- Research
- Open access
- Published:
Biotechnology for Biofuels and Bioproducts volume 18, Article number: 78 (2025) Cite this article
Abstract
Background
The increasing global demand for fuel, driven by the unchecked extraction and consumption of fossil fuels, has intensified the search for sustainable energy alternatives. Recent advancements in biodiesel production techniques highlight the potential of microbial processes. Lipase-mediated whole-cell biocatalysts for biodiesel production offer a sustainable and economical route that eliminates the need for enzyme purification. These biocatalysts use microbial cells that express lipase to catalyze the transesterification of oils into biodiesel. Their good efficiency, reuse, and operational simplicity make them a new promising alternative to green energy solutions.
Result
This work employs the marine bacterial strain Bacillus licheniformis to develop a whole-cell biocatalyst for the enzymatic transesterification process of Madhuca indica oil in order to produce biodiesel. Optimal conditions for achieving a biodiesel yield of 95.3% were identified as a methanol-to-oil molar ratio of 7.5:1 and a catalyst concentration of 30 wt%. The performance and emission characteristics of biodiesel blends MB30 and MB50 were evaluated in comparison to conventional diesel. Results indicated that MB30 and MB50 blends reduced CO emissions by 11.71% and 27.93%, respectively, compared to diesel. Additionally, MB30 showed decreases in hydrocarbon emission (HC) and smoke opacity by 23.53% and 3.02%, respectively, while MB50 exhibited reductions of 36.47% and 15.42%, respectively. The nitrous oxide emission is enhanced while using biodiesel blends MB30 and MB50 by 13.34% and 15.96% respectively.
Conclusion
The analysis indicates the lipolytic activity of this bacterial strain Bacillus licheniformis, is efficient in converting Madhuca indica oil into biodiesel by a sustainable process. The produced biodiesel had better fuel properties and reduced emissions during engine analysis with respect to CO and particulate matter. This further strengthens its potential to be considered a green alternative to conventional fossil fuels. The process will make use of naturally occurring catalytic properties of bacteria and, hence, would be comparatively green and cheap. This brings to note the possibilities that bio-based resources have opened up for cleaner and more sustainable energy production.
Highlights
-
This is the first research to use marine bacteria as a whole-cell biocatalyst for the production of Madhuca indica biodiesel.
-
The bacterial strain was isolated from a marine sponge Tedania anhelans.
-
Parameters for the synthesis of biodiesel were optimized using the RSM approach.
-
The maximum yield of biodiesel produced was 95.3%.
-
In engine study, the biodiesel blends MB30 and MB50 demonstrate a decrease in CO, HC, and smoke emissions.
Graphical Abstract

Introduction
According to the 2022 global energy forecast, fossil fuels account for approximately 80% of global energy demand [1]. Coal, oil, and gas accounted for around 84% of worldwide primary energy in 2019 [2]. Over the next several decades, we must rapidly reduce this fraction by replacing it with low-carbon energy sources. Around 64% of the electricity is produced by using fossil fuels [3]. Petroleum diesel, a popular fossil fuel, has been critical in powering transportation, industries, and numerous sectors of the global economy. However, its widespread use has had serious environmental effects, raising worries about its long-term viability and impact on the world [4]. Due to their extensive usage of fossil fuels, the developed world’s high demand for energy has resulted in issues with environmental contamination in both the residential and commercial sectors. Burning fossil fuels poses several health dangers to the general public as well as environmental issues that could have long-lasting effects on global warming. As a consequence, there are now more worries about how things will affect the environment, which has led to a closer look at other sources of energy [5]. Wind and solar, hydropower, bioenergy, and biofuels are examples of typical renewable energy sources. Due to economic and environmental factors, each of these contributions is significant, and biodiesel may be one of the alternatives [6].
First-generation biodiesel was mostly derived from edible plant and animal sources. Second-generation biodiesel has been produced utilizing non-edible feedstock such as discarded vegetable oils, non-edible vegetable oils, and animal fats as worry about food security has increased. Several plant-based second-generation biodiesel feedstocks have been reported recently, including castor oil, Jatropha, Calophyllum inophyllum, Karanja, cottonseed, neem, and used sunflower oil. One such biomass, formally called “Madhuca indica“, is the subject of the current study and is widely farmed in the center of India. Approximately 300 distinct kinds of non-edible seeds are being grown in India, which is further facilitated by the country’s varied meteorological conditions [7]. A significant non-edible plant is M. indica. In the southern and central regions of India, this plant is extensively distributed. With 20–200 kg of seed produced per tree each year; it is predicted that 0.5 million tons of M. indica seed will be grown [8]. Oil content in M. indica seeds ranges from 45 to 60% [9, 10]. They are useful for the generation of biodiesel because of their high oil content. The oil from the seeds is dissolved using a chemical solvent during solvent extraction [11]. Similar characteristics exist between M. indica biodiesel and other biodiesel fuels.
Methanolysis, also known as transesterification, is a method that can be employed with or without a catalyst to produce biodiesel from vegetable oil. The mechanism of replacing an ester’s organic group R′′ with an alcohol’s organic group R′ is referred to as an equilibrium reaction [12]. There are two types of catalyst-assisted transesterification processes: homogenous and heterogeneous. A catalytic-free procedure known as Supercritical methanolysis is used in the non-catalytic transesterification method [13]. Base-catalyzed transesterification is a common process for making biodiesel from triglyceride-rich feedstocks including vegetable oils and animal fats. A strong base catalyst, often sodium or potassium hydroxide, is employed in this method to facilitate the reaction between the triglycerides and an alcohol (commonly methanol or ethanol). Unlike the more common base-catalyzed transesterification, acid-catalyzed transesterification uses an acid catalyst, such as sulfuric acid, to facilitate the conversion of triglycerides into fatty acid alkyl esters (biodiesel) and glycerol [14]. Lipases are used in the transesterification process, as a biocatalyst, to change the triglycerides found in feedstock oils into fatty acid methyl esters (FAMEs) with alcohol. An alternative to chemical methods, the enzymatic approach is environmentally benign, works in moderate circumstances, and avoids saponification. To produce biodiesel as efficiently and sustainably as possible, lipases from microorganisms and other sources are used as biocatalysts.
Using whole microbial cells as catalysts for different chemical reactions is known as whole-cell biocatalysts. These cells can be naturally occurring microorganisms or strains created through genetic engineering that have the capability to produce lipase enzymes. In comparison to isolated enzymes or conventional chemical catalysts, whole-cell biocatalysts have several advantages, making them a desirable method in a variety of commercial applications [15]. This gives raw material selection freedom and enables the conversion of various feedstocks into desired outputs. Whole cells often display superior stability and robustness under varied reaction circumstances compared to isolated enzymes. Whole-cell biocatalysis can be easily included in already-existing fermentation processes or combined with after-facility recovery and purification operations. Because of this integration, production procedures may be made more efficient, with fewer stages overall and lower costs. In contrast to conventional chemical processes, it makes use of renewable starting materials, operates in comfortable settings, and produces little waste [16].
There are few research works reported in the literature for whole-cell catalyzed biodiesel production. Nain et al. investigated the conversion of cottonseed oil to biodiesel using Aspergillus sp. as a whole-cell catalyst. The transesterification reaction was at 30 °C with agitation of 150 rpm for 120 h. At two distinct alcohol ratios (acyl-acceptor), 1:4 and 1:6 M, biodiesel yield was calculated to be 91% and 86%, respectively [17]. Yang et al. used a whole-cell biocatalyst to develop a novel method for regioselective acylation of helicid with fatty acid vinyl esters. Among the microbial stains utilized, pseudomonas aeruginosa GDM exhibited the highest conversion [18]. Pseudomonas mendocina immobilized in Fe3O4-chitosan microspheres is used as a whole-cell biocatalyst in the production of biodiesel from soybean oil by Chen et al., It was discovered that the greatest biodiesel yield of 87.32% was obtained under the following conditions: 10% biocatalyst concentration, 10% water content, 35 °C, and a methanol-to-oil molar ratio of 4:1 [19]. Using the pPICZB vector from E. coli and the Cal A and Cal B genes from Candida antarctica JCM 10318, Iyyappan et al., created a recombinant Pichia pastoris that was employed as a whole-cell biocatalyst for the conversion of Caulerpa racemosa algal oil to biodiesel. They optimized the reaction using the Plackett–Burman design (PBD) with the variable parameters methanol-to-oil ratio, combined whole cell biocatalyst concentration, water content, temperature, reaction time, agitation speed, and solvent to oil ratio. After 30 h of reaction with a methanol-to-oil ratio of 4.1, a total whole-cell biocatalyst concentration of 38.88% (w/v), and a temperature of 38 °C, the highest biodiesel production of 93.64% was attained [20]. Sena et al., generated recombinant Pichia pastoris by incorporating the lipase-generating gene of Rhizomucor miehei into the PIR-1 vector and using the same as a whole-cell biocatalyst. They evaluated this biocatalyst on two distinct substrates, soybean fatty acid distillate (SFAD) and palm fatty acid distillate (PFAD) for biodiesel production and discovered that SFAD has the greatest biodiesel conversion of 79.1% with a catalyst concentration of 10% and a reaction duration of 48 h [21]. Liu et al. investigated biodiesel generation from waste frying oil in a magnetically fluidized bed reactor packed with beads made up of ferroferric oxide-polyvinyl alcohol that were immobilized with Bacillus subtilis cells. The ideal conditions were predicted using response surface methods and the Box-Behnken design, and the greatest yield of biodiesel was 89% at 48 h under the optimized conditions [22].
The immobilization of bacteria may be accomplished with success using polyurethane matrices [23]. To keep bacteria in a stable position while still allowing them to carry out their biological tasks, they are often enclosed within a solid matrix or support material. This procedure is known as bacterial immobilization [24]. In general, polyurethane is biocompatible and does not significantly harm or negatively affect bacterial cells. This characteristic makes it appropriate for promoting bacterial activity and proliferation. It may be designed to have a variety of permeabilities and porosities [25]. This enables the effective interchange of substances between the environment and the immobilized microorganisms, including nutrients, gasses, and metabolites. The mechanical characteristics of polyurethane can be adjusted to provide the immobilization matrix with structural stability. This stability facilitates the development of bacteria over an extended length of time and guarantees the lifespan of the matrix [26]. Polyurethane matrices provide for the flexible design of immobilization systems for diverse purposes since they may be molded into a variety of forms and sizes. Bacterial cells may adhere to polyurethane well, which makes it easier for them to stick to the matrix surface. This adherence encourages bacterial colonization and improves the effectiveness of immobilization in general [27]. Through several techniques, including direct inoculation, impregnation, or trapping during the matrix creation process, bacterial cells can be introduced into the matrix [28].
This study obtained a bacterium strain from a marine sponge that exhibited greater lipolytic activity and evaluated its lipolytic activity. The selected bacterial strain was rendered immobile within the matrix of reticulated polyurethane. Using response surface methods, the immobilized strain of M. indica was used to produce biodiesel by optimizing process parameters including catalyst concentration and methanol-to-oil molar ratio. Furthermore, the MB30 and MB50 biodiesel blends’ performance and emission analysis of the produced biodiesel were evaluated.
Materials and methods
Materials
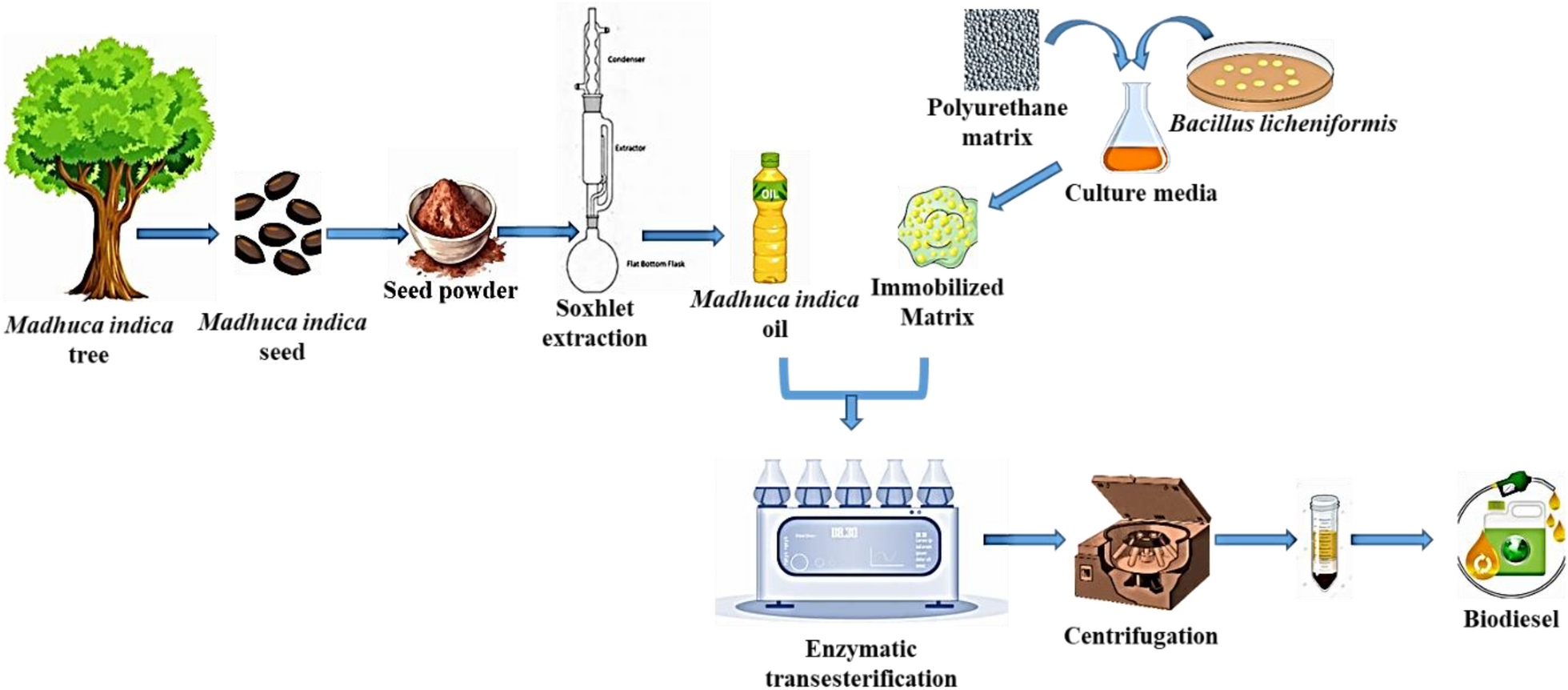
M. indica seeds were obtained from neighborhood farmers in Thanjavur. The seeds were dried, ground into a powder, and then subjected to the oil extraction process (Fig. 1). The Soxhlet extraction method was employed to extract M. indica oil using hexane (Sisco Research Laboratory Pvt. Ltd) as the solvent, resulting in an oil yield of 38.6%. The fatty acid profile of the M. indica oil is shown in Table 1. Polyurethane foams were obtained from the local market and cut in the dimensions of 0.5 cm × 0.5 cm × 0.5 cm. From Himedia, the tributyrin agar base medium was procured. Carbinol, 99% (Sisco Research Laboratory Pvt. Ltd) was procured for transesterification process.

a M. indica seed; b M. indica oil cake; c M. indica oil and Biodiesel
Marine bacterial isolation and lipolytic activity screening
The bacterial strains used in the present study were isolated from the marine sponge Tedania anhelans [29]. Four different strains (S1–S4) were sub-cultured on Zobell Marine Agar (ZMA) and were screened to identify lipase-producing strains using tributyrin agar as the selective medium. These strains were streaked in tributyrin agar plate and incubated at 30 °C for 24 h. Isolates having clearing zone were grown in the liquid culture and the level of lipase production was determined from the cell free culture supernatant fluid. Full-length sequencing of the 16S rRNA gene revealed that all four sponge-associated bacteria had the closest match to Bacillus sp. Figure 2 represents the process flowsheet of biodiesel production using the whole-cell biocatalyst.

Process flow sheet of biodiesel production from M. indica seed using whole-cell biocatalyst
Immobilization of bacterial cells in polyurethane matrix
The polyurethane matrices were reticulated using 1N NaOH solution. The matrices were then repeatedly cleaned with distilled water and allowed to air dry for 24 h. During submerged culture in Zobell marine broth, the bacterial strain’s immobilization within polyurethane foam occurred on its own [30]. A 500 mL Erlenmeyer flask containing 250 mL of Zobell marine broth and 100 biomass supporting particles (BSPs) was used for the bacterial strain’s cultivation, which was done on a rotary shaker at 220 rpm and 28 ℃ for 72 h. Then the immobilized polyurethane matrices were filtered and air dried for 24 h. Approximately, 0.15 g of bacterial cells were immobilized in the single matrix. The matrices were subjected to a scanning electron microscope (SEM) to determine the presence of cells [31, 32]. The enzymatic activity of the bacterial cell was determined using the olive oil emulsion assay [33,34,35]. The standard lipase (1 mg/mL) was acted as a positive control for the assay and the enzymatic activity was calculated using the Eq. 1.
$$text{Enzyme activity }= frac{text{KOH }(text{mL})times text{molarity of KOH }}{text{Assay mixture}}*1000$$
(1)
Enzymatic transesterification
The antimicrobial activity of M. indica oil was tested using agar well diffusion method. The enzymatic transesterification of M. indica oil is catalyzed by using immobilized bacterial cells [36,37,38,39]. The reaction was performed in a 50 mL Erlenmeyer flask with the molar ratio of methanol: oil of 3:1 and the catalyst concentration of 15% w/w. The water content was fixed to 15-wt%. Methanol was split into three parts and added at regular intervals of 0 h, 24 h, and 48 h. Then the resulting biodiesel was recovered by centrifuging the mixture at 6000 rpm for 20 min and heating the supernatant (to remove the unreacted methanol). The biodiesel is then subjected to FTIR and GC–MS analysis [40].
Optimization of biodiesel production using central composite design (CCD)
The traditional experimental approach, which includes factor optimization one at a time, takes a long period. Additionally, traditional optimization cannot discover the interacting effects of all independent factors over the dependent variable. To adjust the operational parameters for improved biodiesel production, interaction effects are studied using the response surface methodology. The most popular technique for examining how process factors interact is central composite design (CCD).
The numerical components are modified in statistical design at three levels (− 1, 0, 1) (Table 2). The total number of options is 2k, + 2k + n0, according to the design. The number of experiment repetitions at the center point is n0, where K is the number of independent components. The methanol-to-oil ratio and the quantity of BSPs were tested at two levels, with zero serving as the midpoint. The set central values for the whole-cell technique of producing biodiesel are 7.5:1 and 15 BSPs. To prevent bias, the experimental design was carried out utilizing coded values. The Minitab® 16.1.1 software version, which generates 13 tests every version, produced a matrix, according to CCD:
$$text{Biodiesel} text{yield} (Y) = {C}_{0}sum_{I=1}^{K}{C}_{i}{X}_{i}+sum_{I=1}^{K}{C}_{ii}{X}_{i}^{2}+sum_{i,j>j}^{k} sum_{i=1}^{k}{C}_{ij}{X}_{i}{X}_{j}+e$$
(2)
where C0 is the regression coefficient of the model and Xi and Xj represent the independent variables in coded values. Using this equation, it is possible to evaluate the linear, quadratic, and interactive effects of the independent variables on the response.
Characterization of M. indica biodiesel
Fourier-Transform Infrared Spectroscopy provides important information about a substance’s chemical composition, molecular structure, and functional groups. The interaction of infrared (IR) light with matter, specifically the vibrations of chemical bonds inside molecules, provides the basis for FTIR spectroscopy. When infrared light gets through a sample, some is absorbed and the rest is analyzed. The resulting spectrum is a depiction of light absorption strength as a function of wavenumber.
NMR analysis is a potent analytical method used to investigate the dynamics, content, and structure of molecules. It is based on nuclear magnetic resonance theories, which describe how atomic nuclei interact with magnetic fields. Nuclear magnetic resonance (NMR) analysis, commonly referred to as proton NMR or 1H-NMR, is a widely used method that focuses on the identification and examination of hydrogen nuclei (protons) in a molecule. It offers important details regarding the connectivity, molecular organization, and chemical setting of hydrogen atoms inside a molecule. Nuclear magnetic resonance (NMR) spectroscopy employs a method known as carbon-13 NMR, also known as 13C-NMR, to analyze the carbon-13 nuclei (carbon isotopes) in molecules. It offers important details regarding the connectivity and chemical environment of the carbon atoms found in a molecule. TopSpin software was used to align the spectra and make baseline adjustments. The data were normalized and the spectral signals were integrated using a Matlab procedure. The JNM-ECZ600R/S1 spectrometer was used to record the 1 H NMR spectra at 600 MHz with a field strength of 14.09636928 Tesla.
Over a sweep width of 12.01923077 kHz, 32,768 data points were gathered throughout the 2.7262976 s capture period. A 5 s relaxation delay and a 16 s receiver gain were used. A constant temperature of 22.1 °C was maintained for the sample. Protons had a 90-degree pulse width of 7.62 ms, a 45-degree pulse angle, and an acquisition duration of 2.7262976 s. A Dante loop of 500 was employed, and the triple resonance and irradiation modes were turned off. A 0.69206016-s collection time, 32,768 data points, and a 14.09636928 Tesla field strength were used to produce the 13 C NMR spectra at 150.91343039 MHz. 1024 scans with a 2-s relaxation delay were performed while the sample temperature was maintained at 22.3 °C. There found a 28.977 dB decoupling attenuation and a 5 mg/L offset at the proton decoupling frequency of 600.1723046 MHz.
Gas Chromatography–Mass Spectrometry (GC–MS) is an effective method of analysis that has proven beneficial in characterizing and evaluating the purity of biodiesel. In GC, a vaporized biodiesel sample is put into a heated column, where components separate depending on their chemical composition. Because each component elutes at a different retention time, both qualitative and quantitative examinations are possible. Individual chemicals eluting from the GC column are identified and quantified using MS. It determines molecular structure by measuring the mass-to-charge ratio of ions formed when molecules are ionized. It produces extensive information in the form of mass spectra. These spectra are evaluated to detect and analyze components in the biodiesel sample. The starting temperature of the column oven used for the GC–MS analysis of biodiesel is 50.0 °C, with ramping as necessary to separate the FAME components. To prevent column overloading while introducing samples in split mode, the injection port is adjusted at 200.00 °C. 3 min after injection, the analysis starts, and it ends at 59.33 min. In order to capture the entire spectrum of biodiesel components, the mass spectrometer works in scan mode, collecting data every 0.40 s at a scan speed of 1666 and scanning a mass range of 40.00–600.00 m/z.
Engine experiment setup
To evaluate the quality of the biodiesel produced, a single-cylinder Kirloskar TV1 type CI engine was used. This four-stroke engine was cooled by a water-cooling system and operated at 1500 rpm. Using commercial diesel and Mahua biodiesel, the engine’s performance was evaluated under five distinct load conditions using an eddy current dynamometer (Fig. 3). The engine’s rated output is 3.75 kW, and its compression ratio is set to 17:1. The engine’s exhaust gas temperature was monitored by installing a K-type thermocouple in the tailpipe. By using AVL DI gas 444 N gas analyzer, the emissions of hydrocarbons (HC), carbon monoxide (CO), and nitric oxide (NO) and by using AVL 437C smoke meter, the emission rate of smoke were assessed.

Schematic representation of engine test
Results and discussion
Screening and identification of marine lipolytic bacteria
The microorganisms which produce lipase are capable of breaking down triglycerides into smaller pieces. Glycerol and three fatty acids make up lipids. These are disintegrated and may be transformed into a range of final products that the organism can utilize for energy production and some other operations. This led to the formation of zones of clearance around the bacterial colonies. On observing the strains inoculated in tributyrin agar, it has been found that the bacterial strains in S1 exhibits greater lipolytic activity, when compared with the other three strains. The 16S rRNA gene sequencing clearly shows that the strain S1 belongs to Bacillus family and matches maximum similarity with the 16S rRNA sequence of Bacillus licheniformis (Genbank accession # JX489608) by 16S rRNA gene sequence analysis. B. licheniformis has been well known for its high lipolytic activity lipases, which is a good candidate in biodiesel production through enzymatic transesterification. The B. licheniformis lipase has demonstrated stability under broad pH and temperature ranges, which are important factors in effective biodiesel synthesis [41]. Additionally, the application of lipases from probiotic origin such as B. licheniformis provides environmental advantages by not producing toxic substances related to certain fungal-derived enzymes [42].
Lipolytic activity of the bacterial cells
The whole-cell biocatalyst was found to have a specific enzyme activity of 31.7 U/mg. The whole-cell biocatalyst exhibited enzyme activity that functions as a catalyst for the generation of biodiesel, as evidenced by its specific enzyme activity, which was almost half that of pure lipase (67.6 U/mg). Even with the lower activity than for pure lipase, the whole-cell biocatalyst is cost-effective and easy to produce [43]. The low activity may be caused by effects such as immobilization of the enzyme within the cell or restriction in substrate accessibility.
SEM analysis of the immobilized cells
Under a scanning electron microscope, bacterial cells that had become immobilized within the matrix were examined. The SEM analysis of the matrix is shown in Fig. 4a and b following 48 and 72 h of immobilization, respectively. The rod-shaped bacterial cells were entrapped inside the matrices can be visualized in the SEM images. It has been noted that after 72 h of immobilization, the bacterial strain immobilization is significantly greater than after 48 h [30].

The SEM image of polyurethane matrix coated with bacterial cells: a after 48 h; b after 72 h
Central composite design (CCD)
The interaction effects of the process-independent variable with the dependent variable are explained by the response surface approach using CCD. The process-independent and dependent variables were computed and analyzed by CCD for the optimization of biodiesel production. The determined and included in the data under analysis was the measured biodiesel yield (Table 3):
$$text{Biodiesel yield}= 16.28times {C}_{M}+2.89 times {C}_{i} -1.14times {C}_{M}^{2}-0.06times {C}_{i}^{2}+0.09times {C}_{M}times {C}_{i}-16.14.$$
(3)
The square correlation and adjusted R2 value play a major role in the accuracy and fitness of the model. R2 and R2adj were determined to be 99.53% and 99.20% for the whole-cell biocatalyst method of producing biodiesel. R2’s value suggests that a model’s fitness is necessary. The P value for every coefficient was found to be less than 0.05, which states that the process is significant. The outcome was a greater F value for the independent variable of catalyst weight, which is the one that has the greatest beneficial impact on FAME yield compared with that of other parameters (Table 4).
The Normal Probability Plot of Residuals shows a good fit of data points along the straight line, which implies that the residuals are normally distributed (Fig. 5). This is an indication that the statistical model is able to capture the underlying patterns in the data. Consequently, this fit gives confidence in the accuracy and reliability of the model’s predictions for biodiesel production. For the two components’ contributions to the synthesis of biodiesel, contour (2D) and response surface plots (3D) were developed using the response data (Fig. 6). The cumulative effects of the process variable across the response element are displayed in contour plots. These graphs’ linear and interactive impacts demonstrate the ideal conditions for biodiesel generation. While retaining other independent variables at the fixed center point, the graph was produced between any two independent variables and the yield on the third axis. The graph’s darkest areas correlate to the highest yield. To fit the response and identify important factors, a regression analysis of the data was created [44].

Observed vs. predicted values of biodiesel production

Response surface graph representing interaction effects between methanol /oil molar ratio and catalyst concentration (Ci %W/W)
By performing experiments for various trials with different working parameters, the process parameter, Methanol: oil ratio of 7.5: 1 and 30% w/w of cell concentration results in maximum conversion of fatty acid methyl esters.
Characterization of Madhuca indica biodiesel
FTIR result of biodiesel
The IR spectra of the M. indica oil and biodiesel samples were identical since the process primarily consists of eliminating glycerol and replacing the methyl group in the hydrocarbon chain. Figure 7 represents the FTIR result of produced biodiesel. There is a substantial variation in the range of 900–1400 cm−1. In the oil sample, the band at 1166 cm−1 represents the expansion vibration of the (C–O) group coupled to (CH2–). In the biodiesel sample, it has been relocated to 1160 cm−1. The presence of ester groups in the sample has been confirmed by the bands at 1742 cm−1 and 1744 cm−1 in both biodiesel and oil samples, respectively [45]. The band at 1468 cm−1 in the oil sample represents the CH3 asymmetric bend that is observed in it because of the presence of the triglyceride group, and the band at 1464 cm−1 is because of the presence of the methane group. In addition, some additional peaks at 1381 cm−1 in biodiesel have been discovered, indicating the presence of a methyl group, which was not observed in the oil sample. This indicates the successful conversion of M. indica oil to biodiesel [46, 47].

FTIR result of oil and biodiesel
NMR analysis
1H NMR spectroscopy was used to characterize the M. indica biodiesel, and the spectrum is shown in Fig. 8. The strong singlet at 1.2 ppm indicated the presence of methylenes in the carbon chain. The production of methyl ester (–CO2CH3) is indicated by the prominent singlet at 3.65 ppm. The presence of the signals at 4.1–4.5 ppm could potentially be attributed to the presence of protons connected to the glycerol triacylglycerols. Protons located in CHl groups adjacent to the methyl or glyceryl ester moieties (–CH2CO2CH3 for methyl esters) provide the signals at 2.31 ppm. Similar chemical shifts occurred at 5.2, 5.3, and 5.4 ppm in the form of triplets, which were olefinic proton properties [48].

1H NMR spectra of the produced biodiesel from M. indica oil
The ester carbonyl (COO) and CAO characteristic peaks are seen in the 13C-NMR spectra of M. indica biodiesel using deuterated chloroform (CDCl3) as a solvent (Fig. 9). These peaks are located at 173.12 and 51.22 ppm, respectively. The peaks at 129.82 and 127.75 ppm demonstrated the unsaturation in methyl esters. Signals at 22–34 ppm are created by methylene carbons in fatty acid methyl esters, while further peaks at 13.95 ppm are caused by the terminal carbons of methyl groups [48].

C13 NMR spectra of the produced biodiesel from M. indica oil
GC–MS analysis
The GC–MS result of the produced M. indica biodiesel shows the presence of diverse group of FAME content (Fig. 10) (Table 5). Hexadecanoic acid, methyl ester (27.69%), 9-Octadecenoic acid (Z)-, methyl ester (26.95%), and Methyl stearate (17.44%) were the predominant compounds, which give biodiesel its oxidative stability and combustion efficiency. The cold flow properties of the fuel are also improved by the presence of 9,12-Octadecadienoic acid (Z,Z)-, methyl ester (8.92%). The presence of derivatives of pentadecanoic acid further proves the lipid origin of the biodiesel. Chlorinated hydrocarbons and nonane derivatives represent examples of so-called minor chemicals which are often created due to degradation operations or minor trace impurities. Under the overall picture, the profile supports the fact that biodiesel of high quality with proper fatty acid methyl ester (FAME). The area percentage of these components in the sample shows that 95.3% of biodiesel has been produced from M. indica oil.

GC–MS analysis of produced biodiesel from M. indica oil
Performance and emission analysis
Performance analysis
Brake-specific fuel consumption (BSFC) and braking thermal efficiency (BTE) are often used metrics to assess engine performance under different workload conditions. For every experimental testing scenario in each of the test fuels used in this investigation, three records have been obtained. With changes ranging from 0 to 100% with diesel and MB30 & MB50 blending, this illustrates how engine capacity impacts BTE and BSFC. Figure 11 represents the effect of biodiesel blends (MB30, MB50) on BTE and BSFC at various loads. Any fuel’s boiling point rises gradually as engine workload increases for all fuel types. Under all loading conditions, diesel uses more BTE than the M. indica biofuels blend (MB30, MB50). The variance is attributed to the lower thermal value, higher density, and increased kinematic viscosity of M. indica biofuel blends, which collectively result in insufficient vaporizing and atomizing. Blends of M. indica biofuels have a lower BTE than diesel fuel, which had the highest BTE levels of any sample that was tested. At 100% load condition, the highest BTE value of 48.17, 38.01, and 36.42 were obtained for diesel, MB30 and MB50 samples, respectively [49]. In engine investigations, BSFC clearly demonstrates the connection between the quantity of fuel consumed per unit of energy generated [50]. M. indica blending has a better BSFC rating than regular diesel fuel for any motor freight. Studies show that the quantity of energy produced is closely correlated with the efficiency of burning. The BSFC of the diesel was found to be 0.2 kg/kWh at the 100% load condition. At the same load condition, the BSFC of MB30 & MB50 was 0.26 kg/kWh and 0.28 kg/kWh, respectively.

Effect of MB30 and MB50 blends on BTE and BSFC at various loads
Emission analysis
The effect of biodiesel blends on the hydrocarbon and carbon monoxide emission is shown in Fig. 12. Hydrocarbon exhaust is mostly caused by malfunctioning engines. Inadequate ignition also shows that BSHC pollutants are greater at lower loading stages and decrease as the load increases. Improper combustion due to an uneven fuel–air mixture results in higher levels of BSHC pollutants. Under full load condition, the HC emission of biodiesel blends MB30 and MB50 is reduced by 23.53% and 36.47%, respectively. When utilizing blend fuels, BSHC pollutants stay lower under all loading conditions since biodiesel has more oxygen than diesel and enhances combustion performance [51]. Fuel is added to the engine with each pass within the engine chamber, increasing the degree of heat of ignition as engine load increases. By achieving consistency, an in-cylinder fuel burns at higher temperatures, reducing the majority of carbon monoxide emissions. M. indica biofuels show a decreasing trend of BSCO because they have a superior oxidation process than diesel [52]. Under the full load condition, the blend MB30 and MB50 shows 11.71% and 27.93% reduction in carbon monoxide emission than that of diesel. Smoke, which is composed of carbon atoms in the form of atoms, is the end result of burning any fuel. The generation of smoke exudation is associated with the intrinsic oxygen content of the studied fuels and complete ignition. It was found that the smoke emission is reduced by 3.02% and 15.42% when operating the engine using MB30 and MB50, respectively. By varying the load and contrasting it with that of commercial diesel, the NOx emissions of the mahua biodiesel blends were studied in the diesel engine. The NOx emissions from the biodiesel blends MB 30 and MB 50 have been shown to be greater than those from diesel. The MB 30 and MB 50 show 13.34% and 15.96% greater NOx emissions than the diesel while operating at full load. It is because of the oxygen concentration in the biodiesel blends that the emission level has increased [53]. Figure 13 represents the effect of biodiesel blends on NOx and smoke emission compared to that of pure diesel.

Effect of MB30 and MB50 blends on HC emission and CO emission at various loads

Effect of MB30 and MB50 blends on smoke emission and NOx emission at various loads
Conclusion
Our research proves the effectiveness of a novel bacterial strain, Bacillus licheniformis, characterized through 16S rRNA sequencing, as a whole-cell biocatalyst for the production of biodiesel from M. indica oil. The response surface methodology for biodiesel production was performed and the optimum condition for attaining maximum yield (95.3%) was obtained at the optimum condition of methanol: oil molar ratio of 7.5:1 and concentration of immobilized cells (BSPs) of 30%w/w. The coefficient of determination (R2) to produce biodiesel from M. indica oil by whole-cell biocatalyst was found to be 99.53%. Additionally, the MB30 and MB50 biodiesel blends’ performance and emission characteristics were assessed in relation to traditional diesel. In comparison to diesel, the results showed that the MB30 and MB50 blends decreased CO emissions by 11.71% and 27.93%, respectively. Furthermore, MB30 shown reductions of 23.53% and 3.02% in hydrocarbon emission (HC) and smoke opacity, respectively, while MB50 demonstrated reductions of 36.47% and 15.42%, respectively. Utilizing biodiesel blends MB30 and MB50 increases nitrous oxide emissions by 13.34% and 15.96%, respectively.
The results of this research underscore the promise of whole-cell biocatalysts in biodiesel synthesis, providing a sustainable and efficient process compared to conventional processes. Immobilized cells not only increase enzyme stability, but also offer an economical means for large-scale biodiesel production. Future studies can aim to scale up this process and identify ways to reduce the rise of nitrous oxide emissions so that biodiesel blends can be an even more eco-friendly source for energy demands.
Data availability
No datasets were generated or analysed during the current study.
Abbreviations
- BSPs:
-
Biomass supporting particles
- GC–MS:
-
Gas chromatography–mass spectrometry
- FTIR:
-
Fourier transformed infrared spectroscopy
- MB30:
-
30% Madhuca indica Biodiesel blend
- MB50:
-
50% Madhuca indica Biodiesel blend
References
-
Schernikau L, Smith WH. Climate impacts of fossil fuels in todays electricity systems. J South Afr Inst Min Metall. 2022;122(3):1–13. https://doi.org/10.17159/2411-9717/1874/2022.
-
Kan S, Chen B, Chen G. Worldwide energy use across global supply chains: decoupled from economic growth? Appl Energy. 2019;250:1235–45. https://doi.org/10.1016/j.apenergy.2019.05.104.
-
Koyamparambath A, Santillán-Saldivar J, McLellan B, Sonnemann G. Supply risk evolution of raw materials for batteries and fossil fuels for selected OECD countries (2000–2018). Resour Policy. 2022;75: 102465. https://doi.org/10.1016/j.resourpol.2021.102465.
-
Holechek JL, Geli HME, Sawalhah MN, Valdez R. A global assessment: can renewable energy replace fossil fuels by 2050? Sustainability. 2022;14(8):4792. https://doi.org/10.3390/su14084792.
-
Nashier I, Lakra P. Integrated journal of social sciences global climate change and its effects. Integr J Soc Sci Integr J Soc Sci. 2020;2020(1):14–23.
-
Yang L, Wang XC, Dai M, et al. Shifting from fossil-based economy to bio-based economy: status quo, challenges, and prospects. Energy. 2021;228: 120533. https://doi.org/10.1016/j.energy.2021.120533.
-
Babazadeh R, Ezati F, Arumugam A. Non-edible feedstock: necessity and societal implications. In: Production of Biodiesel from Non-Edible Sources. Elsevier; 2022:81–116. https://doi.org/10.1016/B978-0-12-824295-7.00002-4
-
Abdul Hakim Shaah M, Hossain MS, Salem Allafi FA, et al. A review on non-edible oil as a potential feedstock for biodiesel: physicochemical properties and production technologies. RSC Adv. 2021;11(40):25018–37. https://doi.org/10.1039/d1ra04311k.
-
Yatish KV, Omkaresh BR, Kattimani VR, Lalithamba HS, Sakar M, Balakrishna RG. Solar energy-assisted reactor for the sustainable biodiesel production from Butea monosperma oil: optimization, kinetic, thermodynamic and assessment studies. Energy. 2023;263: 125768. https://doi.org/10.1016/j.energy.2022.125768.
-
Chowdhury S, Dhawane SH, Jha B, et al. Biodiesel synthesis from transesterified Madhuca indica oil by waste egg shell–derived heterogeneous catalyst: parametric optimization by Taguchi approach. Biomass Convers Biorefinery. 2021;11(4):1171–81. https://doi.org/10.1007/s13399-019-00512-3.
-
Geow CH, Tan MC, Yeap SP, Chin NL. A review on extraction techniques and its future applications in industry. Eur J Lipid Sci Technol. 2021;123(4):2000302. https://doi.org/10.1002/ejlt.202000302.
-
Khan Z, Javed F, Shamair Z, et al. Current developments in esterification reaction: a review on process and parameters. J Ind Eng Chem. 2021;103:80–101. https://doi.org/10.1016/j.jiec.2021.07.018.
-
Athar M, Zaidi S. A review of the feedstocks, catalysts, and intensification techniques for sustainable biodiesel production. J Environ Chem Eng. 2020;8(6): 104523. https://doi.org/10.1016/j.jece.2020.104523.
-
Salaheldeen M, Mariod AA, Aroua MK, Rahman SMA, Soudagar MEM, Fattah IMR. Current state and perspectives on transesterification of triglycerides for biodiesel production. Catalysts. 2021;11(9):1121. https://doi.org/10.3390/catal11091121.
-
Song JW, Seo JH, Oh DK, Bornscheuer UT, Park JB. Design and engineering of whole-cell biocatalytic cascades for the valorization of fatty acids. Catal Sci Technol. 2020;10(1):46–64. https://doi.org/10.1039/C9CY01802F.
-
Jeandet P, Sobarzo-Sánchez E, Silva AS, et al. Whole-cell biocatalytic, enzymatic and green chemistry methods for the production of resveratrol and its derivatives. Biotechnol Adv. 2020;39: 107461. https://doi.org/10.1016/j.biotechadv.2019.107461.
-
Nain P, Jaiswal SK, Prakash NT, Prakash R, Gupta SK. Influence of acyl acceptor blends on the ester yield and fuel properties of biodiesel generated by whole-cell catalysis of cottonseed oil. Fuel. 2020. https://doi.org/10.1016/j.fuel.2019.116258.
-
Yang RL, Zhao XJ, Wu TT, et al. A novel and highly regioselective biocatalytic approach to acetylation of helicid by using whole-cell biocatalysts in organic solvents. Catal Commun. 2019. https://doi.org/10.1016/j.catcom.2019.105707.
-
Chen G, Liu J, Qi Y, Yao J, Yan B. Biodiesel production using magnetic whole-cell biocatalysts by immobilization of Pseudomonas mendocina on Fe3O4-chitosan microspheres. Biochem Eng J. 2016;113:86–92. https://doi.org/10.1016/j.bej.2016.06.003.
-
Iyyappan J, Jayamuthunagai J, Bharathiraja B, Saravanaraj A, Praveen Kumar R, Balraj S. Production of biodiesel from Caulerpa racemosa oil using recombinant Pichia pastoris whole cell biocatalyst with double displayed over expression of Candida antartica lipase. Bioresour Technol. 2022. https://doi.org/10.1016/j.biortech.2022.127893.
-
Sena RO, Carneiro C, Moura MVH, et al. Application of Rhizomucor miehei lipase-displaying Pichia pastoris whole cell for biodiesel production using agro-industrial residuals as substrate. Int J Biol Macromol. 2021;189(April):734–43. https://doi.org/10.1016/j.ijbiomac.2021.08.173.
-
Liu J, Chen G, Yan B, Yi W, Yao J. Biodiesel production in a magnetically fluidized bed reactor using whole-cell biocatalysts immobilized within ferroferric oxide-polyvinyl alcohol composite beads. Bioresour Technol. 2022. https://doi.org/10.1016/j.biortech.2022.127253.
-
Mehrotra T, Dev S, Banerjee A, Chatterjee A, Singh R, Aggarwal S. Use of immobilized bacteria for environmental bioremediation: a review. J Environ Chem Eng. 2021;9(5): 105920. https://doi.org/10.1016/j.jece.2021.105920.
-
Daniel LMC, Pozzi E, Foresti E, Chinalia FA. Removal of ammonium via simultaneous nitrification–denitrification nitrite-shortcut in a single packed-bed batch reactor. Bioresour Technol. 2009;100(3):1100–7. https://doi.org/10.1016/j.biortech.2008.08.003.
-
Pavlova M, Draganova M. Biocompatible and biodegradable polyurethane polymers. Biomaterials. 1993;14(13):1024–9. https://doi.org/10.1016/0142-9612(93)90196-9.
-
Zhong L, Feng Y, Wang G, et al. Production and use of immobilized lipases in/on nanomaterials: a review from the waste to biodiesel production. Int J Biol Macromol. 2020;152:207–22. https://doi.org/10.1016/j.ijbiomac.2020.02.258.
-
Saleemi MA, Lim V. Overview of antimicrobial polyurethane-based nanocomposite materials and associated signalling pathways. Eur Polym J. 2022;167: 111087. https://doi.org/10.1016/j.eurpolymj.2022.111087.
-
Bhattacharjee B, Ghosh S, Patra D, Haldar J. Advancements in release-active antimicrobial biomaterials: a journey from release to relief. WIREs Nanomed Nanobiotechnol. 2022. https://doi.org/10.1002/wnan.1745.
-
Balakrishnan D, Bibiana AS, Vijayakumar A, Santhosh RS, Dhevendaran K, Nithyanand P. Antioxidant activity of bacteria associated with the marine sponge Tedania anhelans. Indian J Microbiol. 2015;55(1):13–8. https://doi.org/10.1007/s12088-014-0490-8.
-
Chen JP, Lin GH. Optimization of biodiesel production catalyzed by fungus cells immobilized in fibrous supports. Appl Biochem Biotechnol. 2010;161(1–8):181–94. https://doi.org/10.1007/s12010-009-8776-8.
-
Yan J, Zheng X, Li S. A novel and robust recombinant Pichia pastoris yeast whole cell biocatalyst with intracellular overexpression of a Thermomyces lanuginosus lipase: preparation, characterization and application in biodiesel production. Bioresour Technol. 2014;151:43–8. https://doi.org/10.1016/j.biortech.2013.10.037.
-
Tamalampudi S, Talukder MDMR, Hama S, et al. Development of recombinant Aspergillus oryzae whole-cell biocatalyst expressing lipase-encoding gene from Candida antarctica. Appl Microbiol Biotechnol. 2007;75(2):387–95. https://doi.org/10.1007/s00253-006-0814-7.
-
Soares CMF, De Castro HF, De Moraes FF, Zanin GM. Characterization and utilization of Candida rugosa lipase immobilized on controlled pore silica. Appl Biochem Biotechnol Part A Enzym Eng Biotechnol. 1999;77–79:745–57. https://doi.org/10.1007/978-1-4612-1604-9_68.
-
Nie K, Xie F, Wang F, Tan T. Lipase catalyzed methanolysis to produce biodiesel: optimization of the biodiesel production. J Mol Catal B Enzym. 2006;43(1–4):142–7. https://doi.org/10.1016/j.molcatb.2006.07.016.
-
Nini L, Sarda L, Comeau LC, Boitard E, Dubès JP, Chahinian H. Lipase-catalysed hydrolysis of short-chain substrates in solution and in emulsion: a kinetic study. Biochim Biophys Acta Mol Cell Biol Lipids. 2001;1534(1):34–44. https://doi.org/10.1016/S1388-1981(01)00172-X.
-
Fukuda H, Hama S, Tamalampudi S, Noda H. Whole-cell biocatalysts for biodiesel fuel production. Trends Biotechnol. 2008;26(12):668–73. https://doi.org/10.1016/j.tibtech.2008.08.001.
-
Soccol CR, Dalmas Neto CJ, Soccol VT, et al. Pilot scale biodiesel production from microbial oil of Rhodosporidium toruloides DEBB 5533 using sugarcane juice: performance in diesel engine and preliminary economic study. Bioresour Technol. 2017;223:259–68. https://doi.org/10.1016/j.biortech.2016.10.055.
-
Chen G, Liu J, Yao J, Qi Y, Yan B. Biodiesel production from waste cooking oil in a magnetically fluidized bed reactor using whole-cell biocatalysts. Energy Convers Manag. 2017;138:556–64. https://doi.org/10.1016/j.enconman.2017.02.036.
-
Surendhiran D, Vijay M, Sirajunnisa AR. Biodiesel production from marine microalga Chlorella salina using whole cell yeast immobilized on sugarcane bagasse. J Environ Chem Eng. 2014;2(3):1294–300. https://doi.org/10.1016/j.jece.2014.05.004.
-
Yoshida A, Hama S, Tamadani N, Noda H, Fukuda H, Kondo A. Continuous production of biodiesel using whole-cell biocatalysts: sequential conversion of an aqueous oil emulsion into anhydrous product. Biochem Eng J. 2012;68:7–11. https://doi.org/10.1016/j.bej.2012.07.002.
-
Neha A, Sethi N, Yadav S, et al. Evaluation of lipase from an indigenous isolated bacillus strain for biodiesel production. Nat Environ Pollut Technol. 2023;22(3):1319–30.
-
Maroju PA, Ganesan R, Ray DJ. Efficient biodiesel production from rice bran oil using magnetite immobilized-recombinant lipase from probiotic Bacillus licheniformis. Green Chem. 2022;24(22):8800–11. https://doi.org/10.1039/D2GC03022E.
-
Chen M, Guo Q, Wang R, et al. Construction of the yeast whole-cell Rhizopus oryzae lipase biocatalyst with high activity. J Zhejiang Univ Sci B. 2011;12(7):545–51. https://doi.org/10.1631/jzus.B1000258.
-
Sun T, Du W, Liu D. Comparative study on stability of whole cells during biodiesel production in solvent-free system. Process Biochem. 2011;46(3):661–4. https://doi.org/10.1016/j.procbio.2010.11.006.
-
Kara K, Ouanji F, Lotfi EM, El MM, Kacimi M, Ziyad M. Biodiesel production from waste fish oil with high free fatty acid content from Moroccan fish-processing industries. Egypt J Pet. 2018;27(2):249–55. https://doi.org/10.1016/j.ejpe.2017.07.010.
-
Devaraj K, Mani Y, Rawoof SAA, Thanarasu A, Dhanasekaran A, Subramanian S. Feasibility of biodiesel production from waste cooking oil: lab-scale to pilot-scale analysis. Environ Sci Pollut Res. 2020;27(20):25828–35. https://doi.org/10.1007/s11356-020-09068-6.
-
Alsaiari RA, Musa EM, Rizk MA. Biodiesel production from date seed oil using hydroxyapatite-derived catalyst from waste camel bone. Heliyon. 2023;9(5): e15606. https://doi.org/10.1016/j.heliyon.2023.e15606.
-
Nautiyal P, Subramanian KA, Dastidar MG. Production and characterization of biodiesel from algae. Fuel Process Technol. 2014;120:79–88. https://doi.org/10.1016/j.fuproc.2013.12.003.
-
Dewangan A, Mallick A, Yadav AK, Ahmad A, Alqahtani D, Islam S. Combined effect of operating parameters and nanoparticles on the performance of a diesel engine: response surface methodology-coupled genetic algorithm approach. ACS Omega. 2023;8(27):24586–600. https://doi.org/10.1021/acsomega.3c02782.
-
Kukana R, Jakhar OP. Performance, combustion and emission characteristics of a diesel engine using composite biodiesel from waste cooking oil—Hibiscus Cannabinus oil. J Clean Prod. 2022;372: 133503. https://doi.org/10.1016/j.jclepro.2022.133503.
-
Rajendran S, Venkatesan EP, Dhairiyasamy R, Jaganathan S, Muniyappan G, Hasan N. Enhancing performance and emission characteristics of biodiesel-operated compression ignition engines through low heat rejection mode and antioxidant additives: a review. ACS Omega. 2023;8(38):34281–98. https://doi.org/10.1021/acsomega.3c03252.
-
Antony Casmir Jayaseelan G, Anderson A, Manigandan S, Elfasakhany A, Dhinakaran V. Effect of engine parameters, combustion and emission characteristics of diesel engine with dual fuel operation. Fuel. 2021;302:121152. https://doi.org/10.1016/j.fuel.2021.121152.
-
Bhave NA, Gupta MM, Joshi SS. Effect of oxy hydrogen gas addition on combustion, performance, and emissions of premixed charge compression ignition engine. Fuel Process Technol. 2022;227: 107098. https://doi.org/10.1016/j.fuproc.2021.107098.
Funding
All of the sources of funding for the work described in this publication are acknowledged below: NA.
Ethics declarations
Ethics approval and consent to participate
Not applicable (manuscript does not report on or involve the use of any animal or human data or tissue)
Consent for publication
I, the Corresponding Author, declare that this manuscript has been published before in the form of patent (Application number: 202341075471).
Competing interests
The authors declare no competing interests.
Ethical responsibilities
I wish to state that all the authors mutually agree that it should be submitted to Biotechnology for Biofuels and Bioproducts. I wish to state that the present work is the original research work of the authors. I wish to state that the manuscript was not previously submitted to Biotechnology for Biofuels and Bioproducts. Results were presented clearly, honestly, and without fabrication, falsification or inappropriate data manipulation. Authors adhere to discipline-specific rules for acquiring, selecting, and processing data.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Rahul, S., Azeez, M.K.A., Nithyanand, P. et al. Optimizing biodiesel production from Madhuca indica oil using marine bacteria as a whole-cell biocatalyst: engine testing and performance analysis. Biotechnol. Biofuels Bioprod. 18, 78 (2025). https://doi.org/10.1186/s13068-025-02642-5
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1186/s13068-025-02642-5