
Introduction
Plants offer an extensive source of bioactive compounds that have significant potential for pharmaceutical and therapeutic applications1. These compounds, commonly referred to as secondary metabolites, exhibit diverse biological activities. Beyond traditional approaches to enhance plant secondary metabolites, advancements in understanding the molecular regulation of plant development—such as m⁶A RNA modifications—have opened new avenues for modulating metabolite biosynthesis through targeted interventions2.
Sonchus arvensis L. is a perennial weed that is widely distributed in various regions3. The traditional use of S. arvensis as a medicinal plant was documented through ethnobiology studies in Bologna4, and in multiethnic areas of the Gansu–Ningxia–Inner Mongolia intersection zone5. Recent scientific studies in this species have demonstrated the presence of valuable secondary metabolites, including flavonoids, phenolic acids and terpenoids6,7,8,9. These findings support its traditional use and emphasises its potential as a source of new therapeutic agents10. Previous studies have clearly demonstrated the antioxidant and antiplasmodial effects of S. arvensis L. leaves7. Nevertheless, the potential for increasing these effects by inducing secondary metabolites has not been adequately examined.
Callus cultures offer a promising platform for large-scale production of secondary metabolites11. However, the yields of these bioactive compounds from in vitro cultures are often lower than those from wild plants12. To overcome this challenge, various in vitro strategies have been developed to increase the production of secondary metabolites13. In addition to classical elicitors, insights from food science now suggest that nanoparticle-like bioactive assemblies—similar to those found in black tea infusions—may influence metabolite composition and stability14, offering a cross-disciplinary perspective on optimizing plant extract qualities and highlighting the breadth of strategies being explored.
Glutamine, an amino acid with a multitude of functions in plant metabolism, has recently been shown to be a potential elicitor15. Glutamine is involved in a number of physiological processes, including stress responses and the regulation of gene expression16,17. Studies have shown that the addition of glutamine to the culture medium together with other elicitors increases the production of plant secondary metabolites, in particular the production of plant antioxidants in Taxus cell cultures18 and withanolides in Withania somnifera cell suspension cultures19. Moreover, treating Ficus benghalensis callus with 150 mg/L glutamine resulted in improved growth and a remarkable 20-fold increase in quercetin content compared to the mother plant20. As research on therapeutic phytochemicals advances, recent studies also emphasize the importance of considering bioavailability and gender-specific metabolic responses when assessing compounds like kokusaginine, highlighting the need for standardized approaches in evaluating plant-derived bioactives21.
Therefore, to address the current lack of knowledge regarding the potential of glutamine in enhancing bioactive compound production in S. arvensis callus cultures, this study was designed to investigate the effect of glutamine supplementation on callus morpho-anatomy, biomass, metabolite profile, in vitro antioxidant activity using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) and 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical scavenging assays and in vitro antiplasmodial potential against Plasmodium falciparum strain 3D7. This is particularly relevant in the context of global efforts to identify safe and effective plant-based agents with antidiabetic or antioxidant roles, as demonstrated by the mechanistic elucidation of polyphenols from Myrica rubra22. The results provide valuable insights into the role of glutamine in improving the therapeutic properties of S. arvensis L. callus. This work may contribute to the development of sustainable and efficient strategies for the production of valuable plant compounds with potential applications in the pharmaceutical and healthcare industries.
Results
Morphology of Sonchus arvensis L. callus under glutamine treatment
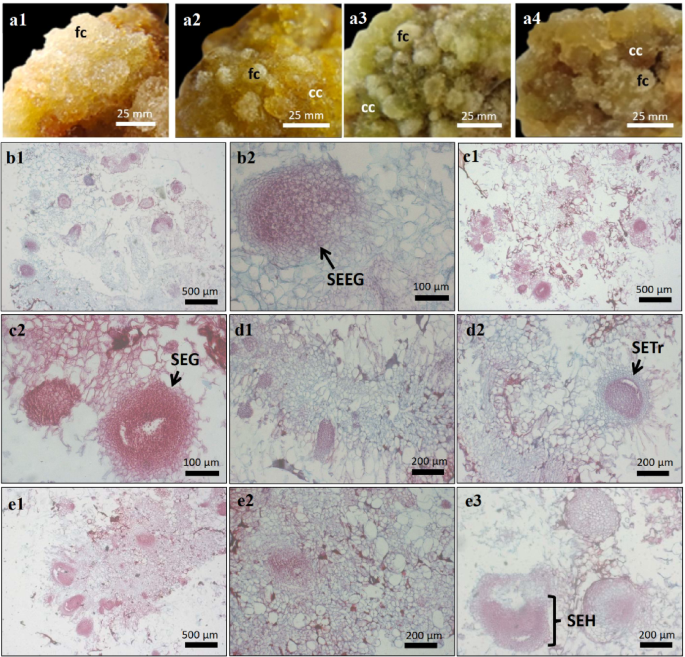
The S. arvensis L. callus obtained in the induction phase by culturing leaves explant on Murashige and Skoog (MS) medium supplemented with 1 mg/L 2,4-Dichlorophenoxyacetic acid (2,4-D) and 0.5 mg/L 6-Benzylaminopurine (BAP), was subcultured on the same media containing 0, 150, 250 and 350 mg/L glutamine. The texture and colour of the callus were monitored over a period of four weeks (Table 1). Glutamine supplementation promoted the formation of both compact and friable callus (Fig. 1). The friable callus observed in all treatments showed a transparent appearance with different colours, including white, yellow, greenish and brownish-yellow. It formed on the surface and consisted of easily separable cells. In the first week of subculture, compact callus was formed, characterised by a brownish-yellow to green colour, a smooth surface and a dense cell structure (Table 1).

Morpho-anatomical characters of S. arvensis L. callus treated with glutamine. a, Morphology of S. arvensis L. callus. (a1), control. (a2), 150 mg/L glutamine. (a3), 250 mg/L glutamine. (a4), 350 mg/L glutamine. (b–e), Cross-section of S. arvensis L. callus. b1-b2, control callus (b1: filled with parenchymatous cells, b2: somatic embryo in early globular stages/SEEG). (c1–c2), callus from 150 mg/L glutamine (c1: filled with parenchymatous cells, c2: somatic embryo in globular stages/SEG). (d1–d3), callus from 250 mg/L glutamine (d1: filled with parenchymatous cells, d2: somatic embryo in transition stage from globular to heart stage/SETr). (e1–e3), callus from 350 mg/L glutamine (e1-e2: filled with parenchymatous cells, e3: somatic embryo in heart stage/SHE).
Anatomical characters of Sonchus arvensis L. callus under glutamine treatment
The cross-section of the callus of S. arvensis L. in the control medium showed friable cell structures and consisted mainly of large parenchymatous cells (Fig. 1). The calli from 150, 250 and 350 mg/L glutamine medium were compact and solid, composed of parenchymatous and meristematic cells (Fig. 1). In addition, somatic embryo structures were observed in all treatments, mostly in globular stage. The somatic embryos in heart stage were only observed on callus from 350 mg/L glutamine medium (Fig. 1).
Biomass of Sonchus arvensis L. callus under glutamine treatment
The S. arvensis L. calli grown on glutamine and control medium were harvested every week until the 4th week and their fresh and dry weights were measured. The result showed that callus biomass varied every week under glutamine treatment (Table 1). None of the treated callus had significantly different fresh and dry weights after one week and two weeks of subculture, except for the 350 mg/L glutamine treatment, which had the highest fresh weight of 1.233 ± 0.055 g one week after subculture. Callus harvested three and four weeks after subculture on the control medium produced the highest fresh weight of 3.228 ± 0.204 g and 4.525 ± 0.228 g, respectively. The highest dry weight was also produced on the control medium at 0.226 ± 0.024 g (three weeks after subculture) and 0.272 ± 0.014 g (four weeks after subculture) and the 150 mg/L glutamine treatment (four weeks after subculture) (Table 1).
The extraction yield and metabolite profile of Sonchus arvensis L. callus under glutamine treatment
The dried callus of S. arvensis L. was extracted with ethanol (100%). The result showed that the extraction yield of the ethanolic extract ranged from 10.00 to 17.86% and that the three-week-old callus extract from the treatment with 150 mg/L glutamine gave the highest extraction yield of 17.86%. The lowest extraction yield (10.00%) was obtained with the one-week-old callus extract from a treatment with 250 mg/L glutamine.
Ethanolic extract of S. arvensis L. leaves, callus on control medium, callus on glutamine medium (150 mg/L glutamine, 250 mg/L glutamine and 350 mg/L glutamine treatments) were analysed by Gas Chromatography-Mass Spectrometry (GC-MS) analysis. Callus ages (1, 3 and 4 weeks) were selected based on callus morphology, anatomy and weight.
In the GC-MS analysis, a total of 72 metabolites were detected in all extracts (similarity index ≥ 80), each with different compounds and percentages (Supplementary file 1). Four metabolites were detected only in callus extracts and 51 metabolites were detected exclusively in callus extracts treated with glutamine. Only two metabolites (1-triacontanol and stigmasterol) were detected in all extracts. In addition, two metabolites were only detected in the control callus and seven in the leaf extract. The remaining six metabolites were only detected in the glutamine-treated callus extracts and the leaf extract.
Methyl alpha-D-glucopyranoside (27.18–62.11%) and methyl beta-D-glucopyranoside (59.21–74.29%) had the highest percentage and were only detected in callus extracts (Table 2). Betulin, on the other hand, showed the highest percentage of 24.41% in the leaf extract. Methyl-alpha-D-glucopyranoside was detected in the four-week-old control callus extract and in the early phase of the callus extracts treated with glutamine. After three weeks (250 mg/L and 350 mg/L glutamine treatment) and four weeks (150 mg/L glutamine treatment), methyl-beta-D-glucopyranoside was detected in the callus extracts instead of methyl-alpha-D-glucopyranoside (Fig. 2).

Chromatogram of S. arvensis L. callus treated with glutamine. (a1), 150 mg/L glutamine treatment in week-1. (a2), 150 mg/L glutamine treatment in week-3. (a3), mg/L glutamine treatment in week-4. (b1), 250 mg/L glutamine treatment in week-1. (b2), 250 mg/L glutamine treatment in week-3. (b3), 250 mg/L glutamine treatment in week-4. (c1), 350 mg/L glutamine treatment in week-3. (c2), 350 mg/L glutamine treatment in week-3. (c3), 350 mg/L glutamine treatment in week-4. (d), control callus (without glutamine) in week-4. (e), leaves of S. arvensis L. wild plant.
Computational analysis of antioxidant and antiplasmodial compounds
A computational approach was employed to analyse the interactions between target proteins and the ligands present within a range of S. arvensis L. callus compounds, which were identified via GC-MS (Table 2). The three-dimensional structure of the compound was displayed with sticks and specific atom colouring such as green (carbon), red (oxygen), and grey (hydrogen). The protein targets selected for investigation were Superoxide dismutase 1 (SOD1) from Homo sapiens and P. falciparum plasmepsin V (PfPMV) from Plasmodium sp. Target protein’s 3D structure was displayed through transparent surfaces, cartoons, and single colour selection (Fig. 3).

Molecular docking and dynamic simulation of ligand and target protein. (a1, b1), Ligand-protein structural visualization from the molecular docking simulation. (a2, b2), Visualization of ligand-protein molecular interactions. (a3, b3), Plot of RMSF values on ligand-protein complexes from molecular dynamics simulations. (a) Stigmasterol_SOD1 (b) Stigmasterol_PfPMV.
Prior to molecular docking, drug-likeness was performed to identify the similarity of properties in query compounds with drug molecules. All compounds subjected to analysis from S. arvensis L. callus extract was predicted to be drug-like molecules with bioavailability values of 0.55, except 9,12-Octadecadienoic acid (Z, Z) and n-Hexadecanoic acid (Table 3). In addition, the prediction of biological activity showed that all compounds have the probability of activation (Pa) score more than 0.3 as antioxidant and antimalaria (Table 3).
This study used the screening docking method to identify the binding ability of ligands from Sonchus arvensis L. callus extract with SOD1 and PfPMV to trigger inhibitory activity. The results showed that stigmasterol from Sonchus arvensis L. callus extract had a more negative binding affinity than other compounds when binding to SOD1 and PfPMV (Table 4). Ligand-protein complexes with the most negative binding affinity are displayed with transparent surfaces, cartoons, and sticks structures with single colouring, red colour for the target and green is dominated by the ligand (Fig. 3).
Ligand interaction at the target domain also showed weak bonds formation. Stigmasterol interacts with the SOD1 domain through hydrogen bonding at the Asn86 residue position, van der Waals at Glu78, Glu77, Glu100, Leu84, Gly85, Gly130, Gly127, and alkyl at Lys70, Ile99, Pro74, Lys128, Lys75 (Fig. 3). Binding to PfPMV, ligands from stigmasterol formed hydrogen bonds at residues Ser113 and Asn207, van der Waals bonds at residues Glu120, Glu121, Phe166, Ser112, Ser151, alkyl/pi-alkyl bonds via Arg167, Pro123, Leu115, Phe124, Leu169, Pro200, and Leu205 (Fig. 3).
Molecular dynamics simulations were performed to identify the stability of interactions formed in ligand-protein complexes, as shown by the root mean square fluctuation (RMSF) values. Stigmasterol_SOD1 (https://biocomp.chem.uw.edu.pl/CABSflex2/job/a4eb99f5f42d406/) and Stigmasterol_PfPMV (https://biocomp.chem.uw.edu.pl/CABSflex2/job/25fa06e3c3ebe22/) complexes formed through molecular dynamic simulations exhibit stable interactions, with an average RMSF value below 3 (Å) (Fig. 3).
Antioxidant activity of Sonchus arvensis L. callus under glutamine treatment
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) and 2,2’-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) assays were performed to evaluate the antioxidant activity of the ethanolic extracts. All extracts showed antioxidant activities. The half-maximal inhibitory concentration (IC50) values of the crude callus and leaf extracts compared to quercetin (positive control) are shown in Table 5. The one-week-old callus of S. arvensis L. treated with glutamine exhibited the strongest antioxidant activity. The IC50 values of ABTS were 78.43 ± 0.9, 96.45 ± 1.26 and 75.83 ± 2.54 µg/mL for the treatment with 150 mg/L, 250 mg/L and 350 mg/L glutamine, respectively. The IC50 values of DPPH for callus treated with 250 mg/L and 350 mg/L glutamine were 79.06 ± 1.46 and 76.71 ± 4.28 µg/mL, respectively. In contrast, the IC50 values of DPPH for callus treated with 150 mg/L glutamine showed a higher IC50 value than ABTS (108.63 ± 2.48 µg/mL).
Antiplasmodial activity of Sonchus arvensis L. callus under glutamine treatment
In this study, the in vitro antiplasmodial activity of the samples against Plasmodium falciparum strain 3D7 was evaluated. The percentage inhibition data were subjected to probit analysis. The result showed that the IC50 of the ethanolic extracts of the control callus, leaves, callus treated with 250 mg/L glutamine and callus treated with 350 mg/L glutamine were 1.248 µg/mL, 9.548 µg/mL, 0.372 µg/mL and 0.982 µg/mL, respectively (Table 5).
Discussion
The increasing incidence of infectious diseases and the rise in free radicals caused by factors such as pollution, unhealthy lifestyles and stress pose a significant threat to human health23. These challenges have strongly driven research to develop novel therapeutic agents from natural sources, especially from medicinal plants24. Among these, S. arvensis L. has emerged as a promising candidate due to its history in traditional medicine and its diverse pharmacological properties25.
Previous studies have provided convincing evidence for the antioxidant and antiplasmodial effects of the leaves of S. arvensis L7. However, the potential to enhance these bioactivities by triggering secondary metabolites has not yet been sufficiently explored. In this study, the release of secondary metabolites in callus of S. arvensis L. was investigated by treatment with glutamine. The callus cultures were grown on MS medium enriched with 1 mg/L 2,4-D and 0.5 mg/L BAP and different glutamine concentrations (0, 150, 250 and 350 mg/L).
The morphology, anatomy and biomass accumulation in the callus of S. arvensis L. in response to glutamine supplementation emphasise the important role of this amino acid in the regulation of plant growth and development in vitro. The friable part on the surface of the callus, characterised by loosely arranged, highly vacuolated parenchymatous cells, is consistent with a cellular state optimised for rapid cell division and expansion. This is confirmed by observations in Boerhaavia paniculata, where friable callus often exhibits increased growth rates and metabolic activities26. The improved nutrient uptake and gas exchange facilitated by the loose cell structure could contribute to the observed increase in biomass in the glutamine-treated cultures, especially in the first weeks of subculture. In addition, the presence of friable callus across all glutamine concentrations suggests a general response to nitrogen availability, as glutamine serves as a primary nitrogen source and precursor for various metabolic processes27. According to Setiawati et al.28 supplementing the medium with glutamine positively influenced the development of friable callus in oil palm callus culture, resulting in structures with meristematic characteristics.
The change in callus texture from friable to compact in glutamine-supplemented media aligns with previous observations in Sonchus arvensis L. callus grown on dolomite media29. Compact and globular-shaped callus was also reported in the Malaysian upland rice (Oryza sativa) cv. Panderas treated with glutamine30. The compact callus has a more organised structure with densely packed parenchymatous and meristematic cells. This denser arrangement combined with increased lignification could indicate a shift towards a more differentiated state, possibly influenced by the role of glutamine in nitrogen signalling and cell wall development31. The presence of somatic embryos, especially at higher glutamine concentrations, supports the role of glutamine in triggering organised cellular differentiation for this term, as embryogenesis is characterised by organised cell division and differentiation32. The exclusive observation of heart-stage embryos at 350 mg/L glutamine aligns with the globular to torpedo-shaped embryo development observed in okra cultures at 300 mg/L glutamine after 3 weeks33. This positive effect of high glutamine concentrations on embryogenesis is further supported by findings in date palm callus culture34.
Interestingly, despite the morphological and anatomical differences, there was no consistent pattern in the total biomass accumulation in the different glutamine treatments. While the 350 mg/L treatment initially resulted in a higher fresh weight, the control medium showed the highest biomass after three and four weeks. This suggests that while glutamine may stimulate early growth, prolonged exposure or higher concentrations may trigger differentiation pathways that prioritise organised development over rapid cell proliferation. In contrast with our finding, a study on Hylocereus costaricensis callus culture found that callus biomass was unaffected by the addition of various glutamine concentrations35.
In relation to the metabolite profile under investigation, the presence of major components in the leaf extract is notable, while the absence or very low relative abundance of these components in the callus extracts is also worthy of attention. This observation lends further support to the hypothesis that in vitro cultures have the potential to produce unique compounds36. The accumulation of specific metabolites in the callus, such as glucopyranoside, could be due to the de-differentiated state of the cells and the altered metabolic pathways associated with callus growth. A substantial number of metabolites, precisely 51, were identified as being exclusive to the callus that had been treated with glutamine. Given the limited data available on the biosynthesis pathways of the identified compounds (as detailed in Supplementary file 1), it would be premature to conclude that glutamine directly triggers their production. Nevertheless, considering glutamine’s known roles as a key nitrogen source and signaling molecule37 its addition to the culture medium might have influenced protein synthesis, potentially leading to the activation of specific biosynthetic routes for these compounds. Further investigation is warranted to elucidate the molecular mechanisms by which glutamine affects S. arvensis L. callus, specifically focusing on the genes and pathways involved in secondary metabolite biosynthesis.
A range of bioactive and pharmacological properties have previously been reported for certain compounds identified via GC-MS from S. arvensis L. callus extract. For instance, methyl-α-D-glucopyranoside has been demonstrated to have anticancer activity38 while methyl-β-D-glucopyranoside has been shown to exhibit anti-quorum-sensing activity39. In addition, stigmasterol and n-hexadecanoic acid have been reported to possess anti-inflammatory, antibacterial, and antioxidant properties40,41,42,43,44. Meanwhile, 9,12-Octadecadienoic acid (Z, Z)- and Octadecanoic acid, 2,3-dihydroxypropyl ester have been documented to display potent antimicrobial properties45.
In the present study, these compounds were subjected to computational analysis using the molecular docking method for antioxidant and antimalarial activities. This analysis aimed to predict the interaction between compounds as drug candidate and target proteins consisting of SOD1 and PfPMV. Superoxide dismutase 1 (SOD1) from Homo sapiens has a 180-mer sequence length with a structure weight of 192.46 kDa, atom number of 12,743, resolution of 1.90 (Å), classification including oxidoreductase, and the experimental method to obtain a three-dimensional structure view is X-ray diffraction46. SOD1 also belongs to an enzyme cluster that acts as an antioxidant with a scavenger mechanism to produce H2O2 by decomposing O2-, this enzyme has metal ions consisting of Zn, Cu, and Mn47.
P. falciparum plasmepsin V (PfPMV) from Plasmodium sp. has a 444-mer sequence length with a structural weight of 103.74 kDa, an atomic number of 6,854, a resolution of 2.37 (Å), a classification including hydrolase, and the experimental method for obtaining a three-dimensional structural view is X-ray diffraction48. Inhibition of PfPMV has been demonstrated to disrupt the export of gametocyte proteins and gametocytogenesis, thereby preventing P. falciparum infection of Anopheles mosquitoes49.
The drug-like molecules compounds from S. arvensis L. callus extract was predicted to have antioxidant and antimalarial activity based on probability of activation (Pa) and probability of inhibition (Pi) through PASS online. Query compounds with Pa > 0.3 (medium confidence) indicates that the predicted activity of the query compound is only theoretically proven and can be validated again through laboratory assays50. Among them, stigmasterol was predicted as potential antioxidant and antimalarial agent which can form a more negative binding affinity value than other compounds. Stigmasterol is predicted to have antioxidant activity by initiating SOD1 activation through interactions on residues Asn86, Glu78, Glu77, Glu100, Leu84, Gly85, Gly130, Gly127, Lys70, Ile99, Pro74, Lys128, and Lys75 with hydrogen, van der Waals, and alkyl bonds. The compound can also be an antimalarial with an inhibitory mechanism on PfPMV through interactions on residues Ser113, Asn207, Glu120, Glu121, Phe166, Ser112, Ser151, Arg167, Pro123, Leu115, Phe124, Leu169, Pro200, and Leu205 with hydrogen bonds.
The antioxidant and antiplasmodial activity of the ethanolic extracts of S. arvensis L. callus was evaluated in vitro. The results showed that the extract of callus stimulated with glutamine exhibited strong to moderate antioxidant activity after one and three weeks of culture (50 µg/mL > IC50 > 150 µg/mL)51. The increased radical scavenging activity by up to two-fold compared to control callus could be due to the successful elicitation of agents with antioxidant activity. In a previous study, glutamine was reported to increase the concentration of total phenolics and flavonoids in Baciparu callus15. Phenolic and flavonoid compounds are known for their role as natural antioxidants52. However, the S. arvensis callus extract treated with glutamine showed weak to very weak antioxidant activity after four weeks of culture (150 > IC50 > 200 µg/mL)51. This result indicates that prolonged exposure or higher concentrations of glutamine could reduce the antioxidant activity of the callus extract (Table 5).
The in vitro antiplasmodial activity of the ethanolic extract from the leaves and callus of S. arvensis L. shows high activity with an IC50 value of less than 10 µg/ml53. Moreover, the callus treated with glutamine showed a three-fold increase in Plasmodium falciparum inhibition compared to the control callus (Table 5). This result showed that the glutamine-enhanced S. arvensis callus has great potential as an antimalarial agent, as the industry standard states that a pure compound is considered antimalarial if the IC50 value is below 10 µg/mL54. This glutamine-enriched ethanolic extract of S. arvensis L. callus also showed higher activity compared to a previous study using a dolomite-enriched ethanolic extract of S. arvensis L. callus with an IC50 value of 5.944 µg/mL29. This result suggests that glutamine stimulation in S. arvensis L. callus culture systems is an effective strategy to improve the production of antimalarial drugs.
Conclusion
Glutamine supplementation influenced callus morphology and anatomy, leading to predominantly compact structures. GC-MS analysis of callus ethanolic extracts revealed diverse metabolites, including 51 unique compounds to the glutamine treatment. In silico predictions suggested that major compounds present possess antioxidant and antimalarial potential. Furthermore, in vitro assays showed that glutamine supplementation, most effectively at 250 mg/L, boosted radical scavenging activity (DPPH/ABTS) up to two-fold and Plasmodium falciparum inhibition up to three-fold compared to the control callus. This study was limited to bioactivity testing of ethanol extracts, and individual metabolites were not isolated, elucidated, quantified, or tested. However, this research provides new insights into the influence of glutamine on callus morpho-anatomy, biomass, metabolite profile, in vitro antioxidant and antiplasmodial activity. For further investigation, it’s necessary to determine the effect of glutamine on the production of potential metabolites by isolating and purifying the compounds from S. arvensis L. callus. Future research should explore the molecular mechanisms of glutamine-induced changes in S. arvensis L. callus, including gene and pathway identification in secondary metabolite biosynthesis. Understanding the genes and pathways affected by glutamine could lead to more precise and effective metabolic engineering techniques for boosting the production of valuable compounds in S. arvensis L.
Methods
Plant material
The young leaves of Sonchus arvensis L., the second and third leaf of the shoot bud, were used as explant source. The samples were collected from Taman Husada Graha Famili, Surabaya, Indonesia. The plant was identified by Purwodadi Botanical Garden, Nasional Research and Innovation Agency, Indonesia.
Media Preparation
Callus induction was performed using Murashige and Skoog (MS) medium supplemented with 3% (w/v) sucrose, 8 g/L agar, 1 mg/L 2,4-Dichlorophenoxyacetic acid (2,4-D) and 0.5 mg/L 6-Benzylaminopurine (BAP) according to a previous study29. The calli from the induction stage were subcultured in MS medium supplemented with 3% (w/v) sucrose, 8 g/L agar, 1 mg/L 2,4-D, 0.5 mg/L BAP and glutamine (0, 150, 250 and 350 mg/L). The pH was adjusted to 5.6–5.8 prior to sterilization in an autoclave at 121ºC for 15 min.
Callus induction and subculture on glutamine media
The leaves were washed with a detergent solution and rinsed three times with running tap water. The sterilisation procedure was then carried out in a cabinet with laminar air flow. The leaves were soaked in a solution of 0.1 g/L fungicide (activated with 70% propineb) for 15 min, rinsed three times with sterilised distilled water, then immersed in 20% (v/v) commercial bleach containing 5.25% sodium hypochlorite (NaClO) for 15 min and rinsed three times with sterilised distilled water. The sterilised leaves were cut into 1 cm2 squares and inoculated onto 30 ml solid medium in a culture jar, which was then covered with aluminium foil. The culture was incubated for 28 days at 25ºC ± 1ºC and continuous illumination (24 h, 650 ± 45 lx light intensity). After 28 days of culture, the callus was harvested and a half of gram callus was subcultured into glutamine treatment medium. The cultures were incubated for 28 days under the same conditions as for callus induction culture. The morphology of the callus, including colour and texture, was examined and documented at weekly intervals. The fresh weight and dry weight of the callus were also determined.
Anatomical characterization of callus
The anatomical structure of the callus of S. arvensis L. was determined using the paraffin envelope method as previously described29. The callus harvested after 21 days of culture was fixed in 70% ethanol and then serially rehydrated with 70% ethanol and water. Infiltration and embedding were performed with a paraffin wax with a melting point of 58–60ºC. The samples were sectioned into thin slices of 10 μm thickness using a Leica RM 2145 rotary microtome and then stained with safranin.
Leaves and callus extraction
The callus and leaves were dried in the air until a constant weight was reached. The dried callus and leaves were ground into powder using a mortar and pestle. The powder was macerated three times for 24 h with 100% ethanol (1:10). The liquid was filtered through filter paper and evaporated until the dry mass of the extract was constant. The extract was stored at 4ºC until further analysis.
Metabolites profiling by gas chromatography-mass spectrometry (GC-MS) analysis
Fifty milligrams of the extract was dissolved in 1.5 ml ethanol and then filtered through a 45 μm filter. One microlitre of the sample was injected into the GCMS-TQ8030 Shimadzu (Triple Quad) with RTX 5-MS capillary column using a continuous flow of helium gas (1 ml/minute) as the mobile phase. The initial oven temperature was held at 80 ºC for 2 min and then increased by 15 ºC per minute until it reached 300 ºC and then held for 6 min. The injector temperature was maintained at 230 ºC for a total run time of 24 min. The interface and ion source for the mass spectra were held at 250ºC and 200ºC respectively. The mass spectra were obtained with a mass scan range of 40–600 m/z. The identification of the compounds was based on the mass spectra of the NIST version 02.L libraries. The compounds with a similarity index (SI) score of 80 or more were selected for this study.
In Silico analysis of antioxidant and antiplasmodial compounds
Sample collection
Chemical compounds from callus extract used in this study consist of methyl alpha-D-Glucopyranoside, methyl beta-D-Glucopyranoside, n-Hexadecanoic acid, 9,12-Octadecadienoic acid (Z, Z)-, Octadecanoic acid, 2,3-dihydroxypropyl ester, 1-Triacontanol, and Stigmasterol. Ligands are retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/) with data consisting of name, CID, SMILE Canonical, link, and structure data format (sdf)55. Superoxide dismutase 1 (SOD1) PDB ID: 5YTU and P. falciparum plasmepsin V (PfPMV) PDB ID: 4ZL4 were used as screening targets for antioxidant and antimalaria candidates in this study, protein databank (https://www.rcsb.org/) was used to retrieve the pdb files of the targets46,48.
Drug-likeness and activity prediction
Swiss-ADME (http://www.swissadme.ch/) is used to identify the similarity of properties of query compounds with drug molecules through druglikeness prediction with parameters based on Lipinski, Ghose, Veber, Egan, and Muegge rules. A compound is categorized as a drug-like molecule if it meets at least one of the five rules. Bioavailability (BA) prediction is done to identify the circulation of the query compound in the body, synthetic accessibility (SA) shows the estimated difficulty in synthesizing the query compound56. Prediction of the probability of query compounds as antioxidant and antimalaria was performed through PASS online (https://www.way2drug.com/passonline/). Parameters of prediction on query compounds are probability activation score (Pa) > 0.3 for medium confidence as antioxidant and antimalaria57.
Virtual screening
Virtual screening methods such as molecular docking to predict the potential of query compounds as antioxidants and antimalaria. Docking preparation was performed on all ligands by converting sdf files into pdb through OpenBabel v3.1.1, target preparation was performed through PyMol v2.5.2 (Schrödinger, Inc., USA) by removing water molecules and native ligands58. Screening docking method with grid setup directed at the entire surface of SOD1 and PfPMV, this simulation was performed through PyRx (v1.0.0; The Scripps Research Institute, La Jolla, CA, USA) with the output of binding affinity score (kcal/mol)59,60. Visualization of the three-dimensional structure of the ligand-protein complex from the docking results was displayed through PyMol v2.5.2 (Schrödinger, Inc., USA) with the structure of cartoons, transparent surfaces, and sticks61.
Ligand-protein interaction
Discovery Studio Visualizer™ v.16.1 (Dassault Systèmes SE, France) was used to identify the bond types and positions of ligand-protein interactions from the docking results with the most negative binding affinity scores. The types of bonding interactions in ligand-protein complexes consist of hydrogen, van der Waals, pi, alkyl, hydrophobic, and electrostatic. Weak bonding interactions contribute to initiating target protein activity when the ligand binds to specific domains8,62.
Dynamic simulation
Stability level of interaction in ligand-protein complex was identified through CABS-flex v2.0 (http://biocomp.chem.uw.edu.pl/CABSflex2/index) by molecular dynamics simulation. The root mean square fluctuation (RMSF) score was reviewed to determine the flexibility of the target protein when interacting with the ligand used in this study, protein domain residues flexibility is an important factor for the determination of ligand-protein interaction stability63.
In vitro antioxidant activity assay
The antioxidant activity was evaluated using 2,2-diphenyl-1-picryl-hydrazyl hydrate (DPPH) and 2,2’-azinobis-3-ethylbenzothiazoline-6-sulfonate (ABTS) inhibition assay64. For the DPPH inhibition assay, 100 µl of the 0.2 mM DPPH reagent was mixed with 100 µl of each sample in methanol at different concentrations (1.075, 3.125, 6.25, 10.00, 15.00, 25.00, 35.00, 50.00, 75.00, 100.00, 150.00 and 200.00 µg/ml) and methanol as a control. After a 30-minute incubation in the dark at room temperature, the absorbance was determined at a wavelength of 517 nm.
For the ABTS inhibition test, ABTS solution (7 mM) was mixed with 2.4 mM potassium persulfate solution to prepare the ABTS reagent. The mixture was stored at room temperature in the dark for 12–16 h before the inhibition test. One hundred microlitres of ABTS reagent was combined with 100 µl of samples at concentrations of 1.075–200 µg/ml. After 5 min incubation at room temperature in the dark, the absorbance was determined at 734 nm. The positive control used for the DPPH and ABTS assay was quercetin.
The percentage inhibition of DPPH or ABTS was calculated using the following formula:
$${text{Percentage~of~inhibition~}}left( {text{% }} right)=left( {{text{A}}{{text{~}}_{{text{control}}}} – {text{~}}{{text{A}}_{{text{sample}}}}} right)/{text{A}}{{text{~}}_{{text{control}}}}{text{~}} times {text{~}}100{text{% }}$$
where Asample is the absorbance of the sample and Acontrol is the absorbance of the DPPH or ABTS reagent. The percentage of inhibition results at various concentrations were plotted and regressed linearly in order to obtain the IC50 values.
In vitro antiplasmodial assay
The antiplasmodial test was performed as described in a previous study29. The leaf/callus extract was dissolved in 10% dimethyl sulphoxide (DMSO) at different concentrations (0.01, 0.1, 1, 10 and 100 µL) and tested in vitro against Plasmodium falciparum strain 3D7 at the ring stage. The P. falciparum cultures were synchronised to the ring stage with 5% sorbitol. DMSO was used as a negative control and chloroquine diphosphate as a positive control. The test was performed in a 24-well microplate and incubated at 37ºC for 48 h. After 48 h, the percentage of parasitemia, inhibition, and growth was calculated using the following formulas:
$${text{Percentage~of~parasitemia~}}left( {text{% }} right)=frac{{{text{infected~eritrocyte}}}}{{5000{text{~of~total~eritrocyte}}}} times 100%$$
$${text{Percentage~of~growth~}}left( {text{% }} right)=~% ~parasitemia – {D_0}$$
$${text{Percentage~of~inhibition~}}left( {text{% }} right)=100{text{% }} – left[ {frac{{Xp}}{{Xk}} times 100% } right]$$
where D0 is the percentage of parasitemia at the 0-hour, whereas Xp and Xk are the percentage of growth in the treatment and negative control, respectively.
Statistical analysis
In this study, all experiments followed a Completely Randomized Design (CRD), with each treatment having three replicates. The callus biomass and IC50 data were expressed as mean ± standard deviation (SD). Statistical analysis of the callus biomass involved a one-way analysis of variance (ANOVA) to compare the means, followed by Duncan’s multiple range test to determine significant differences at a probability level of p ≤ 0.05. The percentage of inhibition of Plasmodium falciparum were statistically analysed using Probit Analysis to determine the IC50. All statistical computations were performed using IBM SPSS version 22.0.
Data availability
The data generated in the present study are included in the figures and/or tables of this article.
References
-
Liu, R. H. Dietary bioactive compounds and their health implications. J. Food Sci. 78, (2013).
-
Shan, C., Dong, K., Wen, D., Cui, Z. & Cao, J. A review of m6A modification in plant development and potential quality improvement. Int. J. Biol. Macromol. 308, 142597 (2025).
-
Brandsæter, L. O., Mangerud, K., Helgheim, M. & Berge, T. W. Control of perennial weeds in spring cereals through stubble cultivation and mouldboard ploughing during autumn or spring. Crop Prot. 98, 16–23 (2017).
-
Sansanelli, S. & Tassoni, A. Wild food plants traditionally consumed in the area of Bologna (Emilia Romagna region, Italy). J. Ethnobiol. Ethnomed. 10, 69 (2014).
-
Xie, J. et al. Study on wild medicinal plant resources and their applied ethnology in multiethnic areas of the Gansu–Ningxia–Inner Mongolia intersection zone. J. Ethnobiol. Ethnomed. 19, 18 (2023).
-
Khan, R. A. Evaluation of flavonoids and diverse antioxidant activities of Sonchus arvensis. Chem. Cent. J. 6, 126 (2012).
-
Wahyuni, D. K. et al. Antioxidant, antimicrobial, and antiplasmodial activities of Sonchus arvensis L. Leaf Ethyl acetate fractions. Pharmacogn J. 14, 993–998 (2023).
-
Wahyuni, D. K. et al. Molecular simulation of compounds from n-hexane fraction of Sonchus arvensis L. leaves as SARS-CoV-2 antiviral through inhibitor activity targeting strategic viral protein. J. Pharm. Pharmacogn Res. 10, 1126–1138 (2022).
-
Wahyuni, D. K. et al. In vitro and in vivo antiplasmodial activities of leaf extracts from Sonchus arvensis L. BMC Complement. Med. Ther. 23, 47 (2023).
-
Dzobo, K. The role of natural products as sources of therapeutic agents for innovative drug discovery. in Comprehensive Pharmacology 408–422 (Elsevier, https://doi.org/10.1016/B978-0-12-820472-6.00041-4. (2022).
-
Efferth, T. Biotechnology applications of plant callus cultures. Engineering 5, 50–59 (2019).
-
Govarthanan, M., Rajinikanth, R., Kamala-Kannan, S. & Selvankumar, T. A comparative study on bioactive constituents between wild and in vitro propagated Centella asiatica. J. Genet. Eng. Biotechnol. 13, 25–29 (2015).
-
Fazili, M. A., Bashir, I., Ahmad, M., Yaqoob, U. & Geelani, S. N. In vitro strategies for the enhancement of secondary metabolite production in plants: a review. Bull. Natl. Res. Cent. 46, 35 (2022).
-
Li, Y. et al. Deciphering composition-structure-taste relationship of black tea-infusion via assessments of nanoparticles by centrifugal treatment. LWT 222, 117601 (2025).
-
Marques, H. P. et al. Proteic and phenolics compounds contents in Bacupari callus cultured with glutamine and nitrogen sources. Brazilian J. Biol. 78, 41–46 (2017).
-
Liao, H. S. et al. Glutamine induces lateral root initiation, stress responses, and disease resistance in Arabidopsis. Plant. Physiol. 195, 2289–2308 (2024).
-
Yin, H. et al. Advances in the functional study of glutamine synthetase in plant abiotic stress tolerance response. Crop J. 10, 917–923 (2022).
-
Perez-Matas, E. et al. Impact of elicitation on plant antioxidants production in Taxus cell cultures. Antioxidants 12, 887 (2023).
-
Sivanandhan, G., Selvaraj, N., Ganapathi, A. & Manickavasagam, M. Enhanced biosynthesis of withanolides by elicitation and precursor feeding in cell suspension culture of Withania somnifera (L.) Dunal in Shake-Flask culture and bioreactor. PLoS One. 9, e104005 (2014).
-
Bandekar, H. & Lele, S. S. Production of flavonol Quercetin from cultured plant cells of Banyan (Ficus benghalensis L). Int. J. Innov. Res. Sci. Eng. Technol. 3, 12150–12157 (2014).
-
Shang, K. et al. An evaluation of Sex-Specific pharmacokinetics and bioavailability of kokusaginine: an in vitro and in vivo investigation. Pharmaceuticals 17, 1053 (2024).
-
Chang, G. et al. Hypoglycemic effects and mechanisms of polyphenols from Myrica rubra pomace in type 2 diabetes (db/db) mice. Mol Nutr. Food Res 69, (2025).
-
Ye, X. et al. Associations of socioeconomic status with infectious diseases mediated by lifestyle, environmental pollution and chronic comorbidities: a comprehensive evaluation based on UK biobank. Infect. Dis. Poverty. 12, 5 (2023).
-
Mushtaq, S., Abbasi, B. H., Uzair, B. & Abbasi, R. Natural products as reservoirs of novel therapeutic agents. EXCLI J. 17, 420–451 (2018).
-
Rafi, M., Suwartiny, N. L., Rohaeti, E. T. & Use Phytochemical composition, and biological activities of Sonchus arvensis. Indones J. Pharm. 540–553. https://doi.org/10.22146/ijp.3823 (2022).
-
Souza, J. M. M., Berkov, S. & Santos, A. S. Improvement of friable callus production of Boerhaavia paniculata rich and the investigation of its lipid profile by GC/MS. Acad. Bras. Cienc. 86, 1015–1027 (2014).
-
Reitzer, L. Amino acid synthesis. in Reference Module in Biomedical Sciences (Elsevier, doi:https://doi.org/10.1016/B978-0-12-801238-3.02427-2. (2014).
-
Setiawati, Y. et al. Characterization and morphological development of oil palm transformed-callus on modified culture media. E-J. Menara Perkeb 92, (2024).
-
Wahyuni, D. K. et al. Growth, secondary metabolite production, and in vitro antiplasmodial activity of Sonchus arvensis L. callus under dolomite [CaMg(CO3)2] treatment. PLoS One. 16, e0254804 (2021).
-
Mohd Din, A. R. J. et al. Improvement of efficient in vitro regeneration potential of mature callus induced from Malaysian upland rice seed (Oryza sativa cv. Panderas). Saudi J. Biol. Sci. 23, S69–S77 (2016).
-
Liao, H. S., Chung, Y. H. & Hsieh, M. H. Glutamate: A multifunctional amino acid in plants. Plant. Sci. 318, 111238 (2022).
-
Kadokura, S., Sugimoto, K., Tarr, P., Suzuki, T. & Matsunaga, S. Characterization of somatic embryogenesis initiated from the Arabidopsis shoot apex. Dev. Biol. 442, 13–27 (2018).
-
Daniel, M. A. et al. Effect of L-glutamine and casein hydrolysate in the development of somatic embryos from cotyledonary leaf explants in Okra (Abelmoschus esculentus L. monech). South. Afr. J. Bot. 114, 223–231 (2018).
-
El-Dawayati, M. M., Ghazzawy, H. S. & Munir, M. Somatic embryogenesis enhancement of date palm cultivar Sewi using different types of polyamines and glutamine amino acid concentration under in-vitro solid and liquid media conditions. Int. J. Biosci. 12, 149–159 (2018).
-
Winson, K. W. S., Chew, B. L., Sathasivam, K. & Subramaniam, S. Effect of amino acid supplementation, elicitation and leds on Hylocereus costaricensis callus culture for the enhancement of betalain pigments. Sci. Hortic. (Amsterdam). 289, 110459 (2021).
-
Mohaddab, M. et al. Biotechnology and in vitro culture as an alternative system for secondary metabolite production. Molecules 27, (2022).
-
Lee, K. T., Liao, H. S. & Hsieh, M. H. Glutamine metabolism, sensing and signaling in plants. Plant. Cell. Physiol. 64, 1466–1481 (2023).
-
Lyantagaye, S. L. Methyl-α-D-glucopyranoside from Tulbaghia violacea extract induces apoptosis in vitro in cancer cells. Bangladesh J. Pharmacol 8, (2013).
-
Davaapurev, B. O. et al. Methyl-β-D-glucopyranoside from Scabiosa Comosa as a Quorum-Sensing inhibitor. Nat Prod. Commun 17, (2022).
-
Aparna, V. et al. Anti-Inflammatory property of n ‐Hexadecanoic acid: structural evidence and kinetic assessment. Chem. Biol. Drug Des. 80, 434–439 (2012).
-
Purushothaman, R., Vishnuram, G. & Ramanathan, T. Isolation and identification of N-Hexadecanoic acid from Excoecaria Agallocha L. and its antibacterial and antioxidant activity. SSRN Electron. J. https://doi.org/10.2139/ssrn.4886224 (2024).
-
Liang, Q. et al. Stigmasterol alleviates cerebral ischemia/reperfusion injury by attenuating inflammation and improving antioxidant defenses in rats. Biosci. Rep. 40, (2020).
-
Sampath, S. J. P., Rath, S. N., Kotikalapudi, N. & Venkatesan, V. Beneficial effects of secretome derived from mesenchymal stem cells with stigmasterol to negate IL-1β-induced inflammation in-vitro using rat chondrocytes—OA management. Inflammopharmacology 29, 1701–1717 (2021).
-
Adnan, S. N. A., Ibrahim, N. & Yaacob, W. A. Transcriptome analysis of methicillin-resistant Staphylococcus aureus in response to stigmasterol and Lupeol. J. Glob Antimicrob. Resist. 8, 48–54 (2017).
-
Rahman, M. M., Ahmad, S. H., Mohamed, M. T. M. & Ab Rahman, M. Z. Antimicrobial compounds from leaf extracts of Jatropha curcas, Psidium guajava, and Andrographis paniculata. Sci. World J. 2014, 1–8 (2014).
-
Manjula, R., Wright, G. S. A., Strange, R. W. & Padmanabhan, B. Assessment of ligand binding at a site relevant to SOD 1 oxidation and aggregation. FEBS Lett. 592, 1725–1737 (2018).
-
Mondola, P., Damiano, S., Sasso, A. & Santillo, M. The cu, Zn superoxide dismutase: not only a dismutase enzyme. Front Physiol. 7, (2016).
-
Hodder, A. N. et al. Structural basis for plasmepsin V Inhibition that blocks export of malaria proteins to human erythrocytes. Nat. Struct. Mol. Biol. 22, 590–596 (2015).
-
Jennison, C. et al. Inhibition of plasmepsin V activity blocks plasmodium falciparum gametocytogenesis and transmission to mosquitoes. Cell. Rep. 29, 3796–3806e4 (2019).
-
Pokharkar, O., Lakshmanan, H., Zyryanov, G. & Tsurkan, M. Silico evaluation of antifungal compounds from marine sponges against COVID-19-Associated mucormycosis. Mar. Drugs. 20, 215 (2022).
-
Syakri, S., Syahrana, N. A., Ismail, A., Tahir, K. A. & Masri, A. A review: testing antioxidant activity on Kawista plants (Limonia acidissima L.) in Indonesia. Open. Access. Maced J. Med. Sci. 9, 281–287 (2021).
-
Brewer, M. S. Natural antioxidants: sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food Saf. 10, 221–247 (2011).
-
Lima, R. B. S. et al. In vitro and in vivo anti-malarial activity of plants from the Brazilian Amazon. Malar. J. 14, 508 (2015).
-
Tajuddeen, N. & Van Heerden, F. R. Antiplasmodial natural products: an update. Malar. J. 18, 404 (2019).
-
Moovarkumudalvan, B., Geethakumari, A. M., Ramadoss, R., Biswas, K. H. & Mifsud, B. Structure-Based virtual screening and functional validation of potential hit molecules targeting the SARS-CoV-2 main protease. Biomolecules 12, 1754 (2022).
-
Abdulredha, F. H., Mahdi, M. F. & Khan, A. K. In Silico evaluation of binding interaction and ADME study of new 1,3-diazetidin-2-one derivatives with high antiproliferative activity. J. Adv. Pharm. Technol. Res. 14, 176–184 (2023).
-
Taibi, M. et al. Assessment of the antioxidant and antimicrobial potential of Ptychotis verticillata Duby essential oil from Eastern morocco: an in vitro and in Silico analysis. Antibiotics 12, 655 (2023).
-
Kannayiram, K. Molecular Docking analysis of flavonoids with aldose reductase. Bioinformation 18, 180–183 (2022).
-
Ji, X. et al. In Silico and in vitro antimalarial screening and validation targeting plasmodium falciparum plasmepsin V. Molecules 27, 2670 (2022).
-
Altunayar-Unsalan, C. & Unsalan, O. Molecular structure, sntioxidant potential, and Pharmacokinetic properties of plant flavonoid blumeatin and investigating its Inhibition mechanism on Xanthine oxidase for hyperuricemia by molecular modeling. ACS Omega. https://doi.org/10.1021/acsomega.3c10083 (2024).
-
Rosignoli, S. & Paiardini, A. Boosting the full potential of PyMOL with structural biology plugins. Biomolecules 12, 1764 (2022).
-
Ansori, A. et al. Bioactive compounds from mangosteen (Garcinia Mangostana L.) as an antiviral agent via dual inhibitor mechanism against SARSCoV- 2: an in Silico approach. Pharmacogn J. 14, 85–90 (2022).
-
Nithin, C. et al. Exploring protein functions from structural flexibility using CABS -flex modeling. Protein Sci. 33, (2024).
-
Wahyuni, D. K. et al. The antioxidant and antimicrobial activity of ethanolic extract in roots, stems, and leaves of three commercial Cymbopogon species. BMC Complement. Med. Ther. 24, 272 (2024).
Acknowledgements
This work was supported by Universitas Airlangga (Penelitian Unggulan Airlangga 2024, grant number 367/UN3.LPPM/PT.01.03/2024).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wahyuni, D.K., Rahmawati, C.T., Syukriya, A.J. et al. Metabolite characterization and in vitro antiplasmodial potential assessment of Sonchus arvensis L. callus under glutamine treatment. Sci Rep 15, 35956 (2025). https://doi.org/10.1038/s41598-025-12617-z
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12617-z