
Introduction
Drought stress is a severe and rising constraint to worldwide agriculture and causes drastic impacts on plant development and physiological processes, and eventually reduces crop yield1,2. Stress caused by drought triggers metabolic imbalances, inhibition of photosynthesis, and retardation of plant growth and is a serious threat to global food security3,4. One of the most vital food crops in the world, wheat (Triticum spp.), is also especially vulnerable to water shortage. The enhanced occurrence and severity of drought associated with climate change also emphasize the importance of developing the molecular mechanisms of drought tolerance in wheat species5,6.
Protein kinases are pivotal in plant stress signaling and are responsible for perceiving and transmitting environmental stimuli. Of importance among them is the Sucrose Non-Fermenting 1-related Kinase (SnRK) family and a subset of it—SnRK3 and the Calcineurin B-like protein-interacting Protein Kinases (CIPKs), also commonly referred to as the SnRK3 subfamily7,8,9. CIPKs and Calcineurin B-like proteins (CBLs) serve as calcium decoders of calcium fluctuations to direct targeted physiological responses10,11. These responses are the regulation of ion homeostasis, osmotic adjustment, stomatal movement, antioxidant defense, and activation of stress-responsive genes12,13,14. Because of their multifunctional capacities at transcriptional and post-translational levels, CIPKs are considered strategic targets to enhance crop drought tolerance15,16.
Several studies have shown the functional significance of specific CIPK genes in crops. OsCIPK23 and AtCIPK23 play a fundamental role in potassium uptake and drought tolerance17,18; StCIPK18 promotes water holding and antioxidant activity in potato; and TaCIPK2 enhances drought tolerance in Arabidopsis19,20. These are involved in ABA-mediated signaling, ROS detoxification, and regulation of stress-related gene expression. CIPKs share standard structural features such as an N-terminal kinase domain and a C-terminal regulatory domain with the NAF/FISL motif, a requirement for CBL binding and formation of a complex21,22,23,24. In addition, specific CIPKs function independently of CBLs as evidenced by CIPK6 from tomato, which regulates ROS production through alternative protein interactions25.
In light of this progress in model plants and advanced crops, the functions and regulation mechanisms of the CIPK gene family in durum wheat (Triticum turgidum ssp. durum) are poorly understood. The lack of such information restrains the application of molecular breeding approaches to promote drought tolerance in the crop, which is of considerable importance to the economy and nutrition. Durum wheat is a good source of dietary fiber, vitamins (B, E), phenolic acids, carotenoids, and other bioactive molecules, and is predominantly utilized in the production of pasta and semolina26,27,28,29. Hence, deciphering the expression and regulatory patterns of CIPKs under drought and ABA stimuli in durum wheat has enormous implications in developing water-stress-compatible genotypes.
Here, we systemically identified and analyzed the CIPK gene family in durum wheat genomes. We compared their structural properties, expression patterns in response to drought and ABA treatment, and putative functions in ABA-dependent and ABA-independent pathways. By closing this knowledge gap, our research contributes to a better understanding of drought tolerance mechanisms and offers valuable targets for breeding climate-resilient durum wheat varieties.
Materials and methods
Identification of CIPK gene family members in T. turgidum genome
The CIPK genes of durum wheat (designated as TtCIPK) were investigated using the Triticum turgidum genome available at the Plant Ensembl database (https://plants.ensembl.org/Triticum_turgidum/Info/Index)30. To perform a homology search in the durum wheat genome, CIPK protein sequences from Arabidopsis and rice were used as reference queries in BLASTp tool with the e-value set to 1 × 10−5. In CIPK protein sequences, the presence of protein kinase and NAF domains was confirmed by checking with the Conserved Domains Database (CDD)31 and Pfam32. The physicochemical properties of TtCIPKs protein sequences, including molecular weight (MW), theoretical isoelectric point (pI), instability index (II), and grand average of hydropathicity (GRAVY) were assessed using the ProtParam prediction tool in the Expasy database (https,//web.expasy.org/protparam/)33. This research identified precise gene locations on chromosomes. It also mapped out exon–intron structure and conserved motifs, utilizing TBtools software (v0.6735)34. In addition, the prediction of TtCIPKs subcellular localization was conducted through the WoLF PSORT online tool (https://wolfpsort.hgc.jp/). Visualization of the TtCIPK gene properties was achieved through a graph created in GraphPad Prism 9.0.
TtCIPKs phylogenetic analysis
The amino acid sequences of CIPK in durum wheat, Arabidopsis, and rice (Oryza sativa) were obtained from the EnsemblPlants and NCBI databases. These sequences were then aligned by ClustalW35. The phylogenetic analysis, which studies the relationships among these plants, was carried out using the maximum likelihood (ML) method. This method was run 1000 times for bootstrap testing and was executed with the standard settings on the IQTREE web server36. The outcome file was transferred to the iTOL online tool to visualize the phylogeny tree37. The grouping of members in the phylogenetic tree was determined based on the location of clusters and the genetic distance between clusters.
Analysis of TtCIPKs duplication events
The duplicated TtCIPK genes were identified by comparing the coding sequences of pairs of TtCIPK genes and selecting those with an identity of 0.85 or higher. The Ka/Ks calculator in TBtools software was used to calculate the non-synonymous (Ka) and synonymous (Ks) values, which measure the purity selection pressure on duplicated genes38. To investigate the gene’s conservation level, the Ka/Ks ratio was calculated39. Similar results regarding the presence of tandem and segmental duplications were obtained by gene duplication location analysis40. The approximate times of duplication events (in Mya) were estimated using the formula T = Ks/2λ; λ is equal to 6.5 × 10–9 substitutions per synonymous site per year and describes the average rate of synonymous substitution in durum wheat41.
Promoter analysis of TtCIPK genes
The Plants Ensembl database was utilized to extract the 1500 bp promoter sequences preceding the ATG start codon for all TtCIPK genes. The analysis of cis-elements related to stresses, light, growth, and hormone responses was carried out through PlantCARE (http://bioinformatics.psb.-ugent.be/webtools/plantcare/html/)42.
Post-translational phosphorylation site prediction in TtCIPK proteins
The reversible post-translation modification of phosphorylation, a significant in vivo event in the signaling pathway related to plant stress responses, was predicted using the NetPhos 3.1 site with a threshold probability of 0.843.
3D structure prediction and possible binding site identification in TtCIPK proteins
The 3D structure of TtCIPK proteins was predicted using the Phyre2 database44. This procedure employs a homology modeling methodology that compares the protein sequence to known structures, generating an optimal 3D conformation. The Phyre investigator tool from the Phyre2 server was used to predict the locations of pocket sites as ligand-binding regions in the three-dimensional structure of TtCIPK proteins.
RNA-seq data analysis
In the present study, the available RNA-seq datasets of durum wheat were investigated to illustrate the expression profile of TtCIPK genes. The raw data, including PRJNA780180 datasets, relating to increased temperature (+ 2 °C) stress, and PRJNA1089221 datasets related to 5 °C temperature45, as cold stress, were investigated. Initially, low-quality regions and adapter sequences were recognized and removed using Trimmomatic, and then RNA-seq data were aligned using HISAT and HTSeq-count46 with the reference genome of durum wheat, Svevo.v1. To show the expression profile, the fold change of TtCIPKs was calculated by the NOISeq package47.
Durum wheat growth conditions
After surface sterilization, durum wheat seeds of the Dena cultivar were planted hydroponically and maintained at a temperature of 25 ± 3 °C, with relative humidity at 35 ± 5% and a 12-h light/dark photoperiod. At 10 days old, the seedlings were fed a half-strength Hoagland solution.
ABA treatment and drought stress
For drought treatment, 45-day-old durum wheat plants were exposed to 15% (w/v) polyethylene glycol (PEG6000)48,49. The pre-treatment with ABA occurred three days before drought stress, administered by external spraying of 100 μM ABA50,51. All treatments were conducted in a greenhouse with three biological replicates. Samples from both shoot and root tissues were collected at 0 (control), 1 (D1), 24 (D24), and 72 h (D72) after the drought treatment, frozen in liquid nitrogen, and stored in a − 70 °C freezer.
Isolation of RNA and production of cDNA
Total RNA was isolated from shoot and root tissues using the Sambio™ RNA Extraction Kit. Complementary DNA (cDNA) was synthesized according to the provided protocol of the Sambio™ cDNA Synthesis Kit, utilizing the supplied components for converting total RNA or mRNA into single-stranded cDNA.
Designing primers for the selected TtCIPK genes
Based on evolutionary analysis, a selection of six TtCIPK genes, which include TtCIPK33, TtCIPK42, TtCIPK12, TtCIPK43, TtCIPK27, and TtCIPK29, were investigated for gene expression. Suitable primers for the selected genes were designed (Supplementary Table 1) using the CDS sequences with the help of the online software Primer3Plus (http://www.bioinformatics.nl/primer3plus)52,53. Subsequently, the specificity of the primers was verified using Primer-BLAST in NCBI.
Real-time PCR analysis
The Sambio™ qPCR Master (EVA) with ROX was employed to assess relative expression levels according to the manufacturer’s instructions. The length of the fragments amplified by the specific primers ranges from 80 to 250 base pairs. The amplification conditions for PCR were 95 °C for 5 min, followed by 25–40 cycles at 95 °C for 15 s, 63 °C for 20 s, and 72 °C for 35 s. In this analysis, relative expression levels of each target gene were determined using the 2-ΔΔCt method and normalized to the housekeeping gene Actin.
Results
Characterization of TtCIPK family members
In the first phase, the homology search using Arabidopsis and rice sequences revealed 58 potential CIPK proteins in durum wheat. Subsequently, the presence of NAF and kinase domains in all members of the TtCIPK family was confirmed by Pfam and NCBI-CDD databases. Their nomenclature was updated to represent their chromosomal locations, from TtCIPK1 to TtCIPK58 (Table 1). Based on physicochemical properties, all the TtCIPK proteins ranged between 1035 to 1608 base pairs in CDS sequence length. The number of exons in TtCIPK genes varied from 1 to 15. TtCIPK10, the smallest protein, has 344 amino acids and a molecular weight of 37.30 kDa, while TtCIPK26, the largest protein, has 535 amino acids and weighs 58.69 kDa. GRAVY values ranged from -0.545 to -0.078, indicating hydrophilicity. Isoelectric points (pI) ranged from 5.95 for TtCIPK1 to 9.51 for TtCIPK28 and TtCIPK48 (Table 1).
Phylogenetic analysis of TtCIPK gene family
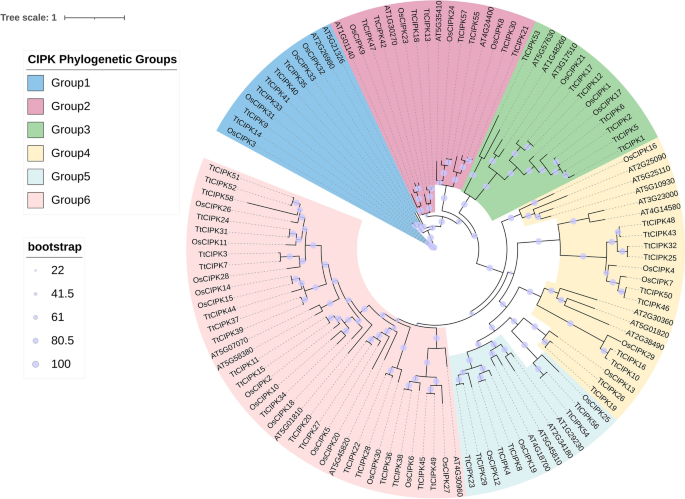
Phylogenetic analysis using the maximum likelihood method, based on protein sequences from durum wheat, rice, and Arabidopsis, clustered the 58 TtCIPK proteins into six evolutionary groups (Fig. 1). The TtCIPK proteins were unevenly distributed across these six groups. Group 1 comprised six proteins: TtCIPK14, TtCIPK9, TtCIPK33, TtCIPK41, TtCIPK40, and TtCIPK35. Group 2 consisted of eight TtCIPK proteins (including TtCIPK47, TtCIPK42, TtCIPK18, TtCIPK13, TtCIPK57, TtCIPK55, TtCIPK30, and TtCIPK21), and Group 3 consisted of seven TtCIPK proteins (including TtCIPK53, TtCIPK17, TtCIPK12, TtCIPK6, TtCIPK2, TtCIPK5, and TtCIPK1). Group 4 consisted of ten TtCIPK proteins (including TtCIPK48, TtCIPK43, TtCIPK32, TtCIPK25, TtCIPK50, TtCIPK46, TtCIPK16, TtCIPK10, TtCIPK26, and TtCIPK19), and Group 5 consisted of six TtCIPK proteins (including TtCIPK56, TtCIPK54, TtCIPK8, TtCIPK4, TtCIPK29, and TtCIPK23). Group 6 stood out as the biggest, housing the other 21 proteins. This hints that group 6 members may have followed a different evolutionary path or have special functional traits; CIPK proteins from rice and Arabidopsis were spread across all groups. This points to a complicated evolutionary background and wide-ranging functional roles.

Phylogeny tree of CIPK gene family in durum wheat (TtCIPK), Arabidopsis (started with AT), rice (started with Os).
The phylogenetic groups of TtCIPKs were compared based on their physicochemical characteristics. The exon number in groups 1, 2, and 3 falls between 12 and 15, with 14 exons being the most common. This points to structural likeness and, as a result, more functional similarity among these three phylogenetic groups. Groups 4, 5, and 6 have one exon, with some cases showing two. This suggests that these groups are related in evolution (Fig. 2a). Based on these results, the members of this gene family can be divided into two groups: low exon and high exon number. The isoelectric point (pI) of proteins in evolutionary groups 2 and 4 covers a broader range and hovers around 9. Groups 5 and 6 have a more limited isoelectric point close to 9 (Fig. 2b). The GRAVY index is negative for all groups, which means they are hydrophilic. Except for group 1, where members mostly share a similar GRAVY index, the other groups display a diverse range of GRAVY values (Fig. 2c). The stability index was measured across different groups, revealing that all members of group 5 are unstable, while all members of group 3 are stable. In the other groups, both stable and unstable members are present. Groups 1, 2, and 6 have more stable than unstable members, while group 4 has more unstable members (Fig. 2d).

Physiochemical properties of phylogeny tree groups of TtCIPK family in terms of exon number (a), pI values (b), GRAVY index (c), and instability index (d).
Structure analysis of TtCIPK family members
All proteins within this gene family possess a conserved kinase domain, which is directly related to their function. It is located in a nearly identical position in all members, shown in blue in Fig. 3a. The NAF domain is a crucial conserved domain in TtCIPKs, playing a regulatory role in the function of these proteins, and is found in a relatively similar position in all proteins, depicted in pink in Fig. 3a. According to gene structure analysis, the genes TtCIPK21 and TtCIPK30 have long introns, which may be associated with functional diversity, gene expression regulation, or their role in evolution (Fig. 3b).

Sequence analysis of TtCIPK family members based on conserved motifs distribution (a), and gene structure (b).
Duplication events and subcellular localization of TtCIPKs
TtCIPK gene family in durum wheat is distributed across all chromosomes with at least one TtCIPK gene in each chromosome. The genomic positions of the CIPK gene family members in durum wheat are well illustrated in Fig. 4a. An analysis of duplication events within this gene family revealed that some members have undergone segmental duplication. We could not detect any tandem duplications in TtCIPKs. The earliest duplication event in these genes occurred approximately 16 million years ago between TtCIPK25 and TtCIPK49. A Ka/Ks ratio less than 0.4 in duplicated genes means Ks is much higher than Ka, which means strong negative selection on these genes (Supplementary Table 3). This implies these genes are functional and have retained their function over time.

Location of TtCIPK genes on the genome of T. turgidum (a). Duplicated genes are linked by gray lines. Subcellular localization of TtCIPK proteins (b).
Subcellular localization of TtCIPK proteins based on their amino acid sequences indicated that these proteins are present in different cellular compartments. Most CIPK proteins are likely to be localized in chloroplast and cytosol respectively. The presence of these proteins in other cellular compartments is possible, with the lowest probabilities being in vacuoles and mitochondria (Fig. 4b). It is predicted that TtCIPK11 and TtCIPK15, belonging to group 6, are more likely to be found in the cell nucleus (Supplementary Table 2).
Pocket site of CIPK proteins
Three-dimensional structure analysis revealed that TtCIPK proteins have a conserved structure and share minimal structural variation (Supplementary Fig. 1). However, there were differences in the binding and interaction sites (pocket site). The nature of amino acids within the active site of proteins determines the chemical properties of this region, thus affecting the ability of the protein to interact with a variety of other ligands and molecules. An analysis of the amino acids in the pocket site of the TtCIPK proteins shows that the most abundant are leucine, valine, aspartic acid, and lysine, while tryptophan and proline are rare in this region (Fig. 5).

Frequency of residues located in the pocket site of TtCIPK proteins.
Post-translational modifications in CIPKs
Phosphorylation-induced conformational modifications are considered an effective mechanism that regulates the activity of CIPKs54. In line with this, we predicted amino acids within TtCIPK structures, which most likely will be phosphorylated. The prediction results indicate that all three major amino acids involved in phosphorylation, serine, threonine, and tyrosine, are likely to be phosphorylated with a probability of 0.8 in TtCIPK proteins. Among these proteins, serine is most likely to be the common amino acid receiving a phosphate group in all the phylogeny groups (Fig. 6). Additionally, the results revealed that there is a great diversity between the TtCIPK proteins in the potential phosphorylation sites, which can affect their function and lifespan.

Prediction of the phosphorylation sites in TtCIPK proteins.
Expression patterns of TtCIPK genes based on RNA-seq data
The expression profile based on RNA-seq data disclosed that the transcription of TtCIPK genes under temperature changes exposed the powerful and versatile role of these genes in signaling pathways related to abiotic stress (Fig. 7). Especially, the TtCIPK49 gene was upregulated under cold stress, which might indicate its functional role in cold tolerance. On the other hand, low expression levels of TtCIPK13 and TtCIPK26 under cold stress may indicate that these genes were downregulated by such a type of stress. Generally, 58 TtCIPK genes showed a decreasing trend in the expression analysis except TtCIPK38, TtCIPK36, and TtCIPK34. Interestingly, both TtCIPK38 and TtCIPK26 were highly upregulated under heat stress; therefore, they are good candidates to be overexpressed to develop improved heat stress tolerance. Further, detailed studies of the expression of such genes are required for a crystal-clear understanding of the underlying molecular mechanisms of abiotic stress tolerance in plants.

Expression profile of TtCIPK genes in response to temperature increasing and cold stress.
Upstream analysis of TtCIPK genes
The analysis of the upstream regions revealed a high diversity among the members of this family based on the frequency and type of cis-element in the promoter of TtCIPK genes. Our finding indicated that, overall, MYB and MYC cis-regulatory elements are the most frequent in the promoters of TtCIPK genes (Fig. 8a). Besides, the results show that most cis-regulatory elements are located upstream of TtCIPK genes associated with stress responses (Fig. 8b). Among the hormonal cis-elements, which are the second most abundant after stress-related cis-elements, those responsive to ABA and methyl jasmonate are more prevalent (Fig. 8c).

Promoter analysis of TtCIPK genes. Distribution of cis-regulatory elements in the promoter region of TtCIPK genes (a). Frequency of cis-regulatory elements based on their function (b) and frequency of cis-regulatory elements related to hormones (c).
Expression levels of TtCIPK genes in shoot tissues in response to drought stress
In the shoot tissues, the relative expression levels of a selected number of TtCIPK genes from each phylogenetic group were tested under drought stress conditions at 1, 24, and 72 h after treatment (indicated as D1, D24, and D72, respectively) with or without the application of ABA (Fig. 9). Upregulation of TtCIPK12, TtCIPK27, TtCIPK29, and TtCIPK33 expression was induced in shoot tissue upon drought treatment; this upregulation was significantly suppressed with the application of ABA at D24 for TtCIPK12, TtCIPK27, and TtCIPK29. For TtCIPK33, significant upregulation was observed upon drought stress at D1 and D24, and with the application of ABA, the transcription of this gene is intensified. This includes expression upregulation in TtCIPK43 following drought exposure at D1 and D24, but downregulated with ABA application at these time points, then upregulated at D72. Taken together, the upregulation of these genes by drought in shoots indicates that they have a vital role in responding to such stress.

Relative expression of candidate TtCIPK genes in shoot tissues in response to drought stress and ABA application. Each column is marked with an asterisk to show a significant variance (p-value < 0.05) from the control group, based on the t-test.
Expression levels of TtCIPK genes in root tissues in response to drought stress
In root tissues, TtCIPK12 was upregulated after 1 (D1) and 24 h (D24) under drought stress (Fig. 10). With ABA treatment, it was up-regulated in control and down-regulated at D1 and D24. The TtCIPK27 gene showed increased expression at D24, which was reduced upon ABA application (Fig. 10). The TtCIPK33 gene demonstrated a significant increase in expression in the control plants treated with ABA. For TtCIPK42, ABA application resulted in elevated gene expression in both control plants and at D24. The TtCIPK43 gene exhibited increased expression at D1 under drought stress, and ABA had a notable effect on its expression in control plants, reducing expression at D1 and increasing it at D72. Roots exhibit a lesser tendency for TtCIPK gene upregulation than shoots under drought stress. Moreover, the effect of ABA in inducing gene upregulation in roots mainly occurred in control plants.

Relative expression of candidate TtCIPK genes in root tissues in response to drought stress and ABA application. Each column is marked with an asterisk to show a significant variance (p-value < 0.05) from the control group, based on the t-test.
Discussion
Given the role of CIPKs in plant physiological processes, including stress responses, hormonal signaling, and growth and development, their study in crop plants seems essential55,56,57. CIPKs interact with CBLs to form a network that establishes a physiological link between plant growth and stress tolerance58. Durum wheat is essential in regions affected by water stress limitations59. The CIPK gene family has been extensively studied in various plant types like Arabidopsis, wheat, tomato, and soybean10,60,61,62. However, so far, the members of this gene family have not been identified and investigated in durum wheat. In this study, 58 members of the TtCIPK family were identified, showing differences in structure and some physicochemical properties. The large number of members of this gene family in durum wheat can be related to the complete doubling of its genome, and considering the role of CIPK in response to environmental stresses, increasing the number of members has been effective in increasing the tolerance of this plant. The study of evolutionary relationships showed that TtCIPKs are separated into six groups, and a high genetic distance was observed between them. This genetic distance and the placement of orthologs of TtCIPKs in monocot and dicot model plants in the phylogeny tree convey the message that the diversity in the members of this gene family occurred both before and after the derivation of dicots and monocots. Domain analysis indicates that TtCIPKs have their kinase domains located always near the N-terminus, while NAF domains are uniformly found close to the C-terminus. The structural analysis of TtCIPK evolutionary groups indicates that groups 1, 2, and 3 are intron-rich, whereas those belonging to groups 4, 5, and 6 are known for their lack of apparent introns. This scenario has also been noted during other plants’ CIPK gene family investigations, such as banana11,63. Two genes, TtCIPK30 and TtCIPK21, belonging to the second evolutionary group, contain long introns. Long introns may indicate the complexity of gene expression regulation64.
Introns harbor regulatory elements that influence transcription, splicing, and mRNA stability. These elements help to precisely regulate gene expression in response to different stimuli across tissues. Additionally, since introns are not translated, they tolerate mutations and contribute to genetic diversity over evolutionary time. Although CIPKs phosphorylate downstream targets in stress response pathways, they are also subject to phosphorylation. For example, phosphorylation of MdCIPK22 has been reported to be induced by ABA and occurs at threonine 41156. We analyzed post-translational modifications in the TtCIPK family and predicted that all TtCIPK proteins are likely phosphorylated at serine residues, with less likelihood at threonine and tyrosine residues. A conserved structure was observed based on the three-dimensional structure prediction in TtCIPK proteins. However, based on the position and type of amino acid, TtCIPK family members differed from each other. The origin of this difference can be related to the mutations that have been created and stabilized during evolution, resulting in functional diversity. In addition, it seems that the amino acids leucine, valine, aspartic acid, and lysine play a key role in the function and interaction of TtCIPKs in the associated signal transmission networks, although further studies are needed.
Of the 58 identified TtCIPK genes, 50 contained the ABRE (Abscisic Acid Responsive Element), a crucial cis-element in stress responses65. ABA has shown a significant role in enhancing tolerance to environmental challenges66,67. Many CIPK genes are regulated by ABA under stress conditions, and some participate in the ABA signaling pathway12,68. The results from the expression analysis of TtCIPKs indicated that these genes were dynamically and differentially expressed in the root and shoot tissues under drought stress. Their regulations were often differentially influenced by ABA treatments. Of these, TtCIPK12 and TtCIPK27 were identified as two major ABA-independent genes playing key roles during the early stages of drought responses (Fig. 11). Both genes were significantly upregulated during the early stages of drought stress, both in root and shoot tissues, suggesting that their participation in rapid stress sensing and mitigation is relevant. Results are in agreement with literature data pointing out the involvement of both factors in early stages of stress responses using calcium ions as second messengers within the stress signal transduction pathways69. On the contrary, TtCIPK33 was strongly induced in roots and shoots, whereas TtCIPK42 was only strongly induced in roots upon ABA treatment; hence, they may take part in the ABA-mediated signaling pathways (Fig. 11). Notably, TtCIPK33 presented a constitutive expression pattern in several tissues and at different time points, which agrees with a dual regulatory role and a candidate function in sustained adaptation responses. These observations are consistent with similar calcium signaling previously reported in other cereals21.

Schematic representation of ABA-dependent and independent pathways and candidate TtCIPK genes involved in them during drought.
These results highlight that the spatial and temporal expression patterns of TtCIPK genes under drought stress in shoot and root tissues reflect differential functional aspects. For instance, specific genes like TtCIPK27 and TtCIPK29 responded promptly to the early drought signal in shoot tissues, whereas others, such as TtCIPK33, continued their expression in longer-term stress. Significantly, these responses were modulated by ABA treatments in either upregulation or downregulation depending on specific genes and tissues, respectively, in agreement with previous studies that have emphasized the vital role of ABA in inter-tissue coordination of various stress responses. The integration of TtCIPK genes into ABA-dependent and ABA-independent pathways points out their varied functions in durum wheat’s mechanisms of drought tolerance. The present work is one of the few pioneering studies to clearly outline the dual regulatory roles of certain CIPKs in durum wheat and has highlighted the space and time patterns of its expression. In addition, a detailed study on TtCIPK12, TtCIPK33, and TtCIPK43 builds a good foundation for future studies on their involvement in cross-talk with stress signaling.
Although the gene expression level alone cannot provide the accurate function of TtCIPKs, TtCIPK33 was considered the most promising candidate for improving drought tolerance since it was always involved in drought stress responses. Other similar members of this CIPK family have already been used in other plant species to enhance drought and abiotic stress tolerance19,55. Significant abiotic stresses were checked along with the expression of other genes in TtCIPK genes; some included being treated with ABA, polyethylene glycol, high temperature, or cold stress. The presented study contributes to a basic understanding of evolutionary development expressed within the CIPK gene family of durum wheat and thus provides the real grounds for functional future investigations among such genes.
The basis of this study is the previous studies that, for the first time, compare ABA-dependent and ABA-independent CIPK pathways directly in durum wheat, integrating promoter element analyses with tissue-specific expression profiling. Unlike the previously reported Arabidopsis and rice studies that mainly focused on a single pathway under specified stress, this research takes a broad approach to unraveling the regulatory mechanisms underlying the activity of CIPK. Furthermore, a few unique phosphorylation sites in durum wheat CIPKs have been identified. They are adding new dimensions to functional diversity and regulatory mechanisms, thus complementing the findings from potato and quinoa studies19,70.
This also suggests that TtCIPKs play a core role in calcium signaling, mediating stress responses that are potential means of improving crop resilience. By mediating stomatal closure, osmotic adjustment, and ROS detoxification, TtCIPKs contribute to water-use efficiency and drought tolerance20,71. Further diversity in cis-regulatory elements is involved in the response of plants to other abiotic stresses, including salinity and cold, positioning these genes as candidates for multi-stress tolerance breeding in crops. Progress in genome-editing technologies has allowed or will allow exact modification of promoter regions, including the recently developed CRISPR-Cas9 system, which might lead to new ways of activating stress-responsive genes and new strategies for crop improvement. This work also presents not only the complex and multi-dimensional nature of the drought and ABA stress response mechanism but also points toward the TtCIPK gene family as a potential key player in enhancing stress tolerance and resilience in durum wheat.
Data availability
The data generated or analyzed in this study are included in this article. Other materials that support the findings of this study are available from the corresponding author upon reasonable request.
References
-
Solomon, K. F. & Labuschagne, M. T. Expression of drought tolerance in F1 hybrids of a diallel cross of durum wheat (Triticum turgidum var. durum L.). Cereal Res. Commun. 31, 49–56 (2003).
-
Diab, A. A. et al. Drought – inducible genes and differentially expressed sequence tags associated with components of drought tolerance in durum wheat. Sci. Res. Essay 3, 9–26 (2008).
-
Joshi, R. et al. Transcription factors and plants response to drought stress : Current understanding and future directions. Front. Plant Sci. 7, 1029 (2016).
-
Gupta, A., Rico-Medina, A. & Caño-Delgado, A. I. The physiology of plant responses to drought. Science (80-) 368, 266–269 (2020).
-
Rampino, P. et al. Novel durum wheat genes up-regulated in response to a combination of heat and drought stress. Plant Physiol. Biochem. 56, 72–78 (2012).
-
Melloul, M. et al. Identification of differentially expressed genes by cDNA-AFLP technique in response to drought stress in Triticum durum. Food Technol. Biotechnol. 52, 479–488 (2014).
-
Zhu, J. Review abiotic stress signaling and responses in plants. Cell 167, 313–324 (2016).
-
Khan, M., Jannat, A., Munir, F., Fatima, N. & Rabia, A. Biochemical and molecular mechanisms of abiotic stress tolerance. In Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives II: Mechanisms of Adaptation and Stress Amelioration 187–230 (Springer, 2020).
-
Xiao, X. et al. The calcium sensor calcineurin B-like proteins -calcineurin B-like interacting protein kinases is involved in leaf development and stress responses related to latex flow in Hevea brasiliensis. Front. Plant Sci. 13, 743506 (2022).
-
Zhu, K. et al. Evolution of an intron-poor cluster of the CIPK gene family and expression in response to drought stress in soybean. Sci. Rep. 6, 28225 (2016).
-
Yang, C. et al. Diverse roles of the CIPK gene family in transcription regulation and various biotic and abiotic stresses: A literature review and bibliometric study. Front. Genet. 13, 1041078 (2022).
-
Kim, K. N., Cheong, Y. H., Grant, J. J., Pandey, G. K. & Luan, S. CIPK3, a calcium sensor-associated protein kinase that regulates abscisic acid and cold signal transduction in arabidopsis. Plant Cell 15, 411–423 (2003).
-
Cheong, Y. H. et al. Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis. Plant J. 52, 223–239 (2007).
-
You, Z., Guo, S., Li, Q., Fang, Y. & Huang, P. The CBL1 / 9-CIPK1 calcium sensor negatively regulates drought stress by phosphorylating the PYLs ABA receptor. Nat. Commun. 14, 5886 (2023).
-
Sanyal, S. K. et al. Alternative splicing of CIPK3 results in distinct target selection to propagate ABA signaling in Arabidopsis. Front. Plant Sci. 8, 1924 (2017).
-
Lu, L. et al. CIPK11: a calcineurin B-like protein-interacting protein kinase from Nitraria tangutorum, confers tolerance to salt and drought in Arabidopsis. BMC Plant Biol. 21, 1–16 (2021).
-
Su, W. et al. New insights into the evolution and functional divergence of the CIPK gene family in Saccharum. BMC Genom. 21, 1–20 (2020).
-
Wang, Q., Zhao, K., Gong, Y., Yang, Y. & Yue, Y. Genome-wide identification and functional analysis of the calcineurin B-like protein and calcineurin B-like protein-interacting protein kinase gene families in Chinese cabbage (Brassica rapa ssp. pekinensis). Genes 13, 1191 (2022).
-
Yang, L. et al. CBL-interacting protein kinases 18 (CIPK18) gene positively regulates drought resistance in potato. Int. J. Mol. Sci. 24, 3613 (2023).
-
Wang, Y. et al. A CBL-interacting protein kinase TaCIPK2 confers drought tolerance in transgenic tobacco plants through regulating the stomatal movement. PLoS ONE 11, e0167962 (2016).
-
Tang, R. J., Wang, C., Li, K. & Luan, S. The CBL–CIPK calcium signaling network: Unified paradigm from 20 years of discoveries. Trends Plant Sci. 25, 604–617 (2020).
-
Paes de Melo, B. et al. Abiotic stresses in plants and their markers: A practice view of plant stress responses and programmed cell death mechanisms. Plants 11, 1100 (2022).
-
Xi, Y., Liu, J., Dong, C. & Cheng, Z. M. M. The CBL and CIPK gene family in grapevine (Vitis vinifera): Genome-wide analysis and expression profiles in response to various abiotic stresses. Front. Plant Sci. 8, 978 (2017).
-
Lu, T., Zhang, G., Sun, L., Wang, J. & Hao, F. Genome-wide identification of CBL family and expression analysis of CBLs in response to potassium deficiency in cotton. PeerJ 5, e3653 (2017).
-
Gutiérrez-Beltrán, E., Personat, J. M., de la Torre, F. & del Pozo, O. A universal stress protein involved in oxidative stress1 is a phosphorylation target for protein kinase CIPK6. Plant Physiol. 173, 836–852 (2017).
-
Arzani, A. & Ashraf, M. Cultivated ancient wheats (Triticum spp.): A potential source of health-beneficial food products. Compr. Rev. Food Sci. Food Saf. 16, 477–488 (2017).
-
Saini, P. et al. Nutritional value and end-use quality of durum wheat. Cereal Res. Commun. 51, 283–294 (2023).
-
De Santis, M. A., Soccio, M., Laus, M. N. & Flagella, Z. Influence of drought and salt stress on durum wheat grain quality and composition: A review. Plants 10, 2599 (2021).
-
Ficco, D. B. M. et al. Durum wheat bread with a potentially high health value through the addition of durum wheat thin bran or barley flour. Plants 12, 397 (2023).
-
Bolser, D. M., Staines, D. M. & Perry, E. K. P. Ensembl plants: integrating tools for visualizing, mining, and analyzing plant genomic data. Plant Genom. Databases Methods Protoc. 1533, 1–31 (2017).
-
Marchler-Bauer, A. et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 43, D222–D226 (2015).
-
Finn, R. D. et al. Pfam: The protein families database. Nucleic Acids Res. 42, 222–230 (2014).
-
Gasteiger, E. et al. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 31, 3784–3788 (2003).
-
Chen, C. et al. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 13, 1194–1202 (2020).
-
Thompson, J. D., Gibson, T. J. & Higgins, D. G. Multiple sequence alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 00, 1–22 (2003).
-
Nguyen, L. T., Schmidt, H. A., Von Haeseler, A. & Minh, B. Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 32, 268–274 (2015).
-
Letunic, I. & Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 47, 256–259 (2019).
-
Yaghobi, M. & Heidari, P. Genome-wide analysis of aquaporin gene family in Triticum turgidum and Its expression profile in response to salt stress. Genes 14, 202 (2023).
-
Hurst, L. D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 18, 486–487 (2002).
-
Cannon, S. B., Mitra, A., Baumgarten, A., Young, N. D. & May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 4, 10 (2004).
-
Liu, D. et al. Phylogenetic analyses provide the first insights into the evolution of OVATE family proteins in land plants. Ann. Bot. 113, 1219–1233 (2014).
-
Lescot, M. et al. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 30, 325–327 (2002).
-
Puresmaeli, F., Heidari, P. & Lawson, S. Insights into the sulfate transporter gene family and its expression patterns in durum wheat seedlings under salinity. Genes 14, 333 (2023).
-
Kelley, L. A., Mezulis, S., Yates, C. M., Wass, M. N. & Sternberg, M. J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 10, 845–858 (2016).
-
Díaz, M. L., Soresi, D. S., Basualdo, J., Cuppari, S. J. & Carrera, A. Transcriptomic response of durum wheat to cold stress at reproductive stage. Mol. Biol. Rep. 46, 2427–2445 (2019).
-
Anders, S., Pyl, P. T. & Huber, W. HTSeq-A python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169 (2015).
-
Tarazona, S., García, F., Ferrer, A., Dopazo, J. & Conesa, A. NOIseq: a RNA-seq differential expression method robust for sequencing depth biases. EMBnet J. 17, 18 (2012).
-
Faraji, S. et al. The AP2/ERF gene family in Triticum durum: Genome-wide identification and expression analysis under drought and salinity stresses. Genes 11, 1–24 (2020).
-
Huang, A. H. et al. Response of maize seed germination characteristics to different concentrations PEG-6000. Southwest China J Agric Sci. 34, 2628–2633 (2021).
-
Sripinyowanich, S., Klomsakul, P. & Boonburapong, B. Exogenous ABA induces salt tolerance in indica rice ( Oryza sativa L.): The role of OsP5CS1 and OsP5CR gene expression during salt stress. Environ. Exp. Bot. 86, 94–105 (2013).
-
Ayadi, M., Cavez, D., Miled, N., Chaumont, F. & Masmoudi, K. Identification and characterization of two plasma membrane aquaporins in durum wheat (Triticum turgidum L. subsp. durum) and their role in abiotic stress tolerance. Plant Physiol. Biochem. 49, 1029–1039 (2011).
-
Untergasser, A. et al. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 35(2), 71–74 (2007).
-
Thornton, B. & Basu, C. Rapid and simple method of qPCR primer design. Methods Mol. Biol. 1275, 173–179 (2015).
-
Chaves-sanjuan, A., Sanchez-barrena, M. J., Gonzalez-rubio, J. M., Moreno, M. & Ragel, P. Structural basis of the regulatory mechanism of the plant CIPK family of protein kinases controlling ion homeostasis and abiotic stress. Proc. Natl. Acad. Sci. 111, E4532–E4541 (2014).
-
Ma, R. et al. Genome-wide identification, characterization and expression analysis of the CIPK gene family in potato ( Solanum tuberosum L.) and the role of StCIPK10 in response to drought and osmotic stress. Int. J. Mol. Sci. 22, 13535 (2021).
-
Niu, L., Dong, B., Song, Z. & Meng, D. Genome-wide identification and characterization of CIPK family and analysis responses to various stresses in apple ( Malus domestica). Int. J. Mol. Sci. 19, 2131 (2018).
-
Li, P., Zheng, T. & Li, L. Identi fi cation and comparative analysis of the CIPK gene family and characterization of the cold stress response in the woody plant Prunus mume. PeerJ 7, e6847 (2019).
-
Mao, J. et al. The CBL-CIPK network is involved in the physiological crosstalk between plant growth and stress adaptation. Plant Cell Environ. 46, 3012–3022 (2023).
-
Martínez-Moreno, F., Ammar, K. & Solís, I. Global changes in cultivated area and breeding activities of durum wheat from 1800 to date: A historical review. Agronomy 12, 1–17 (2022).
-
Zhang, Y. et al. Identification and functional analysis of tomato CIPK gene family. Int. J. Mol. Sci. 21, 110 (2020).
-
Yu, Y., Xia, Æ. X. & Yin, Æ. W. Comparative genomic analysis of CIPK gene family in Arabidopsis and Populus. Plant Growth Regul. 35, 101–110 (2007).
-
Sun, T. et al. Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.). BMC Plant Biol. 15, 269 (2015).
-
Narwal, P., Singh, R. K., Pathak, A. K., Kumar, D. & Negi, N. P. Genome-wide profiling of CBL interacting protein kinases (CIPKs) in banana unveils their role in abiotic stress signaling and stress tolerance enhancement. Plant Stress 11, 100417 (2024).
-
Sharifi, S. A., Zaeimdar, M., Jozi, S. A. & Hejazi, R. Effects of soil, water and air pollution with heavy metal ions around lead and zinc mining and processing factories. Water Air Soil Pollut. 234, 760 (2023).
-
Zhao, J. et al. Foxtail millet (Setaria italica (L.) P. Beauv) CIPKs are responsive to ABA and abiotic stresses. PLoS ONE 14, e0225091 (2019).
-
Sah, S. K., Reddy, K. R. & Li, J. Abscisic acid and abiotic stress tolerance in crop plants. Front. Plant Sci. 7, 571 (2016).
-
Aryadeep, R., Saikat, P. & Basu, S. Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress. Plant Cell Rep. 32, 985–1006 (2013).
-
Zhu, L. et al. Genome-wide investigation and expression analysis of the Nitraria sibirica Pall. CIPK gene family. Int. J. Mol. Sci. 23, 11599 (2022).
-
Kumar, M. et al. Integration of abscisic acid signaling with other signaling pathways in plant stress responses and development. Plants 8, 1–20 (2019).
-
Xiaolin, Z., Baoqiang, W., Xian, W. & Xiaohong, W. Identification of the CIPK – CBL family gene and functional characterization of CqCIPK14 gene under drought stress in quinoa. BMC Genom. 23, 447 (2022).
-
Ravi, B., Foyer, C. H. & Pandey, G. K. The integration of reactive oxygen species (ROS) and calcium signalling in abiotic stress responses. Plant Cell Environ. 46, 1985–2006 (2023).
Funding
Funding was provided by Ferdowsi University of Mashhad, Iran (Grant No. 3/60500).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Pouya, H.S.H., Cheniany, M. & Heidari, P. Insights into TtCIPK gene family and its roles in durum wheat in response to PEG and ABA treatments. Sci Rep 15, 25419 (2025). https://doi.org/10.1038/s41598-025-11099-3
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-11099-3