
Introduction
The increased threat of antibiotic resistance, recently dubbed “The Silent Pandemic”, is a rapidly growing public health concern1. Wastewater treatment utilities are one of the main recipients, reservoirs, and resources of antibiotic-resistant bacteria (ARB) and antibiotic resistance genes (ARGs)2,3. Additionally, wastewater utilities receive a wide variety of antibiotics, metals, and other antimicrobials that can promote the horizontal transfer of ARGs4,5. In wastewater treatment trains, a large fraction of the ARB, ARGs, and mobile genetic elements end up in the biosolids6.
In the United States, 31% of biosolids are used as amendments to agricultural soils to enhance soil fertility (e.g., nutrient cycling) and bolster plant defenses against pathogens5,7,8,9. This practice is regulated under the U.S. EPA’s Part 503 Rule, which establishes standards for pathogen and pollutant reduction10. With over 2.3 million dry metric tons applied annually to cropland, pasture, and rangeland, biosolids land application is widespread across agricultural regions7. Its prevalence highlights the need to evaluate long-term microbial and public health impacts across diverse soil systems. Environments receiving biosolids have been shown to contain a large diversity of ARB (including human pathogens) and ARGs and are considered reservoirs for antibiotic resistance propagation6,11,12,13. Additionally, biosolids contain diverse microorganisms, with large prevalences of Proteobacteria and Bacteroidetes which contain pathogenic species14,15. Accordingly, there are concerns that biosolids land application may increase the prevalence of human pathogens in soils and further propagate clinical antibiotic resistance.
Among the human pathogens of greatest concern are the ESKAPE pathogens: Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp., named for their ability to ‘escape’ the effects of antimicrobial agents. These organisms are characterized by multidrug resistance and widespread clinical prevalence, making them central to global antimicrobial resistance surveillance efforts16. ESKAPE pathogens have been detected in wastewater and receiving environments (e.g., rivers, lakes, agricultural soils that receive treated wastewater effluent or biosolids), including multidrug-resistant Acinetobacter and Pseudomonas aeruginosa, and methicillin-resistant Staphylococcus aureus (MRSA)17,18. While recent research has reported on ESKAPE taxa in wastewater influent and effluent19,20, there is no comprehensive understanding of the fate of these pathogens in environments amended with biosolids.
In a broader context, longitudinal studies have examined the impact of biosolids amendment on the soil microbiome and ARG abundance. Many of these studies rely on targeted analyses, such as qPCR, to characterize the soil resistome21,22,23. While qPCR analysis provides an accurate quantitative assessment of the abundances of targeted ARGs, they are limited in their ability to examine the breadth of diversity in the resistome24. Metagenomic approaches have been applied to characterize microbiome and resistome dynamics in soils impacted by manure, wastewater irrigation, heavy metals, and antibiotics25,26,27,28,29,30. These studies demonstrate the utility of high-throughput sequencing for identifying microbial shifts and resistance profiles in complex environments.
While metagenomic studies have examined biosolids-amended soils, most have focused on the biosolids themselves or the effects of treatment processes, rather than their influence on soil microbiomes and resistomes during crop cultivation31,32,33,34. A recent review of high-throughput sequencing approaches highlighted the need for replicated, longitudinal studies to assess how biosolids amendments shape soil microbiomes and resistomes in agricultural systems35. To address this gap, we conducted a replicated greenhouse experiment using shotgun metagenomics to test the hypotheses that biosolids application (1) alters soil microbial community composition, (2) increases the abundance and diversity of ARGs and virulence factors (VFs), and (3) enriches for clinically relevant pathogens, including members of the ESKAPE group, during carrot cultivation. Carrots were selected as a model crop due to their direct soil-root contact and relevance to human consumption risks. While root vegetables are typically excluded from biosolids-amended fields under the U.S. EPA’s Part 503 regulations10, their inclusion in this study provides a conservative scenario for evaluating potential microbial transfer and public health implications. Additionally, we applied network analysis to identify statistically significant co-occurrences between microbial taxa and ARGs, providing insight into potential host associations and ecological interactions. Sampling time points included weeks 0, 6, and 11, corresponding to key stages of carrot cultivation, pre-planting baseline, mid-growth, and harvest, allowing us to capture temporal shifts in microbial composition, ARG and VF dynamics, and pathogen persistence. These methodological advances offer a more comprehensive understanding of biosolids’ influence on soil microbial ecology and antimicrobial resistance surveillance.
Results
Microbial community diversity and profile
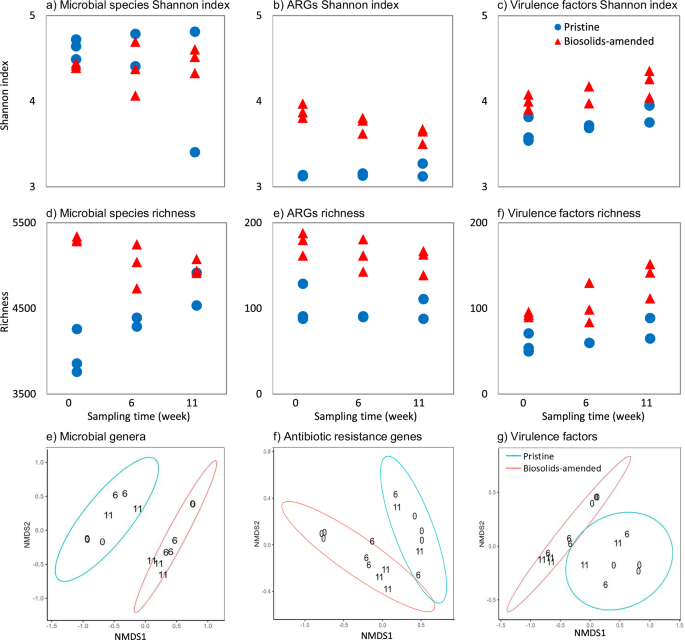
Biosolids generally increase microbial biomass and alter community composition36. In our replicated, controlled, greenhouse study, the impact of biosolids amendment on the microbiome diversity was identified using alpha and beta diversity indices (Fig. 1). This analysis was conducted to test our first hypothesis, which posited that biosolids amendment would alter soil microbial community composition. We report that while there were no significant differences in Shannon indices—which integrate both species richness and evenness—between pristine and biosolids-amended soils, the latter had significantly higher microbial species richness. Including all time points, there were no significant differences between the Shannon indices of microbial genera in the pristine (4.5 ± 0.2) and biosolids-amended (4.4 ± 0.1) soils (one-sided t-test, p > 0.05; Fig. 1a). These Shannon indices are generally lower than reports on agricultural soils from Arizona and California (6.0-7.3) but were comparable to those from biosolids from conventional wastewater treatment facilities (3.4−5.3)37,38. The observed increase in richness in biosolids-amended soils (5,310.7 ± 16.5) compared to pristine soils (3,959.7 ± 153.3; one-sided t-test, p <0.001; including all the time points; Fig. 1d) indicates a greater number of unique taxa, even though dominance by a subset of taxa may have reduced overall evenness. This explains why Shannon diversity remained unchanged despite clear compositional shifts. Over time, this richness declined, suggesting that some bacteria from the biosolids did not naturalize into the soil. In a previous field study with short- and long-term applications of alkaline-treated biosolids, soils with high biosolids application rates showed lower alpha-diversity indices (Chao1 richness, Simpson evenness, and Shannon diversity), whereas lower application rates resulted in increased indices39. Another controlled mesocosm study demonstrated similar diversity indices between biosolids-amended soil and the controls after 28 days of crop growth40. Similar results of the gradual return from microbiome enrichment in biosolids-amended soils to comparable conditions with control soil have been previously documented41.

Alpha diversity indices (a−c) Shannon index and (d−f) richness for (a) and (d) microbial species, (b) and (e) antibiotic resistance genes (ARGs), and (c) and (f) virulence factors, and ordination of the Bray-Curtis dissimilarity between the (e) microbial genera, (f) ARGs, and (g) virulence factors in pristine and biosolids-amended soils at weeks 0, 6, and 11 in a replicated greenhouse study. In e-g, labels indicate the sampling time (i.e., weeks 0, 6, and 11) and the ellipses denote the 95% confidence level for each treatment condition (i.e., pristine and biosolids-amended soils).
The relative abundances of microbial phyla and genera in each sample were determined to understand the impact of biosolids amendment and cultivation time on the microbial community structure. While overall patterns were broadly similar across treatments, Bacteroidetes were statistically enriched in biosolids-amended soils, while Proteobacteria remained the most abundant phylum in both pristine and amended soils. Across all sample types, there was an average of 35.3 ± 1.7% of reads unmapped to any phyla in the database. Of the mapped reads, the most abundant phylum in both pristine and biosolids-amended samples was Proteobacteria throughout the cultivation period (Fig. S1). Bacteroidetes was the second most abundant phyla in all samples at week 0. Previous studies have also observed these phyla to be dominant in both pristine soils and gut microbiomes14,15. Biosolids-amended samples (n = 9) contained a statistically larger enrichment of the phylum Bacteroidetes (14.5 ± 1.1%) as compared to pristine soil samples (n = 7; 6.3 ± 1.0%; one-sided t-test, p <0.01). Although the absolute change in relative abundance was modest, the consistent enrichment of Bacteroidetes across time points suggests a biologically relevant shift. Bacteroidetes remained the second most abundant phylum in biosolids-amended samples for the duration of the study, while in pristine samples, Actinobacteria was the second most dominant phylum during weeks 6 and 11. Because Bacteroidetes are relatively prevalent in both human guts and natural soils, the enrichment of this phylum after soil amendment throughout the cultivation period could be due to their ability to secrete a wide range of carbohydrate-active enzymes capable of targeting diverse and complex glycans present in the soil42.
While short-term studies show significant changes in microbial communities, long-term impacts may vary36,40,43. We report here distinct differences between pristine and amended soils at the genera level after application. These differences diminished over the 11-week cultivation period, indicating a lasting yet gradually stabilizing impact of biosolids on soil microbial communities. At the genera level, we identified Massilia, Ralstonia, Mucilaginibacter, Nocardioides, Streptomyces, Sphingomonas, and Janthinobacterium (clusters A and C) as the dominant genera in pristine soils at Week 0 (Fig. 2a). Biosolids amendment, however, altered the microbial communities so that the dominant genera at week 0 included Dechloromonas, Sulfuritalea, Flavobacterium, Candidatu, Accumulibacter, and Pseudomonas (Clusters E and F). The enrichment of Dechloromonas in the biosolids amended soil is consistent with the prevalence of these genera in biosolids and its association with the human microbiome44,45. By the end of the study (i.e., Week 11), the abundance of these notable genera became more similar between pristine and biosolids-amended soils. The main differentiating genera between the two treatments at Week 11 were Ralstonia (2.25-3.01% in pristine soils vs. 0.26-0.95% in biosolids-amended soils), Gemmatirosa (1.12-2.95% vs. <0.1%), and Gemmatimonas (0.98-2.37% vs. 0%)—all enriched in pristine soils—compared to Rhodanobacter (12.14-24.72% in biosolids-amended soils vs. 0-0.11% in pristine soils), Dyella (3.51-4.34% vs. 0.11-0.13%), and Thermomonas (2.23-3.06% vs. 0.14-0.51%), which were more abundant in biosolids-amended soils. These distinguishing genera between the two treatments at different cultivation points can explain the statistical dissimilarities between the pristine and biosolids-amended soils (PERMANOVA, p <0.001; Fig. 1e). The “legacy effects” of biosolids amendments in other studies are marked by increased abundances of specific bacterial groups, such as Streptomycetaceae and Clostridiaceae, which can form resilient, desiccation-resistant spores22.

Heatmaps of (a) relative abundance (%) of the top 30 most abundant genera, and (b) abundance of 30 most abundant antibiotic resistance gene (ARG) subtypes (log (gene copies/16S rRNA)) detected in at least three samples in pristine and biosolids-amended soil samples collected at weeks 0, 6, and 11 in a greenhouse study. The dendrogram was generated using Euclidean distance.
Antibiotic resistance genes profile and diversity
Biosolids amendments enrich the soil resistome, which encompasses the collection of ARGs present in the soil11,12,13,41. While a few studies have utilized high-throughput DNA sequencing to explore the impact of biosolids amendment on soil resistome31,32,33,34, there is a lack of comprehensive longitudinal replicated shotgun metagenomics studies in this area35. In our replicated, controlled, greenhouse study, we employed shotgun metagenomics to assess the effects of biosolids amendment on the soil resistome. This analysis was conducted to test our second hypothesis, which posited that biosolids amendment would increase the abundance and diversity of ARGs in soil. Our findings revealed that biosolids-amended soils exhibited significantly higher diversity (Fig. 1b) and richness (Fig. 1e) of ARG subtypes compared to pristine soils (one-sided t-test, p <0.001), with 165 unique ARG subtypes (genotypes) in biosolids-amended samples versus 98 in pristine soils. Additionally, the resistome of the pristine and biosolids-amended soils was statistically distinct (PERMANOVA, p <0.001, Fig. 1f). Higher diversity of ARGs can confer ecological resilience to microbial communities, allowing them to adapt to changes in environmental conditions, including antibiotic exposure46. However, higher diversity also raises concerns about the potential for the horizontal gene transfer of these resistance genes, which could contribute to the spread and persistence of antibiotic resistance in agricultural settings47.
Resistance to 19 different classes of antibiotics, including multidrug resistance, was observed, with multidrug resistance being the most prevalent (in all but one sample, sample S014, pristine soil on Week 0) (Fig. S3). Across all samples, the next most abundant classes of resistance were vancomycin (0.021 ± 0.004 gene copies/16S rRNA), bacitracin (0.018 ± 0.002 gene copies/16S rRNA), fosmidomycin (0.013 ± 0.002 gene copies/16S rRNA), and sulfonamides (0.009 ± 0.002 gene copies/16S rRNA). Comparing pristine and biosolids-amended samples, vancomycin resistance was the only class of resistance more abundant in pristine soils (an average relative abundance of 0.035 ± 0.006 gene copies/16S rRNA copy in pristine samples and 0.010 ± 0.003 gene copies/16S rRNA copy in biosolids-amended samples; Fig. 2b). Actinobacteria and Proteobacteria, identified as prevalent phyla in our study, have been previously recognized as potential hosts of vancomycin resistance genes. Findings demonstrate that biosolids amendment increased the abundance of resistance genes for several antibiotic classes, including sulfonamide (sul1 and sul2), tetracycline (tetC and tetX2), fosmidomycin (rosA and rosB), macrolides (ermF), and Class A beta-lactams, an impact that persisted throughout the 11 weeks (Fig. 2b). Similar to these results, the application of biosolids has been associated with increased relative abundances of tetracycline, sulfonamide, and macrolide resistance genes in soil13,48,49. Additionally, previous studies have highlighted increases in resistance genes to aminoglycosides, rifamycin, and fluoroquinolones13,31. The detection of beta-lactam resistance genes in biosolid-amended soils likely reflects their known prevalence in activated sludge, where selective pressures from antibiotic residues and high microbial density promote their persistence and horizontal gene transfer50. Biosolids derived from activated sludge systems can retain these resistance determinants, and land application may facilitate their introduction into soil microbial communities6. It is evident that, in addition to the impact on resistome diversity, biosolids amendment changes the classes of resistance genes in soils.
The temporal impact of biosolids amendments was evident, with notable shifts in ARG subtype abundances over the cultivation period. At Week 0, the most abundant ARG subtypes in pristine soils included vancomycin resistance gene vanR and bacitracin bacA, while in biosolids-amended soils, bacitracin bacA, sulfonamide sul1, and multidrug qacE∆1 were dominant. By Week 11, the resistome of pristine soils remained relatively unchanged, whereas biosolids-amended soils showed a shift to multidrug mexF, fosmidomycin rosA, and a multidrug transporter gene. These findings demonstrate the significant (PERMANOVA, p <0.001; Fig. 1f) and lasting impact of biosolids amendment on the soil resistome, with some effects persisting throughout the 11 weeks. Persistence of biosolids-derived ARG subtypes and bacteria harboring these genes has been previously documented21,51,52,53,54. The concentration of the resistant bacteria and associated resistance genes tends to decline over time53,54.
Virulence factors profile and diversity
Biosolids-amendment increased the soil’s alpha and beta diversities of VFs, which stayed higher throughout the 11 weeks of the study. These analyses were performed to evaluate our second hypothesis, which predicted that biosolids amendment would increase the abundance and diversity of VFs in soil. A total of 285 VFs were annotated across all samples. Including all time points, biosolids amendment resulted in significantly higher alpha diversity of VFs measured by the Shannon index (4.11 ± 0.05) compared to pristine soils (3.72 ± 0.05; one-sided t-test, p <0.001, Fig. 1c). The soil richness of VFs was also significantly higher in biosolids-amended (110.9 ± 8.2) compared to pristine samples (64.1 ± 4.9; one-sided t-test, p <0.001, Fig. 1f). The difference in VFs between pristine and biosolids-amended soils (i.e., treatments) was statistically larger than the differences observed within treatments (PERMANOVA, p <0.001; Fig. 1g), which demonstrates the difference between the two treatments lasted throughout the study.
ESKAPE pathogens profile
The amendment of biosolids led to increased prevalence of ESKAPE pathogens in soil. Here, we tested our third hypothesis, which posited that biosolids amendment would enrich clinically relevant pathogens, including members of the ESKAPE group, during carrot cultivation. Across all time points (i.e., Weeks 0, 6, and 11), biosolids-amended soils (0.015 ± 0.003%) had higher relative abundances of all six ESKAPE pathogens compared to pristine soils (0.007 ± 0.001%; p <0.01) (Fig. S2). At the time of planting, biosolids-amended soils showed higher abundances of each ESKAPE pathogen except for Staphylococcus aureus, which had comparable relative abundance in pristine (0.0033%) and biosolids-amended (0.0035%) soils. At the time of harvest, biosolids amendment still had significantly higher enrichment of the six ESKAPE pathogens.
The amendment of biosolids to soil resulted in a significant increase in the prevalence of all six ESKAPE pathogens, posing a substantial threat to human health and exerting a lasting impact throughout the 11-week cultivation period of our study. Relative to pristine samples at harvest, biosolid-amended soils harbored higher abundances of the six pathogens. Enterococcus faecium was enriched 3.32-fold (soil: 0.0003 ± 0.0002% vs biosolids amended soil: 0.0011 ± 0.0002%; one-sided t-test, p = 0.009), Staphylococcus aureus 1.56-fold (soil: 0.0022 ± 0.0008% vs biosolids amended soil: 0.0035 ± 0.0002%; p = 0.088), Klebsiella pneumoniae 1.58-fold (soil: 0.0045 ± 0.0006% vs biosolids amended soil: 0.0071 ± 0.0006%; p = 0.005), Acinetobacter baumannii 3.41-fold (soil: 0.0028 ± 0.0006% vs biosolids amended soil: 0.0096 ± 0.0005%; p <0.001), Pseudomonas aeruginosa 2.64-fold (soil: 0.0214 ± 0.0016% vs biosolids amended soil: 0.0565 ± 0.0032%; p <0.001), and Enterobacter spp. 1.39-fold (soil: 0.0098 ± 0.0008% vs biosolids amended soil: 0.0135 ± 0.0006%; p = 0.002). While ESKAPE pathogens were present and enriched in our study, they remained at a low relative abundance in soil (0.007 ± 0.001% in pristine and 0.015 ± 0.003% in biosolids-amended soils), similar to previously reported data22.
Associations between the microbiome and resistome
Co-occurrence analyses within and between the microbial genera and ARGs (Fig. 3) identified statistically large (p <0.01 and Spearman coefficient > 0.8) associations (i.e., edges) within and between the microbial genera and ARGs (i.e., nodes). In pristine soil, eight clusters of microbial genera with at least three nodes were identified, while all other observed associations contained only two nodes (Fig. 3a). One large cluster included six microbial genera (Solitalea, Sulfuriferula, Algoriphagus, Rufibacter, Saprospira, and Echinicola) and the tetracycline resistance gene tetO. Of these six microbial genera, five belonged to the Bacteroidota phylum while the other one (Sulfuriferula) belonged to Pseudomonadota. A previous study on target ARGs and amplicon-sequenced microbial communities reported co-occurrences of tetracycline resistance genes (e.g., tetG and tetX) with microbial genera belonging to the Bacteroidota phylum (e.g., Parapedobacter, Sphingobacteriaceae, Sphingobacterium, and Flavobacterium) following the land application of sludge composts41.

Associations between the microbiome (at the genus level present in at least three samples) and resistome in (a) pristine and (b) biosolids-amended soils collected at weeks 0, 6, and 11 in a greenhouse study. All associations shown by connections (i.e., edges) represent significant Spearman correlations at p <0.01. Nodes represent microbial genera (gray circles) and antibiotic resistance subtypes (red circles). Edge thickness is uniform and does not represent correlation strength. The layout was optimized to reduce visual congestion.
Additionally, in pristine soils, the co-occurrence associations of two Proteobacteria genera (Undibacterium and Fimbriimonas) with the multidrug resistance gene oprJ and a chloramphenicol transport gene, respectively, were observed (Fig. 3a). Three multidrug resistance pairs were identified including mexD and oprC, ermB and a multidrug transport gene, and oprA and mdtE. Multidrug resistance prevalence was also associated with resistance to bacitracin (bacA) and fosmidomycin (rosB). Tetracycline resistance gene, otrA was associated with the prevalence of vanA gene resisting vancomycin. The sulfonamide resistance sul1 prevalence clustered with the quinolone resistance abaQ. Despite these associations, none of the identified taxa in the clusters were abundant at levels greater than 0.1% in pristine samples.
In biosolids-amended samples, two large and interconnected clusters of microbial genera were found (Fig. 3b). In one of these clusters, all genera were from the Bacteroidota phylum except Sulfuriferula from the Pseudomonadota phylum. The second large microbial cluster contained genera from the Bacteroidota and Pseudomonadota phyla as well as Casimicrobium from the Firmicutes phylum. The Bacteroidota phylum is the primary degrader of complex carbohydrates found in soils42, therefore their presence is likely originated from the soil microbiome. Sulfuriferula (from Pseudomonadota phylum) are sulfur-oxidizing chemolithoautotrophs—organisms that derive energy from chemical reactions of reduced compounds of mineral origin, making soil the preferred environment for them55.
In addition to the two large clusters of microbial genera in biosolids-amended soil, two other clusters of genera contained more than three nodes, one of which included a beta-lactam resistance gene oxa17. Two pairs of genera and ARG subtypes co-occurrences were identified including the multidrug efflux pump sdiA and Caldilinea as well as the multidrug resistance gene mexW and Caulobacter. Two beta-lactam resistance genes ges14 and ges15 and two vancomycin resistance vanA and vanH also co-occurred in biosolids-amended soils. Additionally, the fosmidomycin resistance rosA co-occurred with the rifamycin resistance adp. While many of the genera identified to have co-occurrences had relatively low abundance (<0.1%) in biosolids-amended soils, the genera Caldilinea represented 0.75% of the microbial community (Fig. 3b). Although most co-occurring genera were low in relative abundance, such taxa may still play ecologically meaningful roles, particularly as reservoirs or vectors of resistance genes56. Rare taxa have been shown to disproportionately contribute to functional gene pools and horizontal gene transfer in soil environments57,58. Nonetheless, we acknowledge that some associations may reflect statistical proximity rather than true ecological linkage, and further validation is needed.
Discussion
This study provides the first replicated, longitudinal metagenomic analysis of biosolids-amended soils during crop cultivation, offering a comprehensive assessment of microbial community shifts, resistome enrichment, VF diversity, and ESKAPE pathogen persistence. Using shotgun metagenomics and network analysis, we demonstrated that biosolids amendment significantly increased microbial richness, ARG and VF diversity, and the relative abundance of all six ESKAPE pathogens throughout an 11 week carrot cultivation period. Importantly, our network analysis revealed statistically supported co-occurrences between microbial taxa and ARGs, suggesting potential host associations and ecological interactions. These findings advance current understanding by moving beyond biosolids characterization to assess their dynamic impact on soil microbiomes and resistomes in an agricultural context, addressing a critical gap identified in recent reviews. The integration of replication, temporal sampling, and ecological network analysis represents a novel approach to evaluating biosolids-associated risks to soil and public health.
Our first hypothesis, that biosolids amendment alters soil microbial community composition, was supported by significant increases in microbial richness and genus-level shifts. Taxa such as Rhodanobacter, Dyella, and Thermomonas were enriched in biosolids-amended soil, with changes persisting over time. These shifts likely reflect biosolids-induced changes in soil chemical properties43, particularly by increasing organic matter, which favors nutrient-responsive taxa such as Pseudomonadota22,40,59. Such shifts influence microbial functions, including nutrient cycling and organic matter decomposition22,39. Biosolids also affect microbial metabolism, promoting pathways such as glycolysis and the reductive tricarboxylic acid cycle39. Abiotic factors, such as pH, texture, and contaminants like heavy metals26,40,59,60, further modulate microbial responses. Although these mechanisms were not directly measured in this study, they warrant further investigation to understand long-term ecological impacts.
Our second hypothesis that biosolids amendment increases the abundance and diversity of ARGs and VFs was strongly supported. Biosolids-amended soils exhibited significantly higher diversity and richness of ARG subtypes and VFs compared to pristine controls. Temporal analysis revealed dynamic shifts in ARG subtypes, with biosolids-amended soils transitioning from dominance by sul1, bacA, and qacEΔ1 at Week 0 to mexF, rosA, and multidrug transporter genes by Week 11. These changes were statistically significant and persisted throughout the study. The enrichment of ARGs observed in our study aligns with previous reports on biosolids-amended soils, reinforcing the role of these environments as reservoirs for antimicrobial resistance dissemination. For example, the sulfonamide resistance gene sul1 was detected at an average relative abundance of 0.006 ± 0.002 gene copies per 16S rRNA, comparable to levels reported in lettuce-cultivated soils and biosolids application sites13,48. Multidrug resistance genes such as qacEΔ1 and mexF were similarly enriched, reflecting patterns observed in wastewater treatment systems and biosolids-amended environments32,54. Continued application of biosolids introduces high levels of antibiotics (e.g., tetracyclines, sulfamethoxazole, ciprofloxacin, and trimethoprim)61,62, heavy metals, and ARB, all of which can exert selective pressure even at low concentrations63,64,65. Notably, ARGs may persist in soil longer than antibiotics themselves. For instance, sulfonamide and other resistance genes have been detected in soils near a pharmaceutical industries and public parks irrigated with reclaimed wastewater, even in absence of detectable antibiotic residues66,67. Environmental factors such as soil pH, moisture, salinity, and organic matter also influence both the microbiome and resistome68,69. Changes in these abiotic conditions can induce cross-adaptive responses in the microbial communities. For example, shifts in soil pH can alter heavy metal solubility and affect microbial susceptibility to antibiotics70,71,72. Biosolids amendment can significantly modify these parameters, potentially increasing the minimum inhibitory concentration of resistant bacteria and promoting resistance in previously susceptible strains73,74. In addition to ARGs, biosolids amendment led to sustained enrichment of diverse VFs with potential ecological and clinical relevance. Biosolids contain a wide array of transcriptionally active pathogens, VFs, and ARGs13,31,75, and their co-occurrence increases the potential for horizontal gene transfer, allowing virulence and resistance genes to spread among different microbial taxa. Taken together, these results support our second hypothesis and demonstrate that biosolids amendment significantly enriched the soil resistome, both in terms of ARG subtype diversity and abundance across multiple antibiotic classes. These findings suggest that ARGs are not only environmentally persistent but may also be more resilient than the compounds that select for them. Overall, our results demonstrate that biosolids amendment significantly enriches the soil resistome and virulome, both in terms of diversity and abundance across multiple antibiotic classes and virulence mechanisms. This underscores the importance of integrated surveillance frameworks that consider both agricultural and wastewater contexts to mitigate the spread of clinically resistant and virulent traits.
Our third hypothesis was supported by the observed enrichment of all six ESKAPE pathogens and their sustained presence throughout the 11-week cultivation period. Elevated alpha diversity indices (Shannon index and richness) of ARG subtypes (Fig. 1b and e; p <0.001) and VFs (Fig. 1c and f; p <0.001), along with the increased relative abundance of ESKAPE pathogens (Fig. S2; p <0.01), underscore the concern. This demonstrates the potential for high-risk pathogenic bacteria to acquire VFs and ARGs following the amendment of biosolids in agricultural soils via opportunistic horizontal gene transfer. For example, Pseudomonas aeruginosa, an ESKAPE pathogen, has been shown to be responsible for 11% of virulence gene expression in biosolids-amended soils31. ESKAPE pathogens can acquire ARGs through horizontal gene transfer as they often host ARG-laden mobile elements such as plasmids and prophages76. Similarly, other pathogens can horizontally transfer clinically relevant ARGs to environmental bacteria when exposed to selective pressures, making biosolids-amended soils a potential reservoir for high-risk ARGs and pathogens11. The significance of these findings lies in the comprehensive assessment of ESKAPE pathogens’ prevalence and dynamics in biosolids-amended soils, offering insights into the long-term implications for soil health and the potential for pathogen proliferation.
Network analysis revealed increased microbiome connectivity following biosolids application, with multiple co-occurrences between ARGs and microbial genera. This pattern of enhanced network connectivity has been previously reported in biosolids-amended soils39, and our findings further support this observation. In this study, we identified multiple co-occurrences of ARGs (conferring resistance to tetracyclines, beta-lactams, chloramphenicol, and multidrugs) with microbial genera in both pristine and biosolids-amended soils. These associations may indicate that specific microbial taxa harbor the co-occurring ARGs, as suggested by similar abundance trends across different environmental reservoirs77,78. While many of the genera involved on these co-occurrences were of relatively low abundance ( <0.1%), such rare taxa may still play disproportionately important roles in resistance gene dissemination and ecological function56,57,58. Their presence in biosolids-amended soils suggests that even low-abundance organisms can act as reservoirs or vectors for ARGs, particularly under selective pressures introduced by biosolids. Importantly, while our study did not directly resolve host-ARG linkages, the co-occurrence of ARGs and VFs with ESKAPE pathogens raises the possibility that these clinically relevant taxa may serve as hosts for enriched resistance and virulence traits. Given that ESKAPE pathogens are known to harbor mobile genetic elements such as plasmids and prophages58, their sustained presence in biosolids-amended soils could facilitate horizontal gene transfer and enhance both antibiotic resistance and pathogenicity. Although these associations are statistically robust, they do not confirm functional linkage; rather, they suggest potential ARG hosts that warrant further investigation. Such co-occurrence patterns may reflect ecological proximity, shared niches, or horizontal gene transfer potential, but should be validated using targeted approaches such as metagenome-assembled genomes, single-cell sequencing, or functional assays.
In summary, our study demonstrates that biosolids amendment significantly increases the diversity and richness of microbial species, ARGs and VFs, resulting in compositional and functional shifts in microbial community structure and resistome profiles. The prevalence of ESKAPE pathogens and the co-occurrence of ARGs with specific microbial genera highlight the potential risks associated with biosolids amendment, including the spread of antibiotic resistance and pathogenic bacteria. These findings underscore the need for evidence-based monitoring, risk assessment, and regulation of biosolids land application to safeguard environmental and public health.
The observed shifts in microbial diversity, ARGs, VFs, and ESKAPE pathogen abundance following biosolids amendment likely reflect multiple interacting mechanisms. First, biosolids are rich in organic matter and nutrients, which can stimulate copiotrophic microbial taxa—organisms that thrive in nutrient-rich environments and often harbor resistance traits14,79. Second, biosolids introduce exogenous microbes, including human-associated taxa and pathogens, which may persist or interact with native soil communities80,81. Third, residual contaminants such as antibiotics, heavy metals, and disinfectants present in biosolids can exert selective pressure, favoring resistant strains and promoting horizontal gene transfer82. Finally, biosolids amendment alters soil physicochemical properties, including pH, moisture, and organic carbon, which in turn shape microbial community structure and functional gene profiles14. These mechanisms likely act synergistically to drive the observed changes and should be further explored through targeted chemical analyses, mobilome characterization, and functional assays.
Our study was limited by the relatively short duration of the cultivation period, which may not fully capture the long-term impacts of biosolids on soil health and microbial dynamics. Long-term field studies are needed to understand the persistent effects of biosolids on soil health and microbial communities. In particular, future research should assess the cumulative risks associated with continuous or repeated biosolids application, including the potential buildup of ARGs, VFs, and clinically relevant pathogens over time. While our metagenomic approach enabled broad characterization of resistome and microbiome profiles, it did not permit confident resolution of their genomic context (e.g., plasmid versus chromosomal origin). Future studies incorporating mobilome analysis or long-read sequencing could address this important dimension of ARG mobility. In addition, our co-occurrence analysis was based on statistical correlations and therefore does not provide definitive evidence of ARG host taxa. Higher-resolution approaches such as metagenome-assembled genomes, Hi-C, or single-cell genomics could more directly resolve host-ARG linkages83,84,85,86. Moreover, the use of DNA-based analyses does not distinguish between live and dead bacteria, potentially affecting the accuracy of our assessments. Future research should incorporate RNA-based approaches to evaluate the viability and transcriptional activity of pathogens and resistance genes in biosolids-amended soils. By addressing these limitations, we can develop more effective strategies for managing biosolids in agricultural settings and protecting soil and human health.
Methods
Sample collection
The greenhouse study design has been described in detail previously53. Briefly, the soil for cultivation was collected from a commercial agricultural field with no history of wastewater irrigation or biosolids amendment in the Willamette Valley area of Oregon. The soil was air-dried and passed through a 5 mm sieve. Biosolids were obtained from a municipal wastewater treatment utility in Oregon employing a conventional activated sludge process. Settled sludge was dewatered by belt press to produce Class B biosolids, which contained 14.4% total solids, 6.3 ± 0.2 log10 CFU/g-TS of Escherichia coli, 6.9 ± 0.0 log10 CFU/g-TS of enterococci53. The biosolids were transferred to the laboratory on ice and applied to six replicated pots at a ratio of 70 g per kg of soil. Similarly, six replicated pots of soil—herein referred to as pristine soil—were used as the control treatment. Four germinated carrot seeds were planted in each of the twelve pots and cultivated for 11 weeks, at which time they were harvested. Pots were randomly assigned to greenhouse positions and rotated periodically to minimize spatial effects, with replication implemented at the pot level as previously described53. At each sampling time point (weeks 0, 6, and 11), three pots per treatment were sampled (n = 3 per treatment per time point), rotating which pots were sampled across occasions to distribute sampling across the six pots in each treatment53. Greenhouse conditions were maintained at overnight temperatures near 15 °C and daytime temperatures ranging from 20–26 °C. Growth trays were enclosed with mosquito netting to prevent insect access, and runoff was contained using steel mesh tables and PVC-framed trays. Deionized water was used to irrigate the plants two to three times per week to maintain a soil moisture content of 80-85% total solids.
Soil core samples were collected from the bulk soil using a soil sampler probe (M.K. Rittenhouse & Sons Ltd., St. Catharines, Ontario, Canada) at the time of planting, on week 6 of the study, and at week 11 at the time of harvest. Soil samples were transferred in WhirlPak bags (Nasco, Fort Atkinson, WI) to the lab where they were homogenized by massaging and shaking for 2 min. From each sample, approximately 0.5 g of soil was stored in 50% v/v ethanol at -20 °C for microbial analysis. Genomic DNA was extracted using the FastDNA Spin Kit for Soil (MP Biomedicals, Irvine, CA). Extracted DNA was stored at -20 °C until submission for sequencing.
DNA purification, shotgun sequencing, and quality analysis
The concentration and quality of extracted DNA were measured with a Qubit fluorometer (Thermo Fisher Scientific, Waltham, MA). To improve the quality of low-concentration samples, extracted DNA was purified with a ReliaPrep DNA Clean-up and concentration kit (Promega, Madison, WI). Purified DNA was submitted to the Center for Quantitative Life Sciences at Oregon State University. Sequencing libraries were prepared for all samples using the NexteraXT kit (Illumina, San Diego, CA), except for the initial biosolids-amended samples, which were prepared using PrepX (IntegenX, Pleasanton, CA). The three biosolids-amended samples required the PrepX kit because the BioAnalyzer results indicated DNA quality issues when using the NexteraXT kit. Paired-end (2×150 bp) sequencing reads were generated on an Illumina HiSeq 3000 (Illumina, San Diego, CA). Read quality was assessed with FastQC87. Subsequently, reads were filtered and trimmed using CutAdapt with quality thresholds of 20 and 15 for forward and reserve reads, respectively, and a minimum length of 3688. Reads were again assessed with FastQC to verify quality before subsequent analysis.
Metagenomic analysis
Taxonomic classification of microbial communities was performed using Kaiju89, which translates metagenomic reads into amino acid sequences in all six possible reading frames and identifies maximum exact matches against the National Center for Biotechnology Information (NCBI) microbial reference protein database89. Matched sequences were assigned to taxa based on the NCBI taxonomy, enabling characterization of community structure from protein-level alignments. Pristine soil samples collected prior to biosolids amendment (week 0) were used to establish baseline microbial community composition and resistance gene profiles. These baseline data enabled comparative analysis of microbiome and resistome changes following biosolids application over time. To assess the presence of clinically relevant pathogens, we specifically examined taxa corresponding to ESKAPE pathogens. Taxonomic assignments and abundance estimates for ESKAPE pathogens were derived from Kaiju-based classification of metagenomic reads against the NCBI microbial reference protein database.
Cleaned reads from each sample were then de novo assembled with MEGAHIT v1.2.990 with k-mer values ranging from 59 to 159. Assembled contigs were directly annotated for ARGs using ARGs-OAP v2.0 pipeline, which applies hidden Markov models to identify ARGs type and subtype levels based on the SARG database, a non-redundant reference integrating the Comprehensive Antibiotic Resistance Database (CARD, version 3.2.4) and the Antibiotic Resistance Database (ARDB, v1.1)91. VFs were identified by BLAST + 2.13.0 searches (with default identity and coverage thresholds) against the VF database (VFDB, v6.0) using assembled contigs as input92.
Statistical analysis
To assess the alpha diversity (richness and evenness) within each treatment (pristine soil and biosolids-amended soil at weeks 0, 6, and 11), the Shannon index and richness for each treatment’s microbiome (species level), resistome, and VFs were calculated in R (version 4.2.2) using the Vegan package93. The impacts of biosolids amendment on the resistome and microbiome diversities were determined by comparing the Shannon indices of pristine samples and those amended with biosolids using the student’s t-test. To determine the differences between sample groups in the microbiome (at the genus level), resistome, and VFs, the beta diversity was determined using the Bray-Curtis dissimilarity and the Vegan package. The variances between treatment groups were determined using the permutational multivariate analysis of variance (PERMANOVA) test Adonis in the Vegan package, which reduces the dimensionality of the data to identify statistically significant differences between biosolids-amended and pristine soil. Hierarchical cluster analysis was used to create heatmaps of the microbial phyla and genera (present above 0.1%) as well as ARGs across samples using the Vegan package. In the heatmaps, dendrograms were generated using Euclidean distances.
The relative abundance of ARGs at the subtype level and microbial taxa at the genus level were aggregated. ARG relative abundances were expressed as gene copies per 16S rRNA gene, enabling normalization across samples. Data was filtered to include microbial genera and ARGs present in at least three samples. To determine co-occurrences within and between the microbiome, resistome, and ESKAPE pathogens, pair-wise Spearman correlations were determined. Statistically correlated (p <0.01) ARG subtypes and microbial genera were visualized as a network with the R package igraph94.
Data availability
All sequenced reads can be found in the National Center for Biotechnology Information (NCBI) Sequence Read Archive under Bioproject accession number PRJNA1049878.
References
-
G7 Health Ministers. G7 Health Ministers’ Communiqué. https://www.g7germany.de (2022).
-
Karkman, A., Do, T., Walsh, F. & Virta, M. Antibiotic-resistance genes in waste water. Trend. Microbiol. 26, 220–228 (2018).
-
Pazda, M., Kumirska, J., Stepnowski, P. & Mulkiewicz, E. Antibiotic resistance genes identified in wastewater treatment plant systems—a review. Sci. Total Environ.697, 134023 (2019).
-
Anjali, R. & Shanthakumar, S. Insights on the current status of occurrence and removal of antibiotics in wastewater by advanced oxidation processes. J. Environ. Manag. 246, 51–62 (2019).
-
Wang, J., Chu, L., Wojnárovits, L. & Takács, E. Occurrence and fate of antibiotics, antibiotic resistant genes (ARGs) and antibiotic resistant bacteria (ARB) in municipal wastewater treatment plant: an overview. Sci. Total Environ. 744, 140997 (2020).
-
Munir, M., Wong, K. & Xagoraraki, I. Release of antibiotic resistant bacteria and genes in the effluent and biosolids of five wastewater utilities in Michigan. Water Res. 45, 681–693 (2011).
-
EPA. Basic Information about Sewage Sludge and Biosolids. https://www.epa.gov/biosolids/basic-information-about-biosolids (2023).
-
Stavridou, E. et al. Biosolid-amended soil enhances defense responses in tomato based on metagenomic profile and expression of pathogenesis-related genes. Plants 10, 2789 (2021).
-
Sullivan, T. S., Stromberger, M. E., Paschke, M. W. & Ippolito, J. A. Long-term impacts of infrequent biosolids applications on chemical and microbial properties of a semi-arid rangeland soil. Biol. Fertility Soils 42, 258–266 (2006).
-
EPA. Standards for the Use or Disposal of Sewage Sludge, 40 CFR Part 503; Environmental Protection Agency. https://www.ecfr.gov/current/title-40/chapter-I/subchapter-O/part-503 (2018).
-
Bondarczuk, K., Markowicz, A. & Piotrowska-Seget, Z. The urgent need for risk assessment on the antibiotic resistance spread via sewage sludge land application. Environ. Int. 87, 49–55 (2016).
-
Markowicz, A., Bondarczuk, K., Cycon, M. & Sulowicz, S. Land application of sewage sludge: Response of soil microbial communities and potential spread of antibiotic resistance. Environ. Pollut. 271, 116317 (2021).
-
Yang, L. et al. Application of biosolids drives the diversity of antibiotic resistance genes in soil and lettuce at harvest. Soil Biol. Biochem. 122, 131–140 (2018).
-
Fierer, N., Bradford, M. & Jackson, R. Toward an ecological classification of soil bacteria. Ecology 88, 1354–1364 (2007).
-
Lozupone, C., Stombaugh, J., Gordon, J., Jansson, J. & Knight, R. Diversity, stability and resilience of the human gut microbiota. NATURE 489, 220–230 (2012).
-
Santajit, S. & Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed. Res. Int. 2016, 2475067 (2016).
-
Kozajda, A. & Jezak, K. Occupational exposure to staphylococcus aureus in the wastewater treatment plants environment. Medycyna Pracy 71, 265–278 (2020).
-
Nishiyama, M. et al. Prevalence of antibiotic-resistant bacteria eskape among healthy people estimated by monitoring of municipal wastewater. Antibiotics-Basel 10, (2021).
-
Ramos, B., Lourenço, A., Monteiro, S., Santos, R. & Cunha, M. Metagenomic profiling of raw wastewater in Portugal highlights microbiota and resistome signatures of public health interest beyond the usual suspects. Sci. Total Environ. 946, 174272 (2024).
-
Raza, S., Shin, H., Hur, H. & Unno, T. Higher abundance of core antimicrobial resistant genes in effluent from wastewater treatment plants. Water Res. 208, 117882 (2022).
-
Cerqueira, F. et al. Antibiotic resistance gene distribution in agricultural fields and crops. A soil-to-food analysis. Environ. Res. 177, 108608 (2019).
-
Schlatter, D. C. et al. Biosolids and tillage practices influence soil bacterial communities in dryland wheat. Microb. Ecol. 78, 737–752 (2019).
-
Zhang, Y.-J. et al. Transfer of antibiotic resistance from manure-amended soils to vegetable microbiomes. Environ. Int. 130, 104912–104912 (2019).
-
Liguori, K. et al. Antimicrobial resistance monitoring of water environments: a framework for standardized methods and quality control. Environ. Sci. Technol. 56, 9149–9160 (2022).
-
Chen, C. et al. Effect of antibiotic use and composting on antibiotic resistance gene abundance and resistome risks of soils receiving manure-derived amendments. Environ. Int. 128, 233–243 (2019).
-
Guron, G. K. P., Arango-Argoty, G., Zhang, L., Pruden, A. & Ponder, M. A. Effects of dairy manure-based amendments and soil texture on lettuce- and radish-associated microbiota and resistomes. mSphere 4, 3 (2019).
-
Keenum, I. et al. A framework for standardized qPCR-targets and protocols for quantifying antibiotic resistance in surface water, recycled water and wastewater. Crit. Rev. Environ. Sci. Technol. 52, 4395–4419 (2022).
-
Bougnom, B. P., Thiele-Bruhn, S., Ricci, V., Zongo, C. & Piddock, L. J. V. High-throughput sequencing data and antibiotic resistance mechanisms of soil microbial communities in non-irrigated and irrigated soils with raw sewage in African cities. Data Brief 27, 104638 (2019).
-
Brown, L.P. et al. Responses of the soil bacterial community, resistome, and mobilome to a decade of annual exposure to macrolide antibiotics. Appl. Environ. Microbiol. 88, e00316-22 (2022).
-
Salam, L. B. Unravelling the antibiotic and heavy metal resistome of a chronically polluted soil. 3 Biotech. 10, 238 (2020).
-
D’Angelo, E. M. Diversity of virulence and antibiotic resistance genes expressed in Class a biosolids and biosolids-amended soil as revealed by metatranscriptomic analysis. Lett. Appl. Microbiol. 76, ovad097 (2023).
-
Guo, J., Li, J., Chen, H., Bond, P. L. & Yuan, Z. Metagenomic analysis reveals wastewater treatment plants as hotspots of antibiotic resistance genes and mobile genetic elements. Water Res. 123, 468–478 (2017).
-
Han, I. & Yoo, K. Metagenomic profiles of antibiotic resistance genes in activated sludge, dewatered sludge and bioaerosols. WATER 12, 1516 (2020).
-
Yergeau, E. et al. Comparison of methods to identify pathogens and associated virulence functional genes in biosolids from two different wastewater treatment facilities in Canada. PLoS ONE 11, e0153554 (2016).
-
Reid, C. J., Farrell, M. & Kirby, J. K. Microbial communities in biosolids-amended soils: A critical review of high-throughput sequencing approaches. J. Environ. Manag. 375, 124203 (2025).
-
Wang, D., Felice, M. L. & Scow, K. M. Impacts and interactions of biochar and biosolids on agricultural soil microbial communities during dry and wet-dry cycles. Appl. Soil Ecol. 152, 103570 (2020).
-
Ma, J., Ibekwe, A., Yang, C. & Crowley, D. Bacterial diversity and composition in major fresh produce growing soils affected by physiochemical properties and geographic locations. Sci. Total Environ. 563, 199–209 (2016).
-
Wolters, B. et al. Biosolids for safe land application: does wastewater treatment plant size matters when considering antibiotics, pollutants, microbiome, mobile genetic elements and associated resistance genes?. Environ. Microbiol. 24, 1573–1589 (2022).
-
Price, G., Langille, M. & Yurgel, S. Microbial co-occurrence network analysis of soils receiving short- and long-term applications of alkaline treated biosolids. Sci. Total Environ. 751, 141687 (2021).
-
Humphries, N. H., Thornton, S. F., Chen, X., Bray, A. W. & Stewart, D. I. Response of soil bacterial populations to application of biosolids under short-term flooding. Environ. Sci. Pollut. Res. 30, 72978–72992 (2023).
-
Zhang, J. et al. Soil types influence the fate of antibiotic-resistant bacteria and antibiotic resistance genes following the land application of sludge composts. Environ. Int. 118, 34–43 (2018).
-
Larsbrink, J. & McKee, L. S. Chapter Two – Bacteroidetes bacteria in the soil: Glycan acquisition, enzyme secretion, and gliding motility. In Advances in Applied Microbiology (eds. Gadd, G. M. & Sariaslani, S.) 63–98 (Academic Press, 2020).
-
Mossa, A.-W., Dickinson, M. J., West, H. M., Young, S. D. & Crout, N. M. J. The response of soil microbial diversity and abundance to long-term application of biosolids. Environmental Pollution 224, 16–25 (2017).
-
Newton, R. et al. Sewage Reflects the Microbiomes of Human Populations. MBIO 6, e02574 (2015).
-
Su, J. et al. Metagenomics of urban sewage identifies an extensively shared antibiotic resistome in China. Microbiome 5, 84 (2017).
-
Klümper, U. et al. Environmental microbiome diversity and stability is a barrier to antimicrobial resistance gene accumulation. Commun. Biol. 7, 706 (2024).
-
Manyi-Loh, C., Mamphweli, S., Meyer, E. & Okoh, A. Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules 23, 795 (2018).
-
Hung, W.-C. et al. Tracking antibiotic resistance through the environment near a biosolid spreading ground: Resistome changes, distribution, and metal(loid) co-selection. Science of The Total Environment 823, 153570 (2022).
-
Lin, H. et al. Fate of tetracycline and sulfonamide resistance genes in a grassland soil amended with different organic fertilizers. Ecotoxicol. Environ. Safety 170, 39–46 (2019).
-
Zhang, T., Zhang, X.-X. & Ye, L. Plasmid metagenome reveals high levels of antibiotic resistance genes and mobile genetic elements in activated sludge. PLoS ONE 6, e26041 (2011).
-
Berendonk, T. U. et al. Tackling antibiotic resistance: the environmental framework. Nat. Rev. Microbiol. 13, 310–317 (2015).
-
Kim, D. & Cha, C. Antibiotic resistome from the One-Health perspective: understanding and controlling antimicrobial resistance transmission. Exp. Mol. Med. 53, 301–309 (2021).
-
Mays, C., Garza, G. L., Waite-Cusic, J., Radniecki, T. S. & Navab-Daneshmand, T. Impact of biosolids amendment and wastewater effluent irrigation on enteric antibiotic-resistant bacteria – a greenhouse study. Water Res. X 13, 100119 (2021).
-
Sorinolu, A. J., Tyagi, N., Kumar, A. & Munir, M. Antibiotic resistance development and human health risks during wastewater reuse and biosolids application in agriculture. Chemosphere 265, 129032 (2021).
-
Kojima, H., Mochizuki, J. & Fukui, M. Sulfuriferula nivalis sp. nov., a sulfur oxidizer isolated from snow and emended description of Sulfuriferula plumbiphila. Int. J. Syst. Evol. Microbiol. 70, 3273–3277 (2020).
-
Pascoal, F., Costa, R. & Magalhaes, C. The microbial rare biosphere: current concepts, methods and ecological principles. FEMS Microbiol. Ecol. 97, fiaa227 (2021).
-
Escalas, A. et al. Macroecological distributions of gene variants highlight the functional organization of soil microbial systems. ISME J. 16, 726–737 (2022).
-
Gu, H. et al. Distinct effects of abundant and rare microbial communities on ecosystem multifunctionality across the soil profiles in agricultural Isohumosols. SOIL Ecol. Lett. 7, 240289 (2025).
-
Sun, T. et al. Heavy metals drive microbial community assembly process in farmland with long-term biosolids application. J. Hazard. Mater. 468, 133845 (2024).
-
Shi, C. et al. Multi-omics integration analysis: Tools and applications in environmental toxicology. Environ. Pollut. 360, 124675 (2024).
-
Ekpeghere, K. I., Lee, J.-W., Kim, H.-Y., Shin, S.-K. & Oh, J.-E. Determination and characterization of pharmaceuticals in sludge from municipal and livestock wastewater treatment plants. Chemosphere 168, 1211–1221 (2017).
-
Subedi, B., Lee, S., Moon, H.-B. & Kannan, K. Emission of artificial sweeteners, select pharmaceuticals, and personal care products through sewage sludge from wastewater treatment plants in Korea. Environ. Int. 68, 33–40 (2014).
-
Chang, Q., Wang, W., Regev-Yochay, G., Lipsitch, M. & Hanage, W. P. Antibiotics in agriculture and the risk to human health: how worried should we be?. Evol. Appl. 8, 240–247 (2015).
-
Della-Negra, O. et al. Temporal dynamic of soil microbial communities and antibiotic resistance markers exposed to increasing concentrations of sulfamethoxazole. Environ. Pollut. 364, 125306 (2025).
-
Wu, J. et al. Antibiotics and antibiotic resistance genes in agricultural soils: A systematic analysis. Crit. Rev. Environ. Sci. Technol. 53, 847–864 (2023).
-
Zhu, Y. et al. Changes in bacterial community structure and antibiotic resistance genes in soil in the vicinity of a pharmaceutical factory. Ecotoxicol. Environ. Saftey 158, 87–93 (2018).
-
Wang, F.-H. et al. Impact of reclaimed water irrigation on antibiotic resistance in public parks, Beijing, China. Environ. Pollut. 184, 247–253 (2014).
-
Gatica, J. & Cytryn, E. Impact of treated wastewater irrigation on antibiotic resistance in the soil microbiome. Environ. Sci. Poll. Res. 20, 3529–3538 (2013).
-
Pan, X. et al. Microbial community and antibiotic resistance gene distribution in food waste, anaerobic digestate, and paddy soil. Sci. Total Environ. 889, 164192 (2023).
-
Nguyen, C. C., Hugie, C. N., Kile, M. L. & Navab-Daneshmand, T. Association between heavy metals and antibiotic-resistant human pathogens in environmental reservoirs: a review. Front. Environ. Sci. Eng. 13, 46 (2019).
-
Roane, T. M. & Kellogg, S. T. Characterization of bacterial communities in heavy metal contaminated soils. Can. J. Microbiol. 42, 593–603 (1996).
-
Wang, S. & Wang, H. Adsorption behavior of antibiotic in soil environment: a critical review. Front. Environ. Sci. Eng. 9, 565–574 (2015).
-
McMahon, M., Xu, J., Moore, J., Blair, I. & McDowell, D. Environmental stress and antibiotic resistance in food-related pathogens. Appl. Environ. Microbiol. 73, 211–217 (2007).
-
Navab-Daneshmand, T., Enayet, S., Gehr, R. & Frigon, D. Bacterial pathogen indicators regrowth and reduced sulphur compounds’ emissions during storage of electro-dewatered biosolids. Chemosphere 113, 109–115 (2014).
-
Zhu, L. et al. Insights into microbial contamination in multi-type manure-amended soils: The profile of human bacterial pathogens, virulence factor genes and antibiotic resistance genes. J. Hazard. Mater. 437, 129356 (2022).
-
Das, S. et al. Genome plasticity as a paradigm of antibiotic resistance spread in ESKAPE pathogens. Environ. Sci. Pollut. Res. 29, 40507–40519 (2022).
-
Forsberg, K. et al. Bacterial phylogeny structures soil resistomes across habitats. Nature 509, 612 (2014).
-
Li, B. et al. Metagenomic and network analysis reveal wide distribution and co-occurrence of environmental antibiotic resistance genes. ISME J. 9, 2490–2502 (2015).
-
Ho, A., Di Lonardo, D. & Bodelier, P. Revisiting life strategy concepts in environmental microbial ecology. FEMS Microbiol. Ecol. https://doi.org/10.1093/femsec/fix006 (2017).
-
Pei, R., Kim, S., Carlson, K. & Pruden, A. Effect of River Landscape on the sediment concentrations of antibiotics and corresponding antibiotic resistance genes (ARG). Water Res. 40, 2427–2435 (2006).
-
Munir, M. & Xagoraraki, I. Levels of antibiotic resistance genes in manure, biosolids, and fertilized soil. J. Environ. Quality 40, 248–255 (2011).
-
Chen, Q. L. et al. Long-term field application of sewage sludge increases the abundance of antibiotic resistance genes in soil. Environ. Int. 92–93, 1–10 (2016).
-
Calderón-Franco, D., van Loosdrecht, M., Abeel, T. & Weissbrodt, D. Catch me if you can: capturing microbial community transformation by extracellular DNA using Hi-C sequencing. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol 116, 667–685 (2023).
-
Risely, A. et al. Host- plasmid network structure in wastewater is linked to antimicrobial resistance genes. Nat. Commun. 15, 555 (2024).
-
Rice, E., Wang, P., Smith, A. & Stadler, L. Determining Hosts of Antibiotic Resistance Genes: A Review of Methodological Advances. Environ. Sci. Technol. Lett. 7, 282–291 (2020).
-
Kim, J. et al. Persistence of antibiotic resistance from animal agricultural effluents to surface water revealed by genome-centric metagenomics. J. Hazard Mater. 457, 131761 (2023).
-
Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (2010).
-
Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J.Vol 17, No 1: Next Gen. Sequencing Data Anal. https://doi.org/10.14806/ej.17.1.200 (2011).
-
Menzel, P., Ng, K. L. & Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 7, 11257 (2016).
-
Li, D., Liu, C.-M., Luo, R., Sadakane, K. & Lam, T.-W. MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 31, 1674–1676 (2015).
-
Yin, X. et al. ARGs-OAP v2.0 with an expanded SARG database and Hidden Markov Models for enhancement characterization and quantification of antibiotic resistance genes in environmental metagenomes. Bioinformatics 34, 2263–2270 (2018).
-
Liu, B., Zheng, D., Jin, Q., Chen, L. & Yang, J. VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nuclear Acid Res. 47, D687–D692 (2019).
-
Oksanen, J. et al. vegan: Community Ecology Package. https://cran.r-project.org/web/packages/vegan/vegan.pdf (2024).
-
Csardi, G. & Nepusz, T. Igraph – Network Analysis Software. https://igraph.org/ (2006).
Acknowledgements
This work was supported by the USDA National Institute of Food and Agriculture, Agricultural and Food Research Initiative Competitive Program, Agriculture Economics and Rural Communities, grant number: 2018-67017-27631. The funder played no role in study design, data collection, analysis and interpretation of data, or the writing of this manuscript.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ste. Marie, J., Mays, C., Guo, B. et al. Longitudinal replicated metagenomic analysis of biosolids-amended soils reveals enrichment of ARGs, virulence factors, and ESKAPE pathogens. npj Antimicrob Resist 3, 96 (2025). https://doi.org/10.1038/s44259-025-00166-4
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s44259-025-00166-4