, followed by final filtration through a 0.45 μm membrane. The supernatants were collected, and part of the methanolic extracts was evaporated to determine extraction yields. The remaining extracts were stored at 4 °C in closed containers until use.
These methanolic extracts were employed for all subsequent phytochemical and bioactivity analyses, including total phenolic and flavonoid content (TPC and TFC), HPLC profiling of rosmarinic acid, antioxidant activity (DPPH and FRAP assays), and enzyme inhibition tests (acetylcholinesterase, etc.). For comparison, wild plant materials collected from natural habitats were subjected to the same drying, grinding, and extraction procedure, and their methanolic extracts were analyzed in parallel with the in vitro samples.
Analysis of total phenolic (TPC) and total flavonoid (TFC) contents
The methanolic extracts prepared as described in Sect. “Extraction of plant materials” were used for TPC and TFC analyses. These extracts were obtained from in vitro-grown C. nepeta seedlings cultured on MS medium supplemented with different concentrations of silver nanoparticles (AgNPs: 0, 1.0, 5.0, 10.0, 20.0, and 50.0 mg/L), as well as from wild plants. The wild materials originated from adult plants collected from the natural population previously described in Sect. “Collection and germination of seeds”(40°59’35.8″N 39°41’35.2″E, Trabzon, Turkey). They were subjected to the same drying, grinding, and methanol extraction procedure as described for the in vitro samples, ensuring methodological consistency for direct comparison.
The total phenolic content (TPC) of the methanolic extracts was determined using the Folin–Ciocalteu assay, with gallic acid as the standard64. Results are expressed as mg gallic acid equivalents (GAE) per g extract. The total flavonoid content (TFC) was measured using the aluminum chloride colorimetric method65, with quercetin as the reference standard (concentration range: 1–0.03125 mg/mL). The calibration curve was constructed from the absorbance values corresponding to each concentration, and the flavonoid content of the samples was calculated from this standard curve.
HPLC analysis of rosmarinic acid
The rosmarinic acid (RA) content in the micropropagated shoots and wild plant was quantified utilizing RP-HPLC-DAD. These measurements were performed on a Thermo Scientific Dionex Ultimate™ 3000 system (Thermo Scientific, Bremen, Germany). Chromatographic decomposition was enforced using a Thermo Scientific™ Hypersil™ ODS C18 HPLC column (250 mm × 4.6 mm × 5 μm) (Thermo Scientific, USA) at a temperature of 30 °C. The mobile phase consisted of 2% (v/v) acetic acid in water (A) and 70% (v/v) acetonitrile in water (B), operating at a flow rate of 1.2 mL/min using gradient elution. The gradient began with 5% B at time zero and rised to 60% B over 26 m. Detection of RA was carried out at 315 nm66.
Antioxidant activity
DPPH assay
The DPPH radical scavenging efficiency was evaluated using the DPPH (2,2-diphenyl-1-picrylhydrazyl) method described by Molyneux67. Briefly, 750 µL of the methanolic extract was mixed with an equal volume of 0.1 M methanolic DPPH solution. After incubation for 50 m at room temperature in the dark, the absorbance was measured at 517 nm. The radical scavenging capacity of the extract was determined by plotting the absorbance values against different extract concentrations, and the SC₅₀ (half maximal scavenging concentration) value was calculated from the resulting dose-response curve. The SC₅₀ value represents the concentration of the extract required to reduce the initial DPPH• concentration by 50%. A lower SC₅₀ value corresponds to higher antioxidant (radical scavenging) activity. All experiments were carried out in three independent replicates.
FRAP assay
The FRAP impact of methanolic extracts were established in accordance with the process submitted previously68. This process is employed for the definition of cumulative antioxidant action. FRAP reagent was prepared by mixing newly prepared TPTZ (0.01 M), FeCl3.6H2O (0.02 M) and acetate buffer (0.3 M, pH: 3.6). The process was implemented by the incubation of 3 mL of reagent (FRAP) with 100 µL of extract at 25 °C for 4 m. Subsequently, the absorbance was recorded at a wavelength of 593 nm. The blanks (reagent and extracts) were recorded and removed from the final absorbance values. Trolox® served as a positive control to establish a reference graph, and the results were reported as µM Trolox equivalents per gram of extract. All tests were performed in three independent replicates.
Acetylcholinesterase inhibitory activity
The in vitro effects of the extracts on acetylcholinesterase were assessed using Ellman’s spectrophotometric method69, with some alterations. To prepare aliquots, acetylcholinesterase was dispersed in a 1% gelatin solution. The aliquots were then adjusted to a concentration of 0.2 U/mL in a phosphate buffer (pH: 7.0, 0.05 M). In test tubes, 0.1 mL of the extract, 1200 µL of phosphate buffer (pH: 6.8, 100 µM), and 100 µL of the enzyme solution (0.2 U/mL) were mixed and set at ambient temperature for 30 m. Thereafter, 50 µL of 5,5’-Dithiobis(2-nitrobenzoic acid) (3 mM, equipped in 50 mM phosphate buffer, pH 7.0) and 50 µL of acetylthiocholine iodide (3 mM, also equipped in 50 mM phosphate buffer, pH 7.0) were joined, and the mixture was incubated for an additional 15 m. Spectrophotometric measurements of absorbance were recorded at a wavelength of 412 nm. Percentage of inhibition versus concentration graphs were plotted based on the evaulated absorbances, and IC50 values were derived from these graphs.
GC-MS analysis of essential oils
Dried and powdered plant materials were used to extract essential oils in the experimental groups, and the headspace solid phase micro-extraction (spme) technique was employed for this purpose. Volatile oils were gathered using a specific fiber assembly, namely the Stable Flex SPME fiber assembly Divinylbenzene/Carboxene/Polydimethylsiloxane (Supelco, 57348-U), which was heated to 150 °C for 20 m. The essential oil composition was subsequently examined using GC-MS, with specific settings for temperature, flow rate, division ratio, ionization energy, and mass/charge range, on a Shimadzu QP-2010 Ultra system equipped with an Rxi-5MS column (30 m x 0.25 mm x 0.25 μm). The GC-MS system was operated with a temperature gradient starting from 40 °C and step by step increasing to 240 °C at a rate of 4 °C per m, followed by a 2-m hold at 240 °C. The injector and detector temperatures were maintained at 250 °C, with helium utilized as the carrier gas (flow rate: 1.44 mL per m, division ratio: 1/25, ionization energy: 70 eV). The detection system of the mass spectrometer operated in scanning process, covering a mass-to-charge ratio range of 40–450. The components of the volatile oil were defined by comparison their mass spectra against those available in the Wiley FFNSC libraries.
Statistical analysis
All trials were executed employing a fully randomized design, and the significance of the datas was assessed using one-way analysis of variance. Subsequently, the treatment means were compared employing Tukey’s t-test, with statistical significance defined at P ≤ 0.05. Additionally, to ensure the reliability and reproducibility of the findings, the trials were performed in three independent replicates and consistent outcomes were obtained.
Data availability
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
-
Couladis, M. & Tzakou, O. Calamintha Nepeta subsp. Glandulosa from Greece. J Essent. Oil Res. 13, 11–12 (2011).
-
Negro, C., Notarnicola, S., De Bellis, L. & Miceli, A. Intraspecific variability of the essential oil of calamintha Nepeta subsp. Nepeta from Southern Italy (Apulia). Nat. Prod. Res. 27, 331–339 (2013).
-
Pacifico, S. et al. Seasonal variation in phenolic composition and antioxidant and anti-inflammatory activities of calamintha Nepeta (L.) Savi. Food Res. Int. 69, 121–132 (2015).
-
Vera, S. M. B., Puente, N., Malafronte, B., Braca, A. & N. & Phenolic compounds from clinopodium tomentosum (Kunth) Govaerts (Lamiaceae). J. Braz Chem. Soc. 25, 2121–2124 (2014).
-
Dunkić, V. et al. Micromorphological and phytochemical traits of four clinopodium L. species (Lamiaceae). S Afr. J. Bot. 111, 232–241 (2017).
-
Moattar, F. S., Sariri, R., Giahi, M. & Yaghmaee, P. Essential oil composition and antioxidant activity of calamintha officinalis Moench. J. Appl. Biotechnol. Rep. 5, 55–58 (2018).
-
Boudjema, K. et al. Phytochemical profile and antimicrobial properties of volatile compounds of satureja calamintha (L.) scheel from Northern Algeria. Trop. J. Pharm. Res. 17, 857–864 (2018).
-
Khodja, N. K. et al. Chemical composition and antioxidant activity of phenolic compounds and essential oils from Calamintha nepeta L. J. Complement. Integr. Med. 15(4), (2018). https://doi.org/10.1515/jcim-2017-0080 PMID: 29794251.
-
Marongiu, B. et al. Chemical composition and biological assays of essential oils of calamintha Nepeta (L.) Savi subsp. Nepeta (Lamiaceae). Nat. Prod. Res. 24, 1734–1742 (2010).
-
Alan, S., Kürkçüoglu, M. & Baser, K. H. C. Composition of essential oils of calamintha Nepeta (L.) Savi subsp. Nepeta and calamintha Nepeta (L.) Savi subsp. Glandulosa (Req.) PW ball. Asian J. Chem. 23, 2357–2360 (2011).
-
Božović, M. & Ragno, R. Calamintha Nepeta (L.) Savi and its main essential oil constituent pulegone: biological activities and chemistry. Molecules 22, 290 (2017).
-
Ambrico, A., Trupo, M., Martino, M. & Sharma, N. Essential oil of calamintha Nepeta (L.) Savi subsp. Nepeta is a potential control agent for some postharvest fruit diseases. Org. Agr. 10, 35–48 (2020).
-
Öztürk, G., Yılmaz, G., Ekşi, G. & Demirci, B. Chemical composition and antibacterial activity of clinopodium Nepeta subsp. Glandulosum (Req.) Govaerts essential oil. Nat. Volatiles Essent. Oils. 8, 75–80 (2021).
-
Çelik, G. et al. Biological activity, and volatile and phenolic compounds from five lamiaceae species. Flavour. Fragr. J. 36, 223–232 (2021).
-
Beddiar, H. et al. Chemical composition, anti-quorum sensing, enzyme inhibitory, and antioxidant properties of phenolic extracts of clinopodium Nepeta L. Kuntze Plants. 10, 1955 (2021).
-
Briskin, D. P. Medicinal plants and phytomedicines. Linking plant biochemistry and physiology to human health. Plant. Physiol. 124, 507–514 (2000).
-
Ncube, B., Finnie, J. F. & Van Staden, J. Quality from the field: the impact of environmental factors as quality determinants in medicinal plants. S Afr. J. Bot. 82, 11–20 (2012).
-
Nascimento, N. C. & Fett-Neto, A. G. Plant secondary metabolism and challenges in modifying its operation: an overview. Methods Mol. Biol. 643, 1–13 (2010).
-
Espinosa-Leal, C. A., Puente-Garza, C. A. & García-Lara, S. In vitro plant tissue culture: means for production of biological active compounds. Planta 248, 1–18 (2018).
-
Zhou, L. G. & Wu, J. U. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Nat. Prod. Rep. 23, 789–810 (2006).
-
Nair, R. in Effects of Nanoparticles on Plant Growth and Development. 1–14 (eds Plant Nanotechnology, C., Kumar, D. & Khodakovskaya, M.) (Springer, 2016).
-
Kim, D. H., Gopal, J. & Sivanesan, I. Nanomaterials in plant tissue culture: the disclosed and undisclosed. RSC Adv. 7, 36492–36505 (2017).
-
Gonçalves, S. et al. Impact of metallic nanoparticles on in vitro culture, phenolic profile and biological activity of two mediterranean lamiaceae species: Lavandula viridis L’Hér and thymus Lotocephalus G. López and R. Morales. Molecules 26, 6427 (2021).
-
Nourozi, E., Hosseini, B., Maleki, R. & Mandoulakani, B. A. Inductive effect of titanium dioxide nanoparticles on the anticancer compounds production and expression of Rosmarinic acid biosynthesis genes in dracocephalum kotschyi transformed roots. Plant. Physiol. Biochem. 167, 934–945 (2021).
-
Ali, A. et al. Silver nanoparticles elicited in vitro callus cultures for accumulation of biomass and secondary metabolites in Caralluma tuberculata. Artif. Cells Nanomed. Biotechnol. 47, 715–724 (2019).
-
Sadak, M. S. Impact of silver nanoparticles on plant growth, some biochemical aspects, and yield of Fenugreek plant (Trigonella foenum-graecum). Bull. Natl. Res. Cent. 43, 38 (2019).
-
El-Temsah, Y. S. & Joner, E. J. Impact of Fe and ag nanoparticles on seed germination and differences in bioavailability during exposure in aqueous suspension and soil. Environ. Toxicol. 27, 42–49 (2010).
-
Sharma, P., Bhatt, D., Zaidi, M. G., Saradhi, P. P. & Khanna, P. K. Silver nanoparticle-mediated enhancement in growth and antioxidant status of brassica juncea. Appl. Biochem. Biotechnol. 167, 2225–2233 (2012).
-
Salama, H. M. H. Effects of silver nanoparticles in some crop plants, common bean (Phaseolus vulgaris L.) and corn (Zea Mays L). Int. Res. J. Biotech. 3, 190–197 (2012).
-
Tymoszuk, A. & Miler, N. Silver and gold nanoparticles impact on in vitro adventitious organogenesis in chrysanthemum, Gerbera and cape primrose. Sci. Hortic. 257, 108766 (2019).
-
Berahmand, A. A. et al. Effects silver nanoparticles and magnetic field on growth of fodder maize (Zea Mays L). Biol. Trace Elem. Res. 149, 419–424 (2012).
-
Vlachou, G., Papafotiou, M. & Bertsouklis, K. F. Studies on seed germination and micropropagation of clinopodium nepeta: a medicinal and aromatic plant. HortScience 54, 1558–1564 (2019).
-
Jiang, H. S., Li, M., Chang, F. Y., Li, W. & Yin, L. Y. Physiological analysis of silver nanoparticles and AgNO₃ toxicity to Spirodela polyrhiza. Environ. Toxicol. Chem. 31, 1880–1886 (2012).
-
Kaveh, R. et al. Changes in Arabidopsis Thaliana gene expression in response to silver nanoparticles and silver ions. Environ. Sci. Technol. 47, 10637–10644 (2013).
-
Dimkpa, C. O. et al. Silver nanoparticles disrupt wheat (Triticum aestivum L.) growth in a sand matrix. Environ. Sci. Technol. 47, 1082–1090 (2013).
-
Nair, P. M. G. & Chung, I. M. Physiological and molecular level effects of silver nanoparticles exposure in rice (Oryza sativa L.) seedlings. Chemosphere 112, 105–113 (2014).
-
Mirzajani, F., Askari, H., Hamzelou, S., Farzaneh, M. & Ghassempour, A. Effect of silver nanoparticles on Oryza sativa L. and its rhizosphere bacteria. Ecotoxicol. Environ. Saf. 88, 48–54 (2013).
-
Vinković, T., Štolfa-Čamagajevac, I., Tkalec, M., Goessler, W. & Domazet-Jurašin, D. Vinković-Vrček, I. Does plant growing condition affect biodistribution and biological effects of silver nanoparticles. Span. J. Agric. Res. 16, e0803 (2018).
-
Nourozi, E., Hosseini, B. & Malek, R. Abdollahi Mandoulakani, B. Iron oxide nanoparticle: a novel elicitor to enhancing anticancer flavonoid production and gene expression in dracocephalum kotschyi hairy-root cultures. J. Sci. Food Agric. 99, 6418–6430 (2019a).
-
Nourozi, E., Hosseini, B. & Maleki, R. Abdollahi Mandoulakani, B. Pharmaceutical important phenolic compounds overproduction and gene expression analysis in dracocephalum kotschyi hairy roots elicited by SiO₂ nanoparticles. Ind. Crops Prod. 133, 435–446 (2019b).
-
Dat, J. et al. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 57, 779–795 (2000).
-
Marslin, G., Sheeba, C. J. & Franklin, G. Nanoparticles alter secondary metabolism in plants via ROS burst. Front. Plant. Sci. 8, 832 (2017).
-
Ranjan, A. et al. Nanoparticles induced stress and toxicity in plants. Environ. Nanotechnol Monit. Manage. 5, 100457 (2021).
-
Jalil, S. U. et al. Nanoparticles and abiotic stress tolerance in plants: synthesis, action, and signaling mechanisms. In Plant Signaling Molecules: Role and Regulation Under Stressful Environments (ed. Khan, M. I. R. Elsevier Inc., 596 (2019).
-
Velusamy, P., Kumar, G. V., Jeyanthi, V., Das, J. & Pachaiappan, R. Biofabrication of ag nanoparticles using Moringa Oleifera leaf extract and their antimicrobial activity. Asian Pac. J. Trop. Biomed. 6, 60–66 (2016).
-
Dobravalskytė, D., Venskutonis, P. & Talou, R. Antioxidant properties and essential oil composition of calamintha grandiflora L. Food Chem. 135, 1539–1546 (2012).
-
Khojasteh, A. et al. New trends in biotechnological production of Rosmarinic acid. Biotechnol. Lett. 36, 2393–2406 (2014).
-
Sahraroo, A., Mirjalili, M. H., Corchete, P., Babalar, M. & Moghadam, F. R. Establishment and characterization of a satureja khuzistanica Jamzad (Lamiaceae) cell suspension culture: a new in vitro source of Rosmarinic acid. Cytotechnology 68, 1415–1424 (2016).
-
Costa, P., Gonçalves, S., Valentão, P., Andrade, P. B. & Romano, A. Accumulation of phenolic compounds in in vitro cultures and wild plants of Lavandula viridis l’her and their antioxidant and anti-cholinesterase potential. Food Chem. Toxicol. 57, 69–74 (2013).
-
Bektaş, E. Changes in essential oil composition, phenylalanine ammonia lyase gene expression, and Rosmarinic acid content during shoot organogenesis in cytokinin-treated satureja spicigera (C. Koch) boiss. Shoots. J. Plant. Biochem. Biotechnol. 29, 450–460 (2021).
-
Zygadlo, J. A., Merino, E. F., Maestri, D. M. & Guzman, C. A. Ariza Espinar, L. The essential oils of satureja Odora and S. parvifolia from Argentina. J. Essent. Oil Res. 5, 549–551 (1993).
-
Zaeem, A. et al. Effects of biogenic zinc oxide nanoparticles on growth and oxidative stress response in flax seedlings vs. in vitro cultures: a comparative analysis. Biomolecules 10, 918 (2020).
-
Choi, O. & Hu, Z. Size dependent and reactive oxygen species related nanosilver toxicity to nitrifying bacteria. Environ. Sci. Technol. 42, 4583–4588 (2008).
-
Venkatachalam, P. et al. Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena Leucocephala seedlings: A physiochemical analysis. Plant. Physiol. Biochem. 110, 59–69 (2017).
-
Javed, R., Usman, M., Yücesan, B., Zia, M. & Gürel, E. Effect of zinc oxide (ZnO) nanoparticles on physiology and steviol glycosides production in micropropagated shoots of stevia rebaudiana Bertoni. Plant. Physiol. Biochem. 110, 94–99 (2017).
-
Tungmunnithum, D., Thongboonyou, A., Pholboon, A. & Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: an overview. Medicines 5, 93 (2018).
-
Debbabi, H. et al. Chemical composition, antifungal and insecticidal activities of the essential oils from Tunisian clinopodium Nepeta subsp. Nepeta and clinopodium Nepeta subsp. Glandulosum. Molecules 25, 2137 (2020).
-
Benkhaira, N. et al. Unveiling the phytochemical profile, in vitro bioactivities evaluation, in Silico molecular Docking and ADMET study of essential oil from clinopodium Nepeta grown in middle atlas of Morocco. Biocatal. Agric. Biotechnol. 54, 102923 (2023).
-
Taibi, M. et al. Phytochemical characterization and multifaceted bioactivity assessment of essential oil from clinopodium Nepeta subsp. Ascendens: potential for the food and pharmaceutical sectors. CyTA – J. Food. 22, 2348735 (2024).
-
El Guerrouj, B. et al. The effect of altitude on the chemical composition, antioxidant and antimicrobial activities of Eucalyptus globulus Labill. Essential oils. Trop. J. Nat. Prod. Res. 7, 5279–5285 (2023).
-
Bektaş, E. & Sökmen, A. In vitro seed germination, plantlet growth, tuberization, and synthetic seed production of Serapias vomeracea (Burm.f.) Briq. Turk. J. Bot. 40, 584–594 (2016).
-
Murashige, T. & Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15, 473–497 (1962).
-
Jadczak, P., Kulpa, D., Bihun, M. & Przewodowski, W. Positive effect of AgNPs and AuNPs in in vitro cultures of Lavandula angustifolia mill. Plant. Cell. Tiss Org. 139, 191–197 (2019).
-
Singleton, V. L. & Rossi, J. A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 16, 144–158 (1965).
-
Chang, C. C., Yang, M. H., Wen, H. M. & Chern, J. C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 10, 178–182 (2002).
-
Bektaş, E., Sahin, H., Beldüz, A. O. & Güler, H. I. HIV-1-RT Inhibition activity of satureja spicigera (C. KOCH) BOISS. Aqueous extract and Docking studies of phenolic compounds identified by RP-HPLC-DAD. J. Food Biochem. 46, e13921 (2022).
-
Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin J. Sci. Technol. 26, 211–219 (2004).
-
Benzie, I. F. & Strain, J. J. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: the FRAP assay. Anal. Biochem. 239, 70–76 (1996).
-
Ellman, G., Courtney, K. & Andres, V. Feather-Stone, R. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 7, 88 (1961).
Acknowledgements
The author would like to thank Huseyin SAHIN (Espiye Vocational School, Giresun University) for his valuable support in conducting the high-performance liquid chromatography (HPLC) analysis.
Funding
This work was supported by Scientific Research Projects, Giresun University (Project No: FEN-BAP-A-250221-29).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Bektaş, E. Influence of silver nanoparticles on the in vitro growth, phenolic profile, antioxidant potential, enzyme inhibition, and essential oil composition of Clinopodium nepeta subsp. spruneri (Boiss.) Bartolucci & F. Conti. Sci Rep 15, 38744 (2025). https://doi.org/10.1038/s41598-025-22586-y
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-22586-y