
Introduction
Vitamin B12 (hereafter referred as B12), also known as cobalamin, is an essential water-soluble vitamin that plays a critical role in human physiology. It is a unique coordination complex of cobalt, characterized by a corrin-ring structure with two axial ligands. Cobalamins are distinguished from other cobamides by the presence of 5,6-dimethylbenzimidazole (DMBI) as the lower ligand, in contrast to adenine found in pseudocobalamins. Although structurally similar, B12 analogues are not utilized by the human body, highlighting the distinct nutritional value of active B121. Cobalamins act as crucial cofactors for enzymes such as cytoplasmic methionine synthase and methylmalonyl-CoA mutase2 and are involved in nucleotide synthesis and myelin formation essential for neural health. A deficiency in B12 can cause severe damage, particularly to the brain and nervous systems, leading to symptoms such as weakness, depression, and immune system disorders3,4. Notably, B12 is produced solely by bacteria and archaea and is not required by plants, making animal-derived food sources the exclusive providers of this vitamin for humans5.
In industrial applications, microbial fermentation is the primary method for producing B126, with Thiopseudomonas denitrificans, and Propionibacterium freudenreichii, being the main organisms used5,6,7. P. freudenreichii, a Gram-positive bacterium from the genus Propionibacterium with a high GC% content (64–70%), belongs to the phylum Actinomycetota8,9. This bacterium has a long history of use in cheese manufacture5. Recent interest has surged in fortifying food with B12 through in situ bacterial fermentation10,11,12,13,14. P. freudenreichii is particularly promising for this application due to its Generally Recognized as Safe (GRAS) status5, its efficient cobalamin production with recent reports showing production rates of up to 58.8 mg/L15,16, and its minimal production of pseudocobalamin17,18. Additionally, P. freudenreichii produces various functional metabolites, including conjugated linoleic acid (CLA), trehalose, and bifidogenic factors19.
The synthesis of cobalamin, which is unique to microorganisms, involves over 30 genes5. Previous research has elucidated the complete cobalamin synthesis pathway for P. freudenreichii20,21,22,23,24,25. Among the complex steps, the activation and attachment of the lower ligand determines whether the end product is active B12 or pseudovitamin B12. The BluB-CobT2 fusion enzyme is responsible for synthesizing the DMBI base and activating it into α-ribazole phosphate, which is then attached as the lower ligand of cobalamin18. Fusing the production and activation of DMBI within the same enzymatic complex results in high propensity toward the production of the active form of B1218. DMBI is derived from reduced flavin mononucleotide (FMN) through the action of the BluB enzyme18,26,27,28. In P. freudenreichii oxygen plays a critical role in the synthesis of the DMBI ligand. Without adequate oxygen, the DMBI ligand cannot be synthesized, leading to the production of inactive forms of B12, such as pseudocobalamin with adenine as a lower ligand base. Conversely, in the presence of excess oxygen, P. freudenreichii shifts from B12 production to heme synthesis29.
In the cobalamin synthesis pathway of P. freudenreichii, riboflavin is a precursor for the formation of DMBI. Riboflavin, or vitamin B2, is another crucial B vitamin, essential for human growth, cellular function, and metabolism. It plays a critical role in converting proteins, fats, and carbohydrates into energy30,31. Deficiency in riboflavin can lead to symptoms such as sore throat, mouth inflammation, and skin conditions31. Riboflavin is also vital for microbial metabolism as it serves as the precursor for the synthesis of the coenzymes FMN and FAD, which are essential for various redox reactions32. In cobalamin synthesis by P. freudenreichii, riboflavin is a foundational molecule for the formation of DMBI through the action to the BluB enzyme, ultimately determining the form of B12 produced. The ribA gene, encoding the GTP cyclohydrolase II enzyme, is central to the riboflavin biosynthetic pathway33. By catalyzing the early steps of riboflavin synthesis, the ribA gene indirectly influences B12 biosynthesis. Given that the key role riboflavin plays in DMBI synthesis and its further influence on B12 synthesis, understanding the relationship between riboflavin synthesis and the ribA gene in P. freudenreichii can provide insights into optimizing B12 production, identifying potential bottlenecks, and highlighting areas for future research.
Materials and methods
Bacterial strains and culture conditions
The strains and plasmids used in this study are listed in Table 1. P. freudenreichii strains were routinely cultured in yeast extract sodium lactate (YEL) medium, a standard medium for propionibacteria37. This medium is devoid of vitamin B12 but contains riboflavin provided by yeast extract. P. freudenreichii cultures were incubated routinely at 30 °C under anaerobic conditions, generated using Oxoid™ AnaeroGen™ 2.5 L Sachets (ThermoFisher) within an anerobic jar. These sachets create an atmosphere with < 1% O₂ within 30 min, and these conditions are referred to as anaerobic (pO2 < 1%) throughout this study. To analyze the effect of riboflavin on growth characteristics, Riboflavin Assay Medium (RAM) (BD™ Difco™, USA) was used. Cysteine or other chemical reductants were not added to the media for P. freudenreichii cultivation. Escherichia coli was grown in Lysogeny Broth (LB) medium. When required, erythromycin (Sigma-Aldrich, USA) was added at 200 µg/mL for E. coli and 10 µg/mL for P. freudenreichii, or ampicillin (Sigma-Aldrich, USA) was added at 50 µg/mL for E. coli.
PCR and other DNA techniques
Platinum SuperFi II DNA Polymerase, restriction enzymes and T4 ligase were obtained from Thermo ScientificTM (Darmstadt, Germany), and were used according to the instructions of the supplier. PCR products were purified with a Qiagen PCR purification kit. Plasmid DNA from E. coli clones was isolated using GenElute Plasmid Miniprep Kit (SIGMA, USA). Genomic DNA was extracted using the MagAttract HMW DNA Kit (Qiagen, Nordic).
Insertional inactivation of the bluB/cobT2 and ribA genes
The protocol for inactivating the bluB and ribA genes in P. freudenreichii was adapted from previous research with some adjustments38. First, a suicide vector was constructed by inserting an erythromycin resistance gene (ermE, GenBank: M11200.1) into a pBluescript SK + plasmid. The ermE gene was amplified by PCR with the primers ermfor and ermrev (Table 2) from genomic DNA of Saccharopolyspora erythraea DSM 40517 (DSMZ, Germany, Acc. No. PDBV01000001.1). The PCR product was then digested with EcoRV (Thermo Scientific™, Germany) and ligated with EcoRV-cut pBluescript SK +. The ligation mixture was transformed into competent E. coli DH5α and transformants were selected on LB agar (Condalab, Spain) (1.5%) containing erythromycin (200 µg/mL). Plasmid DNA was isolated from selected clones, and its identity was confirmed first by restriction digestion and agarose gel-electrophoresis, and finally by Nanopore sequencing (See below). This resulting plasmid was named pBsErmE.
For the inactivation of bluB/cobT2 two integration constructs were prepared. Primer pairs bluBcobtfor1/bluBcobtrev1 and bluBcobtfor2/bluBcobtrev2 were used to PCR amplify 550-bp and 545-bp internal bluB/cobT2 DNA fragments (Table 2), which were designed to be homologous to nucleotides 77–625 and 1151–1695 of the bluB/cobT2 coding region, respectively. These PCR products were then cloned as a EcoRI-XbaI fragment into pBsErmE, resulting in the plasmids pBsErmEbluB and pBsErmEcobT. Similarly, 607-bp ribA DNA fragment was amplified with the primers ribAfor and ribArev, which were designed to be homologous to nucleotides 63–669 of the ribA coding sequence. This PCR product was cloned as an XbaI-EcoRI fragment into pBsErmE resulting in pBsErmEribA for the inactivation of ribA.
P. freudenreichii electrocompetent cells were prepared as described previously with some modifications38. Colonies of the P. freudenreichii DSM 4902 from YEL agar plate were used to inoculate YEL medium supplemented with 0.5 M sucrose and 2% glycine and incubated at 30 °C until the OD600 reached 0.1. The cells were then harvested by centrifugation (4000×g, 15 min, 4 ℃) and washed three times with ice-cold 0.5 M sucrose. After washing, the cells were resuspended in an electroporation buffer (0.5 M sucrose, 10% glycerol, and 1 mM potassium acetate, adjusted to pH 5.5). For the electroporation step, 90 µL of the electrocompetent cells were mixed with 3–4 µg of pBsErmEbluB, pBsErmEcobT, or pBsErmEribA plasmid DNA in cooled electroporation cuvettes. The electroporation of P. freudenreichii DSM 4902 was performed with Gene Pulser XcellTM (Bio-Rad Laboratories, Richmond, CA, United States) at 20 kV/cm, 200-Ω resistance, and 25-µF capacitance followed by mixing the cells with 900 µL of YEL medium containing 0.5 M sucrose, 20 mM MgCl2, and 2 mM CaCl2 and incubation at 30 ℃ for 24 h. To select the mutants harboring inserted plasmids, the transformed cells were plated on YEL plates containing 10 µg/mL erythromycin and for pBsErmEbluB and pBsErmEcobT also 1 mg/L of vitamin B12 was added. The plates were cultivated for 7 days at 30 ℃ under anaerobic (pO2 < 1%) conditions. Despite several attempts, transformant colonies were obtained only with pBsErmEbluB and pBsErmEribA plasmids, but not with pBsErmEcobT. The mutant strains were further confirmed by colony PCR and Illumina genome sequencing (Sect.2.9).
Vitamin B12 analysis
The content of B12 in cell pellets was determined in its cyanocobalamin form and analysed using previously reported extraction and ultra-high-performance liquid chromatographic (UHPLC) methods11,12. P. freudenreichii DSM 4902 strain, P.f ∆bluB mutant, and P.f ∆ribA mutant were grown at 30 ℃ in 10 mL of medium for 4 days in triplicates. Cells were harvested by centrifugation, and both the pellet and the supernatant were stored at −20 °C. The cell pellet suspended in 10 mL of the extraction buffer (8.3 mM sodium hydroxide and 20.7 mM acetic acid, adjusted to pH 4.5) and supplemented with 1% sodium cyanide was subjected to boiling water extraction (for 30 min) followed by centrifugation upon cooling to obtain 25 mL of the final extract. The extract was then analyzed with a Waters Acquity UPLC system (Milford, MA, US) equipped with a photodiode array detector (PDA; 210 ~ 600 nm) using an Acquity HSS T3 C18 column (2.1 × 100 mm, 1.8 μm). The mobile phase was a gradient flow of MilliQ water and acetonitrile, both containing 0.025% TFA. The chromatogram was obtained by recording the absorbance at 361 nm. Six cyanocobalamin standards (0.015 ~ 0.075 ng/µL) were injected (10 µL in duplicate) to create a calibration curve for each sample set. Cellular B12 yield (µg/g fresh cell biomass) for each strain is reported as the average of three biological replicates.
Furthermore, the identity of the cobamide in the extracts (as cyanocobalamin) was confirmed with UPLC-MS/MS using a high resolution quadrupole time-of-flight mass spectrometer (Q-TOF, Synapt G2-Si; Waters, MA, USA) following the instrumental settings reported previously12.
Measurement of riboflavin content
Riboflavin was extracted from the supernatants of the cultures mentioned above (See Vitamin B12 analysis) using the European standard method (EN 14152:2014) with some modifications and analysed with a UHPLC method11. Briefly, the culture supernatant (1mL) was mixed with 15 mL of 0.1 M hydrochloric acid and extracted in a boiling water bath for 60 min. After cooling to the room temperature, the pH was adjusted to 4.5 with 2.5 M sodium acetate and the extract was incubated for 24 h at 37℃ with Taka-Diastase (50 mg; Pfaltz and Bauer, CT, USA) and β-amylase (5 mg; Sigma-Aldrich). The analysis was performed with a Waters UPLC system equipped with a fluorescence detector using a Waters Acquity BEH C18 column (2.1 mm × 100 mm; 1.7 μm). The mobile phase contained 20 mM ammonium acetate dissolved in 30% aqueous methanol and was eluted at a constant flow of 0.2 mL/min. The excitation and emission wavelengths were set at 432 nm and 520 nm respectively. An external calibration curve obtained by injecting six riboflavin standards (0.01 ~ 1 ng/µL) in duplicate was used for the quantitation.
Analysis of acids
The supernatants from the strains and control media were diluted with MilliQ water and filtered (0.45 μm; Pall, USA) to HPLC vials. Succinic acid, lactic acid acetic acid and propionic acid were quantified using the HPLC method reported previously11,39. The analysis was performed using an HPLC system equipped with a pump (Waters 515), an autosampler, a UV detector (Waters 717) and a refractive index detector (HP 1047 A, HP, USA) on a Hi-Plex H column (Agilent, CA, USA; 300 mm × 6.5 mm) as a pre-column and HiPlex H (Agilent, CA, USA; 50 mm × 7.7 mm) as a guard column. 10 mM H2SO4 was used as a mobile phase with the flow rate set at 0.5 mL/min for 30 min with the column temperature maintained at 40 °C.
Impact of vitamin B12 on the metabolism of P. freudenreichii: an isothermal calorimetric analysis
The heat flow measurements were conducted following the protocol established by40 with some modifications. Both P. freudenreichii DSM 4902 and P.f ∆bluB (with 10 µg/mL erythromycin addition) inoculated from 3-day liquid cultures were cultured in YEL media at 30 °C overnight. The cultures were diluted in fresh YEL media to achieve OD600 0.05, preparing them for subsequent analyses under two distinct environments: a micro-oxygen condition, which maintained an anaerobic atmosphere without explicitly removing oxygen, and a fully anaerobic setting, wherein oxygen was actively excluded by normalising the media in an anaerobic chamber (Whitley A85 anaerobic workstation, Don Whitley Scientific, UK) over night. Additionally, both strains were grown in the presence and absence of B12 supplementation (1 µg/mL).
300 µL of the inoculated cultures were then transferred into plastic inserts (calWells, Symcel, Sweden) in four biological replicates, housed within titanium vials of a 48-well calPlate (Symcel, Sweden). The calPlate was incubated at 30 °C for 72 h within the calScreener (Symcel, Sweden), where heat flow signals were continuously recorded. For concurrent optical density measurements, the cultures (200 µL) were loaded to a 96-well microtiter plate (82.1581001, SARSTEDT, Germany), with the anaerobic culture prepared anaerobically (Whitley A85 anaerobic workstation, Don Whitley Scientific, UK) in one half of the plate and sealed with a clear Adhesive Sealing Sheet (Thermo-Fischer Scientific), and the micro-oxygen cultures were inoculated outside the anaerobic chamber. This plate was then incubated in the Varioskan Flash (Thermo Fisher Scientific, USA) at 30 °C for 72 h, during which OD600 readings were taken every two h.
For determination of growth-limiting levels of vitamin B12, both the DSM 4902 strain and its mutant derivative were prepared in the same way as above, but the P.f ∆bluB inoculum was supplemented with a gradient of vitamin B12 (cyanocobalamin) concentrations, specifically at 1.00 µg/mL, 0.50 µg/mL, 0.10 µg/mL, 0.05 µg/mL, 0.01 µg/mL, and 0.005 µg/mL to examine the dose-dependent effects of B12 on the metabolic activity of P.f ∆bluB compared to unsupplemented wild type, under fully anaerobic conditions.
Impact of riboflavin on the growth of P. freudenreichii
The growth curves were determined by following optical density at 600 nm wavelength in cultures using a Varioskan LUX Multimode Microplate Reader (Thermo Fisher Scientific, USA). Both P. freudenreichii DSM 4902 and P.f ∆ribA (with 10 µg/mL erythromycin addition) were cultured in 10 mL of YEL media under anaerobic (pO2 < 1%) conditions at 30 °C for 3 days. The cultures were then centrifuged; washed with RAM medium and resuspended in fresh RAM medium to an OD600 value of 0.03.
For the subsequent growth curve measurements, 200 µL of the cultures were prepared in 6 replicates in a 96 well microplate (Thermo Fisher Scientific, USA) under micro-oxygen condition (10% O2; maintained by N2 gas flushing). P.f ∆ribA strains were inoculated with 10 µg/mL erythromycin. To determine the growth-limiting levels of riboflavin, both the DSM 4902 strain and P.f ∆ribA inoculum were supplemented with a gradient of riboflavin concentrations: 0.00 µg/mL, 0.01 µg/mL, 0.05 µg/mL, 0.10 µg/mL, 0.50 µg/mL, 1.00 µg/mL. This was done to examine the impact of different riboflavin doses on growth of P.f ∆ribA compared to supplemented wild type.
The inoculated cultures were then transferred into plastic inserts Nunc™ Microwell 96-Well Microplate with Nunclon™ Supra Surface (Thermo Fisher Scientific, USA). The microplate was incubated in the Varioskan at 30 °C, 10% O2 for 72 h, during which OD600 readings were taken every two h.
Sequencing and bioinformatics
For plasmid sequencing, E. coli strain carrying pBsErmE was cultivated overnight at 37 °C with shaking at 225 rpm (Forma Orbital Shaker, Thermo-Fischer Scientific). The cells were harvested via centrifugation at 8000 × g for 1 min (Accu Spin 17, Thermo-Fischer Scientific, Germany). Plasmid DNA was extracted from the pelleted cells using a GenElute Plasmid Miniprep Kit (Sigma-Aldrich, USA) according to the manufacturer’s instructions. Libraries were prepared using Native Barcoding Kit 96 V14 from Oxford Nanopore, UK (SQK-NBD114.96) and barcoding protocol NBE_9171_v114_revO_15sep2022. Libraries were loaded and sequenced on a PromethION 2 Solo (PRO-SEQ002) for 72 h using standard settings. Sequencing data were base-called and assembled using the workflow (https://github.com/yanhui09/ONTrapid) with some modifications. Flye assembler was used for assembly with the parameters: –nano-corr, –plasmid, –min-overlap 5000. The plasmid was annotated with pLannotate41.
For the P.f ∆bluB and P.f ∆ribA mutants DNA sequencing was performed at CeGaT (Germany). Briefly, 100 ng of genomic DNA was used for library preparation with Illumina DNA Prep kit (Illumina) and sequenced as 2 × 100 bp reads on NovaSeq 6000 sequencer (Illumina). The reads were demultiplexed using Illumina bcl2fastq v2.20 and the adapters were trimmed with Skewer v.0.2.2. The quality of the reads was assessed by FastQC v.0.11.5-cegat. The reads were then assembled with SPAdes genome assembler v3.14.142. The resulting contigs were searched for the vector insertion sites using Megablast and annotated with PROKKA43. Additionally, the trimmed reads were mapped to the reference genome LT618787 through alignment via Burrows-Wheeler transformation tool BWA v.0.7.17-r118844 to search for deletion site and exclude the possibility of off-target mutations.
Riboswitch Scanner45,46 was used for detecting riboswitches in the genome sequence of P. freudenreichii DSM 490236.
Results and discussion
Insertional inactivations of bluB/cobT2 and ribA genes
An integrative plasmid suitable for gene inactivation in P. freudenreichii was constructed by cloning ermE from S. erythraea DSM 40517 into the E. coli vector pBluescript SK+ (Additional Fig. 1). The ermE has previously been successfully used as a selection marker in P. freudenreichii47. In this study, homologous regions in the integration constructs ranged from 545 to 607-bp. Since the E. coli vector containing the internal fragments of P. freudenreichii genes cannot persist in P. freudenreichii, erythromycin-resistant colonies are expected to arise exclusively through single crossover recombination events with the corresponding chromosomal allele. Despite repeated attempts, no erythromycin-resistant transformants were obtained using the construct with the shortest homologous region (545 bp) located within the cobT region of the bluB/cobT2 gene. Although a shorter homologous region (520 bp) was previously sufficient for insertional inactivation of the slpB in P. freudenreichii38the absence of transformants in the current study with a 545 bp homologous region may reflect lower recombination efficiency compared to other constructs with slightly longer homologous regions. Another possible explanation is that mutations in the cobT region might be lethal to the strain, even when the vitamin B12 is supplemented. In contrast, with the other two integration constructs, erythromycin-resistant transformants were obtained following electroporation. Subsequent genomic DNA sequencing and bioinformatics analyses confirmed that integration events occurred at predicted genomic locations (Additional Fig. 2).

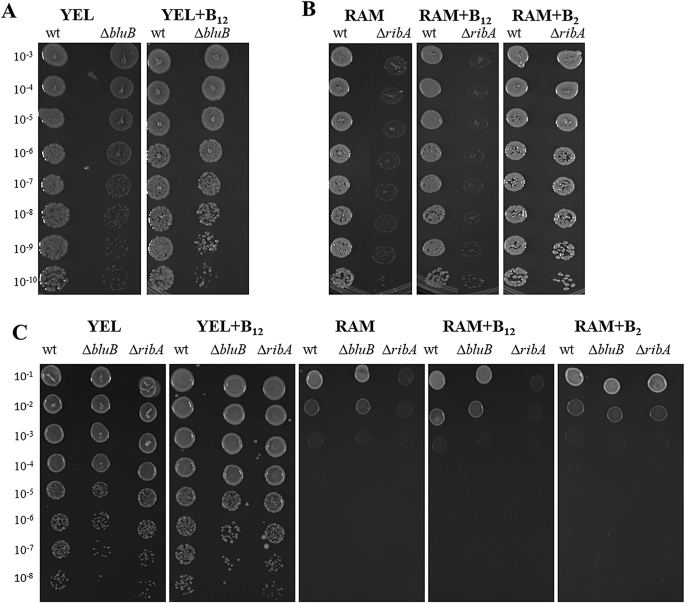
Comparison of growth of DSM 4902 (wt), P.f ∆ribA (∆ribA), and P.f ∆bluB (∆bluB) under anaerobic (pO2 < 1%) (< 1% oxygen) (A and B) and aerobic conditions (~ 20% oxygen) (C) on YEL and RAM agar without and with supplementation of vitamin B12 (+ B12), or riboflavin (+ B2) after 4–6 days of incubation at 30 °C. Cell cultures were serially diluted and spotted onto the different agar media.

(A): B12 production of DSM 4902 (wt), P.f ∆ribA (∆ribA), and P.f ∆bluB (∆bluB) in YEL medium supplemented with CoCl2 at 30 °C under normal atmospheric conditions after 7 days of fermentation. (B): Organic acid production by the same strains under identical conditions. (B): Organic acids of DSM 4902 (wt), P.f ∆ribA (∆ribA), and P.f ∆bluB (∆bluB) in YEL medium supplemented with CoCl2 at 30 °C under normal atmosphere in presence of O2 for 7 days. All results are shown as mean ± standard deviation (SD) from three biological replicates (n = 3). Bars with the same letters indicate no significant differences between groups; bars with different letters denote significant differences (p < 0.05).
Sequence analyses of the genomic arrangement in the P. freudenreichii bluB/cobT mutant, named as P.f ∆bluB, revealed that integration created two truncated copies of the bluB/cobT2 gene. The first copy lacks 1251 bp from the 3’ end of the coding region, while the second copy lacks the first 93 nucleotides of the coding region at the 5’ end, in addition to the upstream control region of the gene. Between these copies, 4388 bp of vector sequence is inserted. In the P. freudenreichii ribA mutant, named as P.f ∆ribA, genomic sequencing revealed two truncated copies of ribA separated by a 4.4 kb vector sequence. The first copy of ribA lacks 786 bp from the 3’ end of the coding region, and the second copy lacks the first 62 nucleotides of the coding region at the 5’ end, as well as the upstream control region of the gene (Additional Fig. 2).
Phenotypic characterization of the P.f ∆bluB and P.f ∆ribA strains
To assess the colony forming ability of the mutants in the absence of B12 and riboflavin, serially diluted cultures were spotted on YEL (riboflavin-containing) or RAM (riboflavin deficient) agar plates. Media were either unsupplemented or supplemented with B12 or riboflavin as appropriate. Plates were incubated at 30 °C for six days under either anaerobic (pO2 < 1%; AnaeroGen system) or in a CO2 incubator (~ 20% O2, 5% CO2) (Fig. 1).
As expected, under anaerobic (pO2 < 1%) conditions (Fig. 1A and B), the wild-type formed visible colonies at all dilutions (10−3 to 10−10) on both media. In contrast, P.f ∆bluB strain showed reduced colony size on YEL agar, exhibiting a partial growth defect that could be restored by B12 supplementation (Fig. 1A). This indicates that P. freudenreichii possesses a transport system capable of importing B12 from the growth media, although this transporter remains to be genetically identified. Conversely, P.f ∆ribA exhibited minimal growth on riboflavin-deficient RAM agar, but its growth was fully restored with riboflavin supplementation (Fig. 1B), suggesting the presence of an as-yet-uncharacterized riboflavin transport system.
Under aerobic (~ 20% O2) conditions, all strains formed colonies of roughly equal size across all dilutions (10−1 to 10−8) on YEL agar, regardless of B12 supplementation (Fig. 1C). On RAM agar, growth was restricted to the lowest dilutions (10−1−10−2) and P.f ∆ribA again grew only when riboflavin was supplemented (Fig. 1C). Interestingly, P.f ∆bluB exihibited wild-type-like growth on both media, regardless of B₁₂ supplementation (Fig. 1C). These results indicate that riboflavin is essential under both anaerobic (pO2 < 1%) and aerobic (~ 20% oxygen) conditions, whereas the growth-promoting effect of B12 is limited to low-oxygen conditions.
The phenotype of the ribA disruption mutant is consistent with the predicted function of ribDEAH operon (locus tags PFR_JS15-1_2050, PFR_JS15-1_2049, PFR_JS15-1_2048, PFR_JS15-1_2047), which likely encodes the riboflavin biosynthesis machinery. An FMN riboswitch located approximately 100 nucleotides upstream may regulate this operon. Another FMN riboswitch was identified upstream of PFR_JS15-1_1083, encoding a putative ABC transporter, whose possible involvement in riboflavin uptake warrants further investigation48.
The partial growth defect of the P.f ∆bluB strain under anaerobic (pO2 < 1%) conditions confirms that that B₁₂ biosynthesis is important, but not strictly essential, for growth in anaerobic (pO2 < 1%) conditions. In contrast, the P.f ∆bluB was indistinguishable from the wild type under aerobic conditions (~ 20% oxygen), suggesting that B₁₂ is dispensable in higher oxygen concentrations. This is consistent with recent reports showing that P. freudenreichii can grow well under aerobic bioreactor conditions while producing minimal or no B₁₂29.
These observations reflect the hybrid nature of B₁₂ biosynthesis in P. freudenreichii, which uses the anaerobic (cobalt-early) pathway for the core corrin ring and cobalt insertion5,25 but relies on the oxygen-dependent enzyme BluB to synthesize DMBI, the lower ligand of cobalamin28. Therefore, B₁₂ biosynthesis in P. freudenreichii depends on oxygen only at the final stage, creating a physiological constraint: under oxygen-limited conditions, demand for B₁₂ increases due to metabolic requirements (e.g., for methylmalonyl-CoA mutase activity), yet the ability to synthesize B₁₂ is reduced due to impaired BluB function. This explains the importance of exogenous B₁₂ under anaerobic conditions and the paradox of low B₁₂ production despite robust growth in oxygen-rich environments.
B12 and organic acids production by wild type, P.f ∆ribA and P.f ∆bluB strains
To further investigate the impact of impaired B12 biosynthesis in P. freudenreichii, we cultivated the wild type, P.f ∆ribA, and P.f ∆bluB strains in YEL medium supplemented with CoCl2. Cultures were incubated statically under atmospheric conditions for seven days. While initially oxygenated, the medium likely became progressively oxygen-depleted over time, simulating oxygen-limited conditions. Given the lack of active O₂ control, we avoid the term “microaerobic”. YEL medium, supported the growth of P.f ∆ribA owing to riboflavin present in the yeast extract. After seven days, the P.f ∆bluB strain reached a cell-density ~ 50% of that of the wild type. B12 concentrations reached 10.91 µg/g (wild type) and 13.5 µg/g (P.f ∆ribA) but remained below the detection limit in the P.f ∆bluB strain (Fig. 2A).
Organic acid profiling showed that lactate was fully consumed by wild type and P.f ∆ribA cultures, while P.f ∆bluB consumed only 50%. Propionate and acetate production by P.f ∆bluB was also ~ 50% lower than by the other strains (Fig. 2B). These results confirm that B₁₂ deficiency impairs both growth and metabolic output under oxygen-limited conditions. Alternatively, trace amounts of B12 below the detection threshold of our assay may suffice to support limited growth.
Prior data29 further confirm that P. freudenreichii can grow well under aerobic conditions without producing substantial B₁₂. This implies that the dependence on B₁₂ is condition-specific, linked to the requirement of certain metabolic reactions (e.g., methylmalonyl-CoA mutase) that are more active or essential under low-oxygen conditions.
Isothermal calorimetric analysis of wild type and P.f ∆blub strains
To gain insight into the role of B12 on the metabolic activity, we conducted isothermal calorimetric analyses comparing the wild type and the P.f ∆bluB mutant strains. Heat flow and OD₆₀₀ were monitored over 72 h under both aerobic and anaerobic conditions, with or without B₁₂ supplementation (Fig. 3A).

(A) Isothermal microcalorimetry profiles and optical density (OD600) measurements of P. freudenreichii DSM 4902 (WT) and P. f ∆bluB strains in the presence of B12 and oxygen; (B) Isothermal microcalorimetry profiles and OD600 measurements of the same strains in the presence of varying concentrations of B12.
When B₁₂ was supplemented, P.f ∆bluB reached OD₆₀₀ and accumulated heat values similar to wild type. Oxygen also enhanced total heat output by ~ 60% in the wild type and ~ 50% in P.f ∆bluB. However, oxygen delayed the peak heat flow by 2–4 h, suggesting a metabolic shift in response to O₂.
To identify the minimal B₁₂ concentration needed to restore P.f ∆bluB growth, we conducted a dose-response experiment under strictly anaerobic conditions. At ≥ 0.1 µg/mL B₁₂, P.f ∆bluB achieved growth comparable to wild type, with peak heat production reached within 25 h (Fig. 3B). Lower concentrations (≤ 0.01 µg/mL) delayed and diminished heat output. These data indicate that at least 0.1 µg/mL of B₁₂ is required to support robust anaerobic growth.
Compared to the vitamin B₁₂ production yields of other bacteria (e.g., Bacillus megaterium, Acetobacter pasteurianus)49,50P. freudenreichii requires relatively high intracellular concentrations to maintain full metabolic activity, supporting the idea that this species evolved efficient B₁₂ biosynthesis to meet high cellular demand.
Impact of riboflavin concentration on growth of wild-type and P. f ∆riba
To determine the minimal riboflavin concentration required for growth, we cultured wild-type and P. f ∆ribA strains in RAM medium under 10% O₂. While RAM is not optimized for P. freudenreichii, its lack of riboflavin makes it suitable for this purpose.
Wild-type growth was unaffected by varying riboflavin concentrations. P.f ∆ribA grew comparably to wild type when supplemented with ≥ 0.05 µg/mL riboflavin. Lower concentrations (0.01 and 0.00 µg/mL) led to reduced growth rates and final OD₆₀₀ (Fig. 4). These results confirm that riboflavin is essential under low-O₂ conditions and that supplementation at ≥ 0.05 µg/mL is sufficient to restore growth in P.f ∆ribA.

Growth curves of P. freudenreichii DSM 4902 (wild type) and P.f ∆ribA mutant in RAM medium supplemented with varying concentrations of riboflavin. Optical density at 600 nm (OD₆₀₀) was measured over time, and values represent the mean of six biological replicates. The ∆ribA strain (ribA) was tested under riboflavin concentrations of 0.00, 0.01, 0.10, and 1.00 µg/mL; wild type (wt) was included with 0.00 and 1.00 µg/mL as controls. The data are presented using a linear y-axis to accurately depict growth dynamics across all phases.
Effect of B12 supplementation on wild type, P.f ∆ribA and P.f ∆bluB strains
Based on isothermal calorimetric results, we next evaluated whether supplementing cultures with 0.1 µg/mL B₁₂ would normalize growth, organic acid production, and intracellular B₁₂ levels across all strains under B₁₂-producing conditions (YEL supplemented with CoCl2 and under a microaerobic atmosphere)29,34,35. After seven days, all strains reached comparable final cell densities and acid production levels (Fig. 5). Lactate was exhausted in all cases, and B₁₂ concentrations were similar across strains. These results confirm that 0.1 µg/mL B₁₂ is sufficient to fully restore growth and metabolism in P.f ∆bluB under these conditions.

(A) B12 production of DSM 4902 (Wild Type), P.f ∆ribA (∆ribA), and P.f ∆bluB (∆bluB) in YEL medium supplemented with CoCl2 and 0.1 µg/mL of vitamin B12 incubated at 30 °C under normal atmosphere for 7 days. (B) Organic acid production by the same strains under identical conditions. All results are shown as mean ± standard deviation (SD) from three biological replicates (n = 3).
Importantly, P.f ∆ribA showed no defect in B₁₂ synthesis or metabolic output when supplemented with riboflavin, confirming that endogenous riboflavin biosynthesis does not affect B12 production when exogenous riboflavin is available.
Conclusions
In this study, we successfully constructed two mutant derivatives of P. freudenreichii DSM 4902, each lacking the ability to synthesize either vitamin B12 or B2, through insertional inactivation of a key biosynthesis gene in each respective pathway. The genetic modifications in the bluB and ribA mutants was confirmed via genome sequencing and bioinformatics analysis. Phenotypic characterization showed that P. freudenreichii can import both vitamins from its environment. We found that riboflavin (B2) is essential for optimal growth under both aerobic and anaerobic (pO2 < 1%) conditions, while B12 becomes particularly important under oxygen-limited conditions.
Our results demonstrated impaired growth and organic acid production in the absence of B12, particularly under low oxygen availability. Supplementation experiments revealed that 0.10 µg/mL of B12 and 0.05 µg/mL are sufficient to restore normal growth in the respective mutants. Notably, the presence of exogenous B₂ eliminated the dependency on de novo riboflavin synthesis for B₁₂ production, confirming that riboflavin does not limit cobalamin biosynthesis when provided externally.
Importantly, P. freudenreichii follows the anaerobic (cobalt-early) pathway for corrin ring and cobalt insertion steps in B₁₂ biosynthesis, which are oxygen-independent. However, it requires oxygen to complete the biosynthesis via the enzyme BluB, which catalyzes the formation of the lower ligand base DMBI. This dependency creates a metabolic bottleneck: under anaerobic conditions, B₁₂-dependent metabolic reactions become more important, yet BluB activity is impaired due to the lack of oxygen. In contrast, under aerobic conditions, the need for B₁₂ diminishes despite favorable conditions for its synthesis. These findings help explain the observed disconnect between oxygen availability and B₁₂ production.
In summary, this study refines our understanding of the interplay between oxygen, vitamin biosynthesis, and metabolic requirements in P. freudenreichii. It highlights how oxygen availability shapes both the need for and the capacity to synthesize B₁₂ and suggests that optimizing cobalamin production in this organism will require tight control of oxygen to balance enzymatic activity and metabolic demand. Future studies employing transcriptomic and metabolic profiling under defined oxygen gradients will be valuable for elucidating the regulatory mechanisms governing this balance.
Data availability
Sequence data of pBsErmE vector constructed in this study has been deposited to GenBank with an accession number PP857440. Other data is provided within the manuscript or supplementary information files or are available from the corresponding author on reasonable request.
References
-
Butler, P. A. & Kräutler, B. Biological organometallic chemistry of B12 in Bioorganometallic Chemistry (ed Simonneaux, G.) 1–55 (Springer, (2006).
-
Nielsen, M. J., Rasmussen, M. R., Andersen, C. B. F., Nexø, E. & Moestrup, S. K. Vitamin B12 transport from food to the body’s cells—a sophisticated, multistep pathway. Nat. Rev. Gastroenterol. Hepatol.9, 345–354. https://doi.org/10.1038/nrgastro.2012.76 (2012).
-
Reynolds, E. Vitamin B12, folic acid, and the nervous system. Lancet Neurol.5, 949–960. https://doi.org/10.1016/S1474-4422(06)70598-1 (2006).
-
O’Leary, F. & Samman, S. Vitamin B12 in health and disease. Nutrients2, 299–316. https://doi.org/10.3390/nu2030299 (2010).
-
Martens, J. H., Barg, H., Warren, M. & Jahn, D. Microbial production of vitamin B12. Appl. Microbiol. Biotechnol. 58, 275–285. https://doi.org/10.1007/s00253-001-0902-7 (2002).
-
Fang, H., Kang, J. & Zhang, D. Microbial production of vitamin B12: A review and future perspectives. Microb. Cell. Fact.16, 15. https://doi.org/10.1186/s12934-017-0631-y (2017).
-
Rudra, B. & S Gupta, R. Phylogenomic And comparative genomic analyses of species of the family pseudomonadaceae: proposals for the genera halopseudomonas gen. Nov. And atopomonas gen. Nov., merger of the genus oblitimonas with the genus thiopseudomonas, And transfer of some misclassified species of the genus Pseudomonas into other genera. Int. J. Syst. Evol. Microbiol. 71, 005011. https://doi.org/10.1099/ijsem.0.005011 (2021).
-
Scholz, C. F. P. & Kilian, M. The natural history of cutaneous propionibacteria, and reclassification of selected species within the genus Propionibacterium to the proposed Nov.l genera Acidipropionibacterium gen. Nov., Cutibacterium gen. Nov. and Pseudopropionibacterium gen. Nov.. Int. J. Syst. Evol. Microbiol.66, 4422–4432. https://doi.org/10.1099/ijsem.0.001367 (2016).
-
Oren, A. & Garrity, G. M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol.71, 005056. https://doi.org/10.1099/ijsem.0.005056 (2021).
-
Capozzi, V., Russo, P., Dueñas, M. T., López, P. & Spano, G. Lactic acid bacteria producing B-group vitamins: A great potential for functional cereals products. Appl. Microbiol. Biotechnol.96, 1383–1394. https://doi.org/10.1007/s00253-012-4440-2 (2012).
-
Chamlagain, B. et al. Effect of the lower ligand precursors on vitamin B12 production by food-grade propionibacteria. LWT72, 117–124. https://doi.org/10.1016/j.lwt.2016.04.023 (2016).
-
Chamlagain, B., Edelmann, M., Kariluoto, S., Ollilainen, V. & Piironen, V. Ultra-high performance liquid chromatographic and mass spectrometric analysis of active vitamin B12 in cells of Propionibacterium and fermented cereal matrices. Food Chem.166, 630–638. https://doi.org/10.1016/j.foodchem.2014.06.068 (2015).
-
Gu, Q., Zhang, C., Song, D., Li, P. & Zhu, X. Enhancing vitamin B12 content in soy-yogurt by Lactobacillus reuteri. Int. J. Food Microbiol.206, 56–59. https://doi.org/10.1016/j.ijfoodmicro.2015.04.033 (2015).
-
Molina, V., Médici, M., Font de Valdez, G. & Taranto, M. P. Soybean-based functional food with vitamin B12-producing lactic acid bacteria. J. Funct. Foods. 4, 831–836. https://doi.org/10.1016/j.jff.2012.05.011 (2012).
-
Wang, P., Zhang, Z., Jiao, Y., Liu, S. & Wang, Y. Improved propionic acid and 5,6-dimethylbenzimidazole control strategy for vitamin B12 fermentation by Propionibacterium freudenreichii. J. Biotechnol.193, 123–129. https://doi.org/10.1016/j.jbiotec.2014.11.019 (2015).
-
Calvillo, Á., Pellicer, T., Carnicer, M. & Planas, A. Bioprocess strategies for vitamin B12 production by microbial fermentation and its market applications. Bioengineering9, 365. https://doi.org/10.3390/bioengineering9080365 (2022).
-
Friedmann, H. C. Partial purification and properties of a single displacement trans-N-glycosidase. J. Biol. Chem.240, 413–418. https://doi.org/10.1016/S0021-9258(18)97664-7 (1965).
-
Deptula, P. et al. Blub/CobT2 fusion enzyme activity reveals mechanisms responsible for production of active form of vitamin B12 by Propionibacterium freudenreichii. Microb. Cell. Fact.14, 186. https://doi.org/10.1186/s12934-015-0363-9 (2015).
-
Poonam, P., Pophaly, S. D., Tomar, S. K., De, S. & Singh, R. Multifaceted attributes of dairy propionibacteria: A review. World J. Microbiol. Biotechnol.28, 3081–3095. https://doi.org/10.1007/s11274-012-1117-z (2012).
-
Kiatpapan, P. & Murooka, Y. Construction of an expression vector for propionibacteria and its use in production of 5-aminolevulinic acid by Propionibacterium freudenreichii. Appl. Microbiol. Biotechnol.56, 144–149. https://doi.org/10.1007/s002530100603 (2001).
-
Falentin, H. et al. The complete genome of Propionibacterium freudenreichii CIRM-BIA1T, a hardy actinobacterium with food and probiotic applications. PLoS One5, e11748. https://doi.org/10.1371/journal.pone.0011748 (2010).
-
Hashimoto, Y., Yamashita, M. & Murooka, Y. The Propionibacterium freudenreichii HemYHBXRL gene cluster, which encodes enzymes and a regulator involved in the biosynthetic pathway from glutamate to protoheme. Appl. Microbiol. Biotechnol.47, 385–392. https://doi.org/10.1007/s002530050945 (1997).
-
Piao, Y. et al. Production of vitamin B12 in genetically engineered Propionibacterium freudenreichii. J. Biosci. Bioeng.98, 167–173. https://doi.org/10.1016/S1389-1723(04)00261-0 (2004).
-
Sattler, I. et al. Cloning, sequencing, and expression of the uroporphyrinogen III methyltransferase CobA gene of Propionibacterium freudenreichii (shermanii). J. Bacteriol.177, 1564–1569. https://doi.org/10.1128/jb.177.6.1564-1569.1995 (1995).
-
Roessner, C. A., Huang, K. X., Warren, M. J., Raux, E. & Scott, A. I. Isolation and characterization of 14 additional genes specifying the anaerobic biosynthesis of cobalamin (vitamin B12) in Propionibacterium freudenreichii (P. shermanii). Microbiology 148, 1845–1853. https://doi.org/10.1099/00221287-148-6-1845 (2002).
-
Collins, H. F. et al. Bacillus megaterium has both a functional BluB protein required for DMB synthesis and a related flavoprotein that forms a stable radical species. PLoS One8, e55708. https://doi.org/10.1371/journal.pone.0055708 (2013).
-
Gray, M. J. & Escalante-Semerena, J. C. Single-enzyme conversion of FMNH2 to 5,6-dimethylbenzimidazole, the lower ligand of B12. Proc. Natl. Acad. Sci. U. S. A.104, 2921–2926. https://doi.org/10.1073/pnas.0609270104 (2007).
-
Taga, M. E., Larsen, N. A., Howard-Jones, A. R., Walsh, C. T. & Walker, G. C. BluB cannibalizes flavin to form the lower ligand of vitamin B12. Nature 446, 449–453. https://doi.org/10.1038/nature05611 (2007).
-
Loivamaa, I. et al. Aerobic adaptation and metabolic dynamics of Propionibacterium freudenreichii DSM 20271: insights from comparative transcriptomics and surfaceome analysis. mSystems 9, e00615–00624. https://doi.org/10.1128/msystems.00615-24 (2024).
-
Powers, H. J. Riboflavin (vitamin B-2) and health. Am. J. Clin. Nutr.77(6), 1352–1360. https://doi.org/10.1093/ajcn/77.6.1352 (2003).
-
Thakur, K., Tomar, S. K., Singh, A. K., Mandal, S. & Arora, S. Riboflavin and health: A review of recent human research. Crit. Rev. Food Sci. Nutr.57, 3650–3660. https://doi.org/10.1080/10408398.2016.1145104 (2017).
-
Hiltunen, H. M. & Thesis, P. D. Functional analysis of RIBA, the introductory enzyme for riboflavin biosynthesis. Humboldt-Universität zu Berlin (2016).
-
Lin, J. W., Chao, Y. F. & Weng, S. F. Riboflavin synthesis genes ribE, ribB, ribH, RibA reside in the Lux Operon of Photobacterium leiognathi. Biochem. Biophys. Res. Commun. 284, 587–595. https://doi.org/10.1006/bbrc.2001.5013 (2001).
-
Dank, A. et al. Propionibacterium freudenreichii thrives in microaerobic conditions by complete oxidation of lactate to CO2. Environ. Microbiol.23, 3116–3129. https://doi.org/10.1111/1462-2920.15532 (2021).
-
Dank, A., Biel, G., Abee, T. & Smid, E. J. Microaerobic metabolism of lactate and propionate enhances vitamin B12 production in Propionibacterium freudenreichii. Microb. Cell. Fact.21, 225. https://doi.org/10.1186/s12934-022-01945-8 (2022).
-
Deptula, P. et al. De novo assembly of genomes from long sequence reads reveals uncharted territories of Propionibacterium freudenreichii. BMC Genomics18, 790. https://doi.org/10.1186/s12864-017-4165-9 (2017).
-
Malik, A. C., Reinbold, G. W. & Vedamuthu, E. R. An evaluation of the taxonomy of Propionibacterium. Can. J. Microbiol.14, 1185–1191. https://doi.org/10.1139/m68-199 (1968).
-
do Carmo, F. L. R. et al. Propionibacterium freudenreichii surface protein SlpB is involved in adhesion to intestinal HT-29 cells. Front. Microbiol.8, 1033. https://doi.org/10.3389/fmicb.2017.01033 (2017).
-
Hugenschmidt, S., Schwenninger, S. M., Gnehm, N. & Lacroix, C. Screening of a natural biodiversity of lactic and propionic acid bacteria for folate and vitamin B12 production in supplemented Whey permeate. Int. Dairy. J. 20, 852–857. https://doi.org/10.1016/j.idairyj.2010.05.005 (2010).
-
Beilharz, K. et al. Protocol to assess metabolic activity of Pseudomonas aeruginosa by measuring heat flow using isothermal calorimetry. STAR. Protoc. 4, 102269. https://doi.org/10.1016/j.xpro.2023.102269 (2023).
-
McGuffie, M. J. & Barrick, J. E. PLannotate: Engineered plasmid annotation. Nucleic Acids Res.49, W516–W522. https://doi.org/10.1093/nar/gkab374 (2021).
-
Prjibelski, A., Antipov, D., Meleshko, D., Lapidus, A. & Korobeynikov, A. Using spades de Novo assembler. Curr. Protoc. Bioinf. 70, e102. https://doi.org/10.1002/cpbi.102 (2020).
-
Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics30, 2068–2069. https://doi.org/10.1093/bioinformatics/btu153 (2014).
-
Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. Preprint At. https://doi.org/10.48550/arXiv.1303.3997 (2013).
-
Mukherjee, S. & Sengupta, S. Riboswitch scanner: An efficient pHMM-based web-server to detect riboswitches in genomic sequences. Bioinformatics32, 776–778. https://doi.org/10.1093/bioinformatics/btv640 (2016).
-
Singh, P., Bandyopadhyay, P., Bhattacharya, S., Krishnamachari, A. & Sengupta, S. Riboswitch detection using profile hidden Markov models. BMC Bioinformatics10, 325. https://doi.org/10.1186/1471-2105-10-325 (2009).
-
Jore, J. P. M., van Luijk, N., Luiten, R. G. M., van der Werf, M. J. & Pouwels, P. H. Efficient transformation system for Propionibacterium freudenreichii based on a novel vector. Appl. Environ. Microbiol.67, 499–503. https://doi.org/10.1128/AEM.67.2.499-503.2001 (2001).
-
Gutiérrez-Preciado, A. et al. Extensive identification of bacterial riboflavin transporters and their distribution across bacterial species. PLoS One. 10, e0126124. https://doi.org/10.1371/journal.pone.0126124 (2015).
-
Mohammed, Y., Lee, B., Kang, Z. & Du, G. Development of a two-step cultivation strategy for the production of vitamin B12 by Bacillus megaterium. Microb. Cell. Fact.13, 102. https://doi.org/10.1186/s12934-014-0102-7 (2014).
-
Bernhardt, C., Zhu, X., Schütz, D., Fischer, M. & Bisping, B. Cobalamin is produced by Acetobacter pasteurianus dsm 3509. Appl. Microbiol. Biotechnol.103, 3875–3885. https://doi.org/10.1007/s00253-019-09704-3 (2019).
Acknowledgements
We thank Miikka Olin for technical assistance during the UHPLC and LC-MS/MS analyses.
Funding
Ruoxi Zhang holds a China Scholarship Council Studentship with the University of Helsinki. Other funding for this study was from University of Helsinki and Novo Nordisk Foundation (project number NNF20OC0065096). Open access funded by Helsinki University Library.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, R., Sha, Y., Chamlagain, B. et al. Oxygen determines the requirement for cobalamin but not riboflavin in the growth of Propionibacterium freudenreichii. Sci Rep 15, 27679 (2025). https://doi.org/10.1038/s41598-025-12983-8
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-12983-8