
Introduction
Spinal cord injury (SCI) is a severe condition that leads to long-term or permanent loss of sensory and motor function below the injury site1,2. This leads to neuronal death and myelin degeneration, which are further exacerbated by secondary injury mechanisms that elevate oxidative stress (OS), promote inflammation, induce neuronal and oligodendrocyte apoptosis, and contribute to cystic cavity formation3,4,5,6,7,8. These pathological events progressively alter neuronal activity and disrupt the structural integrity of the damaged tissue by enhancing lipid peroxidation (LPO) and generating reactive aldehydes9,10,11,12. Notably, experimental studies have demonstrated that therapeutic approaches targeting oxidative stress, by reducing reactive oxygen species (ROS) and LPO, can significantly improve histological and functional outcomes following SCI3,13. This therapeutic potential is attributed to the high content of unsaturated fatty acids in spinal cord neurons, which renders them particularly vulnerable to LPO-induced damage14,15. Consequently, considerable attention has been directed toward the development of novel treatment strategies for SCI, with particular focus on bioactive compounds and cell-based therapies as promising antioxidant interventions4,7,16,17,18,19.
There has been a recent focus on electrospun fiber scaffolds as a potential option for treating SCIs. These nanofibers have advantages over traditional biological materials in terms of their structure and porosity. They resemble the natural extracellular matrix, which can help with cell attachment and development20,21,22. To facilitate spinal cord regeneration, various types of polymers, particularly polyesters, have been developed for scaffolds. Polyesters such as polycaprolactone (PCL) possess desirable qualities, such as strength, biocompatibility, and biodegradability23. However, the low hydrophilicity of PCL limits its effectiveness in terms of cell adhesion, proliferation, migration, and differentiation20,24. One solution involves increasing the wettability of PCL fibers by incorporating hydrophilic materials. Multiwalled carbon nanotubes (MWCNTs) have been widely used in regenerating different tissues. Their needle-like structure enhances scaffold strength, and their inner hollow space and high surface area allow for binding with bioactive molecules25. Owing to their conductivity, CNTs also have good compatibility with interneuronal cellular communication26. However, their hydrophobic nature leads to severe aggregation in aqueous solutions, limiting their biological applications. To overcome this, functionalizing of MWCNTs (f-MWCNTs) is necessary27.
In addition to providing a suitable substrate for axon growth and neuronal recovery, biologically active substances play a crucial role for controlling the oxidative environment of SCI3,6. By incorporating an antioxidant drug into a polymer system near the target tissue, it is possible to increase the local drug concentration and reduce tissue exposure to stress20. Ellagic acid (EA), a polyphenol, has gained significant attention due to its various physiological and pharmacological effects28,29,30. EA has been found to regulate cyclooxygenase (COX) enzyme activity, exhibiting strong anti-inflammatory properties in addition to modulating oxidative status and LPO31,32,33,34. However, EA suffers from poor water solubility and limited therapeutic efficacy and clinical applicability because of its high log P35. This issue can be resolved by encapsulating the drug into liposomes. They protect bioactive compounds from various environmental factors, such as enzymatic degradation, oxidation, molecular interactions, and pH changes36. Liposomes are not cytotoxic and do not stimulate the immune system37. Moreover, liposomes can carry both hydrophilic and hydrophobic drugs and enhance surface hydrophilicity to support cell attachment36. Microencapsulation of EA within nanoliposomes provides better absorption for tissue regeneration, longer release, and increased drug half-life with regulated distribution35.
The need for cell therapy is intensified by the inability of the spinal cord to heal itself and replace lost native cells18. Recent data have shown that mesenchymal stem cell (MSC) therapy is effective and safe for improving spinal cord dysfunction4. However, there are ethical concerns and safety issues associated with the various cell sources used in experimental research, such as bone marrow-derived MSCs, hematopoietic stem/progenitor cells, and cells of embryonic origin38. The use of adipose-derived MSCs (ADMSCs) has distinct advantages, including minimally invasive harvesting and an unlimited supply from in vitro culture. Compared with MSCs derived from bone marrow, ADMSCs exhibit stronger antioxidant, anti-inflammatory, and immunomodulatory functions7,16,39. These properties strengthen the antioxidant system, leading to the removal of free radicals and the self-production of reactive aldehydes, which can increase neuron activity after injury4.
Thus, our hypothesis is that by implanting ADMSCs on a modified scaffold that targets the suppressed oxidative state through enhancement of the antioxidant system by reducing reactive oxygen species (ROS) and LPO, we can preserve the integrity of microanatomical structures and facilitate the recovery of motor function following SCI.
Materials and methods
Synthesis and characterization of liposomes
EA@lip was prepared via the “lipid film hydration and extrusion” method35. The detailed synthesis procedure, along with comprehensive physicochemical characterizations, is provided in the Supplementary File 1 (Sect. 1.5). These include particle size, polydispersity index (PDI), surface charge (zeta potential) measured by Dynamic Light Scattering (DLS), morphological evaluation by Transmission Electron Microscopy (TEM), phospholipid quantification via the Bartlett phosphate assay, and entrapment efficiency (EE) of EA determined spectrophotometrically at 276 nm.
Drug release study
Ellagic acid release from the PCL/f-MWCNT/EA@lip scaffolds was evaluated using the dialysis method in PBS buffer (pH = 7.4, 37 °C) under shaking incubation (100 rpm). Briefly, 5 × 5 cm2 PCL/f-MWCNT/EA@lip scaffolds contained approximately 6.6 mg of entrapped EA were placed in a petri dish and 75 mL of buffer was added. The release of ellagic acid was assessed during 7 days. 1 mL of buffer solution was taken out and replaced with an equal amount of fresh buffer medium at defined time-intervals (0, 0.5, 1, 2, 4, 8, 12, 24, 36, 48, 72, 96, 120, 144, and 168 h). Comparing the experimental results and calibration curve in the UV/Vis spectrophotometer measurement (UV-2600, Shimadzu, Japan) at a wavelength of 276 nm, the quantity of EA in the reserved samples was ascertained. The initial EA concentration in the buffer was approximately 88 µg/mL.
Electrospinning and fabrication of the scaffolds
The production of nanofibers was achieved via the electrospinning technique, with further information provided in the supplementary file.
Morphology of nanofibers
The morphology and size of the fibers were analyzed via field-emission scanning electron microscopy (FESEM), and the dispersion and morphology of the f-MWCNTs within the fibers were assessed via transmission electron microscopy (TEM). More information on these methods is available in the supplementary file.
Characterization of the scaffolds
Mechanical properties
Microtensile testing was performed using a TA Plus machine (USA) equipped with a 100 N load cell at a crosshead speed of 10 mm/min. Rectangular scaffold samples (5 × 1 cm, n = 5) were tested in dry conditions. The initial gauge length was set to 3 cm, and the ultimate tensile strength and elongation were recorded (n = 5).
Chemical characterization
Fourier transform infrared spectroscopy (FTIR) and attenuated total reflection (ATR) were used to assess chemical composition. The f-MWCNT spectrum was acquired using the KBr disk method, while PCL-based samples were evaluated using ATR-FTIR (Thermo Nicolet Avatar 370, USA) within a scanning range of 400–4000 cm⁻¹ at 4 cm⁻¹ resolution.
Hydrophilicity
Hydrophilicity of the scaffolds was assessed by measuring the water contact angle (WCA) using an optical contact angle apparatus (JC2000A, China). A 3 µL drop of deionized water was placed on scaffold samples (1 × 1 cm), and images were captured at 1, 5, and 10 s (n = 3). ImageJ software was used for analysis.
Degradation and swelling behavior
To evaluate in vitro biodegradation, sterilized scaffolds (5 × 5 cm, n = 3) were incubated in PBS (pH 7.4, 40 mL) at 37 °C. Weight loss was measured over 56 days at weekly intervals using Eq. (1).
$${text{Weight loss }}left( % right){text{ }} = {text{ }}left[ {left( {{text{W}}0 – {text{W1}}} right)/{text{ W}}0} right]{text{ }} times {text{ 1}}00$$
(1)
To evaluate the water absorption and hydrophilicity of each group, the swelling ratio (SR) of the PCL and PCL/f-MWCNT scaffolds were tested (n = 3). After 1, 3, 5, 7, 12, 16, 20 and 24 h of immersion in PBS solution (pH = 7.4), the dry weight and the wet weight of the scaffolds were assessed to evaluate the swelling ratio (SR) of the scaffolds. The following equation was used to get the SR% (2):
$${text{SR }}left( % right){text{ }} = left[ {left( {{text{W2}} – {text{W}}0} right)/{text{W}}0} right]{text{ }} times {text{ 1}}00$$
(2)
W0 is the weight of dried samples before soaking and W2 is the weight of soaked samples.
2.6. In vitro biological and biochemical assessments
The ADMSCs used in this study were originally isolated from human subcutaneous adipose tissue obtained from healthy female donors who underwent aesthetic liposuction procedures. The cells were provided by the Matin Laboratory (Ferdowsi University of Mashhad, Iran)40.The cells were used at passage 4 and cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco) supplemented with 10% fetal bovine serum at 37 °C in humidified air with 5% CO2. After they reached confluence, the ADMSCs were passaged with 0.25% trypsin, and the cells at passage 4 were used for the following experiments.
MTT and 1,1-diphenyl-2-picrylhydrazyl (DPPH) assays were used to determine the half-maximal inhibitory concentration (IC50) for the cytotoxicity and radical scavenging of EA and EA@lip (n = 3). For both assays, the following concentrations were tested: 7.18, 14.3, 28.7, 57.5, 115, 230, and 460 µg/ml. The optimal concentration of EA was subsequently determined on the basis of the concentrations that resulted in the highest survival rates and maximum radical scavenging rates at 1, 3, and 7 days after treatment. Further information can be found in the supplementary file 1.
The MTT assay was employed to assess cell viability and proliferation by seeding cells on the samples on days 1, 3, and 7 (n = 3). The attachment of cells on the samples was evaluated via DAPI staining, which relies on the ability to stain intact DNA on day 3 (n = 3). The expansion and connection of cells with the scaffolds were examined via FE-SEM on day 3 (n = 3). Further information can be found in the supplementary file 1.
Biochemical parameters for measuring OS, such as LPO, reactive oxygen species (ROS), total antioxidant capacity (TAC), and total thiol molecules (TTM), were examined by exposing scaffolds containing 1 × 104 ADMSCs to H2O2 for 24 h. The control groups for all the assays consisted of ADMSCs cultured on plates with or without H2O2. Prior to H2O2 treatment, the ADMSCs were cultured in the scaffolds and plates for three days (n = 3). More information can be found in the supplementary file 1.
In vivo studies
Surgical procedure
Sixty female Wistar rats (weighing 250–300 g) were obtained from the Animal House Center of the Faculty of Medical Sciences, Mashhad University of Medical Sciences (MUMS). All rats were housed in separate cages under a 12-hour light-dark cycle, with water and food provided ad libitum. Animal welfare was monitored, and all surgical procedures were conducted in accordance with the institutional guidelines of Mashhad University of Medical Sciences (MUMS) under ethical approval (IR. MUMS. REC.1399.571). An intraperitoneal (IP) injection of a mixture containing xylazine (5 mg/kg) and ketamine (100 mg/kg) was administered to anesthetize the rats. The dorsal skin was shaved and disinfected with betadine solution. A midline incision was made, followed by T9 laminectomy using a rongeur. The exposed dura mater was incised longitudinally and retracted. Afterward, a 1 mm section of dorsal hemisection spinal tissue between the T8 and T9 vertebral levels was removed using microdissection scissors and a 22-gauge needle. Before implantation, the PCL/f-MWCNT scaffold was sterilized by UV exposure for one hour. Scaffolds with or without ADMSCs and EA@lip were implanted into the lesion site, and muscle and skin were closed separately using absorbable vicryl sutures. The dimensions of the implanted scaffolds were 6 × 3 mm, and the volume of EA@lip was 50 µl. The animals were divided randomly into six groups as follows (n = 10/group):
- (a)
Sham-operated group: Laminectomy without SCI (referred to as the sham group).
- (b)
SCI without treatment (referred to as SCI).
- (c)
SCI with PCL/f-MWCNTs without cells or liposomes (referred to as PC).
- (d)
Laminectomized rats receiving scaffolds containing EA@lip (referred to as PC/N). In this group, EA@lip at a concentration of 57.5 µg/ml was applied to the scaffold surface for three days. Additionally, half an hour prior to implanting the scaffold onto the spinal cord, liposomes with the same concentration and volume were also applied.
- (e)
SCI with PCL/f-MWCNT scaffolds containing ADMSCs (referred to as PC/C). For a period of three days, 104 cells were seeded on the scaffold before grafting.
- (f)
SCI with PCL/f-MWCNT/EA@lip scaffolds containing ADMSCs (referred to as PC/N/C). In this group, after the scaffold was covered with liposomes, the cells were seeded onto it for three days. Then, half an hour before grafting, EA@lip at the same concentration and volume was placed on the scaffold. Further information about postoperative care and behavioral assessment can be found in supplementary file 1.
Assessment of OS markers
During the second week post-surgery, biochemical analyses were conducted to assess OS. This involved measuring LPO and TTM levels in the blood of the animals (n = 6–8). More information about these investigations can be found in the supplementary file 1.
Gene expression analysis
At the end of the 6-week experimental period, the remaining animals in each group were euthanized under deep anesthesia, followed by transcardial perfusion with 10% neutral buffered formalin. Real-time reverse transcription polymerase chain reaction (RT‒PCR) was used to assess the mRNA expression of particular genes (n = 5). Additional information regarding the procedures can be found in the supplementary file 1 (Table S1).
Tissue fixation and histological Preparation
After the animals were sacrificed, tissue blocks were prepared for stereological examination using cresyl violet (CV) and Luxol fast blue (LFB) staining. Quantitative evaluations involved analyzing viable neurons in the ventral and dorsal horn of the gray matter, as well as determining the axon density percentage in the dorsal column (n = 5). More information about these examinations can be found in the supplementary file 1.
Statistical analysis
The experimental data were analyzed via GraphPad Prism v 9.0, and the data are presented as the means ± standard deviations (SDs). Unpaired t-tests were used for fiber diameter and mechanical strength comparisons; two-way ANOVA with Tukey’s test was applied for time-based behavioral and contact angle data; and Brown-Forsythe and Welch ANOVA with Dunnett’s T3 multiple comparisons test was used for in vivo biochemical, histochemical and gene expression data; one-way ANOVA with Tukey’s post hoc test was used for rest of in vitro assays. A significance threshold of p < 0.05 was used throughout.
Results
Morphology of the scaffolds
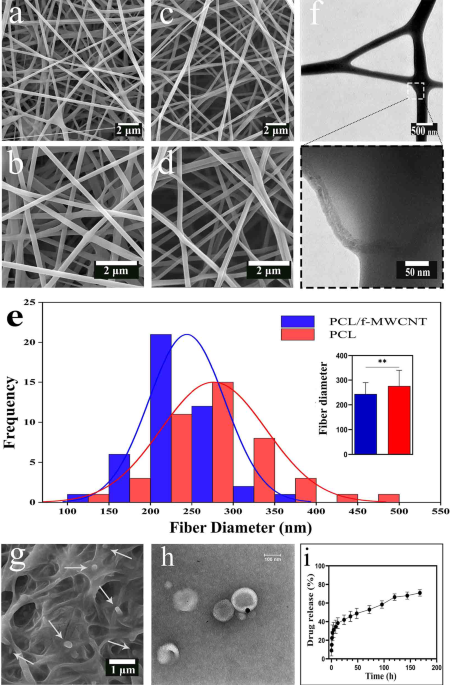
The FE-SEM images of the PCL and PCL/f-MWCNT scaffolds (Fig. 1a-d) indicated that both electrospun scaffolds were composed of randomly oriented, bead-free and uniform nanofibers. The surface of the nanofibers was smooth, and no aggregates of MWCNTs were observed on the surface of the loaded scaffolds. This confirmed the homogeneous distribution of the f-MWCNTs within the PCL nanofibers. The mean diameter of the PCL nanofibers was 276 ± 63 nm. The addition of f-MWCNTs to PCL fibers decreased the average fiber diameter to 243 ± 45 nm (Fig. 1e). Figure 1f shows TEM images of the MWCNTs embedded within the composite fibers. It can be clearly observed that the f-MWCNTs are well embedded along the PCL nanofibers, in which the MWCNTs appear darker than the PCL matrix. Figure 1g illustrates the application of liposomes on the scaffold surface, showing a consistent and thin layer of the liposome solution on the fibers, with no signs of aggregation. TEM images of EA@lip revealed that the liposomes had approximately spherical shape with a homogeneous size of around 100 nm (Fig. 1h), consistent with their physicochemical characterization (Table S2). An in vitro release study of EA from liposome-coated scaffolds is illustrated in Fig. 1i. The release of EA from liposomes occurred rapidly and in a burst-like manner within the first 12 h. However, the release rate then decreased sharply and gradually reached 70% after 7 days.

FE‒SEM micrographs of (a and b) PCLs and (c and d) PCL/f-MWCNTs. (e) Histogram showing the fiber diameter distribution for both PCL and PCL/f-MWCNT scaffolds; the inset displays the calculated average fiber diameters. (f) TEM image of PCL/f-MWCNT fibers. Asterisks indicate statistically significant differences (P < 0.01). (g) FE-SEM image of the PCL/f-MWCNT/EA@lip scaffold surface. (h) TEM images of EA-loaded liposomes, showing a nearly spherical morphology with a uniform size of approximately 100 nm. (i) Cumulative release profile of ellagic acid from the PCL/f-MWCNT/EA@lip scaffold over time. A rapid, burst release was observed during the first 12 h, followed by a markedly slower release phase, reaching approximately 70% by day 7.
Inhibiting OS and enhancing the antioxidant defense system with PCL/f-MWCNT/EA@lip/ADMSC implantation following SCI
SCI led to a significant increase in MDA levels and a decrease in TTM (p < 0.0001 and p < 0.0001, respectively) compared with those in the sham group after 2 weeks (Fig. 2a). Compared with the SCI group, the implantation of nanofibers alone did not alter the levels of MDA or TTM. However, the PC/N, PC/C, and PC/N/C groups showed significant reductions in MDA levels (p < 0.05, p < 0.05, and p < 0.001, respectively) and increases in TTM levels (p < 0.001, p < 0.05, and p < 0.0001, respectively) compared with the SCI group. Furthermore, the EA@lip and ADMSC-treated groups exhibited significantly lower MDA levels compared with the PC, PC/N, and PC/C groups (p < 0.001, p < 0.05, and p < 0.05, respectively). However, no significant differences were observed in TTM levels among the treatment groups.

(a) Concentrations of malondialdehyde (MDA) and total thiol molecules (TTM) in serum samples collected two weeks after spinal cord injury (SCI). (b) Relative mRNA expression levels of COX2 and GPx1 in spinal cord tissue six weeks post-SCI. Data are presented as the means ± standard deviations (n = 5). Asterisks denote significant differences compared with the SCI group, # indicates differences compared with the Sham group, $ indicates significant differences compared with the PC group, and + indicates significant differences compared with the PC/N/C group.
To evaluate the impact of dorsal hemisection on spinal cord oxidative status, we conducted a qPCR analysis (Fig. 2b). Our findings revealed that COX2 expression was significantly higher in the SCI group than in the sham group (p < 0.01), whereas GPx1 expression was significantly reduced following SCI (p < 0.001). Interestingly, COX2 expression significantly decreased after nanofiber implantation in the PC/N, PC/C, and PC/N/C groups (p < 0.05, p < 0.05, and p < 0.01, respectively), whereas nanofibers alone did not induce any change in this gene. Moreover, the reduction in COX2 expression was significantly greater in the PC/N/C and PC/N groups compared with the PC group (p < 0.01 and p < 0.05, respectively). Compared with the SCI group, only the groups that contained cells presented significantly increased GPx1 expression levels, which were restored to levels comparable to those of the sham group.
Boosting neuronal activity and histochemical integrity in Gray matter Horns with PCL/f-MWCNT/EA@lip/ADMSCs following SCI
The neuroprotective effects of the scaffolds following SCI were investigated through LFB and CV staining of the anterior and posterior horns of the spinal cord (Fig. 3a and c). The PC/N, PC/C, and PC/N/C groups exhibited an improved tissue matrix in the anterior horn and reduced cavity formation in the host tissue. Notably, the PC/N/C group showed no cyst formation in the anterior horn, in contrast to the other SCI groups. Histological analysis revealed that rats in the SCI group had fewer neurons in the anterior horns of the gray matter within a 3 mm block surrounding the injury center (Fig. 3b). Compared with SCI group, scaffold treatment significantly increased the neuronal density in the anterior horn across all treatment groups. Moreover, the number of viable neurons in the PC/N/C group was significantly higher than that in the PC group (p < 0.05). The histological analysis also showed extensive structural damage in laminae I–IV of the posterior horn of the gray matter in both the SCI and PC groups (Fig. 3d). In contrast, the PC/N, PC/C, and PC/N/C groups demonstrated improved matrix integrity and neuron preservation in the posterior horn. Nevertheless, matrix reconstruction in lamina I was not well restored in the PC/N group. Compared with the PC and SCI groups, the PC/N/C group exhibited the greatest degree of neuronal preservation in the posterior horn (p < 0.001 for both), with only a slight difference compared to the sham group (p < 0.05).

(a) Histological sections of the spinal cord six weeks post-SCI showing cresyl violet/Luxol fast blue staining. % indicates cyst formation and vascular rupture. The red arrow denotes a representative viable neuron in the anterior horn. (b) Stereological analysis of the percentage of viable neurons in the anterior horn, compared with the sham group. (c) Histological sections of laminae I–IV in the dorsal horn of the gray matter at six weeks post-SCI. The numbers 1–4 correspond to laminae I–IV, respectively. The black arrow marks a representative viable neuron in the posterior horn. (d) Stereological quantification of viable neurons in the posterior horn, compared with the sham group. (e) Relative mRNA expression levels of Slc17a6 and Slc17a7 in spinal cord tissue six weeks following SCI. Data are presented as the means ± standard deviations (n = 5).
To assess neuronal excitability in the posterior and anterior horns, expression levels of Slc17a6 and Slc17a7 were measured (Fig. 3e). Dorsal hemisection led to a significant reduction in the expression of both genes in the SCI group compared with the sham group (p < 0.001 and p < 0.05, respectively). However, the PC/N, PC/C, and PC/N/C groups showed significantly increased expression of both genes compared with the SCI group (p < 0.05, p < 0.05, and p < 0.01 for Slc17a6 and p < 0.05, p < 0.05, and p < 0.05 for all groups for Slc17a7). Notably, the scaffold alone group did not show any significant changes in the expression of these genes compared to the SCI group.
Revitalizing motor function after SCI: A PCL-based platform enhances myelination in posterior limb pathways
The hind limb motor function of the animals after SCI was assessed using the BBB behavioral test over a period of six weeks to evaluate the impact of scaffold implantation. Additionally, the test was conducted one day post-surgery to confirm successful SCI induction and associated posterior limb paralysis (Fig. 4a). The results revealed that all the animals, except those in the sham group, exhibited severe posterior limb impairment one day after surgery. During the first week, the PC/N and PC/N/C groups demonstrated faster recovery of motor function compared with the other groups. After the third week, the recovery slope for the PC/N and PC/N/C groups decreased and plateaued, whereas the performance recovery speed in the PC/C group remained unchanged. The scaffold alone group showed minimal improvements in hindlimb function. By the end of the sixth week, the PC/N/C group exhibited the greatest improvement in motor performance.

(a) Graphs showing the results of the Basso, Beattie, and Bresnahan (BBB) behavioral test and bladder capacity scores at six weeks post-spinal cord injury (SCI; n = 10). (b) Histological sections of the dorsal column stained with cresyl violet/Luxol fast blue. Numbers 1, 2, and 3 denote the gracile fasciculus, dorsal corticospinal tract, and postsynaptic dorsal column pathway, respectively. % indicates cyst formation and vascular rupture. (c) Stereological quantification of axon density in the dorsal column, expressed as a percentage relative to the sham group. (d) Relative mRNA expression levels of the myelin basic protein (MBP) gene in spinal cord tissue six weeks post-SCI. Data are presented as the means ± standard deviations (n = 5).
Evaluation of bladder emptying, as part of the functional recovery, revealed that all dorsal hemisection groups initially had low bladder capacity scores (Fig. 4a). However, beginning in the second week, the PC/N/C group demonstrated improved bladder function, achieving complete emptying by the third week. Other groups reached normal bladder function by the fifth week.
Photomicrographs in Fig. 4b show spinal cord sections from the postsynaptic dorsal column pathway, dorsal corticospinal tract, and gracile fasciculus, which were used to assess myelination. SCI caused severe damage to these dorsal pathways. Compared with the SCI group, treatment with nanofibers alone led to improved myelin staining and a significant increase in axon density (p < 0.05, Fig. 4c). However, in the PC group, the specific pathways could not be clearly distinguished. Treatment in the PC/N, PC/C, and PCNC groups preserved the dorsal myelin structure and enabled pathway differentiation. These groups also exhibited significantly increased axon density compared with the SCI group (p < 0.01, p < 0.01, and p < 0.0001, respectively), and the EA@lip-treated groups were significantly different from the PC group (p < 0.05 for PC/N and p < 0.01 for PC/N/C). Moreover, vacuole formation and vascular disruption were notably reduced in the PC/N/C group compared with the PC/N and PC/C groups. However, no significant differences in axon density were observed among the PC/N, PC/C, and PC/N/C groups.
Compared with the sham group, MBP mRNA expression—a key marker of myelin integrity—was significantly reduced in the SCI group (p < 0.001, Fig. 4d). Treatment with the PCL/f-MWCNT scaffold alone partially restored MBP transcription (p < 0.05 vs. SCI), but it remained significantly lower than in the sham group (p < 0.01). Intervention in the PC/C, PC/N, and PC/N/C groups fully restored MBP expression to levels comparable to the sham group, and were significantly different from the PC group (p < 0.01). Among these, the PC/N/C group demonstrated the strongest effect, showing greater differences than the SCI group (p < 0.001), while the PC/N and PC/C groups also showed significant but lower effects (p < 0.01). The utilization of the PCL/f-MWCNT/EA@lip/ADMSC platform effectively protected against tissue matrix degradation in the dorsal column and resulted in increased axonal density and MBP expression. These positive outcomes have the potential to enhance myelin formation and ultimately improve motor function in rats with SCI.
Discussion
The increase in OS following SCI, coupled with the activation of inflammatory cells, leads to a loss of antioxidant capacity12. During the early phase of secondary damage, the surge in OS promotes the expression of COX2 via the NF-κB signaling pathway41, resulting in the destruction of basement membranes and neural structures3. The upregulation of COX2 is associated with the nerve tissue degradation and an increase in LPO of neuronal membranes14, which, in turn, produces toxic byproducts such as 4-HNE and MDA. These lipid peroxidation products initiate secondary cascades that impair cellular homeostasis and further suppress the expression of key antioxidant enzymes such as GPx and superoxide dismutase (SOD)42. Therefore, we detected the levels of MDA, a cytomembrane oxidation product, and TTM as important biomarkers of OS to assess whether the PCL/f-MWCNT/EA@lip/ADMSC platform ameliorated secondary cascade damage. Compared with the sham group, elevated blood MDA levels and reduced TTM levels in all experimental groups were associated with lower hindlimb motor function scores up to two weeks post-surgery. These findings suggest that dorsal hemisection SCI induces significant imbalances in mitochondrial antioxidant distribution and function in neurons, contributing to spinal cord dysfunction. Furthermore, while monitoring the animals’ behavioral performance, which stabilized by the sixth week, the SCI groups exhibited a reduction in GPx1 expression and an increase in COX2 expression.
The ability of ADMSCs to increase the expression of endogenous antioxidant enzymes (e.g., GPx1) and reduce inflammatory mediators plays a key role in restoring redox homeostasis43. Likewise, EA, by directly scavenging ROS and modulating signaling cascades such as Nrf2/ARE and MAPK, contributes to the suppression of oxidative and inflammatory pathways44. Nrf2 activation increases the transcription of antioxidant genes, while inhibiting ROS-mediated activation of COX2 and other pro-inflammatory genes45. These molecular mechanisms jointly underlie the therapeutic efficacy of our combinatorial platform.
In accordance with previous studies, the formation of a cellular structure depends on a an optimal balance between the strength and plasticity of the polymer matrix46. In the context of SCI, the use of a scaffold carrier with high mechanical strength is essential to support cellular functions and create a stable three-dimensional (3D) matrix35. A study conducted by Wu et al.47 demonstrated that f-MWCNTs with carboxylic acid groups exhibited more favorable mechanical properties than nonfunctionalized MWCNTs, primarily due to their improved dispersion within PCL fibers. The findings of our study indicate that the f-MWCNTs were uniformly dispersed along the PCL fibers, resulting in an increase in the ultimate tensile strength (UTS), Young’s modulus, and cell adhesion in the composite fibers compared with pure PCL fibers (Fig S 1–2). In accordance with Świętek et al.48, the cells were well spread on a high-UTS scaffold with prominent and well-developed focal adhesions.
Hierarchical fibrous morphology is highly important for synthetic neural scaffolds, as this structure mimics the architecture of the native ECM49. Preclinical models of SCI have demonstrated that PCL composite fiber implants are effective in preserving tissue integrity23,50,51. Additionally, the degree of cell adhesion depends on fiber diameter52. The reduced fiber diameter of the PCL/f-MWCNTs compared with that of the neat PCL scaffold is attributed to the higher conductivity of the electrospinning solution53, as other parameters (such as the solution composition, applied voltage, nozzle-to-collector distance, and flow rate) were kept constant. The results of the present study indicate that the obtained fiber diameter of both scaffolds fall within the acceptable range for mimicking the native ECM in the central nervous system (CNS)54. However, smaller fiber diameters provide a greater surface area, offer more sites for cell attachment, and exhibit superior tensile properties55, a condition that is clearly observed in the PCL/f-MWCNT scaffolds. In vitro studies have also shown that the incorporation of CNTs into PCL fibers enhances their electrical conductivity, which is crucial for nerve tissue regeneration56,57. However, in vivo results indicate that the PCL/f-MWCNT scaffold alone lacks regenerative capacity in our SCI model, suggesting that it does not significantly influence the behavior of endogenous neural stem cells or contribute to the repair of primary damage within the studied timeframe. Furthermore, our findings confirm that the modulatory effects of PCL scaffolds on SCI are primarily due to the substrate optimization for the delivery of therapeutic agents and cells, rather than any direct involvement of endogenous stem cells in the repair process50,51.
Based on the FE-SEM images, the scaffold alone does not provide a suitable environment for cell niche formation, growth, and adhesion. The ECM plays a crucial role in cell adhesion, proliferation, and migration22. Therefore, the initial step toward improving the OS environment is to enhance cell adhesion to the culture substrate21. The in vitro results indicate that the incorporation of EA@lip into the scaffold enhances its hydrophilicity (Fig S1) and promotes stem cell attachment (Fig S2). This finding is consistent with that of Mohammadi et al.58, who demonstrated that the WCA of poly-L-lactic acid scaffolds decreased from hydrophobic (130°) to completely hydrophilic (0°) upon the addition of similar liposomal components. Additionally, liposome incorporation significantly increased the OH stretching peak in the FTIR spectrum (Fig S1), which correlates with the WCA results. The molecular mechanism by which EA enhances the antioxidant system involves both direct and indirect pathways. EA can directly scavenge free radicals via hydrogen atom transfer and electron donation due to its multiple hydroxyl groups. Indirectly, it can activate the Nrf2 signaling pathway, which upregulates antioxidant genes30,32. Nrf2 binds to the antioxidant response element (ARE) in the promoter regions of these genes, promoting cytoprotective responses against oxidative and electrophilic stress45. Moreover, EA has been shown to inhibit the NF-κB pathway, thereby reducing the transcription of pro-inflammatory genes such as COX2, TNF-α, and IL-1β59.
The encapsulation of antioxidant drugs in liposomes improves their stability, solubility, bioavailability, and circulation time, while also reducing their toxicity and side effects60. Our findings demonstrate that, over time, encapsulated EA exhibits stronger antioxidant activity than its free form does (Table S3). Moreover, drug encapsulation was associated with a higher cell survival and enhanced antioxidant potential compared to the free form (Table S3). These results suggest that treatment with PCL/f-MWCNT/EA@lip increases TTM levels and reduces MDA levels. However, the MTT assay on day 1 showed no significant difference between the control group and the PCL/f-MWCNT/EA@lip scaffold group. As observed in the drug release profile, the release of EA occurred in a burst-like manner on the first day, which may explain the low cell proliferation at that time point.
Under inflammatory conditions, glial cells become activated and express elevated levels of COX2; these cells contribute to ECM degradation and damage the lipid membranes of neurons and oligodendrocytes61. In contrast, ADMSCs exert anti-inflammatory effects through paracrine secretion of bioactive factors and exosomal transfer of regulatory miRNAs, which suppress pro-inflammatory cytokines and enhance antioxidant defenses62. Our results showed that ADMSCs increased the levels of TTM and GPx1 while reducing LPO and COX2 expression in vivo. This supports the concept that MSCs can modulate redox-sensitive transcription factors to restore homeostasis16. Furthermore, while EA reduced LPO and MDA levels31 it did not significantly upregulate GPx1 expression in our model. This finding may reflect distinct but complementary mechanisms of action between ADMSCs and EA. EA primarily acts by directly scavenging radicals and inhibiting COX2, whereas ADMSCs activate endogenous enzymatic pathways through cell signaling. These data highlight that COX2 suppression is a shared target of both agents, and its inhibition represents a key strategy for attenuating secondary injury in SCI. In light of these antioxidant and anti-inflammatory findings, understanding the molecular signaling events following SCI becomes critical.
Following SCI, the exacerbation of OS not only disrupts redox homeostasis but also activates intracellular signaling cascades such as the NF-κB and MAPK pathways, which upregulate pro-inflammatory mediators including COX2, TNF-α, and IL-1β41,63. These molecular events intensify lipid peroxidation and glial activation, ultimately impairing neuronal survival and axonal integrity. The current study suggests that the antioxidant-rich environment provided by EA@lip, particularly when combined with ADMSCs, may interfere with these detrimental cascades by both scavenging ROS and attenuating pro-inflammatory signaling. This dual modulatory role is evidenced by the suppression of COX2 expression and restoration of GPx1 levels in vivo, indicating a shift toward an anti-inflammatory and neuroprotective phenotype. Such molecular interventions are crucial for mitigating secondary damage and facilitating remyelination and synaptic preservation, which were corroborated by histological and gene expression analyses.
Controlling extensive damage to neurons and oligodendrocytes after SCI is crucial for coordinating motor function recovery and spinal cord regeneration. Although strategies to prevent gliosis have been successful in limiting the spread of neuronal death and secondary damage64, the main challenge remains in preserving functional neurons and promoting remyelination in the perilesional area. The proteins Slc17a7 and Slc17a6, encoding vesicular glutamate transporters VGLUT1 and VGLUT2, respectively, are vital for excitatory neurotransmission. Both are downregulated after SCI, likely due to excitotoxicity, axonal degeneration, and oxidative stress1,6. Enhancing their expression may facilitate synaptic plasticity and support the maintenance of neuronal function.
Our results show that incorporation of EA@lip and ADMSCs into PCL/f-MWCNT scaffolds not only enhanced the expression of Slc17a6/7 but also increased MBP, a myelin-associated marker, suggesting the preservation of excitatory neurons and promotion of oligodendrocyte integrity. Part of the observed improvement in our SCI model following PCL/f-MWCNT/EA@lip treatment may be attributed to the pro-myelinating effects of EA. This property has previously been reported in various models of spinal cord demyelination28,29. Our earlier studies also showed that scaffolds containing EA@lip can enhance neuronal plasticity and electrophysiological function in regions remote from the injury site35. One likely mechanism is the anti-inflammatory and antioxidant action of EA, which suppresses microglial activation via COX2 downregulation, thereby mitigating chronic demyelination5. In line with this, our current results show that EA@lip treatment induced the expression of Slc17a6, Slc17a7, and MBP, while limiting neuronal and oligodendrocyte loss, suppressing COX2 expression, and preventing SCI-induced demyelination. Ardah et al. attributed the neuroprotective effects of EA in an Alzheimer’s disease model to its potent antioxidant properties34. Our findings strongly support the role of EA’s antioxidant activity in mitigating secondary injury cascades following SCI. However, we acknowledge that additional mechanisms—independent of antioxidant defense—may also contribute to the neuroprotective effects of EA.
Although MSC differentiation into neurons or glia has been demonstrated on PCL scaffolds51, our study did not evaluate lineage-specific differentiation. Instead, we hypothesize that the modulation of oxidative and inflammatory microenvironments by the scaffold components facilitated neuronal survival and remyelination. By suppressing ROS and LPO, and enhancing TAC and TTM, the treatment limited secondary neurodegeneration, a key pathological feature of SC4,65. Our findings demonstrated that the antioxidant properties of ADMSCs play a key role in mitigating both inflammation and OS, as evidenced by decreased MDA and COX2 levels. These effects contributed to increased axon density, enhanced survival of excitatory and native neurons in both the posterior and anterior horns, and improved remyelination, as indicated by elevated MBP expression. Additionally, the upregulation of GPx1 expression represents a notable advantage of the PC/C group compared with the PC/N group, highlighting the critical contribution of cell-based therapy to the observed neuroprotective effects.
Our results confirm that early burst release of EA during the acute phase (< 48 h) coincides with a critical window of ROS accumulation and inflammatory signaling activation, including TNF-α, IL-1β, and COX2 expression10. The initial release may have counteracted these pathways, reducing tissue damage. Meanwhile, the sustained release profile observed during the subacute and early chronic phases likely prevented delayed neuroinflammation and reactivation of glial responses, as described in chronic SCI models15. This pharmacokinetic profile enhanced axonal density, neuronal survival in both horns, and MBP expression, resulting in superior outcomes in the PC/N/C group.
Despite these promising results, a limitation of our study was that mechanical properties were only tested under dry conditions. In vivo, hydration significantly alters scaffold compliance and mechanical behavior. Future studies should consider evaluating the mechanical performance of scaffolds under physiological wet conditions to provide a more accurate representation of their functional integrity post-implantation. Lastly, while the in vitro degradation profile suggested slow hydrolysis, in vivo results indicated complete scaffold degradation. This might be due to enhanced macrophage infiltration, hydrophilic modification by f-MWCNTs, and enzymatic degradation within the injured tissue. However, despite these positive features, full functional recovery was not achieved, indicating the need for advanced bioactive materials with tailored degradation, controlled spatiotemporal release of neuroprotective agents, and multi-modal therapeutic potential. The synergistic action of antioxidant-loaded nanofibers and stem cells contributes to the preservation of spinal architecture, reduction of oxidative damage, suppression of COX2-driven inflammation, and enhancement of remyelination, offering a promising foundation for future translational approaches in SCI repair.
Conclusion
The objective of this study was to develop a biomimetic scaffold incorporating ADMSCs and enabling the controlled release of EA to improve tissue structure and mitigate oxidative cascade damage in SCI. The EA@lip exhibited favorable particle size and zeta potential. Moreover, the encapsulation of EA enhanced its radical scavenging capacity (increased IC₅₀) and significantly reduced cytotoxicity. We successfully fabricated PCL/f-MWCNT nanofibrous scaffolds with interconnected, bead-free networks via electrospinning. The integration of f-MWCNTs resulted in a decrease in fiber diameter, while their alignment along the nanofibers contributed to enhanced mechanical strength, swelling ratio, and accelerated degradation of the PCL matrix. The hydrophilicity of the scaffolds was markedly improved by the incorporation of EA@lip. Among all tested formulations, the PCL/f-MWCNT/EA@lip scaffold exhibited the highest levels of ADMSC attachment and proliferation, as demonstrated by MTT assay, DAPI staining, and FE-SEM analysis. In vivo, this platform effectively regulated oxidative stress by reducing LPO and COX2 levels and enhancing the antioxidant defense system, as evidenced by increased TTM and GPx1 levels. Improvement in behavioral performance was supported by histological and cellular analyses, along with upregulated MBP expression. Furthermore, the increased expression of Slc17a6 and Slc17a7 contributed to the restoration of neuronal excitability in the posterior and anterior horns of the spinal cord. The scaffold’s high hydrophilicity, mechanical robustness, biodegradation profile, and excellent biological performance—including antioxidant and anti-inflammatory properties, and support for stem cell proliferation, adhesion, and differentiation—collectively suggest that this platform is a promising candidate for future applications in neural tissue engineering.
Data availability
The raw data supporting the findings and conclusions of this study are available from the corresponding author upon reasonable request.
References
-
Fukuoka, T. et al. Neurod4 converts endogenous neural stem cells to neurons with synaptic formation after spinal cord injury. iScience [Internet]. ;24(2):102074. Available from: https://www.sciencedirect.com/science/article/pii/S2589004221000420 (2021).
-
Romanelli, P. et al. Enhancing functional recovery through intralesional application of extracellular vesicles in a rat model of traumatic spinal cord injury. Front. Cell. Neurosci. 15, 795008 (2022).
-
Xi, J. et al. Tetrahydrocurcumin protects against spinal cord injury and inhibits the oxidative stress response by regulating FOXO4 in model rats. Exp. Ther. Med. 18 (5), 3681–3687 (2019).
-
Park, S. S. et al. Functional recovery after spinal cord injury in dogs treated with a combination of matrigel and neural-induced adipose-derived mesenchymal stem cells. Cytotherapy 14 (5), 584–597 (2012).
-
Macchi, M. et al. Mature oligodendrocytes bordering lesions limit demyelination and favor Myelin repair via Heparan sulfate production. Elife 9, e51735 (2020).
-
Coyoy-Salgado, A. et al. Gene expression and locomotor recovery in adult rats with spinal cord injury and plasma-synthesized polypyrrole/iodine application combined with a mixed rehabilitation scheme. Front. Neurol. 14, 1124245 (2023).
-
Kim, C. et al. Mesenchymal stem cell transplantation promotes functional recovery through MMP2/STAT3 related astrogliosis after spinal cord injury. Int. J. Stem Cells. 12 (2), 331–339 (2019).
-
McTigue, D. M., Tripathi, R., Wei, P. & Lash, A. T. The PPAR gamma agonist Pioglitazone improves anatomical and locomotor recovery after rodent spinal cord injury. Exp Neurol [Internet]. ;205(2):396–406. Available from: https://www.sciencedirect.com/science/article/pii/S0014488607000891 (2007).
-
Beck, K. D. et al. Quantitative analysis of cellular inflammation after traumatic spinal cord injury: evidence for a multiphasic inflammatory response in the acute to chronic environment. Brain [Internet]. ;133(2):433–47. Available from: https://doi.org/10.1093/brain/awp322 (2010).
-
Kong, X. & Gao, J. Macrophage polarization: a key event in the secondary phase of acute spinal cord injury. J. Cell. Mol. Med. 21 (5), 941–954 (2017).
-
Alizadeh, A., Dyck, S. M. & Karimi-Abdolrezaee, S. Traumatic spinal cord injury: an overview of pathophysiology, models and acute injury mechanisms. Front. Neurol. 10, 282 (2019).
-
Visavadiya, N. P., Patel, S. P., VanRooyen, J. L., Sullivan, P. G. & Rabchevsky, A. G. Cellular and subcellular oxidative stress parameters following severe spinal cord injury. Redox Biol [Internet]. ;8:59–67. Available from: https://www.sciencedirect.com/science/article/pii/S2213231715300264 (2016).
-
Cao, J. et al. ROS filter coating scaffold protects 3D mesenchymal stem cell spheroids for dual-phase treatment of spinal cord injury. Chem. Eng. J. 462, 142192 (2023).
-
Adibhatla, R. M. & Hatcher, J. F. Lipid oxidation and peroxidation in CNS health and disease: from molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 12 (1), 125–169 (2010).
-
Allison, D. J. & Ditor, D. S. Immune dysfunction and chronic inflammation following spinal cord injury. Spinal Cord [Internet]. ;53(1):14–8. Available from: https://doi.org/10.1038/sc.2014.184 (2015).
-
Chen, S., He, Z. & Xu, J. Application of adipose-derived stem cells in photoaging: basic science and literature review. Stem Cell. Res. Ther. 11, 1–15 (2020).
-
Labombarda, F. et al. Effects of progesterone on oligodendrocyte progenitors, oligodendrocyte transcription factors, and myelin proteins following spinal cord injury. Glia [Internet]. ;57(8):884–97. Available from: https://doi.org/10.1002/glia.20814 (2009).
-
Vismara, I., Papa, S., Rossi, F., Forloni, G. & Veglianese, P. Current options for cell therapy in spinal cord injury. Trends Mol. Med. 23 (9), 831–849 (2017).
-
Popovich, P. G. et al. A reassessment of a classic neuroprotective combination therapy for spinal cord injured rats: lps/pregnenolone/indomethacin. Exp. Neurol. 233 (2), 677–685 (2012).
-
Li, X., Wang, C., Yang, S., Liu, P. & Zhang, B. Electrospun pcl/mupirocin and chitosan/lidocaine hydrochloride multifunctional double layer nanofibrous scaffolds for wound dressing applications. Int. J. Nanomed. ;5287–5299. (2018).
-
Wu, R. X., Ma, C., Liang, Y., Chen, F. M. & Liu, X. ECM-mimicking nanofibrous matrix coaxes macrophages toward an anti-inflammatory phenotype: Cellular behaviors and transcriptome analysis. Appl Mater Today [Internet]. ;18:100508. Available from: https://www.sciencedirect.com/science/article/pii/S2352940719306274 (2020).
-
Cai, S. et al. Recent advance in surface modification for regulating cell adhesion and behaviors. Nanotechnol. Rev. 9 (1), 971–989 (2020).
-
Wang, Z. et al. A functionalized self-assembling peptide containing E7 and YIGSR sequences enhances neuronal differentiation of spermatogonial stem cells on aligned PCL fibers for spinal cord injury repair. Theranostics 12 (17), 7567 (2022).
-
Díaz, E., Sandonis, I. & Valle, M. B. In vitro degradation of Poly (caprolactone)/nHA composites. J. Nanomater. 2014, 185 (2014).
-
Patel, K. D. et al. Coating biopolymer nanofibers with carbon nanotubes accelerates tissue healing and bone regeneration through orchestrated cell- and tissue-regulatory responses. Acta Biomater [Internet]. ;108:97–110. (2020). Available from: https://www.sciencedirect.com/science/article/pii/S1742706120301446
-
Yousefi Talouki, P., Tehrani, P. & Shojaei, S. The relationship between thermomechanical properties with morphology in PCL/PHBV/MWCNT biodegradable nanocomposites with application in neural tissue engineering. Razi J. Med. Sci. 27 (11), 25–38 (2021).
-
Mahmoodi, M. et al. Highly osteogenic and mechanically strong nanofibrous scaffolds based on functionalized multi-walled carbon nanotubes-reinforced electrospun keratin/poly(ε-caprolactone). Mater Today Commun [Internet]. 27, 102401. Available from: https://www.sciencedirect.com/science/article/pii/S2352492821003937 (2021).
-
Busto, R. et al. Ellagic acid protects from myelin-associated sphingolipid loss in experimental autoimmune encephalomyelitis. Biochim Biophys Acta – Mol Cell Biol Lipids [Internet]. , 1863(9), 958–67. Available from: https://www.sciencedirect.com/science/article/pii/S1388198118301021 (2018).
-
Kiasalari, Z. et al. Ellagic acid ameliorates neuroinflammation and demyelination in experimental autoimmune encephalomyelitis: Involvement of NLRP3 and pyroptosis. J Chem Neuroanat [Internet]. , 111, 101891. Available from: https://www.sciencedirect.com/science/article/pii/S0891061820301605 (2021).
-
Khanduja, K. L., Gandhi, R. K., Pathania, V. & Syal, N. Prevention of N-nitrosodiethylamine-induced lung tumorigenesis by ellagic acid and quercetin in mice. Food Chem Toxicol [Internet]. 37(4), 313–8. Available from: https://www.sciencedirect.com/science/article/pii/S0278691599000216 (1999).
-
Aslan, A. et al. The preventive effect of ellagic acid on brain damage in rats via regulating of Nrf-2, NF‐kB and apoptotic pathway. J. Food Biochem. 44 (6), e13217 (2020).
-
García-Niño, W. R., Ibarra-Lara, L., Cuevas-Magaña, M. Y., Sánchez-Mendoza, A. & Armada, E. Protective activities of ellagic acid and urolithins against kidney toxicity of environmental pollutants: A review. Environ Toxicol Pharmacol [Internet]. 95, 103960.Available from: https://www.sciencedirect.com/science/article/pii/S1382668922001533 (2022).
-
Sanadgol, N. et al. Ellagic acid ameliorates cuprizone-induced acute CNS inflammation via restriction of microgliosis and down-regulation of CCL2 and CCL3 pro-inflammatory chemokines. Cell. Mol. Biol. 62 (12), 24–30 (2016).
-
Ardah, M. T., Bharathan, G., Kitada, T. & Haque, M. E. Ellagic acid prevents dopamine neuron degeneration from oxidative stress and neuroinflammation in MPTP model of Parkinson’s disease. Biomolecules [Internet], 10(11), 1–17. Available from: https://pubmed.ncbi.nlm.nih.gov/33172035 (2020).
-
Abroumand Gholami, A. et al. Effect of polycaprolactone/carbon nanotube scaffold implantation along with liposomal ellagic acid in hippocampal synaptogenesis after spinal cord injury. Nanomedicine J [Internet].10(3), 197–209. Available from: https://nmj.mums.ac.ir/article_22560.html (2023).
-
Cardoso, R. V., Pereira, P. R., Freitas, C. S. & Paschoalin, V. M. F. Trends in drug delivery systems for natural bioactive molecules to treat health disorders: the importance of nano-liposomes. Pharmaceutics 14 (12), 2808 (2022).
-
Chi, Y. et al. Redox-sensitive and hyaluronic acid functionalized liposomes for cytoplasmic drug delivery to osteosarcoma in animal models. J Control Release [Internet]. ;261:113–25. Available from: https://www.sciencedirect.com/science/article/pii/S0168365917306922 (2017).
-
Cofano, F. et al. Mesenchymal stem cells for spinal cord injury: current options, limitations, and future of cell therapy. Int. J. Mol. Sci. 20 (11), 2698 (2019).
-
Abdelbaset-Ismail, A. et al. Transplantation of adipose-derived mesenchymal stem cells ameliorates acute hepatic injury caused by nonsteroidal anti-inflammatory drug diclofenac sodium in female rats. Biomed Pharmacother [Internet].155, 113805. Available from: https://www.sciencedirect.com/science/article/pii/S0753332222011945 (2022).
-
Naderi-Meshkin, H. et al. Injectable hydrogel delivery plus preconditioning of mesenchymal stem cells: exploitation of SDF‐1/CXCR4 axis toward enhancing the efficacy of stem cells’ homing. Cell. Biol. Int. 40 (7), 730–741 (2016).
-
Iordanis, V. et al. The impact of nuclear factor kappa B on the response of microglia in spinal cord injuries. Cureus ;17(2). (2025).
-
Zhang, C. et al. Research progress of antioxidants in oxidative stress therapy after spinal cord injury. Neurochem Res. 48 (12), 3473–3484 (2023).
-
Shen, Y. J., Huang, Y. C. & Cheng, Y. C. Advancements in Antioxidant-Based therapeutics for spinal cord injury: A critical review of strategies and combination approaches. Antioxidants 14 (1), 17 (2024).
-
Yu, C., Xu, Y., Zhao, M., Song, P. & Yu, J. New insights into mechanism of ellagic acid alleviating arsenic-induced oxidative stress through MAPK/keap1-Nrf2 signaling pathway response, molecular Docking and metabolomics analysis in HepG2 cells. Ecotoxicol. Environ. Saf. 285, 117029 (2024).
-
Xiao, C. et al. tong, jun, Nrf2 signaling pathway: focus on oxidative stress in spinal cord injury. Mol Neurobiol. 62(2). 2230–49. (2025).
-
Li, J. et al. Morphologies and electromagnetic interference shielding performances of microcellular epoxy/multi-wall carbon nanotube nanocomposite foams. Compos Sci Technol [Internet].129, 70–8. Available from: https://www.sciencedirect.com/science/article/pii/S0266353816301324 (2016).
-
Wu, D., Zhang, Y., Zhang, M. & Yu, W. Selective Localization of Multiwalled Carbon Nanotubes in Poly(ε-caprolactone)/Polylactide Blend. Biomacromolecules [Internet].10(2), 417–24. Available from: https://doi.org/10.1021/bm801183f (2009).
-
Świętek, M. et al. Carbon nanotube/iron oxide hybrid particles and their PCL-based 3D composites for potential bone regeneration. Mater Sci Eng C [Internet].104, 109913. Available from: https://www.sciencedirect.com/science/article/pii/S0928493118337639 (2019).
-
Hu, Y. et al. Scaffolds with anisotropic structure for neural tissue engineering. Eng. Regen. 3 (2), 154–162 (2022).
-
Wang, P. et al. Novel cytokine-loaded PCL-PEG scaffold composites for spinal cord injury repair. RSC Adv. 10 (11), 6306–6314 (2020).
-
Babaloo, H. et al. PCL/gelatin nanofibrous scaffolds with human endometrial stem cells/schwann cells facilitate axon regeneration in spinal cord injury. J. Cell. Physiol. 234 (7), 11060–11069 (2019).
-
Tian, F. et al. Quantitative analysis of cell adhesion on aligned micro-and nanofibers. J. Biomed. Mater. Res. Part. A. 84 (2), 291–299 (2008).
-
Liao, G. Y. et al. Electrospun aligned PLLA/PCL/functionalised multiwalled carbon nanotube composite fibrous membranes and their bio/mechanical properties. Compos Sci Technol [Internet].72(2). 248–55.Available from: https://www.sciencedirect.com/science/article/pii/S0266353811004015 (2012).
-
Jain, D., Mattiassi, S., Goh, E. L. & Yim, E. K. F. Extracellular matrix and biomimetic engineering microenvironment for neuronal differentiation. Neural Regen Res. 15 (4), 573 (2020).
-
Akrami-Hasan-Kohal, M., Tayebi, L. & Ghorbani, M. Curcumin-loaded naturally-based nanofibers as active wound dressing mats: morphology, drug release, cell proliferation, and cell adhesion studies. New. J. Chem. 44 (25), 10343–10351 (2020).
-
Zhou, Z. et al. Effective nerve cell modulation by electrical stimulation of carbon nanotube embedded conductive polymeric scaffolds. Biomater. Sci. 6 (9), 2375–2385 (2018).
-
Hu, X. et al. Electric conductivity on aligned nanofibers facilitates the transdifferentiation of mesenchymal stem cells into Schwann cells and regeneration of injured peripheral nerve. Adv. Healthc. Mater. 9 (11), 1901570 (2020).
-
Mohammadi, M. et al. Fabrication of hybrid scaffold based on hydroxyapatite-biodegradable nanofibers incorporated with liposomal formulation of BMP-2 peptide for bone tissue engineering. Nanomedicine Nanotechnology, Biol Med [Internet].14(7). 1987–97.Available from: https://www.sciencedirect.com/science/article/pii/S154996341830474X (2018).
-
El-Shitany, N. A., El-Bastawissy, E. A. & El-desoky, K. Ellagic acid protects against carrageenan-induced acute inflammation through Inhibition of nuclear factor kappa B, inducible cyclooxygenase and Proinflammatory cytokines and enhancement of interleukin-10 via an antioxidant mechanism. Int. Immunopharmacol. 19 (2), 290–299 (2014).
-
Isuru Wijesiriwardana, C. & Jacobsen, N. W. R. Encapsulation of antioxidants in liposomes: a review focusing on mechanisms of antioxidant activity and health benefits. Nutr J. ;20(1). (2021).
-
Zhao, W. et al. Astrocytic Nrf2 expression protects spinal cord from oxidative stress following spinal cord injury in a male mouse model. J. Neuroinflamm.. 19 (1), 1–14 (2022).
-
El-Sawah, S. G. et al. Investigation of the antioxidant defensive role of both AD-MSCs and BM-MSCs in modulating the alteration in the oxidative stress status in various STZ-diabetic rats’ tissues. Biocell 45 (6), 1561 (2021).
-
Lin, F., Gu, H. & yun, He, W. MAPK signaling pathway in spinal cord injury: Mechanisms and therapeutic potential. Exp Neurol [Internet]. ;383:115043. Available from: https://www.sciencedirect.com/science/article/pii/S0014488624003698 (2025).
-
Anjum, A. et al. Spinal cord injury: pathophysiology, multimolecular interactions, and underlying recovery mechanisms. Int. J. Mol. Sci. ;21(20). (2020).
-
Fakhri, S. et al. Effects of polyphenols on oxidative stress, inflammation, and interconnected pathways during spinal cord injury. Oxid Med Cell Longev. ;2022. (2022).
Acknowledgements
The authors would like to thank Mashhad University of Medical Sciences for the financial support of this work [grant NO. 990834]. We also sincerely thank Prof. Hossein Haghir and Prof. Reza Kazemi Oskuee for their valuable scientific guidance and early-stage consultation during the conceptual development of the study.
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
All methods were carried out in accordance with relevant guidelines and regulations. All animal experiments were conducted and reported in compliance with the ARRIVE guidelines. Ethics approval was obtained from the institutional guidelines of Mashhad University of Medical Sciences (MUMS), with a COD number (IR. MUMS. REC.1399.571).
Consent for publication
All authors have given their consent to publish the results.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abroumand Gholami, A., Rahmani, S., Moharreri, P. et al. Liposomal ellagic acid enhances the regenerative potential of ADMSC-laden nanofibrous PCL scaffolds in a rat model of spinal cord injury. Sci Rep 15, 30202 (2025). https://doi.org/10.1038/s41598-025-15789-w
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-15789-w