
Introduction
AAV-based gene therapies have shown success in treating retinal disease [1], spinal muscular atrophy [2], and hemophilia [3], leading to increased research and development for other diseases [4]. AAV vectors can deliver genes expressing therapeutic protein with sustained expression in target tissues, addressing the root cause of certain diseases [4]. When establishing preclinical safety and efficacy of AAV vector-based therapies, the biodistribution profile is one of the crucial determinants that shapes the decisive “go/no-go” verdict for the gene therapy product [5]. The choice of AAV serotype is guided by capsid tropism. AAV2 has applicability towards retina [6] and hepatocyte cell [7] transduction, AAV5 can target vascular endothelial cells [8] and non-mitotic central nervous system cells [9], while AAV8 exhibits delivery to skeletal muscle [10] and renal epithelium [11]. This article focuses on AAV9, which can partially cross the blood-brain barrier [12] and transduce various cell types in the central nervous system [13].
The AAV9 biodistribution tissue profile has been extensively studied in animal studies supporting clinical trials for spinal muscular atrophy, Parkinson’s disease, and lysosomal storage diseases [14]. Here, research grade biodistribution data were generated using cynomolgus macaques dosed with ssAAV9 or scAAV9 vectors. The constructs contained transgenes encoding the fluorescent reporter green fluorescence protein (GFP), mCherry, or the human SMN1 protein. Metadata analysis of vector genome (vg) concentration and transgene expression provided unique insights into transgene expression efficiency across various primate tissues. The ratio of transgene RNA or protein expressed per viral vector DNA unit within the same tissue sample was used to measure expression efficiency. The analysis encompassed biodistribution datasets of fluorescent reporter protein constructs in non-human primate (NHP) hippocampus, heart, and liver across multiple timepoints and ROAs, as illustrated in Fig. 1. The study also includes human SMN1 protein constructs in NHP liver, heart, and spinal cord across multiple timepoints and ROAs, as illustrated in Fig. 2. Interestingly, the expression efficiency in NHP liver was significantly lower compared to other tissues, regardless of viral construct, ROA, animal age, or dosing regimen. Mouse liver dosed with a similar viral construct did not display this trend of decreased transgene expression efficiency (Supplementary Fig. 1), consistent with literature tropism assessments of various AAV serotypes in C57BL/6 mice vs. cynomolgus macaques. [15]

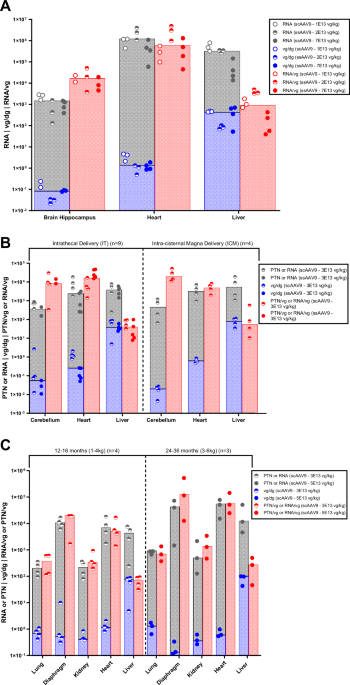
A Analysis of NHP (n = 10, ages: 6–50 months) hippocampus, heart, and liver tissues following IV delivery of ssAAV9 CMV173-mCherry or scAAV9-CBA-mCherry (dosed at 2.0E13–7.0E13 vg/kg). B The vector expression efficiency of ss or scAAV9-CBA-GFP (dosed at 3.0E13 vg/Animal) following different routes of administrations (IT, ICM) was assessed across NHP (n = 13, ages: 12–48 months) cerebellum, heart, and liver tissues. C Intrathecal administration of scAAV9-CBA-GFP (dosed at 3.0E13–5.0E13 vg/Animal) across younger (n = 4, ages: 12–16 months) and older (n = 3, ages: 24–36 months) NHPs. All studies had a 4-week evaluation period, and transgene expression was measured in cDNA copies per µg Total RNA, labelled RNA, or picogram (pg) mCherry/GFP per mg Total Protein, labelled PTN. Vector genome concentration was normalized to the endogenous diploid gene CFTR and labelled vg/dg. Transgene expression efficiency was assessed as GFP cDNA copies per µg RNA divided by vg per dg concentration, and pg of GFP per mg protein divided by vg per dg concentration. Bar graph overlay is used to highlight the median value for each grouped dataset, corresponding to RNA or PTN, vg/dg, and expression efficiency.

A The biodistribution assessment of NHP (n = 22, ages: 13–19 months) liver and heart tissues following IV (1.1E14 vg/kg) or IT (dosed at 1.2E13 vg/Animal) administration of scAAV9-CBA-SMN1 demonstrates reduced expression efficiency in liver tissue over the duration of the study periods. B Vector expression efficiency in spinal cord remained consistent over 6 weeks post IT delivery. C Vector expression efficiency post IV delivery was consistent over the 6-week interim endpoint and 26-week terminal endpoint. SMN1 vector genome, transgene expression, and expression efficiency ratio biodistribution datasets were assessed as described in Fig. 1.
It is widely known that systemic administration of AAV9 leads to major vector trafficking to the NHP liver [13, 16, 17] and that intracranial ROA also leads to significant delivery to the liver [18]. However, our findings show that elevated vg concentration in liver may not correlate with elevated transgene expression as would be seen in other tissues, irrespective of the vector sequence. To better understand the underlying mechanisms of reduced expression in NHP liver, epigenetic evaluation of in vivo viral DNA was performed. To this end, Cleavage Under Targets and Tagmentation (CUT&Tag) assessment was performed to investigate vector chromatin state association. Our findings, as shown in Fig. 3, indicate the potential involvement of transgene expression silencing and repression mechanisms.

A CUT&Tag assessment H3-K27me3 association with ssAAV9-CMV173-mCherry (IV delivery, 7.0E13 vg/kg, n = 4, ages: 25–50 months). For Animal 2 and Animal 3 liver biological replicates are displayed in tracks 1 through 4, and track 5 represents vector in Animal 5 heart (IV delivery, 1.0E14 vg/kg). B The H3-K9me3 histone association to vector in Animal 2 and Animal 3 liver biological replicates are displayed in tracks 1 through 4, and track 5 consists of H3-K9me3 association with vector in Animal 5 heart. C CUT&Tag assessment of ssAAV9-CBA-H3_HA (IV delivery, 5.0E13 vg/kg, n = 2, ages: 24–50 months) association to H3-K9me3 in Animal 1 and Animal 2 liver is displayed in tracks 1 and 2, while association to H3-K9me3 in Animal 1 and Animal 2 heart is displayed in tracks 3 and 4. Construct design map for the CUT&Tag assessment is provided under the gene coordinates for each figure.
Materials and methods
Animal test system
This analysis collated data from 51 NHPs (ages: 6–50 months) dosed with AAV9 across ten studies, as listed in Table 1. Of the ten studies, three used male NHPs, three used females, and the rest used both genders. The NHPs were sourced from Envigo Global Services, Inc. (Alice, TX), Mannheimer Foundation, Inc. (Homestead, FL), Bioculture Mauritius Ltd. (Immokalee, FL), and Worldwide Primates, Inc. (Miami, FL). The in-life portion of NHP studies was performed at Labcorp Early Development Laboratories Inc. (Madison, WI) and Mannheimer Foundation, Inc. (Homestead, FL). Animals were preselected for low pre-existing anti-AAV9 antibody titers. All in-life procedures, including dosing, monitoring, and terminal collection, were conducted following a consistent protocol across facilities. NHP animals were given cage enrichment devices and fruits, vegetables, and/or dietary enrichment.
Female C57BL/6 mice (age: 6 weeks) were dosed with AAV9 vector at a local facility, Explora BioLabs (San Diego). The study was approved by the Institutional Animal Care and Use Committee (IACUC) at the testing facility, and procedures were in accordance with an Animal Care and Use Protocol.
AAV9 vectors
All studies utilized vectors with genetic payloads consisting of self-complementary (4 studies) or single-stranded genome (6 studies). Expression of the transgenes GFP, mCherry, or human SMN1 was driven by CMV173 or chicken β-actin (CBA) promoters. The vectors were produced using preclinical research standards (for reporter genes) or Good Manufacturing Practice conditions (for GLP studies, including the CBA promoter with human cDNA). Dose formulations were prepared by diluting the respective drug substance in vehicle control article Tangential Flow Filtration [TFF] 3 buffer.
Dosing procedures
For IV administration, AAV vectors or control articles were dosed at the testing facility by slow IV infusion via saphenous vein (or alternatively via cephalic vein) over approximately 20 min using a motorized syringe press or infusion pump, followed by saline flush.
For IT and ICM administration, animals were anesthetized at the time of dosing (10.0 mg/kg of ketamine and dexmedetomidine) at the testing facilities. Animals were administered buprenorphine before anesthesia for ICM or after anesthesia for lumbar puncture (LP). Prior to dosing, 1.0 mL of CSF was collected from each animal. Doses were administered via ICM over 2–3 min or via lumbar puncture into the intervertebral space of L5 to L6 over at least 1 min, followed by artificial CSF flush. Immediately after dosing, IT injected animals were maintained in a Trendelenburg-like position for 10 min. Atipamezole (0.2 mg/kg for LP or 0.1 mg/kg for ICM) was administered as an anesthesia reversal agent.
In-life and terminal procedures
General daily observations included but were not limited to twice-daily mortality and distress checks and daily checks for behavioral abnormalities (health concerns, behavioral/activity level changes, postural changes, relative skin color changes, injuries, signs of pain or distress). Weekly detailed observations for signs of toxicity were also performed by removing the animals from their cages and obtaining body weights.
Prior to euthanasia, animals were anesthetized with sodium pentobarbital and exsanguinated. Tissue samples were collected, flash-frozen in liquid nitrogen, and sent to Novartis for analysis.
Biodistribution analysis
Vector genome biodistribution analysis was performed using droplet digital polymerase chain reaction (ddPCR) in accordance with the method specified in Hudry et al. [17] Sequences for all vg assay primers and probes used in ddPCR assays with a 60 °C annealing temperature are listed in Table 2. Vg values were normalized to the cystic fibrosis transmembrane conductance regulator (CFTR) diploid reference gene. Data points that did not meet a pre-established acceptance criteria of 18, 21, or 26 vg copies/ 20 μL reaction for GFP, mCherry or SMN1, respectively, were below the lower limit of quantification (BLQ) and omitted from data sets.
Two comparable methods were used for RNA purification and analysis. For studies F, G, I, and K the tissue samples were homogenized using TRI reagent (Zymo Research, R2050-1-200) and a TissueLyser II (Qiagen, 85300), then RNA was purified using the Direct-zol™ RNA MiniPrep Plus kit (Zymo Research, R2073), quantified using NanoDrop (Thermo Fisher, ND8000) and analyzed in triplicate using the One-Step RT-ddPCR Advanced Kit for Probes (Bio-Rad, 1864022) with both + and − reverse transcriptase (RT) reactions. The signal in the – RT reaction was used to subtract the background per sample. For studies A, B, C, tissue samples were homogenized using TRI Reagent and a TissueLyser II, then the organic phase was separated using Phasemaker™ tubes (Thermo Fisher, A33248) with chloroform. The aqueous phase was collected and processed using the Maxwell® RSC SimplyRNA tissue kit and Maxwell RSC 48 Instrument (Promega Corporation, AS1340). Purified RNA samples were treated with Turbo DNase (Thermo Fisher, AM2238) followed by ReliaPrep RNA Clean-up (Promega Corporation, Z1071) and quantified using NanoDrop. Samples were analyzed in triplicate using RT-ddPCR and a + RT reaction only. Sequences for all RNA assay primers and probes used in RT-ddPCR assays with a 62 °C annealing temperature are listed in Table 2. RNA concentration values were reported as cDNA copies per µg total RNA. Samples with insufficient RNA concentration for analysis were labeled as quantity not sufficient (QNS) and excluded from the data set.
For studies D and E, protein expression was quantified using Meso Scale Discovery (MSD) electrochemiluminescence immunosorbent assay (ECLIA) as previously described [19]. For the GFP ECLIA assay, capture GFP monoclonal antibody (3E6) (Invitrogen, A11120), detection GFP polyclonal antibody (Abcam, Ab290), and secondary MSD SULFO-TAG™ labeled polyclonal antibody (MSD, R32 AH) were used.
A subset of protein expression and vector genome data from studies D, F, and G has been previously reported [17, 19, 20] and was incorporated into the metadata analysis.
Statistical analysis
Statistical analyses were performed using GraphPad Prism 10. Sample size for each experimental group is provided in Figs. 1 and 2. Two-way ANOVA multiple comparison was used for each vg and RNA biodistribution dataset under assessment. A Wilcoxon and Mann-Whitney test was employed in Fig. 2A comparing the 6-week and 26-week RNA/vg datasets for liver tissues from IV dosed NHPs, and no statistically significant difference was observed (P value = 0.1320). Similarly, the IT dosed group assessment over 6-weeks and 52-weeks also showcased no statistically significant differences amongst the two datasets (P value = 0.9143). Variance amongst the IV dosed groups and the IT dosed groups was not significantly different according to an F-test (F-ratio approximation was 1.23 for IV, and 1.72 for IT datasets).
Ethical approval
All animal experiments were performed in accordance with the Guide for the Care and Use of Laboratory Animals and the Animal Welfare Act. Studies were approved by the Institutional Animal Care and Use Committee (IACUC) at each testing facility used, Labcorp Early Development Laboratories Inc. (Madison, WI), and Mannheimer Foundation, Inc. (Homestead, FL).
Results
NHP liver metadata analysis after IV, IT, or ICM administration of AAV9 reporter vectors
Vector distribution and expression results from three studies were combined. These studies involved NHPs (n = 10, ages: 6–50 months) administered with ssAAV9-CMV173-mCherry or scAAV9-CBA-mCherry (dose range: 2.0–7.0E13 vg/kg) via intravenous administration with a 4-week terminal endpoint. Insights into expression efficiency across different tissues (Fig. 1A) were obtained by studying the ratio of RNA expression to vg per diploid genome (dg). Although RNA levels were comparable among peripheral tissues, vector expression efficiency in NHP liver was strikingly lower when compared to brain and heart tissue despite vg concentration in liver being significantly higher than brain and heart tissues.
Biodistribution evaluation was also performed on NHP (n = 13, ages:12–48 months) brain, heart, and liver tissues following IT or ICM administration of ss or scAAV9-CBA-GFP. Lower reporter protein expression efficiency in NHP liver was observed following the two ROAs relative to cerebellum and heart peripheral tissue at different ages (12–16 months vs. 24–36 months) (Fig. 1B, C). Therefore, reduced reporter transgene expression efficiency in the NHP liver was observed following IV, IT, and ICM administration across different timepoints and ages compared to brain and heart.
Expression efficiency of scAAV9-CBA-SMN1 following IT or IV delivery
The duration of the evaluation period was assessed with a different vector to further evaluate the expression efficiency in NHP liver relative to other tissues. Liver and heart biodistribution of scAAV9-CBA-SMN1 following IV (1.1E14 vg/kg, n = 10, ages: 13–19 months) or IT administration (1.2E13 vg/Animal, n = 12, ages: 13–19 months) was assessed over two extended timepoints (Fig. 2A). Both groups shared a 6-week interim timepoint, while the IT and IV groups had 52- and 26-week terminal endpoints, respectively. The RNA expression per vg was lower in liver compared to heart at both early and late timepoints for both IT and IV ROAs. Vector expression efficiency in spinal cord was consistent over the entirety of the timepoints in IT and IV groups (Fig. 2B, C, respectively). Data from the early and late timepoints showed a consistent liver RNA/vg ratio and no statistically significant difference for liver for either the IV or IT ROA (P values > 0.05).
Epigenetic evaluation of ssAAV9-CMV173-mCherry and ssAAV9-CBA-H3_HA in NHP liver and heart after IV delivery
Epigenetic profiling was performed on NHPs dosed with ssAAV9-CMV173-mCherry (7.0E13 vg/kg, n = 4, ages: 25–50 months) or ssAAV9-CBA-H3_HA (HA-tagged histone H3) (5.0E13 vg/kg, n = 2, ages: 24–50 months) each with a 4-week endpoint. Vector DNA from liver and heart tissues was isolated and assessed for chromatin suppressor association, namely H3-K9me3 and H3-K27me3, using CUT&Tag analysis. Peak association intensity across the vector sequence coordinates was organized into 50 bp bins (Fig. 3A, B). The results indicate enrichment of H3-K9me3 and H3-K27me3 across the mCherry sequence. Vector DNA isolated from liver displayed significantly higher chromatin suppressive modification compared to heart tissue isolates, with a greater than 10-fold enrichment in H3-K9me3 and H3-K27me3 association. H3-K9me3 association was also evaluated via CUT&Tag (Fig. 3C) on in vivo vector DNA from NHPs dosed with a construct expressing a novel reporter protein, HA-tagged human histone H3C1, observing similar results. These findings demonstrate greater H3-K9me3 association to the transgene in liver tissue relative to heart tissue, further supporting evidence of transgene repression in NHP liver.
Discussion
The retrospective analysis of AAV9 biodistribution presented here includes ten NHP studies with a total of 51 animals. By combining vg per dg biodistribution with transgene RNA or protein expression, insights were gathered into transgene expression efficiency in several NHP tissues. The quantified RNA was presumed to be representative of each tissue under investigation, without considering the potential variation in transduction across different cell types or heterogeneity in epigenetic silencing amongst distinct cell populations, which can be further investigated. Other studies have showcased reduced transgene expression over time in cynomolgus hepatocytes via spatial genomics and ISH studies [21].
The metadata findings reveal that while AAV9 mediated transgene expression in the liver is observed, a consistent trend of lower transgene expression per vg in NHP liver compared to brain, spinal cord, and heart exists across multiple AAV9 study conditions. This phenomenon carries implications for the success of liver-directed gene therapies, as the liver may require a lower or higher percentage of transduction depending on the extent needed to treat the particular disease [22,23,24]. Hemophilia treatment, for example, requires only a relatively low level of transgene expression to achieve therapeutic benefit [23]. This modest threshold makes liver-directed gene therapy a promising strategy for hemophilia treatment, as even a small percentage (1–5%) of hepatocyte transduction can yield clinically meaningful outcomes [25]. Conversely, inherited metabolic diseases, such as urea cycle disorders or glycogen storage diseases, often demand a much higher percentage of hepatocyte transduction to correct systemic metabolites [22, 25]. In these cases, the dilution of episomal vector and a low amount of integrated vector during hepatocyte turnover and growth may create challenges, particularly for pediatric applications [24, 26].
Studies with longer evaluation periods have identified a trend of temporary expression silencing in extrahepatic tissues, which is partially rescued at a later timepoint by enduring episomal vector genomes [27, 28]. In contrast, the hepatic tissue does not exhibit a rescue in transgene expression, pointing to cell-type-specific epigenetic silencing factors unique to the cynomolgus liver [21, 29]. These factors potentially complicate the transduction process due to elements like host anti-viral defense mechanisms learned through the gut virome [30, 31], differences in vector hypermethylation driven by NP220 and HUSH availability in distinct cell-types [32], and the influence of viral serotype structure on transgene epigenetic silencing in liver [33]. The observed trend of reduced expression efficiency was absent in mouse liver, which aligns with previously reported findings Supplementary Fig. 1 [21]. Further studies are needed to confirm the interspecies variability in AAV9 transgene expression efficiency in liver tissues for other animal models.
Epigenetic profiling was used to assess the impact of chromatin modification on transgene expression silencing in NHP liver tissue. H3-K9me3 modification, associated with heterochromatin-mediated gene silencing [34], and H3-K27me3 modification, involved in repressing genes [33, 35, 36], were assessed by CUT&Tag. Analysis revealed a striking increase in H3-K9me3 and H3-K27me3 association with the mCherry transgene in NHP liver compared to heart tissue at a similar dose. CUT&Tag analysis of an additional construct (HA-tagged histone H3) also showed a similar trend of elevated H3-K9me3 association with the viral transgene in liver tissues compared to heart. Our metadata observations and epigenetic evaluations suggest increased silencing/repression of AAV9 transgene expression in cynomolgus liver tissue. These findings in AAV9-dosed NHP liver are consistent with those reported by Greig et al. [21] regarding complications of using AAV8 or AAVrh10 vectors in NHP liver for long-term transgene expression.
Viral gene therapy for inherited liver diseases would benefit from enhancing viral expression efficiency in primate liver. Although hepatocytes are transduced with AAV vector, [37] sustained in vivo gene expression is more challenging to achieve in primates than in mice. Grieg et al. [21] showed that vector expression decline occurs in NHP liver tissue within 2 to 6 weeks, with RNA expression decrease more pronounced than that of vector DNA loss [21]. Clinical evidence suggests this decline is unrelated to transgene immunogenicity or liver inflammation [38]. Overcoming this unique hurdle is critical for successful translation to clinical applications.
Despite the challenge that epigenetic silencing presents to liver gene therapy development, there are methods to mitigate chromatinization, although each carries associated drawbacks. For instance, histone deacetylase (HDAC) inhibitors increase rAAV gene expression [39], but a global reduction in silencing can lead to various unintended clinical toxicities [40]. Valproic acid enhances viral gene expression by promoting histone demethylation on episomal transgene DNA and influencing histone acetylation pathways [41], but excessive use is associated with therapeutic risks including hepatic steatosis and severe liver injury [42]. CpG motifs are primary targets for DNA methyltransferase activity, resulting in repression of transgene expression [43]; hence, exploration of functionally active CpG-free vectors is a promising strategy for preventing vector chromatinization and silencing [44]. While challenges remain, innovative approaches such as these continue to pave the way toward unlocking the full potential of liver gene therapies.
Data availability
All data generated in this study are included as part of the published article and supplementary information.
References
-
Testa F, Bacci G, Falsini B, Iarossi G, Melillo P, Mucciolo DP, et al. Voretigene neparvovec for inherited retinal dystrophy due to RPE65 mutations: a scoping review of eligibility and treatment challenges from clinical trials to real practice. Eye. 2024;38:2504–15.
-
Ogbonmide T, Rathore R, Rangrej SB, Hutchinson S, Lewis M, Ojilere S, et al. Gene therapy for spinal muscular atrophy (SMA): a review of current challenges and safety considerations for onasemnogene abeparvovec (Zolgensma). Cureus. 2023;15:e36197.
-
Shah J, Kim H, Sivamurthy K, Monahan PE, Fries M. Comprehensive analysis and prediction of long-term durability of factor IX activity following etranacogene dezaparvovec gene therapy in the treatment of hemophilia B. Curr Mes Res Opin. 2022;39:227–37.
-
Pupo A, Fernández A, Low SH, François A, Suárez-Amarán L, Samulksi RJ. AAV vectors: The Rubik’s cube of human gene therapy. Mol Ther. 2022;30:3515–41.
-
Assaf BT, Whiteley LO. Considerations for preclinical safety assessment of adeno-associated virus gene therapy products. Toxicol Pathol. 2018;46:1020–7.
-
Petrs-Silva H, Dinculescu A, Li Q, Deng W, Pang J, Min S, et al. Novel properties of tyrosine-mutant AAV2 vectors in the mouse retina. Mol Ther. 2011;19:293–301.
-
Logan G, Dane A, Hallwirth C, Smyth CM, Wilkie EE, Amaya AK, et al. Identification of liver-specific enhancer–promoter activity in the 3′ untranslated region of the wild-type AAV2 genome. Nat Genet. 2017;49:1267–73.
-
Chen S, Kapturczak M, Loiler SA, Zolotukhin S, Glushakova OY, Madsen KM, et al. Efficient transduction of vascular endothelial cells with recombinant adeno-associated virus serotype 1 and 5 vectors. Hum Gene Ther. 2005;16:235–47.
-
Pietersz KL, Martier RM, Baatje MS, Liefhebber JM, Brouwers CC, Pouw SM, et al. Transduction patterns in the CNS following various routes of AAV-5-mediated gene delivery. Gene Ther. 2021;28:435–46.
-
Charan RA, Niizawa G, Nakai H, Clemens PR. Adeno-associated virus serotype 8 (AAV8) delivery of recombinant A20 to skeletal muscle reduces pathological activation of nuclear factor (NF)-κB in muscle of mdx mice. Mol Med. 2012;18:1527–35.
-
Rubin JD, Barry MA. Improving molecular therapy in the kidney. Mol Diagn Ther. 2020;24:375–96.
-
Merkel SF, Andrews AM, Lutton EM, Mu D, Hudry E, Hyman BT, et al. Trafficking of adeno-associated virus vectors across a model of the blood-brain barrier; a comparative study of transcytosis and transduction using primary human brain endothelial cells. J Neurochem. 2017;140:216–30.
-
Saraiva J, Nobre RJ, Pereira de Almeida L. Gene therapy for the CNS using AAVs: the impact of systemic delivery by AAV9. J Control Release. 2016;241:94–109.
-
Kang L, Jin S, Wang J, Lv Z, Xin C, Tan C, et al. AAV vectors applied to the treatment of CNS disorders: clinical status and challenges. J Control Release. 2023;355:458–73.
-
Fang K, Yang X, Liu Y, Xia J, Wu R, Yang F, et al. A comprehensive study of AAV tropism across C57BL/6 mice, BALB/c mice, and crab-eating macaques. Mol Ther Methods Clin Dev. 2025;33:101434.
-
Giannelli SG, Luoni M, Iannielli A, Middeldorp J, Philippens I, Bido S, et al. New AAV9 engineered variants with enhanced neurotropism and reduced liver off-targeting in mice and marmosets. iScience. 2024;27:109777.
-
Hudry E, Aihara F, Meseck E, Mansfield K, McElroy C, Chand D, et al. Liver injury in cynomolgus monkeys following intravenous and intrathecal SCAAV9 gene therapy delivery. Mol Ther. 2023;31:2999–3014.
-
Hinderer C, Bell P, Vite CH, Louboutin JP, Grant R, Bote E, et al. Widespread gene transfer in the central nervous system of cynomolgus macaques following delivery of AAV9 into the cisterna magna. Mol Ther Methods Clin Dev. 2014;1:14051.
-
Meseck EK, Guibinga G, Wang S, McElroy C, Hudry E, Mansfield K. Intrathecal sc-AAV9-CB-GFP: systemic distribution predominates following single-dose administration in cynomolgus macaques. Toxicol Pathol. 2022;50:415–31.
-
Tukov FF, Mansfield K, Milton M, Meseck E, Penraat K, Chand D, et al. Single-dose intrathecal dorsal root ganglia toxicity of onasemnogene abeparvovec in cynomolgus monkeys. Hum Gene Ther. 2022;33:740–56.
-
Greig JA, Martins KM, Breton C, Lamontagne RJ, Zhu Y, He Z, et al. Integrated vector genomes may contribute to long-term expression in primate liver after AAV administration. Nat Biotechnol. 2023;42:1232–42.
-
Brunetti-Pierri N, Lee B. Gene Therapy for inborn errors of liver metabolism. Mol Genet Metab. 2005;86:13–24.
-
Nathwani AC, Reiss UM, Tuddenham EGD, Rosales C, Chowdary P, et al. Long-term safety and efficacy of factor IX gene therapy in hemophilia B. N Engl J Med. 2014;371:1994–2004.
-
Puzzo F, Kay MA. The delivered promises of gene therapy: past, present, and future of liver-directed gene therapy. Mol Ther. 2025;33:1966–87.
-
Chuecos MA, Lagor WR. Liver directed adeno-associated viral vectors to treat metabolic disease. J Inherit Metab Dis. 2024;47:22–40.
-
Piccolo P, Brunetti-Pierri N. Current and emerging issues in adeno-associated virus vector-mediated liver-directed gene therapy. Hum Gene Ther. 2025;36:77–87.
-
Herzog RW, Mount JD, Arruda VR, High KA, Lothrop CD Jr. Muscle-directed gene transfer and transient immune suppression result in sustained partial correction of canine hemophilia B caused by a null mutation. Mol Ther. 2001;4:192–200.
-
Duan D, Sharma P, Yang J, Yue Y, Dudus L, Zhang Y, et al. Circular intermediates of recombinant adeno-associated virus have defined structural characteristics responsible for long-term episomal persistence in muscle tissue. J Virol. 1998;72:8568–77.
-
Journou M, Devaux M, Jaulin N, Pichard V, Segovia M, Moreau A, et al. Immune-driven gene expression loss following intramuscular AAV delivery to non-human primates is only transient. Mol Ther Methods Clin Dev. 2025;33:101409.
-
Lang S, Demir M, Martin A, Jiang L, Zhang X, Duan Y, et al. Intestinal virome signature associated with severity of nonalcoholic fatty liver disease. Gastroenterology. 2020;159:1839–52.
-
Cao Z, Sugimura N, Burgermeister E, Ebert MP, Zuo T, Lan P. The gut virome: a new microbiome component in health and disease. EBioMedicine. 2022;81:104113.
-
Das A, Vijayan M, Walton EM, Stafford VG, Fiflis DN, Asokan A. Epigenetic silencing of recombinant adeno-associated virus genomes by NP220 and the HUSH complex. J Virol. 2022;96:e0203921.
-
Gonzalez-Sandoval A, Pekrun K, Tsuji S, Zhang F, Hung KL, Chang HY, et al. The AAV capsid can influence the epigenetic marking of rAAV delivered episomal genomes in a species dependent manner. Nat Commun. 2023;14:2448.
-
Wang Z, Fan R, Russo A, Cernilogar FM, Number A, Schirge S, et al. Dominant role of DNA methylation over H3K9me3 for IAP silencing in endoderm. Nat Commun. 2022;13:5447.
-
Cai Y, Zhang Y, Loh YP, Tng JQ, Lim MC, Cao Z, et al. H3K27me3-rich genomic regions can function as silencers to repress gene expression via chromatin interactions. Nat Commun. 2021;12:719.
-
Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, et al. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–26.
-
Palazzi X, Pardo ID, Sirivelu MP, Newman L, Kumpf SW, Qian J, et al. Biodistribution and tolerability of AAV-PHP.B-CBh-SMN1 in Wistar Han rats and cynomolgus macaques reveal different toxicologic profiles. Hum Gene Ther. 2022;33:175–87.
-
D’Antiga L, Beuers U, Ronzitti G, Brunetti-Pierri N, Baumann U, Di Giorgio A, et al. Gene therapy in patients with the Crigler–Najjar syndrome. N Engl J Med. 2023;389:620–31.
-
Okada T, Uchibori R, Iwata-Okada M, Takahashi M, Nomoto T, Nonaka-Sarukawa M, et al. A histone deacetylase inhibitor enhances recombinant adeno-associated virus-mediated gene expression in tumor cells. Mol Ther. 2006;13:738–46.
-
Subramanian S, Bates SE, Wright JJ, Espinoza-Delgado I, Piekarz RL. Clinical toxicities of histone deacetylase inhibitors. Pharmaceuticals. 2010;3:2751–67.
-
Fan S, Maguire CA, Ramirez SH, Bradel-Tretheway B, Sapinoro R, Sui Z, et al. Valproic acid enhances gene expression from viral gene transfer vectors. J Virol Methods. 2005;125:23–33.
-
He X, Yuan R, Chen Y, Huang W, Xu Z, Wang B, et al. Mechanism of valproic acid-induced hepatic steatosis via enhancing NRF2-FATP2-mediated fatty acid uptake. Theranostics. 2025;15:5258–76.
-
Chanda D, Hensel JA, Higgs JT, Grover R, Kaza N, Ponnazhagan S. Effects of cellular methylation on transgene expression and site-specific integration of adeno-associated virus. Genes. 2017;8:232.
-
Subkhankulova EO, Tolmachov T. Silencing of transgene expression: a gene therapy perspective. Gene Ther. Tools Potential Appl. https://doi.org/10.5772/53379 (2013).
Acknowledgements
We thank Biologics Research Center (BRC), Pharmacokinetic Sciences (PKS), and PreClinical Safety (PCS) colleagues for collaborations. Studies were funded by Novartis Gene Therapies, Inc.
Funding
This research study was conducted without receiving any financial assistance or sponsorship from external organizations or entities. All costs associated with the study, including data collection, analysis, and publication, were covered by Novartis Biomedical Research.
Ethics declarations
Competing interests
M.S., J.R.S., T.d.R., and F.O. are all employees of Novartis Biomedical Research. All authors own Novartis stock or other equities.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Shahrukh, M., Sweeney, J.R., del Rio, T. et al. Metadata assessment of non-human primate studies of AAV9 uncovers potential tissue specific variation in expression efficiency. Gene Ther (2026). https://doi.org/10.1038/s41434-025-00589-8
-
Received:
-
Revised:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41434-025-00589-8