Introduction
Production of nutritious and safe food in sufficient quantities for ensuring food availability and nutritional security for the expected 9.8 billion people in 2050 is one of the ultimate goals of modern agriculture. Recently there is growing awareness of the relationship between diet and health, leading to an increasing demand for food products which support health beyond providing basic nutrition. Sustainable agricultural growth and eradication of hunger and poverty, ensuring healthiness and safety are the first three of the UN sustainable development goals1. Vitamins and mineral deficiencies (VMDS) – particularly of vitamin A, iron, iodine, zinc and folate pose a public health problem and affects over 2 billion people globally2. Micronutrient malnutrition, often called hidden hunger by the WHO3is a major problem and a challenge to researchers, public health officials and policy makers worldwide4. Cereals comprise the staple food for the majority of the world population and represent an important source of carbohydrates, proteins, fibers, minerals, vitamins, and phytochemicals and their regular consumption appears to be associated with many health benefits5.
Zinc is an essential mineral for humans, being the integral part of many enzymes6 and important for growth, cell differentiation, metabolism and disease responses. Zinc is the only metal that is part of all the six important enzyme classes7. In addition to this zinc is known as an important co-factor for 300 enzymes like alcohol dehydrogenase, RNA polymerases, carbonic anhydrase, superoxide dismutase, and involved in the synthesis of lipids, carbohydrates, proteins and nucleic acids8,9. The human body does not contain storage system for zinc and thus dietary intake is crucial for maintaining the exchangeable zinc pool, especially during childhood, adolescence and pregnancy10. Iron and Zinc are the most prevalent micronutrients deficient in the diet11affecting about a quarter of the world’s population, particularly women and children in developing countries12. Similarly, it is estimated by the WHO that approximately 35% of children between 0 and 5 years of age suffer from Fe and zinc deficiency13. Zinc deficiency alone accounts for approximately 1.9% of the global disease burden14.
Wheat (Triticum aestivum L.) belonging to family poaceae is the staple crop for majority of world population15including Pakistan16. Wheat plays critical role in ensuring global food security and ranks fifth in provision of global food calories and proteins17,18. Despite being an important component of human diet, wheat grains have a low concentration of micronutrients19 including Zinc (Zn) and Iron (Fe)20,21. The cereal based diets are not only low in zinc content but also contain higher amount of substances, like phenols and phytic acids which substantially reduce its bioavailability and absorption. Thus, in areas where wheat is consumed as a major dietary component, micronutrient malnutrition is prevalent. In addition, the milling of wheat grains into white flours usually removes the zinc -rich parts (mainly the embryo and aleurone layer) retaining only the zinc-poor endosperm, which contains about 5–10 mg/kg zinc22 depending on the extraction rate, and thus, cannot meet dietary zinc requirement (15–30 mg/kg in flour)23. Therefore, an increase in the wheat quality, such as the improved micronutrient is also essential beside an increase in the calories required to feed the growing population. Various possible techniques to overcome the micronutrient deficiency have been suggested including agronomic biofortification, a major agricultural strategies to enhance the grain concentration of micronutrients24,25,26. Isotopically exchangeable Zn in the calcareous soils of Pakistan is extremely low. Being an essential micronutrient for plant growth and development, its deficiency negatively impact crop yield and quality. Similarly, the high pH, low organic matter and coarse texture of the calcareous soils make zinc availability is limited, impacting plant uptake.
Microbial assisted biofortification involves the use of rhizospheric and endophytic bacteria for increasing the micronutrient concentration in the edible part of the plants. Rhizobacteria also assists in plant growth, development and consequently improve the yield in cereal crops27. The rhizobacteria have been reported to promote plant growth after colonizing the roots through different pathways6. The rhizobacteria may develop a synergistic interaction with plants and promoting its growth are termed as plant growth promoting rhizobacteria (PGPR). These bacteria give benefits to the plants either through nitrogen fixation, mobilization of mineral nutrients or the production of plant hormones. PGPR have the potential to reduce the application of chemical fertilizers and promote plant growth in a sustainable manner and offset the economic and environmental cost of the conventional farming. Several rhizobacteria have been reported to solubilize zinc in the soil and make it available for plant growth. Zn solubilizing rhizobacteria are involved in solubilization of insoluble zinc through production of different organic acid, proton exclusion and lowering of soil pH28. Further, these bacteria may also be involved in the expression of different zinc transporter genes29. Thus, augmenting the soils with acclimatized native Zn solubilizing rhizobacteria (ZSB) can be a promising approach to sustainably improve Zn nutrition of crops under deficient soils. However, microorganisms differ significantly in establishing a mutual partnership with their phytobionts depending on the crops and their varieties, soil types, and other environmental parameters30. Therefore, the mechanism and optimum inoculation conditions for root colonization and synergism with the host plant need elaboration to ensure reproducibility under different soil and environmental conditions.
In the present study, we reported the in vitro zinc solubilization potential of three bacterial strains and their synergistic interaction with roots of wheat genotypes, bacterial assisted zinc uptake, translocation and grains accumulation as well as the expression of zinc transporter genes under low zinc, supplementation of ZnSO4 or inoculation with rhizobacteria for developing a sustainable biofortification strategy.
Materials and methods
Experimental layout and location
The experiment was conducted at the Institute of Biotechnology and Genetic Engineering (IBGE), The University of Agriculture Peshawar Khyber Pakhtunkhwa Pakistan. The experiment was carried out in pots using 2-factorial completely randomized design (CRD) under greenhouse conditions.
Bacterial source
Three rhizobacterial strains, isolated from fields previously cultivated with maize in Dir, Khyber Pakhtunkhwa and identified for their plant growth promotion potential has been used during this study31. The selected bacterial species were characterized by traditional phenotypic and biochemical methods as well as 16 S rRNA gene sequencing and exhibited good plant growth promotion potential under in vitro conditions. These pre-characterized bacterial strains were used to study the kinetics of zinc solubilization and their synergism with different wheat genotypes. Furthermore, these rhizobacterial strains were used to study the deviation in zinc uptake, transportation and accumulation from the control plants as well as the expression of selected zinc transporters in the wheat.
In vitro study of zone of solubilization
In vitro study was performed to determine zinc solubilization potential of the selected rhizobacteria in semi-solidified tris minimal media containing 0.1% Zinc oxide (ZnO) as a source of non-soluble form of Zn (D-glucose–10 g, Tris HCl-6.06 g, NaCl-4.68 g, KCl-1.49 g, NH4Cl-1.07 g, Na2SO4−0.43 g, MgCl2.2H2O-0.2 g, and CaCl2.2H2O–30 mg). Different concentrations of bacterial inocula (6, 12 and 18 µl) from a culture with OD600 of 0.8 were applied to a well on plates. Zinc solubilization was checked by measuring the diameter of the halo zone after 1, 4 and 7 days of inoculation at 28oC in dark. The experiment was performed in triplicate.
Measurement of Gluconic acid and modulation of culture pH by the bacterial isolates
To detect and quantify gluconic acids following bacterial inoculation, tris-minimal salt broth was supplemented with 0.1% (w/v) Zn and autoclaved at 121 °C for 15 min. Starter cultures of each strain were prepared by culturing overnight in LB media to achieve an OD600 of 0.5 and 1 ml of starter culture was used to inoculate 100 ml of medium. Flasks were incubated at 28 °C in the dark and agitated at 100 rpm for up to 7 days. Samples of culture for each strain were withdrawn after 7 days for measurement of gluconic acid content. For determining the potential of gluconic acid production by the different bacterial isolates, 5 ml was centrifuged at 12,000 rpm for 20 min. A filter-sterilized culture filtrate was collected and analyzed using the HPLC system (Model e2695, Water, USA). The chromatographic separation of gluconic acid was achieved at ambient temperature using an Agilent ZORBAX SB-Aq column (4.6 × 300 mm ID, 5 μm particle size; Agilant Technologies, USA). Similarly, the clear supernatant was used for determination of pH using a pH meter (Orion model 250 A).
Scanning electron microscope (SEM)
The bacterial biofilm formations were observed through SEM30,32. For this purpose, wheat genotype cv Atta Habib, Sirin and Pak China were uprooted after 15 days of inoculation with bacterial strain DJ13 and sectioned with sterilized scissor into 1–2 cm pieces. Root samples were washed with 2.5% glutaraldehyde. The samples were then dehydrated with a series of ethyl alcohol (5, 10, 20, 50, 70, 90 and 100%). Finally the root samples were coated with gold for visualization with SEM.
Inoculum preparation seed treatments and maintenance of wheat genotypes
Wheat Zn uptake, translocation and grain accumulation after inoculation with the selected bacterial strains, with ZnSO4.7H2O application33 or without Zn was checked in a pot experiment in the Greenhouse of the Institute of Biotechnology and Genetic Engineering, The University of Agriculture Peshawar, Pakistan. Eight elite wheat genotypes Atta-Habib, Sirin, Pak-China 17, Zincol-16, Shahkar-13, Khaista, Pir Sabak-2013 and Pir Sabak-2015 were used during this experiment. The experiment was arranged in a completely randomized design with 3 replications. The control plants were grown under similar conditions but were not supplemented with bacteria or Zn fertilizer. Rhizobacterial cultures were initiated in aluminum foil covered flask containing LB medium in an orbital shaker at 120 rpm at 28 ± 2 °C in the dark until an OD600 of 0.8-1.0 was obtained34. Approximately 5.0 ml of the bacterial culture was harvested by centrifugation and suspended in 1% agar, mixed with 100 g surface sterilized wheat seeds of different genotypes and air dried in a laminar flow hood. Eight seeds were planted and five plants were maintained in each pot.The recommended dose of nitrogen, phosphorus and potassium (120: 90: 60 kg ha−1, respectively) were applied. Half doses of nitrogen and full dose of phosphorus and potassium were applied at the time of sowing, while remaining nitrogen was applied at booting stage with irrigation water. Routine plant management and irrigation were followed throughout the experiment.
Determination of mineral nutrients by atomic absorption spectroscopy
The micronutrient concentrations of samples from during the grain formation stage (root, shoot and leaves) or after threshing (grains) were determined by atomic absorption spectrometry35,36. For determination of mineral nutrient content, the samples were collected, washed with tap water and dried in an oven at 70oC for 5 h. After oven drying, 0.2 g of the sample was ground and digestion was obtained by mixing HNO3:HClO4 (3:1, v/v). The mixture was heated on hot plate until the solution became transparent. The digestate was diluted to 50 ml with distilled water and filtered through Whatman 42 filter paper and analyzed for total Zn concentration using atomic absorption spectrophotometer37. Zn translocation from root to shoot and accumulation index in grains was determined by the formula:
$${rm Z_{n}: Translocation: = Z_{n} :content :in :shoot / (Z_{n}: content :in :shoot + Z_{n}: content: in: root)}$$
$${rm Z_{n} :Accumulation: index = Z_{n} :content: in: grains / Z_{n} :content: in: shoot}$$
.
Expression analysis of Zn transporter and transcription factor genes
The expression and accumulation of transcripts of the different ZIP transporter and bZIP transcription factor genes were checked through the semi-quantitative reverse transcriptase PCR using specific primers (Table 1). Wheat, Actin-2 was used a loading control for normalization of expression. The wheat genotypes were grown in a hydroponic culture for 15 days and the expression of different genes was measured 4 days after addition of ZnO along with inoculation of Zn solubilizing bacteria (DJ13), ZnSO4 or without Zn supplementation. Samples were obtained with a sterile scissor and immediately stored at −80ºC until further processing. About (100 to 150 mg) of leaves samples were cut into small pieces, followed by crashing in liquid nitrogen38. Trizol (1 ml) was added to samples for obtaining homogenous mixture then about 200µL chloroform was added and vortexed for proper mixing. Incubation of solution was made on ice for 15 min, then centrifugation was carried out on 12,000 rpm for 15 min at 4ºC (Model Thermo scientific sorval ST16). The aqueous phase (400 µl) was taken into a fresh tube and 500 µl isopropanol was added to the samples. Incubation was repeated on ice for 10 min, the samples were centrifuged for 10 min on 12,000 rpm at 4ºC. After centrifugation the pellet was washed with 1 ml of 70% ethanol by flicking carefully while supernatant was removed. Final centrifugation was performed on 10,080 rpm for 10 min at 4ºC then the supernatant was discarded. RNA was air dried, and then 50 µl of RNase free water was added to the samples. RNA storage was carried out at −80ºC. The RNA concentration and purity was determined through spectrophotometric analysis with Nano Drop (Model Thermo scientific NANODROP 2000).
Total extracted RNA (1 µg) was reverse transcribed for cDNA synthesis using Thermo Fisher Revert Aid cDNA synthesis Kit. About 20 µl reaction mixture was prepared for the first strand cDNA synthesis. 1 µl of oligo (dT) primer, 1 µg of RNA and required amount of RNase free water was mixed carefully. The incubation of reaction mixture was performed at 65ºC for 5 min and was cooled on ice, then 1 µl RNase inhibitor, 4 µl reaction buffer, 1 µl of Revert Aid and 2 µl dNTP were added for making 20 µl mixture. Brief centrifugation and incubation were carried out at 42ºC for 1 h and termination at 70ºC for 5 min. Using gene specific PCR the final product obtained during cDNA synthesis was used as a template. 1 µl of cDNA was used in performing PCR with gene-specific primers.
Data analysis
Mean ± standard deviation of the bacterial Zn solubilization zone and medium pH were calculated using Microsoft Excel. Agronomic and biochemical data regarding various parameters were analyzed according to 2-factorial CRD using Origin 2018 software. Upon obtaining significant differences, least square difference (LSD) test was used for classification of genotypes, treatments and their interactions into different groups. The significant levels for all the parameters were set at p < 0.05.
Results
Zn solubilization activity
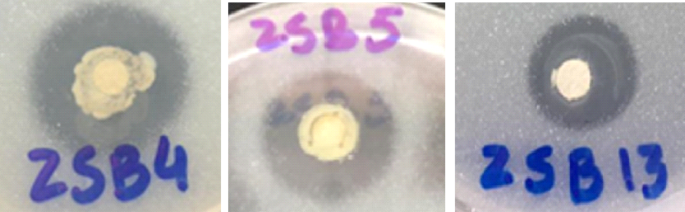
The appearance of halo zone on media supplemented with insoluble form of zinc exhibited the presence of specific zinc solubilizing activity of the bacterial strains (Fig I). Furthermore, mean value of halo zone diameters for different isolates showed that maximum Zn was solubilized by DJ13 (1.46 cm) followed by DJ4 and DJ5 (1.03 cm, respectively). Similarly, halo zone diameter increased with the days after inoculation in all the three isolates. Maximum halo zone (1.90 cm) which exhibited maximum Zn solubilization was noted in DJ4 after 7 days of inoculation (Table 2). In addition, maximum Zn solubilization was noted in DJ13 that was evident with a halo zone of 1.58 cm, followed by DJ4 (1.43 cm) and DJ5 (1.33 cm) after inoculation with bacterial strains.
Production of gluconic acid by rhizobacterial strains
Gluconic acid is supposed to play a predominant role in the inorganic nutrients solubilization activity of different rhizobacterial strains, and it was hypothesized that it also has an impact on the Zn solubilization activity of these strains. HPLC analysis of the gluconic acid standard revealed a peak after 2.49 min of filtration. Analysis of the culture filtrates exhibited 3 peaks after 1.96, 2.20 and 2.50 min of elution, the last two peaks representing glucuronic and gluconic acids, respectively. Peak height of gluconic acid chromatograph of bacterial strains indicated that DJ4 could produce 195.56 µg·l−1, DJ5 198.11 µg·l−1 and DJ13 could produce the maximum 242.34 µg·l−1 gluconic acid in the presence of insoluble Zn (Fig. 2). The higher production of gluconic acid by the isolates support the potential of a higher Zn solubilization as well as synergism with host plants by suppressing plant immunity. Furthermore, HPLC peak at retention time of 2.20 min indicated the production of 71.23 98.41 and 108.08 µg·l−1 glucuronic acid by DJ4, DJ5 and DJ13, respectively in the presence of insoluble Zn.

Halo zone formation by three bacterial strains (Pseudomonas aeruginosa (DJ5), Pseudomonas fluorescens (DJ13) and Pseudomonas lurida (DJ4).

High performances liquid chromatography (HPLC) chromatogram for gluconic acid standard or production by DJ4, DJ5 and DJ13, respectively.
Medium pH variation after different days by the bacteria
Consequent to the qualitative and quantitative assays all the three isolates were found to be efficient zinc solubilizers and HPLC confirmation of gluconic and glucuronic acids production, it was assumed that pH decrease could play a crucial role in the solubilization process. To measure the effect of bacterial inoculation on medium pH, they were grown on tris minimal broth in the presence of ZnO and the decline in pH was measured. Maximum release of soluble Zn occurred in the liquid medium supplemented with zinc oxide at the lowest mean pH value of 4.63, suggesting that solubilization is dependent on reduction of pH by organic acid production (Fig. 3). Furthermore, the higher gluconic and glucuronic acids production and lowest mean pH value of the liquid medium supplemented with ZnO after inoculation with DJ13 suggested that solubilization is reliant with pH reduction due to production of organic acid. Therefore, this bacterium was used in the further experiments to determine root colonization, zinc content and partitioning in wheat tissues.

Medium pH variation by the bacteria at different days after inoculation of media supplemented with ZnO.
Scanning electron microscope (SEM) analysis of wheat root colonization by DJ13 ( Pseudomonas fluorescens )
An effective plant growth promoting rhizobacteria has to colonize the roots and establish a synergistic relationship, besides persisting in the soil against autochthonous microorganisms living in environment through its rhizo-competence traits. During this study, the colonization and biofilm formation by the most potent ZSB strain DJ13 on roots of wheat genotypes (Atta Habib, Sirin and Pak China) was determined through Scanning electron microscopy (SEM). SEM light micrographs of the root surface (adjacent to the root tips) of all the three wheat genotypes revealed colonization by DJ13. Both individual cells and colonies of the rod shaped cells were noted on the root surface of all the three genotypes. However, no bacterial colonies were observed on the root surface of the plants maintained without inoculum application in hydroponic culture (Fig. 4).

SEM microgram of root surface of wheat seedlings of genotypes Atta Habib, Sirin and Pak China seven days after inoculation. (A, B and C) SEM micrograms of wheat genotypes Atta Habib, Sirin and Pak China respectively inoculated with P. fluorescens isolate DJ13; (D, E and F) Root surface of non-inoculated seedlings of the same genotypes but not inoculated with the rhizobacteria.
Zinc content in different tissues of wheat genotypes
The tissue Zn content of the eight wheat genotypes exhibited significant variation when grown in pots without zinc supplementation. Under these conditions, the root zinc content ranged between 29.64 mg·kg−1 (Pir Sabak 2013) to 43.32 mg·kg−1 (Sirin). However, root zinc content does not necessarily translated into a higher shoot or leaves zinc content, exhibiting variation in genotypic potential for translocation. Though Zincol 2016 has a moderate zinc content in the roots and shoots (29.85 and 31.83 mg·kg−1, respectively), it accumulated a maximum zinc in the leaves (41.16 mg·kg−1), exhibiting higher potential for sequestering zinc in the leaves. Consequently, lowest zinc accumulation in the wheat grains was noted in this genotype (21.12 mg·kg−1) when grown in pots under control conditions without zinc supplementation (Fig. 5). Application of ZnSO4.7H2O to the pots significantly increased the zinc content in different plant tissues (Fig. 6). Root zinc content in the wheat genotypes ranged between 44.59 (Zincol 2016) to 48.31 mg·kg−1 (Sirin) under zinc sufficiency conditions. Sirin also sequestered the highest concentration in the leaves (58.31 mg·kg−1) but not translated into accumulation in the grains (47.69 mg·kg−1). In the conditions of sufficient zinc provision, Zincol 2016 exhibited the highest potential to accumulate zinc in the grains (42.48 mg·kg−1). However, the differences in the amount of zinc in different plant tissues show a variable potential of zinc uptake, translocation and accumulation in the grains of wheat genotypes.

Tissue zinc content and relative distribution in different tissues (roots, shoots and grains; blue, orange and grey colours, respectively) of wheat genotypes when grown in pots without zinc supplementation.

Tissue zinc content and relative distribution in different tissues (roots, shoots and grains; blue, orange and grey colours, respectively) of wheat genotypes after supplementation with ZnSO4.
Inoculation with rhizobacterium DJ13 also resulted in a significant increase in the tissue zinc content as well as distribution. Maximum root zinc content was noted in Sirin (53.01 mg·kg−1) followed by Pir Sabak 2013 (37.25 mg·kg−1). Roots of Zincol 2016, the control variety, contained 39.59 mg·kg−1 zinc (Fig. 7). However, unlike Pir Sabak 2013, which translocated a high amount of zinc to the shoot, translocation was less in Sirin and Zincol 2016 (36.510 and 43.86 mg·kg−1, respectively). Highest shoot zinc content was noted in Shahkar (54.00 mg·kg−1). Leaves zinc was relatively stable in the wheat genotypes except Shahkar, Pak China and Zincol 2016 exhibiting higher content (51.54, 49.39 and 43.29 mg·kg−1, respectively). During this study the highest accumulation of zinc in the grains was noted in Pak China and Zincol 2016 (43.34 and 57.87 mg·kg−1, respectively).

Tissue zinc content and relative distribution in different tissues (roots, shoots and grains; blue, orange and grey colours, respectively) of wheat genotypes after inoculation with rhizobacterium DJ13.
Dynamics of Root to Shoot translocation and Zn accumulation index in wheat genotypes under different treatment conditions
Data on root-to-shoot zinc (Zn) translocation in various wheat genotypes under different treatment conditions revealed maximum variation in Zincol-2016 and Shahkar, while Pak China and Pir Sabak-2015 exhibited less variability (Fig. 8A). The highest Zn translocation from root to shoot was recorded in the genotype Sirin, achieving a value of 2.17 ± 0.06 after incubation with DJ5, followed closely by the same genotype after ZnSO₄ supplementation, which yielded 2.05 ± 0.02. Conversely, the lowest translocation (0.76 ± 0.02) occurred in Pir Sabak-2013 following incubation with DJ13. Additionally, one outlier above the 1.5 interquartile range (IQR) for Zn translocation was identified in Pak China cultivated under control conditions (1.38 ± 0.05), while two outliers below the 1.5 IQR were noted: one in Pak China after DJ13 inoculation (0.89 ± 0.02) and another in Pir Sabak-2015 under control conditions (1.18 ± 0.03). Grain zinc accumulation is a primary objective for wheat biofortification and serves as a critical target for both breeding and agronomic interventions. The translocation of zinc to the grains is influenced by the source-sink relationship during the grain-filling period, which may also interact with photo-assimilate dynamics. Variable grain zinc accumulation was observed among different wheat genotypes under various treatment conditions, with the greatest variation noted in wheat genotypes Pak China, Shahkar and Khaista. Specifically, maximum zinc accumulation was achieved in Pak China (0.635 ± 0.015) and Khaista (0.608 ± 0.014) following inoculation with strain DJ4. Conversely, the lowest grain zinc accumulation (0.308 ± 0.014) was recorded in Shahkar after inoculation with DJ13 (Fig. 8B).

Effect of different treatments on Zinc translocation from root to shoot (A) Grain Zn accumulation (B) of eight wheat varieties at maturity.
Effect of bacterial inoculation on expression of different zinc transporters and b ZIP transcription factor in wheat genotypes
The ZIP (Zn-regulated, iron-regulated transporter-like protein) transporter plays an important role in regulating the uptake, transport, and accumulation of microelements in plants. The expression of eight ZIP genes was examined in the leaves of wheat genotypes. A relatively similar expression of ZIP transporter and bZIP family genes was reported in the root and shoot of wheat. Significant modulation was noted in the expression of the TaZIP genes in the leaves of various wheat genotypes by different treatments compared to control plants in this study (Fig. 9).

Transcript abundance of different TaZIP and TabZIP transcription factor in leaves of wheat varieties under different treatment conditions.
TaZIP1 expression was induced in Pak China, Zincol-2016 and Shahkar following ZnSO₄ supplementation and Shahkar and Pir Sabak-2013 after inoculation with DJ13. Maximum TaZIP1 expression was noted in the leaves of the wheat genotype Zincol-2016 increasing 9.0-fold following ZnSO₄ treatment, while Pak China exhibited a 6.1-fold increase under the same conditions. In contrast, a decrease in TaZIP1 transcript levels was observed in Pir Sabak-2015 after DJ13 inoculation and in Atta Habib, Khaista, Pir Sabak-2013 and Pir Sabak-2015 after ZnSO₄ treatment (Fig. 9A). The expression of the TabZIP3 gene was differentially induced in the leaves of wheat genotypes 14 days after exposure to various treatment conditions (Fig. 9B). The highest expression of TabZIP3 was observed in Pak China following ZnSO₄ supplementation, with an increase of 13.2-fold, while Zincol-2016 exhibited a 4.1-fold increase under the same treatment. Additionally, significant induction of TabZIP3 (4-fold increase in transcript levels) was recorded in Pak China after inoculation with the rhizobacterial strain DJ13. Overall, inoculation with DJ13 led to the induction of the TabZIP3 gene in four genotypes: Pak China, Khaista, Shahkar, and Pir Sabak-2013. In contrast, ZnSO₄ supplementation resulted in increased TabZIP3 transcript levels in three genotypes: Pak China, Zincol-2016, and Shahkar.
The various treatment differentially induced expression of TaZIP4 gene in the leaves of wheat genotypes and the highest expression of TaZIP4 was observed in Shahkar following ZnSO₄ supplementation, with a 3.9-fold increase, while Khaista exhibited a 3-fold increase after inoculation with the rhizobacterial strain DJ13 (Fig. 9C). Overall, inoculation with DJ13 led to significant induction of the TaZIP4 gene in four genotypes: Khaista, Zincol-2016, Shahkar, and Sirin. Similarly, ZnSO₄ supplementation also resulted in an increase in TaZIP4 transcripts in four genotypes: Shahkar, Zincol-2016, Sirin, and Pak China. The expression of the TaZIP5 gene was significantly induced by DJ13 treatment in leaves of 4 wheat genotypes; Atta Habib, Sirin, Pak China and Pir Sabak 2013. Similar, higher expression of TaZIP5 relative to control conditions was noted in 3 wheat genotypes; Atta Habib, Pir Sabak 2013 and Pir Sabak 2015 after supplementation with ZnSO4. The highest relative expression of TaZIP5 was observed in wheat genotype Sirin inoculated with DJ13, showing a 5.9-fold increase over the control plants. Similarly, TaZIP5 expression in Pir Sabak-2015 exhibited a 4.1-fold increase in comparison with control plants after ZnSO₄ supplementation. Conversely, a decrease in TaZIP5 transcript levels was recorded in Zincol-2016, Shahkar and Khaista after inoculation with DJ13 or application of ZnSO4 (Fig. 9D).
The expression of the TaZIP7 gene in the leaves of various wheat genotypes was significantly influenced by different treatment conditions. The highest relative expression of TaZIP7 was observed in Shahkar, which exhibited a 6.2-fold increase over the control plants following ZnSO₄ supplementation, while Pir Sabak-2013 demonstrated a 2.4-fold increase after inoculation with the rhizobacterial strain DJ13. Compared to control plants, TaZIP7 expression was induced in Atta Habib, Zincol-2016, Shahkar, Khaista, and Pir Sabak-2013 following DJ13 inoculation; however, minimal expression was noted in Sirin, Pak China, and Pir Sabak-2015. Conversely, an increase in TaZIP7 transcripts was observed in Pak China and Shahkar after ZnSO₄ supplementation, while a repression of transcripts occurred in Atta Habib, Sirin, Zincol-2016, Khaista, Pir Sabak-2013, and Pir Sabak-2015 under the same treatment conditions (Fig. 9E). The expression of TaZIP9 transcripts in the leaves of eight wheat genotypes was significantly influenced by ZnSO₄ supplementation and rhizobacterial inoculation. Notably, TaZIP9 exhibited the highest expression in Pir Sabak-2013, with a 4.2-fold increase, followed by Shahkar, which showed a 3.4-fold increase after ZnSO₄ treatment. Additionally, Khaista and Shahkar demonstrated elevated TaZIP9 expression following inoculation with the rhizobacterial strain DJ13. In contrast, the lowest expression levels were recorded in Atta Habib and Sirin after ZnSO₄ supplementation. Overall, TaZIP9 transcription was induced in Shahkar, Khaista, and Pir Sabak-2013 following DJ13 inoculation, as well as in Shahkar, Khaista, Pir Sabak-2013, and Pir Sabak-2015 after ZnSO₄ treatment (Fig. 9F).
The expression of the TaZIP13 gene in the leaves of eight wheat genotypes was significantly influenced by different treatment conditions. Notably, the highest expression was observed in Pak China, with a 13.5-fold increase following ZnSO₄ supplementation, and in Khaista, which exhibited a 12-fold increase after inoculation with the rhizobacterial strain DJ13. The expression pattern of TaZIP13 was notably elevated in Pak China, Zincol-2016, and Khaista following DJ13 inoculation, as well as in Pak China and Zincol-2016 after ZnSO₄ treatment. In contrast, very low expression levels of TaZIP13 were recorded in other genotypes under varying treatments (Fig. 9G). The expression of the TabZIP1 gene in the leaves of eight wheat genotypes was notably low across various treatment conditions. The highest expression level was recorded in Pak China, with a 6.9-fold increase following ZnSO₄ supplementation. Significant expression was also observed in Zincol-2016 (6.2-fold increase) and Shahkar (4-fold increase) under the same treatment. Additionally, Pak China exhibited a transcript level of 3.9 ± 0.13 after inoculation with the rhizobacterial strain DJ13. In contrast, very low levels of TabZIP1 expression were detected in the wheat genotypes Khaista and Pir Sabak-2013 following ZnSO₄ supplementation (Fig. 9H).
Discussion
Pakistan faces a significant malnutrition burden, with widespread micronutrient deficiencies, particularly zinc (Zn), which is critical for human health but often lacking due to inadequate dietary intake39. Wheat, a staple food for over 40% of the global population, has been bred primarily for yield rather than nutritional quality, resulting in low grain Zn content, especially in developing countries40. Wheat is a Zn in-efficient plant under both Zn-deficient or sufficient conditions and a rational manipulation to establish a well-coordinated source–sink relationship is required to increase grain Zn content. To address this, strategies like commercial Zn fortification and biofortification through breeding and agronomic practices have been explored to enhance wheat grain Zn content41. Zinc-solubilizing bacteria (ZSB), such as Bacillus species, in combination with chemical fertilizers, could increase Zn bioavailability by converting insoluble zinc42. The positive impact of the zinc-solubilizing bacteria and increased soil availability has been reported in wheat43 and soybean44. The seed priming with Bacillus45 is considered as an efficient way for plant growth promotion46. Moreover, the microbial potential to modulate gene expression to facilitate Zn transport and accumulation in the cereal grains has also been documented47. These bacteria, used in combination with chemical fertilizers or seed priming, offer a sustainable approach to biofortification, improving wheat flour’s nutritional quality and addressing human micronutrient deficiencies through microbial-facilitated nutrient enhancement.
In the present study three zinc solubilizing rhizpspheric bacterial strains were chosen, namely, DJ4 (Pseudomonas lurida), DJ5 (Pseudomonas aeruginosa) and DJ13 (Pseudomonas fluorescens) to determine whether bacterial inoculation can be successful in increasing zinc accumulation in diverse wheat genotypes. Members of Pseudomonas genus have been reported as plant growth promoters with zinc solubilization properties, IAA production and phytohormone production48,49. Different bacterial species from the rhizosphere belonging to Bacillus, Pseudomonas, Rhizobium, Azospirillium, Burkholderia, Gluconacetobacter and Serretia sp etc. have been reported to show zinc solubilization50. The bacterial strains used during the current study also exhibited a high Zn solubilization index for insoluble ZnO as well as the production of gluconic and glucouronic acid, subsequently in reduction of medium pH. Furthermore, the approach of detecting the presence of diverse bacterial species using a hydroponic plant-maintenance assay has been reported51 and providing an alternative to sequencing-based bacterial community studies. Scanning electron microscopic (SEM) analysis of root sections of three wheat genotypes grown in hydroponic culture revealed attachment of DJ13 (Pseudomonas fluorescens) with the root surface of 7 days old seedlings. These findings of colonization are consistent with other studies reporting wheat root colonization and biofilm formation by various species of Pseudomonas having plant growth promoting activities through SEM analysis52.
Zinc enters plants primarily via root absorption of Zn2+ from the soil solution, which is a dynamic and complex process. The interception of nutrients by roots is an important uptake mechanism for soil immobile nutrients such as Zn. Thus, root interception (i.e., root growth and root surface area) is also an important factor in determining plant availability of Zn. This differentiate the varieties of a single crop specie in zinc acquisition. Therefore, the concentration and distribution of zinc in eight wheat genotypes were measured in roots, shoot and grains after inoculation with DJ13, supplementation with ZnSO4 or without Zn application. Though genotypic variation were evident, enhanced zinc content in the different plant tissues were noted after inoculated with Zinc solubilizing rhizospheric bacteria and ZnSO4, compared to control plants. Wheat genotype Sirin acquired maximum zinc from the soil in plants grown without zinc application, whereas maximum content in the grains was noted in Zincol 2016. Furthermore, Zincol 2016 could accumulate the maximum zinc in the leaves. Similar variation in zinc uptake, translocation and concentration in the grains was noted in the genotypes after supplementation with ZnSO4. Reports have revealed that most of the nutrients derived from vegetative tissues especially 70% of Zn accumulated at the grain filling stage53. Though the genetic mechanism of grain zinc concentration in wheat has been studied54these results indicate the necessity of a more detailed study of the genetic variation of the genotypes efficient in zinc uptake, translocation as well as grain deposition for developing varieties with high grain zinc content under variable availability in the soil The Zn biofortification in wheat grains through ZSB inoculation may be a promising approach to reduce its deficiency in humans55. The results from this study also demonstrated an increased concentration in grain zinc content after inoculation with DJ13. Similar results of an increased Zn content in grain in response to ZSB inoculation was noted by other studies56. However, genotypic differences in zinc uptake, phloem loading, sequestering, and grain deposition were evident. These variations may be attributed to the variable synergism between host plant and bacterial strains to stimulate biological processes, mineralization and solubilization57. Consequently, the expression of zinc transporter genes may be changed to maintain cellular homeostasis which lead to variable uptake of zinc in crops58. Thus, appropriate host plant bacterial combination or a consortia of bacteria shall be identified by understanding the molecular mechanism of zinc uptake from the soil and its translocation to the grains so that biofortification can be achieved more efficiently.
One of the primary objectives of this study was to investigate the impact of synergistic microbial applications on the expression of genes associated with zinc uptake and translocation in plants. The ZIP (ZRT, IRT-related proteins) family transporters are known for zinc and iron transport in plants. The expression of ZIP genes plays a critical role in zinc grain accumulation in wheat by regulating zinc uptake, transport, and distribution within the plants59. ZIP transporter facilitate Zn uptake through the roots increasing influx and subsequently its transport to the shoot and grains. Studies on rice and barley has shown that overexpression of ZIP genes such as OsZIP4 or HvZIP7 significantly increased zinc uptake, with downstream effects on seed zinc content. Most of ZIP family genes are activated in zinc deficient condition and their expression in shoot and root varies60. In the present study, the induction of different zinc transporters in wheat plants inoculated with zinc solubilizing DJ13, supplemented with ZnSO4 or without Zn supplementation (control) were checked. In this study, high expression levels of TaZIP1, TaZIP4, TaZIP13, and TabZIP1 were observed in wheat genotype Zincol 2016 following ZnSO₄ supplementation, while elevated expression of TaZIP4 and TaZIP13 was noted after bacterial inoculation. The TaZIP5 gene exhibited increased expression, accompanied by elevated root zinc content, in the wheat genotype Sirin following bacterial inoculation, however, no TaZIP genes were induced in this genotype following ZnSO4 supplementation. Thus the differences in zinc uptake from the soil after bacterial inoculation or ZnSO4 treatment by Sirin may be attributed to diverse genetic mechanism. Additionally, the cultivars Atta Habib and Pak China exhibited significantly increased root zinc and high expression levels of ZIP5, ZIP7, and ZIP9 genes and ZIP3, ZIP5, ZIP13, and bZIP1 genes, respectively. An increase in the expression of TaZIPs indicated that TaZIP5, − 6, and − 13in roots are primarily involved in Zn uptake when the medium has elevated Zn levels61. The observed variations in ZIP genes expression and root zinc content among wheat genotypes, such as Zincol 2016, Sirin, Atta Habib, and Pak China, suggest that differences in the genetic mechanisms regulating zinc uptake from soil may underlie their differential responses to ZnSO₄ supplementation and bacterial inoculation. Notably, ZIP5 expression was most frequently associated with instances of higher root zinc concentrations in wheat.
After uptake, zinc moves from roots to shoots via the xylem, a process also influenced by ZIP transporters in root vascular tissues. However, elevated pool of zinc in the roots translated into variable translocation or accumulation in the grains in the studied wheat genotypes. While ZIP genes are best known for uptake, some family members may assist in loading zinc into the xylem by transporting it from root parenchyma cells into vascular bundles62. Pak China and Zincol 2016 were noted to be the most efficient wheat varieties in term of zinc transport from root to shoot after rhizospheric elevation through ZnSO4 application. TaZIP1, TaZIP13 and TabZIP1 were commonly induced in these genotypes after ZnSO4 application. Further, the shoot zinc pool was significantly elevated in Shahkar that could be attributed to higher transport from roots to shoot after inoculation with DJ13. The TaZIP3, TaZIP4 and TaZIP9 were induced and may be responsible for higher root to shoot movement of zinc in Shahkar after inoculation with DJ13. Furthermore, only TabZIP1 gene was noted to be induced in the three genotypes under both ZnSO4 and DJ13 application. In wheat, TaZIP7 or TaZIP13 are implicated in phloem loading and unloading processes. Zinc sequestering in the leaves was also variable in the genotypes. Maximum zinc pool in the leaves was noted in Sirin, Shahkar and Pir Sabak 2013 after supplementation with ZnSO4. Only TaZIP9 was induced in these genotypes after supplementation and may be responsible for increased sequestering of zinc in the leaves. This zinc can later be remobilized to the grain during senescence, a key pathway for grain zinc accumulation in wheat. Research indicates that 67–100% of grain zinc in wheat can come from remobilized sources, and efficient xylem loading supported by ZIP genes is a prerequisite. Maximum accumulation in the grains as fraction of shoot zinc was noted in Sirin after DJ13 application. During grain filling, zinc is transported to the developing seeds via the phloem. ZIP transporters expressed in phloem tissues or grain-surrounding cells. The higher grain zinc accumulation in Sirin may be associated with expression TaZIP5 after inoculation with DJ13. Thus TaZIP5 may be the key gene in both the root uptake and translocation of zinc in wheat playing a significant role in homeostasis and transport. Up-regulation of ZIP expression in phloem tissues increases zinc mobility into the grain, especially during post-anthesis when direct uptake and translocation occur63. The differential uptake and translocation kinetics may be related to genetic differences between the different varieties of wheat, consequently affecting the grain zinc accumulation64. This study has provided some preliminary information on the role of ZIP transporters in zinc uptake, translocation and grain accumulation in wheat. An extensive genomic analysis will provide a more clear understanding of the molecular basis of genetic variability for zinc uptake, root to shoot translocation and accumulation in grains of the wheat and could provide valuable insights into enhancing zinc bioavailability in wheat and other crops65.
Conclusion
In conclusion, this study elucidates the positive impact of zinc-solubilizing bacteria (ZSB) on wheat zinc uptake, translocation, sequestering, and content in the grains. Notably, a significant variation was observed in the synergistic interactions between different wheat genotypes and the bacterial strains employed, indicating a genotype-specific development of mutualistic relationships between wheat and rhizobacteria. This variability underscores the necessity for employing bacterial consortia to fully harness their biofortification potential. High expression levels of TaZIP1, TaZIP4, TaZIP13 and TabZIP1 were correlated with increased root uptake, TaZIP1and TabZIP1 were correlated with zinc translocation and TaZIP5 with grain acculmulation. In conclusion it was found that the selected bacterial strains have the potential to solubilize zinc and establish synergistic relationship with wheat, impact zinc uptake, translocation, mobilization and modulate ZIP genes expression resulting in higher partionioning of zinc in the grains. This finding provides critical insights into mechanisms that may facilitate increased zinc uptake and translocation within the grains. These results collectively suggest that integrating ZSB into agricultural practices could significantly improve zinc biofortification in wheat, addressing micronutrients deficiencies and enhancing overall crop quality. Future research should focus on optimizing bacterial consortia tailored to specific wheat genotypes to maximize these beneficial interactions.
Data availability
The datasets generated and/or analysed during the current study are not publicly available, but are available from the corresponding author on reasonable request.
References
-
Yadav, R. C. et al. Modulation in biofertilization and biofortification of wheat crop by inoculation of zinc-solubilizing rhizobacteria. Front. Plant. Sci. 13, 777771 (2022).
-
Kiani, A. K. et al. Main nutritional deficiencies. J. Prev. Med. Hyg. 17 (2 Suppl 3), E93–E101 (2022).
-
Muthayya, S. et al. The global hidden hunger indices and maps: an advocacy tool for action. PLoS ONE. 12 (6), e67860 (2013).
-
Ali, M. et al. Growth improvement of wheat (Triticum aestivum) and zinc biofortification using potent zinc solubilizing. Front. Plant. Sci. 14, 1140454 (2023).
-
Garutti, M. et al. Definition of High-Risk early Hormone-Positive HER2 – Negative breast cancer: A consensus review. Cancers 14 (8), 1898 (2022).
-
Bhatt, K. & Maheshwari, D. K. Zinc solubilizing bacteria (Bacillus megaterium) with multifarious plant growth promoting activities alleviates growth in Capsicum annuum L. 3 Biotech. 10, 36 (2020).
-
Maret, W. Molecular aspects of human cellular zinc homeostasis: redox control of zinc potentials and zinc signals. Biometals 22, 149–157 (2009).
-
Maret, W. Zinc biochemistry: from a single zinc enzyme to a key element of life. Adv. Nutr. 4, 2–91 (2013).
-
Palmer, C. & Guerinot, M. L. Facing the challenges of cu, Fe and Zn homeostasis in plants. Nat. Chem. Biol. 5, 333–340 (2009).
-
Sinclair, S. A. & Kramer, U. The zinc homeostasis network of land plants. Biochim. Biophys. Acta. 1823, 1553–1567 (2012).
-
Sangeetha, V. J., Dutta, S., Moses, J. A. & Anandharamakrishnan, C. Zinc nutrition and human health: overview and implications. eFood 3 (5), e17 (2022).
-
Smith, M. R., Golden, C. D. & Myers, S. S. Potential rise in iron deficiency due to future anthropogenic carbon dioxide emissions. Geo Health. 1, 248–257 (2016).
-
Li-na, J. et al. Grain zinc and iron concentrations of Chinese wheat landraces and cultivars and their responses to foliar micronutrient applications. J. Integr. Agric. 21 (2), 532–541 (2022).
-
Das, A., Singh, S. K., Kumar, M. & Kumar, O. Evaluation of Different Methods of Zinc Application on Growth, Yield and Biofortification of Zinc in Rice (Oryza sativa L.) JISSS. 67, (2019).
-
Sheirdil, R. A. et al. Exploring potential soil Bacteria for sustainable wheat (Triticum aestivum L.) production. Sustainability 11, 3361 (2019).
-
Longin, C. F. H. et al. Mineral and phytic acid content as well as phytase activity in flours and breads made from different wheat species. Int. J. Mol. Sci. 24, 2770 (2023).
-
Erenstein, O. et al. Global Trends in Wheat Production, Consumption and Trade. In: Reynolds, M.P., Braun, HJ. (eds) Wheat Improvement. (Springer, Cham. (2022). https://doi.org/10.1007/978-3-030-90673-3-4
-
FAO. Food outlook-Biannual report on global food markets. Food outlook. Rome.June (2023).
-
Cakmak, I. Enrichment of cereal grains with zinc: agronomic or genetic biofortification? Plant. Soil. 302, 1–17 (2008).
-
Alloway, B. J. Zinc in Soils and Crop Nutrition, 2nd edition. IZA Brussels, Belgium and IFA Paris, France (2008).
-
Basit, A., Shah, S. T., Ullah, I., Muntha, S. T. & Mohamed, H. I. Microbe-assisted phytoremediation of environmental pollutants and energy recycling in sustainable agriculture. Arch. Microbiol. 203, 5859–5885 (2021).
-
Brown, K. H., Hambridge, K. M. & Ranum, P. Zinc fortification of cereal flours: current recommendations and research needs. FoodNutr. Bull. 31, S62–S74 (2010).
-
Cakmak, I. et al. Biofortification and localization of zinc in wheat grain. J. Agric. Food Chem. 58, 9092–9102 (2010).
-
Ku, Y. S., Rehman, H. M. & Lam, H. M. Possible roles of rhizospheric and endophytic microbes to provide a safe and affordable means of crop biofortification. Agron. J. 9 (11), 764 (2019).
-
Kumssa, D. B. et al. Dietary calcium and zinc deficiency risks are decreasing but remain prevalent. Sci. Rep. 5, 10974 (2015).
-
Ramzan, Y. et al. Ation with zinc and Iron improves the grain quality and yield of wheat crop. Int. J. Plant. Prod. 14, 1–510 (2020).
-
Serteyn, L. et al. Induced systemic resistance by a plant Growth-Promoting rhizobacterium impacts development and feeding behavior of aphids. Insects 11(4), 234 (2020).
-
Athukorala, A. D. Solubilization of micronutrients using Indigenous microorganisms. In Microbial Technology for Sustainable Environment (eds Bhatt, P. et al.) 365–417 (Springer, 2021).
-
Selvaraj, K. & Dananjeyan, B. Expression of Zn transporter genes in rice as influenced by Zn-solubilizing Enterobacter cloacae strain ZSB14. Front. Plant. Sci. 7, 446 (2016).
-
Singh, S. et al. Restructuring the cellular responses: connecting microbial intervention with ecological fitness and adaptiveness to the maize (Zea mays L.) grown in saline sodic soil. Front. Microbiol. 11, 568325 (2021).
-
Rahman, A. et al. Isolation and screening of Zn (Zn) solubilizing rhizosphere bacteria from different vegetations for their ability to improve growth, Zn uptake, and expression of Zn transporter genes in tomato. Curr. Microbiol. 81, 83 (2024).
-
Ansari, F. A., Ahmad, I. & Pichtel, J. Synergistic effects of biofilm-producing PGPR strains on wheat plant colonization, growth and soil resilience under drought stress. Saudi J. Biol. Sci. 30, 103664 (2023).
-
Khalid, S., Ahmed, I. & Amanullah & Enhancing zinc biofortification of wheat through integration of zinc, compost, and zinc-solubilizing bacteria. Agriculture 12, 968 (2022).
-
Khan, S. I. et al. Cloning,expression and biochemical characterization of lignin-degrading DyP-type peroxidase from Bacillus sp. Strain BL5. Enzyme Micro Technol. 151, 109917 (2021).
-
Jorhem, L. Determination of metals in foods by atomic absorption spectrometry after dry ashing: NMKL collaborative study. J. AOAC Int. 83, 1204–1211 (2000).
-
Okafor, S. N., Okafor, C. J., Ogbonna, E. T., Ezenwafor, C. & Osadebe, P. O. Quantitative determination of selected heavy metals and micronutrients in branded dairy products sold in Nigeria using atomic absorption spectroscopy (AAS). Afr. J. Pharm. Res. Dev. 16 (1), 33–38 (2024).
-
Wei, C. et al. Hormetic effects of zinc on growth and antioxidant defense system of wheat plants. Sci Total Environ. In Press 150992 (2021).
-
Portillo, M., Fenoll, C. & Escobar, C. Evaluation of different RNA extraction methods for small quantities of plant tissue: combined effects of reagent type and homogenization procedure on RNA quality-integrity and yield. Physiol. Plant. 128, 1–7 (2006).
-
Sharma, R. & Pal, V. Zinc Bio-Fortification in Food Crops to Alleviate zinc Malnutrition. In: Thakur, M. (eds) Sustainable Food Systems (Volume I). World Sustainability Series. Springer, Cham. (2024). https://doi.org/10.1007/978-3-031-47122-3_20
-
Singh, S., Kaur, J., Ram, H., Singh, J. & Kaur, S. Agronomic bio-fortification of wheat (Triticum aestivum L.) to alleviate zinc deficiency in human beings. Reviews Environmental Sci. Bio/Technology. 22 (1), 1–22 (2023).
-
Velu, G., Ortiz-Monasterio, I., Cakmak, I. & Hao, Y. Singh, R. P. Biofortification strategies to increase grain zinc and iron concentrations in wheat. J. Cereal Sci. 10, 1016 (2013).
-
Garcia, J. & Kao-Kniffin, J. Microbial group dynamics in plant rhizospheres and their implications on nutrient cycling. Front. Microbiol. 11 (9), 1516 (2018).
-
Kushwaha, P. et al. Enhancement in plant growth and zinc biofortification of Chickpea (Cicer arietinum L.) by Bacillus altitudinis. JPNSS 21, 922–935 (2021).
-
Sharma, S. K., Sharma, M. P., Ramesh, A. & Joshi, O. P. Characterization of zinc solubilizing Bacillus isolates and their potential to influence zinc assimilation in soybean seeds. JMB 22, 352–359 (2012).
-
Akinrinlola, R., Gary, Y., Rhae, D. & Tony, A. Evaluation of Bacillus strains for plant growth promotion and predictability of efficacy by in vitro physiological traits. Int J. Microbiol. 2018, 5686874 (2018).
-
Choudhary, S. et al. Molecular characterization and validation of zinc solubilization potential of bacteria isolated from onion (Allium Cepa L.) rhizosphere. Microbe 4, 100145. https://doi.org/10.1016/j.microb.2024.100145 (2024).
-
Krithika, S. & Balachandar, D. Expression of zinc transporter genes in rice as influenced by zinc-Solubilizing Enterobacter cloacae strain ZSB14. Front. Plant. Sci. 7, 446 (2016).
-
Pallavi et al. Isolation and characterization of halotolerant plant growth promoting rhizobacteria from Mangrove region of sundarbans, India for enhanced crop productivity. Front. Plant. Sci. 14, 1122347 (2023).
-
Singh, S., Chhabra, R., Sharma, A. & Bisht, A. Harnessing the power of Zinc-Solubilizing bacteria: A catalyst for a sustainable agrosystem. Bacteria 3 (1), 15–29 (2024).
-
Kamran, S. et al. Contribution of zinc solubilizing bacteria in growth promotion and zinc content of wheat. Front. Microbiol. 8, 2593 (2017).
-
Harris, S. L., Pelaez, C. A. & Shank, E. A. Monitoring bacterial colonization and maintenance on Arabidopsis thaliana roots in a floating hydroponic system. J. Vis. Exp. 147, e59517 (2019).
-
Tank, N. & Saraf, M. Enhancement of plant growth and decontamination of nickel-spiked soil using PGPR. J. Basic. Microbiol. 49, 195–204 (2009).
-
Wani, S. H. et al. Improving zinc and iron biofortification in wheat through genomics approaches. Mol. Biol. Rep. 49, 8007–8023 (2022).
-
Velu, G. et al. Genetic dissection of grain zinc concentration in spring wheat for mainstreaming biofortification in CIMMYT wheat breeding. Sci. Rep. 8, 13526 (2018).
-
Ramesh, A., Sharma, K. S., Sharma, M. P., Yadav, N. & Joshi, O. P. Inoculation of zinc solubilizing Bacillus aryabhattai strains for improved growth, mobilization and biofortification of zinc in soybean and wheat cultivated in vertisols of central India. Appl. Soil. Eco. 73, 87–96 (2014).
-
Abaid-Ullah, M. et al. Plant growth promoting rhizobacteria: an alternate way to improve yield and quality of wheat (Triticum aestivum). Int. J. Agric. Biol. 17, 51–60 (2015).
-
Yadav, R. C. et al. Zinc-solubilizing Bacillus spp. In conjunction with chemical fertilizers enhance growth, yield, nutrient content, and zinc biofortification In wheat crop. Front. Microbiol. 4, 1210938 (2023).
-
Van de Mortel, J. E. et al. Large expression differences in genes for iron and zinc homeostasis, stress response, and lignin biosynthesis distinguish roots of Arabidopsis thaliana and the related metal hyperaccumulator Thlaspicaerulescens. Plant Physiol. 142 (3), 1127–1147 (2006).
-
Tiong, J. et al. Increased expression of six ZIP family genes by zinc (Zn) deficiency is associated with enhanced uptake and root-to-shoot translocation of Zn in barley (Hordeum vulgare). New. Phytol. 207, 1097–1109 (2015).
-
Chen, W. R., Feng, Y. & Chao, Y. E. Genomic analysis and expression pattern of OsZIP1, OsZIP3, and OsZIP4 in two rice (Oryza sativa L.) genotypes with different zinc efficiency. Russ. J. Plant Physiol. 55, 400–409 (2008).
-
Li, G., Li, C., Rengel, Z., Liu, H. & Zhao, P. Excess Zn-induced changes in physiological parameters and expression levels of TaZIPs in two wheat genotypes. Environ. Exp. Bot. 177, 104133 (2020).
-
Lee, S. et al. Redundant roles of four ZIP family members in zinc homeostasis and seed development in Arabidopsis thaliana. Plant J. 108 (4), 1162–1173 (2021).
-
Stanton, C., Sanders, D., Kramer, U. & Podar, D. Zinc in plants: integrating homeostasis and biofortification. Mol. Plant. 15 (1), 65–85 (2022).
-
Hacisalihoglu, G., Hart, J. J. & Kochian, L. V. High- and low-affinity zinc transport systems and their possible role in zinc efficiency in bread wheat. Plant Physiol. 125 (1), 456–463 (2001).
-
Velu, G., Ortiz-Monasterio, I., Cakmak, I. & Hao, Y. Singh, R. P. Biofortification strategies to increase grain zinc and iron concentrations in wheat. J. Cereal Sci. 59 (3), 365–372 (2014).
Acknowledgements
Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2025R39), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
We all declare that manuscripts reporting studies do not involve any human participants, human data, or human tissue. So, it is not applicable.
Complies with international, national and/or institutional guidelines
Experimental research and field studies on plants (either cultivated or wild), comply with relevant institutional, national, and international guidelines and legislation. Experimental studies were carried out in accordance with relevant institutional, national or international guidelines or regulation.
Permissions or licenses
The experiment was started, after taking permission from The University of Agriculture, Peshawar, Khyber Pakhtunkhwa, Pakistan.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Malik, S., Iqbal, A., Munir, I. et al. Microbial assisted zinc biofortification of wheat germplasm for the amelioration of zinc malnutrition. Sci Rep 15, 24555 (2025). https://doi.org/10.1038/s41598-025-09946-4
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-09946-4