
Introduction
Wheat (Triticum aestivum L. AABBDD), one of the most important staple food crops, provides 20% of the global energy and dietary protein1. Nevertheless, the emergence of pests and diseases poses a significant threat to wheat production2. Powdery mildew caused by fungal pathogen Blumeria graminis f. sp. tritici (Bgt) is one of the most damaging wheat diseases worldwide3. Deployment of disease-resistance genes is one of the most effective strategies to mitigate yield loss caused by crop diseases. However, genetic resistance is often overcome due to the rapid evolution of pathogen virulence. Thus, the continuous arms race between plants and pathogens necessitates the continuous identification and deployment of resistance genes to enhance wheat disease resistance.
To date, over a hundred powdery mildew resistance genes/alleles in wheat and its wild relatives have been identified, with 20 of these resistance genes having been successfully cloned. More than half of the cloned genes encode nucleotide-binding leucine-rich repeat (NLR) receptor proteins, including Pm1a4, Pm25, Pm36, Pm5e7, Pm88, Pm129, Pm1710, Pm2111,12, Pm4113, Pm5514, Pm6015, Pm6916, and Pm6Sl17. Pm4 is a chimeric protein of a serine/threonine kinase and multiple C2 domains18, and Pm13 is a mixed lineage kinase domain-like protein19. Pm24 (WTK3)20, WTK421, Pm36 (WTK7-TM)22, and Pm57 (WTK6b-vWA)23 are tandem kinase proteins recently found in the Triticeae family. The partial and adult plant resistance (APR) genes Pm38/Yr18/Lr34/Sr5724 and Pm46/Yr46/Lr67/Sr5525 encode an ABC transporter and a hexose transporter, respectively. Although numerous powdery mildew resistance genes have been identified, the rapid mutation of the pathogenic fungus renders existing resistance genes vulnerable to breakdown, highlighting the urgent need to clone more resistance genes and apply them to maintain effective disease control.
Crop wild species represent a vital reservoir for modern genetic improvement. Wild emmer wheat (T. turgidum ssp. dicoccoides, WEW, AABB), as the wild progenitor of both modern cultivated tetraploid and hexaploid wheat, has a wide range of morphological and genetic variation, especially rich in disease resistance genes26. Hence, WEW is regarded as an ideal source for the mining of excellent disease resistance genes. To date, more than 20 powdery mildew resistance genes/alleles have been documented in WEW, but only Pm3622, Pm4113, Pm60 alleles MlWE1827/MlIW17228/TdPm6029, and Pm6916 have been characterized. Consequently, the cloning of powdery mildew resistance genes from wild emmer wheat has great potential for wheat breeding programs.
Plants have evolved sophisticated immune systems to defend against diverse pathogens, primarily mediated by intracellular nucleotide-binding leucine-rich repeat (NLR) receptors30. These NLRs function as singletons, pairs, or within complex signaling networks31. While singleton NLRs can independently recognize pathogen-derived effectors and activate immune responses, many NLRs act cooperatively32,33,34. NLR pairs often exhibit conserved genomic arrangements, most notably head-to-head orientation, as seen in Arabidopsis RPS4/RRS135, rice RGA4/RGA536, and Pik-1/Pik-237. Recent studies in wheat have revealed similar arrangements, with NLR gene pairs such as RXL/Pm5e38, TdCNL1/TdCNL539, and TdNLR1/TdNLR240,41 functioning jointly to confer resistance. Additionally, tandem kinases such as WTK3 (Pm24), Rwt4, and Sr62 have been shown to operate together with associated NLRs as “sensor-executor” pairs, mediating immune responses against fungal pathogens42,43.
In this study, we clone a powdery mildew resistance locus, PmWR183, which is controlled by a pair of NLR proteins originating from wild emmer wheat. Stable genetic transformation and CRISPR/Cas9-mediated knockout experiments show that the PmWR183–NLR1/PmWR183–NLR2 pair coordinately confers resistance to powdery mildew in common wheat. Protein interaction assays demonstrate that PmWR183-NLR1 and PmWR183-NLR2 engage in both homomeric and heteromeric interactions, suggesting their cooperative role in immune signaling pathways. In addition, we investigate the natural variation of the PmWR183 locus in diverse worldwide wheat germplasm collections, including tetraploid and hexaploid wheat.
Results
PmWR183 is a developmental stage-dependent resistance to powdery mildew in hexaploid wheat
Our previous study has identified a powdery mildew resistance gene, Pm64, originating from wild emmer wheat and mapped on the chromosome arm 2BL, from the WEW-hexaploid wheat introgression line WE3544. The homozygous line WR183, derived from the F5 progeny of a cross between WE35 and the common wheat line S2199, was susceptible to powdery mildew at the two-leaf stage, exhibited partial resistance at the three- and four-leaf stages, and showed high resistance at the adult plant stage under both greenhouse and field conditions (Supplementary Fig. 1a–c). Molecular marker analysis revealed the absence of the Pm64 resistance gene in line WR183 (Supplementary Fig. 1d). Given that WE35 is highly resistant and S2199 is highly susceptible to powdery mildew at both seedling and adult plant stages (Supplementary Fig. 1b, c), we deduced that the adult plant stage resistance to powdery mildew in WR183 is inherited from the resistant parent WE35.
To investigate the genetic basis of APR to powdery mildew in WR183, we crossed WR183 with Fielder, a common wheat cultivar highly susceptible to powdery mildew at both seedling and adult plant stages (Fig. 1a). The F1 hybrids are highly resistant to Bgt isolate E20 at the adult plant stage, indicating the dominant nature of the powdery mildew resistance in WR183 (Supplementary Data 1). The F2 plants segregated as 338 resistant: 120 susceptible, as expected of a 3:1 ratio (χ2 = 0.35, P > 0.05). The F3 families segregated as 115 homozygous resistant: 223 segregating: 120 homozygous susceptible, fitting a 1:2:1 segregation ratio (χ2 = 0.42, P > 0.05) (Supplementary Data 1). These results suggest that the powdery mildew resistance at the adult plant stage in WR183 is controlled by a single dominant gene, provisionally designated PmWR183.

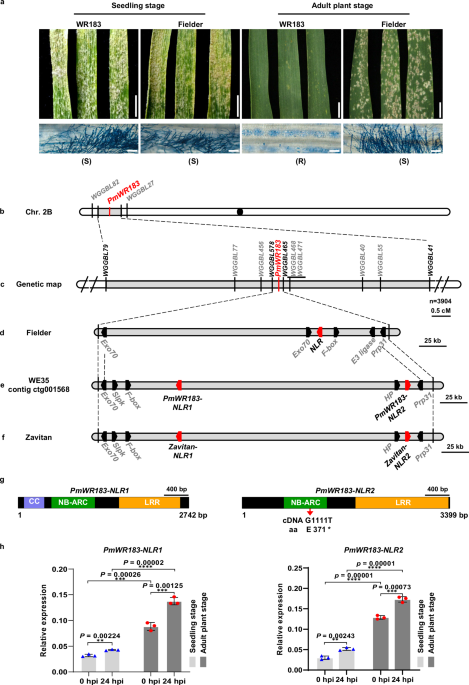
a Seedling and adult plant responses of parental lines WR183 and Fielder to Bgt isolate E20. Plant leaves were detached and photographed at 10 days post-inoculation (dpi) of the two-leaf (seedling) and 20 dpi of the jointing (adult) stage. Scale bar, 0.5 cm. R and S under pictures indicate the resistance and susceptibility of the plants to Bgt, respectively. Fungal structures of Bgt isolate E20 at 5 dpi (seedling stage) or 10 dpi (adult stage) were stained by Coomassie Brilliant Blue, respectively. Scale bar, 100 μm. b, c Genetic map of PmWR183 on the chromosome arm 2BS. d–f Physical map of PmWR183 locus on Fielder, contig ctg001568 of WE35, and Zavitan (WEW v1.0) reference genome, respectively. The red boxes indicate NLR genes. g Genomic structure and variations of PmWR183–NLR1 and PmWR183–NLR2 genes in WR183 and Zavitan. The nucleotide and corresponding amino acid change in PmWR183-NLR2 are shown as indicated. h The relative expression levels of PmWR183–NLR1 and PmWR183–NLR2 in WR183 at both seedling and adult plant stages under mock (uninoculated) and Bgt isolate E20 inoculation. Leaves from WR183 were collected 24 h after Bgt inoculation at both the two-leaf (seedling) and jointing (adult) stages. TaActin was used as the endogenous control. Data are means ± SEM (n = 3 biologically independent samples). Data were analyzed by two-tailed Student’s t-test (**p < 0.01, ***p < 0.001, ****p < 0.0001). Source data are provided as a Source data file.
Fine mapping of PmWR183
Bulked segregant RNA sequencing (BSR-Seq) was utilized to map PmWR183. A pair of RNA bulks generated by pooling homozygous resistant and homozygous susceptible F3 families derived from WR183 × Fielder was subjected to RNA sequencing (RNA-Seq) using the Illumina HiSeq 4000 platform. A total of 34,896,582 and 32,739,558 clean reads were obtained from the resistant pool and the susceptible pool, respectively. The clean reads were aligned to the Chinese Spring reference genome IWGSC RefSeq v2.145, and 299,731 variants were identified from the uniquely mapped reads, with 843 of these variants being trait-associated (P value < 1e-06 and AFD > 0.5). The highest frequency SNPs (283/843) associated with PmWR183 was identified on the end of the short arm of chromosome 2B (2BS) (Supplementary Fig. 2). Based on the BSR-Seq results, four molecular markers, WGGBL82, WGGBL79, WGGBL41 and WGGBL27 were designed according to the Fielder reference genome46 and showed linkage to PmWR183 after genotyping the 458 F2 plants from cross WR183 × Fielder (Supplementary Data 2). PmWR183 was mapped in an 8.52 cM genetic interval of chromosome 2BS flanked by markers WGGBL79 and WGGBL41 (Fig. 1b).
To fine mapping PmWR183, we screened a population of 3904 F2 plants derived from the cross between WR183 and Fielder using PmWR183 flanking markers WGGBL79 and WGGBL41 and identified 405 recombinant plants between these two markers. Eight molecular markers between markers WGGBL79 and WGGBL41 were further developed according to the Fielder reference genome to construct a high-density linkage map of PmWR183 (Fig. 1c). Finally, PmWR183 was mapped to a 0.37 cM genetic interval delimited by the closest markers WGGBL578 and WGGBL456, which corresponds to a 260.1 kb genomic region of the Fielder reference genome46. This region contained six genes, including two exocyst component of 70-kDa (TraesFLD2B01G044400, Exo70; TraesFLD2B01G044500, Exo70), a nucleotide-binding and leucine-rich repeat protein (TraesFLD2B01G044600, NLR), an F-box protein (TraesFLD2B01G044700, F–box), an E3 ubiquitin-protein ligase (TraesFLD2B01G044800, E3 ligase), and a U4/U6 small nuclear ribonucleoprotein Prp31 (TraesFLD2B01G044900, Prp31) (Fig. 1d). We designed specific primers based on Fielder reference genome to amplify the six candidate genes within the PmWR183 interval in WR183 and Fielder. The results revealed that all six candidate genes produced amplification bands in Fielder, while only the Prp31 gene showed amplification bands in WR183, with the remaining five genes showing no amplification (Supplementary Fig. 3). This suggests that there may be genomic structural variations at the PmWR183 locus between WR183 and Fielder.
Long-read genome sequencing of line WE35
To facilitate the cloning of PmWR183, we generated a de novo genome assembly of line WE3544 carrying Pm64 and PmWR183 using the PacBio SMRT long-read sequencing approach. High-fidelity (HiFi) reads were obtained, achieving an approximate 9-fold genome coverage of hexaploid wheat (Supplementary Data 3). After the assembly of the sequences, we obtained a total of 6606 contigs, with the N50 contig length being 4.4 Mb, the N90 contig length 1.3 Mb, and the longest contig spanned 27.3 Mb in length (Supplementary Data 3).
A 2.5 Mb contig, designated ctg001568, was identified by the closest flanking markers WGGBL578 and WGGBL456. This contig was found to encompass the entire 295.6 kb physical interval of PmWR183. Within this region, seven genes were annotated: an exocyst component of 70-kDa (Exo70), an S-locus lectin protein kinase family member (Slpk), an F-box gene (F–box), two nucleotide-binding site (NBS) leucine-rich repeat proteins (PmWR183–NLR1 and PmWR183–NLR2), a hypothetical protein (HP) and a U4/U6 small nuclear ribonucleoprotein Prp31 (Prp31) (Fig. 1e). Genomic sequence comparison and micro-collinearity analysis of the PMWR183 locus revealed significant genomic structural variations between WR183 and both Fielder and Chinese Spring, while exhibiting a perfect collinearity with WEW Zavitan (Fig. 1f and Supplementary Fig. 4). These findings suggest that the genomic interval of PmWR183 is likely derived from wild emmer wheat, in agreement with that WE35 is a WEW-common wheat introgression line. RNA-Seq analysis revealed that, among the seven annotated genes within the genomic interval of PmWR183, only PmWR183–NLR1, PmWR183–NLR2, and Prp31 were expressed in the leaves of WR183 at the adult plant stage after inoculation with Bgt isolate E20 (Supplementary Data 4). In addition, DNA sequence alignment of the three candidate genes, including 1.5 kb upstream and downstream regions of the candidate genes, revealed no sequence variation for PmWR183–NLR1 and Prp31 between WR183 and Zavitan (Supplementary Data 5 and 6). Notably, PmWR183–NLR2 contained a single nonsynonymous SNP (c. G1111T) in Zavitan, resulting in a premature stop codon (Fig. 1g and Supplementary Data 7). This mutation correlated perfectly with Zavitan’s susceptible phenotype after inoculation with Bgt isolate E20 at both the seedling and adult plant stages (Supplementary Fig. 5). Furthermore, PmWR183–NLR2 expression was significantly induced upon Bgt infection in both the seedling and adult plant stages (Fig. 1h). Together, these results suggested that PmWR183–NLR2 is the causal gene responsible for PmWR183 locus-mediated resistance.
Functional validation of PmWR183
To further investigate the role of PmWR183–NLR2 in resistance to Bgt, we employed the dual gRNA CRISPR/Cas9 system to generate knockout mutations of PmWR183–NLR2 in WR183. Firstly, 5′- and 3′- rapid amplification of cDNA ends (RACE) experiments were performed to obtain the full-length cDNA of PmWR183–NLR2 in WR183 (Supplementary Fig. 6). The PmWR183–NLR2 gene is a single-exon gene encoding a 1232 amino acids nucleotide-binding leucine-rich repeat (NLR) protein, which includes typical NBS and leucine-rich repeat (LRR) domains but lacking a typical coiled-coil (CC) domain (Fig. 1g). Two gRNA target sites near the 5′-terminus of the PmWR183–NLR2 were selected to guide Cas9 to simultaneously cleave two sites (Fig. 2a). The recombinant CRISPR/Cas9 construct targeting PmWR183–NLR2 was introduced into WR183 via Agrobacterium-mediated transformation47. We obtained three homozygous independent T1 of PmWR183–NLR2 knock-out (KO) plants (PmWR183–NLR2-KO#1-#3). Specific primer PmWR183–NLR2-2g was designed to amplify the target regions of the PmWR183–NLR2 gene in T1 KO plants, followed by Sanger sequencing (Fig. 2a). When challenged with Bgt isolate E20, three homozygous KO plants exhibited susceptibility to powdery mildew at the adult plant stage, in contrast with WR183 (Fig. 2a). These results show that PmWR183–NLR2 is essential for conferring adult plat resistance to powdery mildew in WR183.

a, b Functional verification PmWR183–NLR1/PmWR183–NLR2 pair using CRISPR-Cas9-induced PmWR183–NLR2-KO and PmWR183–NLR1-KO lines in T1 generations, respectively. Target sites for sgRNAs are marked in red boxes. The sgRNA sequences used for CRISPR-Cas9-mediated mutagenesis are as shown. The blue letters indicate the PAM site. The “−” indicates the number of deleted bases. The WR183, PmWR183–NLR2, and PmWR183–NLR1 knockout mutants were inoculated with Bgt isolate E20 at the adult plant stage. Scale bar, 0.5 cm. Photographs of representative leaves were taken at 20 dpi. c Infection phenotypes of T1 transgenic lines expressing ProUbi:PmWR183–NLR1, ProUbi:PmWR183–NLR2, and ProUbi:PmWR183–NLR1 + ProUbi:PmWR183–NLR2 at the seedling and adult plant stages. Detached leaves were photographed at 10 dpi (seedling stage) and 20 dpi (adult stage) after inoculation with Bgt isolate E20. Scale bar, 0.5 cm. Transgene presence in T1 plants was confirmed by PCR. “+” indicates the positive transgenic individuals; “−” indicates the negative individuals. d The relative expression levels of PmWR183–NLR1 and PmWR183–NLR2 in WR183 and PmWR183–NLR1 + PmWR183–NLR2 overexpressing transgenic plants. The leaves of seedlings and adult plants were harvested at the two-leaf stage and jointing stage, respectively, with TaActin serving as the endogenous control. Data are means ± SEM (n = 3 biologically independent samples). Data were analyzed by two-tailed Student’s t-test (**p < 0.01, ***p < 0.001, ****p < 0.0001). e Resistance phenotypes of T0 transgenic lines expressing ProPmWR183–NLR1:PmWR183–NLR1, ProPmWR183–NLR2:PmWR183–NLR2, and ProPmWR183–NLR1:PmWR183–NLR1 + ProPmWR183–NLR2:PmWR183–NLR2 following inoculation with Bgt isolate E20 at the adult plant stage. Representative leaves were photographed at 20 dpi. Scale bar, 0.5 cm. T0 plants were genotyped by PCR to verify transgene integration. “+” indicates the positive transgenic individuals; “−” indicates the negative individuals. Source data are provided as a Source data file.
To further validate the function of PmWR183–NLR2, we generated an overexpression construct, ProUbi:PmWR183–NLR2, driven by the maize (Zea mays L.) Ubiquitin (Ubi) promoter and introduced it into the susceptible hexaploid wheat cultivar Fielder by Agrobacterium-mediated transformation47. The transformation of the ProUbi:PmWR183–NLR2 construct yielded five positive T0 plants with the confirmed transgene sequence by the specific marker PmWR183–NLR2-DC. The positive T0 individuals of PmWR183–NLR2-OE were advanced to generate the T1 generation (Supplementary Data 8). Surprisingly, all of the positive plants from the five independent PmWR183–NLR2-OE T1 transgenic families, as well as the negative control plants, were highly susceptible to Bgt isolate E20 at the adult plant stage (Supplementary Fig. 7). The introduction of the PmWR183–NLR2 gene alone into Fielder does not confer resistance to powdery mildew at the adult plant stage, suggesting that complete resistance mediated by the PMWR183 locus may require additional genetic component from WR183.
Previous findings revealed that some NLRs cooperate with adjacent, genetically linked NLRs for disease resistance in plants35,36,37,38,48. We observed that the PmWR183–NLR1 gene is located approximately 197 kb from the PmWR183–NLR2 gene within the genetic interval of PmWR183. qRT-PCR results indicated that both PmWR183–NLR1 and PmWR183–NLR2 were highly expressed in the leaves of WR183 at the adult plant stage, in comparison with the seedling stage (Fig. 1h). Therefore, we deduced that PmWR183–NLR1 may also be involved in the resistance to powdery mildew in WR183, potentially functioning cooperatively with PmWR183–NLR2 to establish complete immunity against Bgt infection. The full-length cDNAs of PmWR183–NLR1 were then obtained from line WR183 by the 5′- and 3′- RACE (Supplementary Fig. 6). PmWR183–NLR1 possesses a single exon and encodes a protein comprising 913 amino acids, which includes CC, NBS, and LRR domains (Fig. 1g). We then constructed the recombinant CRISPR/Cas9, which carries two gRNA target sites of PmWR183–NLR1 and transformed it into WR183 using the Agrobacterium-mediated transformation method44. Specific primers, PmWR183–NLR1–2g, were designed to amplify the target regions of PmWR183–NLR1 in the three individual T1 KO plants, followed by Sanger sequencing (Fig. 2b). The three homozygous PmWR183–NLR1-KO plants exhibited susceptibility to Bgt isolate E20 at adult plant stage, similar to the PmWR183–NLR2-KO plants (Fig. 2b). These results indicate that PmWR183–NLR1 also indeed plays a role in the APR to powdery mildew in WR183.
To determine whether PmWR183–NLR1, in conjunction with PmWR183–NLR2, contributes to APR to powdery mildew in WR183, we generated the overexpression construct ProUbi:PmWR183–NLR1 driven by the maize (Zea mays L.) Ubi promoter and co-transformed it along with ProUbi:PmWR183–NLR2 into Fielder by Agrobacterium-mediated transformation. We obtained 15 independent T0 transgene-positive plants by PCR analysis using specific primers of PmWR183–NLR1 and PmWR183–NLR2, including 6 transgene-positive plants for PmWR183–NLR1, 4 transgene-positive plants for PmWR183–NLR2, and 5 transgene-positive plants for both PmWR183–NLR1 and PmWR183–NLR2. These transgene-positive plants were self-pollinated to produce the T1 generation. All of the 6 PmWR183–NLR1 and 4 PmWR183–NLR2 transgene-positive T0 plants and their T1 families are highly susceptible to Bgt isolate E20 at seedling and adult plant stages (Fig. 2c and Supplementary Data 8). In contrast, all transgene-positive plants carrying both PmWR183–NLR1 and PmWR183–NLR2 were highly resistant to Bgt isolate E20 at the adult plant stage (Fig. 2c; Supplementary Fig. 8). In addition, we found that transgene-positive plants carrying both PmWR183–NLR1 and PmWR183–NLR2 showed moderate resistance to Bgt isolate E20 at the seedling stage, while transgene-positive plants carrying only PmWR183–NLR1 or PmWR183–NLR2 were highly susceptible (Fig. 2c and Supplementary Data 8). Furthermore, qRT-PCR analysis revealed that the expression levels of PmWR183–NLR1 and PmWR183–NLR2 in co-transgenic plants were significantly higher than those in WR183 at both the seedling and adult stages (Fig. 2d). This elevated expression likely accounts for the enhanced disease resistance observed in co-transgenic plants at the seedling stage, consistent with the patterns observed in WR183 (Figs. 1h and 2d). In addition, we constructed independent complementation constructs containing the full genomic sequences of PmWR183–NLR1 (7427 bp) and PmWR183–NLR2 (8145 bp), including their native promoter regions and complete coding sequences. The resulting constructs were then individually or simultaneously introduced into the susceptible wheat cultivar Fielder via Agrobacterium-mediated transformation. Subsequently, T0 transgenic plants were evaluated for resistance at the adult plant stage (Fig. 2c and Supplementary Data 8). The disease resistance evaluation results of PmWR183–NLR1– and PmWR183–NLR2-complemented transgenic plants were consistent with those of PmWR183–NLR1– and PmWR183–NLR2-overexpressing transgenic plants: transgene-positive plants carrying both PmWR183–NLR1 and PmWR183–NLR2 were resistant to Bgt isolate E20, whereas plants carrying either PmWR183–NLR1 or PmWR183–NLR2 remained susceptible (Fig. 2e and Supplementary Data 8). Taken together, these results show that the PmWR183–NLR1 and PmWR183–NLR2 pair work together to confer powdery mildew resistance in WR183.
PmWR183-NLR1 and PmWR183-NLR2 exhibit both self and mutual interactions
To investigate the functional and molecular interactions between PmWR183-NLR1 and PmWR183-NLR2, the two proteins were transiently expressed in Nicotiana benthamiana leaves. Expression of PmWR183-NLR1 alone triggered a slight hypersensitive response (HR)-like cell death (Fig. 3a). In contrast, PmWR183-NLR2 expression did not induce any visible response, suggesting that PmWR183-NLR2 lacks cell death-inducing activity (Fig. 3a). Furthermore, co-expression of PmWR183-NLR1 and PmWR183-NLR2 did not significantly enhance or suppress PmWR183-NLR1-triggered cell death (Fig. 3a). Previous studies have shown that substitutions affecting the acidic inner lining of the funnel formed by α1-helices can abolish cell death in planta49,50,51. To further investigate the interaction between PmWR183-NLR1 and PmWR183-NLR2 while minimizing the confounding effects of cell death, we introduced L to E mutation at the α1-helix of PmWR183-NLR1 (PmWR183-NLR1L7E/L8E) (Supplementary Fig. 9). These mutations effectively suppressed the HR-like cell death phenotype without affecting PmWR183-NLR1 protein expression (Supplementary Fig. 10). To investigate the physical interactions between PmWR183-NLR1 and PmWR183-NLR2 in vivo, the PmWR183-NLR1L7E/L8E mutant was used in co-immunoprecipitation (Co-IP) and split firefly luciferase complementation (SFLC) assays. Using WTN1Δ23 (WTN124-1038) as a negative control43, we found that PmWR183-NLR1 and PmWR183-NLR2 are capable of both self-association and mutual interaction (Fig. 3b, c). Western blot analysis confirmed proper expression of all fusion proteins (Fig. 3). These findings provide strong evidence for an interaction between PmWR183-NLR1 and PmWR183-NLR2, highlighting their cooperative role in conferring resistance to powdery mildew in wheat.

a PmWR183-NLR1-HA and PmWR183-NLR2-MYC were transiently expressed in N. benthamiana, either individually or in combination. Cell death phenotypes were recorded at 3 days post-infiltration. The number of leaves tested and hypersensitive response (HR) occurrences are indicated in parentheses. Data are representative of three independent experiments, with similar results. b Co-IP analysis of PmWR183-NLR1L7E/L8E and PmWR183-NLR2 interactions when expressed individually or co-expressed in N. benthamiana. WTN1△23 (WTN124-1038) served as a negative control. Data are representative of three independent experiments, with similar results. c Luciferase complementation assay showing the interaction between PmWR183-NLR1L7E/L8E and PmWR183-NLR2 when expressed individually or together in N. benthamiana. WTN1Δ23 was used as a negative control. Data are presented as mean ± SEM (n = 5 independent biological replicates). Statistical significance was determined by one-way ANOVA followed by Tukey’s multiple comparisons test (p < 0.05). Black, blue, and purple asterisks indicate WTN1Δ23-HA-nLUC, cLUC-PmWR183-NLR2-HA, and cLUC-PmWR183-NLR1L7E/L8E-HA, respectively. Source data are provided as a Source data file.
Origins analysis of PmWR183–NLR1 and PmWR183–NLR2 in ancestral species
Bread wheat (Triticum aestivum) is an allohexaploid species (2n = 6x = 42, AABBDD genome) that evolved through the hybridization of three distinct wild grass species. The wild emmer wheat (Triticum turgidum subsp. dicoccoides, 2n = 4x = 28) emerged from an initial hybridization event between Triticum urartu (2n = 2x = 14) and an unknown B genome donor closely related to the goatgrass Aegilops speltoides. Therefore, we utilized the sequenced Aegilops species to trace the origins of PmWR183–NLR1 and PmWR183–NLR2, including Ae. speltoides, Ae. sharonensis, Ae. longissima, Ae. bicornis and Ae. searsii52,53. The protein and genomic sequences of PmWR183–NLR1 and PmWR183–NLR2 were used as queries to identify their orthologs and homologs. Subsequently, we identified PmWR183–NLR1 orthologs exclusively in Ae. bicornis (TB01) (Nucleotide BLAST: coverage = 95%, percent identity = 96%) and Ae. sharonensis (TH02) (Nucleotide BLAST: coverage = 94%, percent identity = 96%), while PmWR183–NLR1 was absent in other Aegilops genomes on the chromosome 2S, as determined by both nucleotide and protein BLAST (coverage > 90%, percent identity > 60%) (Fig. 4a–d). Interestingly, no orthologs of PmWR183–NLR2 were found in any sequenced Aegilops species, including the two species that contain PmWR183–NLR1. Moreover, three PmWR183–NLR2 homologs were identified exclusively on the 2S chromosmme of Ae. sharonensis (AS 1644), positioned near the expected location of PmWR183–NLR2, and each has a corresponding ortholog in wild emmer wheat (Fig. 4d, e). We hypothesize that the genomic region of the PmWR183–NLR2 locus may experience an NLR duplication. In addition, a protein encoding the exocyst component of 70-kDa (Exo70) in this duplication event, similar to that observed in the NLR genes (Fig. 4d, e). Therefore, we hypothesize that PmWR183–NLR2 may have arisen from a duplication event in a specific Aegilops species, which was subsequently followed by hybridization and recombination with another Aegilops species containing the PmWR183–NLR1 gene, leading to the formation of an NLR gene pair.

Genes are shown as colored boxes along the physical maps of each species. The micro-collinearity relationship showed that some neighboring genes of PmWR183–NLR1/PmWR183–NLR2 were conserved across Aegilops species genomes. PmWR183-NLR1 alleles are represented by blue boxes, PmWR183–NLR2 alleles by red boxes, genes encoding other NLR receptor proteins by pink boxes, genes encoding an exocyst component of 70-kDa by yellow boxes, and all other genes by black boxes. The genes included in the displayed genomic regions are as follows: a Ae. longissima (AEG-6782-2: AE.LONG.r1.2SG0076750-AE.LONG.r1.2SG0077050), b Ae. sharonensis (TH02: Ae.sharonensis.TH02.2S01G0038000 –Ae.sharonensis.TH02.2S01G0038600), c Ae. bicornis (TB01: Ae.bicornis.TB01.2S01G0040400 –Ae.bicornis.TB01.2S01G0041800), d T. turgidum ssp. dicoccoides (Zavitan V1.0: TRIDC2BG003690 –TRIDC2BG004030), and e Ae. sharonensis (AS 1644: AE.SHARON.r1.2SG0084690 –AE.SHARON.r1.2SG0085000), respectively. Orthologous genes are connected by solid lines, while homologous genes are represented with dashed lines.
Haplotype analysis and geographic distribution of PmWR183–NLR1 and PmWR183–NLR2
To evaluate the natural variation of PmWR183–NLR1 and PmWR183–NLR2 in wheat, two sequence tagged site (STS) markers, PmWR183–NLR1-FM for PmWR183–NLR1 and PmWR183–NLR2-FM for PmWR183–NLR2, were developed to screen the wheat germplasm collected from geographically diverse regions worldwide (Supplementary Data 2), which comprises 588 tetraploid wheat accessions and 1292 hexaploid wheat accessions, respectively (Supplementary Data 9). Consequently, PmWR183–NLR1 and PmWR183–NLR2 were detected in 57 wheat accessions and were always present together (Supplementary Data 9). Haplotype analysis was subsequently performed on these 57 accessions by amplifying and sequencing the coding sequences of both PmWR183–NLR1 and PmWR183–NLR2.
For PmWR183–NLR1, we identified five haplotypes (PmWR183–NLR1Hap1 to PmWR183–NLR1Hap5) (Fig. 5a). Notably, PmWR183–NLR1Hap2 and PmWR183–NLR1Hap3 each differ from PmWR183–NLR1Hap1 by only a single amino acid variant. The three-dimensional (3D) model of the PmWR183-NLR1 protein predicted by AlphaFold3 revealed distinct CC, NBS, and LRR domains, with haplotypic sequence variations predominantly localized in the NBS and LRR domains (Fig. 5b). PmWR183–NLR2 exhibits seven haplotypes (PmWR183–NLR2Hap1 to PmWR183–NLR2Hap7) (Fig. 5c), showing greater sequence diversity compared to PmWR183–NLR1, with more than 50% of these variations localized within the LRR domain (Fig. 5d). Additionally, the 3D model indicated that a sequence variation resulting in premature translation termination causes PmWR183-NLR2Hap3 (i.e., Zavitan) to encode only a truncated NBS domain (Fig. 5e).

a Amino acid sequence variations among different haplotypes of PmWR183-NLR1. Compared with PmWR183-NLR1, the haplotypes exhibiting amino acid sequence variations are highlighted in red. b Protein structure of PmWR183-NLR1 predicted by AlphaFold3. The CC, NB-ARC, and LRR domains of PmWR183-NLR1 are highlighted in purple, green, and orange, respectively. c Amino acid sequence variation among different haplotypes of PmWR183-NLR2. In comparison to PmWR183-NLR2, haplotypes displaying amino acid sequence variations are marked with red. “*” indicates translation termination. d, e Protein structure of PmWR183-NLR2 predicted by AlphaFold3. The NB-ARC and LRR domains of PmWR183-NLR2 are highlighted in green and orange, respectively. f Geographic distribution of PmWR183–NLR1 and PmWR183–NLR2 in wild emmer wheat, durum, and common wheat accessions. The haplotype combinations of PmWR183–NLR1 and PmWR183–NLR2 are labeled with different symbols. The T. turgidum ssp. dicoccoides, T. turgidum ssp. durum, T. turgidum ssp. dicoccum, and T. aestivum are highlighted with red, yellow, blue, and purple, respectively. The right figure is a 20× enlargement of the red box region in the left image. The geographical locations were plotted on a map generated using rworldmap (v1.3-6) in R (v4.4.2). NLR1 and NLR2 in the bottom box represent PmWR183-NLR1 and PmWR183-NLR2, respectively.
Based on sequence variations in PmWR183–NLR1 and PmWR183–NLR2, the different combinations of these two genes were classified into 10 distinct haplotypes (PmWR183–NLR1Hap1/PmWR183–NLR2Hap1, PmWR183–NLR1Hap1/PmWR183–NLR2Hap2, PmWR183–NLR1Hap1/PmWR183–NLR2Hap3, PmWR183–NLR1Hap1/PmWR183–NLR2Hap6, PmWR183–NLR1Hap1/PmWR183–NLR2Hap7, PmWR183–NLR1Hap2/PmWR183–NLR2Hap1, PmWR183–NLR1Hap2/PmWR183–NLR2Hap3, PmWR183–NLR1Hap3/PmWR183–NLR2Hap1, PmWR183–NLR1Hap4/PmWR183–NLR2Hap5 and PmWR183–NLR1Hap5/PmWR183–NLR2Hap4) (Supplementary Data 9 and 10). The PmWR183–NLR1 and PmWR183–NLR2 isolated from WR183 exhibited the PmWR183–NLR1Hap1/PmWR183–NLR2Hap1 combination.
The 45 WEW accessions contained all the identified ten distinct haplotype combinations except PmWR183–NLR1Hap1/PmWR183–NLR2Hap2, which was detected only in six durum wheat accessions. For cultivated emmer wheat, four accessions were identified carrying the PmWR183–NLR1Hap4/PmWR183–NLR2Hap5 haplotype combination. In the 1,292 common wheat accessions analyzed, only one accession was found to carry the haplotype PmWR183–NLR1Hap4/PmWR183–NLR2Hap5 (Supplementary Data 9 and 10), suggesting the PmWR183 and its alleles have been rarely utilized in modern wheat breeding.
Geographic origins information was available for 42 of the 57 wheat accessions, which include nine distinct haplotype combinations (Supplementary Data 9). WEW accessions containing PmWR183 alleles were predominantly identified in the southern Levant WEW populations (i.e., Israel), with five accessions found in northern Levant populations (Türkiye) (Fig. 5f). Durum wheat accessions carrying PmWR183 alleles were distributed across Bulgaria, Greece, Türkiye, Iraq, and Azerbaijan (Fig. 5f). Cultivated emmer wheat accessions appear to have spread farther than durum wheat, with distributions extending to regions such as Iran, Russia, and Spain (Fig. 5f). Interestingly, only one common wheat accession, Barbilla, containing the PmWR183 alleles, was collected from Spain. The functional characterization of those different PmWR183 haplotypes in the future could provide more allelic variants and options in future wheat powdery mildew resistance breeding programs.
Discussion
WEW is considered a valuable gene pool for disease-resistance genes that can contribute to wheat resistance breeding. Several disease resistance genes have been cloned from WEW, including powdery mildew resistance genes Pm2639, Pm3622, Pm4113, Pm60 alleles MlWE1827/MlIW17228/TdPm6029, and Pm6916; and stripe rust resistance genes Yr1554, Yr3655, and Yr84/YrTD12140,41. In this work, we cloned a powdery mildew resistance locus PMWR183, originating from WEW, through the combination of map-based cloning, PacBio SMRT long-read genome sequencing, stable transformation, and CRISPR/Cas9-mediated genome editing. Among the reported disease resistance genes derived from WEW, Pm41, Pm60 alleles MlWE18/MlIW172/TdPm60, and Pm69 were found to be singleton NLR proteins; Yr15 (WTK1) and Pm36 (WTK7-TM) are tandem kinase resistance proteins; Yr36 encodes a serine/threonine kinase integrated steroidogenic acute regulatory protein-related lipid-transfer (START) domain. All these disease-resistant genes cloned from wild emmer wheat are controlled by single gene loci, with the exception of PM2639 and YR84/YRTD12140,41, which encode NLR pairs. Recently, the PM5e locus from the Chinese wheat cultivar Fuzhuang 30 was also characterized as paired NLR RXL/Pm5e38. In this paper, we show that the PMWR183 locus is controlled by a pair of NLR genes, PmWR183–NLR1/PmWR183–NLR2. Screening a diverse range of global tetraploid and hexaploid wheats using PmWR183–NLR1/PmWR183–NLR2 diagnostic markers revealed that PMWR183 originated from WEW and is rarely found in cultivated emmer, durum, and common wheat, suggesting that it has not been fully utilized in modern wheat breeding. Sequence analysis showed that the PmWR183–NLR2 gene exhibits extremely high amino acid diversity in wheat populations, especially in the LRR region. This diversity may be maintained through continuous mutual selection between host and pathogen. Our research underscores the importance of WEW as an excellent gene reservoir, highlighting the potential of its genetic diversity for further exploration and application in wheat breeding.
The majority of disease resistance genes in plants encode NLR proteins, responsible for detecting pathogen invasion and initiating defense responses56. Typical NLRs are characterized by a central NBS domain, a C-terminal leucine-rich repeat (LRR) domain, and a notably variable N-terminal domain, which can be either a Toll/interleukin-1 receptor (TIR) or a coiled-coil (CC) domain30. This distinction divides plant NLRs into two major types: TNL (TIR-NLR) and CNL (CC-NLR). Some individual NLRs are capable of both detecting the presence of pathogen effectors and initiating the subsequent immune response within the plant cell. Wheat powdery mildew resistance gene Pm36, stem rust resistance genes Sr3549, Sr5057, and Arabidopsis RPP1358 encode NLR proteins, which induce resistance upon recognition of isolate-specific avirulence (AVR) effectors from pathogens. Recent findings demonstrate that NLR-mediated immunity is more intricate than previously understood, with NLRs requiring collaboration with other NLRs, either genetically linked or unlinked, to initiate an immune response. Arabidopsis NLR pair, RRS1/RPS435, CHS3/CSA159 pairs, which belong to TNLs, cooperate genetically and molecularly in resistance to different pathogens. Examples of CNL pairs include RGA4/RGA536, Pik-1/Pik-260, and Pi1/Pii248 in rice and RXL/Pm5e38, TdCNL1/TdCNL539 and TdNLR1/TdNLR240,41 in wheat. Additionally, the pioneering discovery that tandem kinases such as WTK3 (Pm24), Rwt4, and Sr62 function with partner NLRs as pairs has uncovered immune modules of fungal pathogen resistance in planta42,43. In our current study, cloning and characterization of the PMWR183 locus indicated that it also consisted of two adjacent NBS-LRR genes, PmWR183–NLR1 and PmWR183-NLR2, providing additional document for cloned powdery mildew resistance genes in wheat.
Paired NLRs are often found in close genetic proximity on chromosomes, one of which functions as a sensor receptor and the other as a helper NLR. Sensor NLRs, which often possess a noncanonical domain known as the integrated domain (ID), can recognize pathogen effectors either directly or indirectly by detecting modifications to host target proteins, but they require helper NLRs to initiate downstream immune signaling61,62,63. Rice RGA5 contains a heavy metal-associated (HMA) domain at the C-terminus after the LRR domain, which interacts with Magnaporthe oryzae effectors Avr-Pia and AVR1-CO3936. Similar to RGA5, Pik-1 also contains an HMA domain, but located between CC and NB-ARC domains64. Arabidopsis RRS1 protein features an integrated WRKY domain near its C-terminus, which serves as a target for bacterial pathogen effectors such as AvrRps4 and PopP265. Interestingly, our research revealed that neither PmWR183-NLR1 nor PmWR183-NLR2 harbors an ID. However, similar to other paired NLRs, they exhibit direct physical interactions35,36,38. Sequence analysis of accessions containing both PmWR183-NLR1 and PmWR183-NLR2 revealed that PmWR183-NLR1 exhibits relatively conserved sequences, while PmWR183-NLR2 displays greater sequence variation. Notably, the variations in PmWR183-NLR2 across different materials were predominantly localized in the LRR domain, which plays a crucial role in pathogen effector recognition in NLR proteins. Even in the absence of Bgt effector AvrPmWR183, PmWR183-NLR1 expression alone in N. benthamiana induced a mild HR, whereas PmWR183-NLR2 failed to trigger cell death in tobacco leaves. Furthermore, mutational analysis revealed that PmWR183-NLR1’s α1-helix constitutes an essential element for HR, as single-residue substitutions (L7E/L8E) completely disabled cell death activity in our assays (Supplementary Figs. 9 and 10), which was similar to the NLR proteins Sr35 and ZAR149,50. Based on the experimental data presented above, PmWR183-NLR2 likely serves as the sensor, while the more conserved PmWR183-NLR1 appears to function as a helper NLR in mediating immune responses. However, definitive validation of this hypothesis requires cloning of the AvrPmWR183 effector protein in the future.
APR refers to the ability of disease resistance genes to provide protection against pathogens at the adult plant stage, but they are ineffective in seedlings. The wheat genes Lr34/Yr18/Sr57/Pm3824 and Lr67/Yr46/Sr55/Pm4625 encode an ABC transporter and a hexose transporter, respectively, which provide resistance to multiple fungal pathogens, including stripe rust, leaf rust, stem rust, and powdery mildew, specifically at the adult stage. The wheat Yr36 gene is a high-temperature APR gene that effectively confers resistance to a wide range of Puccinia striiformis f. sp. tritici (Pst) races55. In rice, the Xa21 gene confers a broad and persistent resistance against bacterial blight in adult plants, while it offers almost no resistance at the seedling stage66. Maize APR gene Hm1 encodes an NADPH-dependent reductase that confers robust resistance to northern leaf spot at maturity67. Our findings demonstrate that the PmWR183 confers strong resistance to powdery mildew at the adult plant stage in WR183, but not at the two-leaf stage, which was further validated by resistance tests using multiple Bgt isolates (Supplementary Data 11). Interestingly, resistance at the seedling stage was observed in tetraploid wheat accessions carrying PmWR183 or its alleles (Supplementary Fig. 11), whereas in WR183, resistance gradually developed with plant growth and became pronounced at the adult stage (Supplementary Fig. 1). This developmental stage-dependent resistance is in accordance with gene expression analysis, which demonstrated that transcript levels of PmWR183-NLR1 and PmWR183-NLR2 gradually accumulated as the plant growth (Supplementary Fig. 12a). The markedly reduced expression of PmWR183-NLR2 in hexaploid WR183 compared to tetraploid wheat (IW60, PmWR183–NLR1Hap1/PmWR183–NLR2Hap1) suggests that it may be transcriptionally repressed (Supplementary Fig. 12b). As PmWR183-NLR2 likely functions as a sensor NLR, its insufficient expression may represent a critical bottleneck for seedling-stage resistance in WR183. We hypothesize that a suppressor locus targeting PmWR183–NLR2 in the genetic background of hexaploid WR183 modulates the expression or activity of PmWR183-mediated resistance during the early developmental stage. Notably, co-overexpression of PmWR183–NLR1 and PmWR183–NLR2 in the susceptible cultivar Fielder appeared to relieve this suppression, thereby restoring seedling-stage resistance to powdery mildew, similar to the resistance conferred by the overexpression of some R genes68,69,70. These results support the functional roles of PmWR183–NLR1 and PmWR183–NLR2 in PmWR183-mediated resistance and suggest that modulating their expression may help bypass stage-specific suppression. This strategy could contribute to the development of cultivars with durable resistance across developmental stages, enhancing disease control and yield stability.
Methods
Plant materials
Common wheat line WE35 is developed by backcrossing a common wheat line 87-1, which is highly susceptible to Bgt, with wild emmer accession G-573-1 (pedigree: 87-1*4/G-573-1)44. WE35 displays seedling (two-leaf stage) and adult stage (grain-filling stage) resistance to Bgt, in contrast to the common wheat line S2199, which is highly susceptible at all plant stages. The homozygous line WR183, an F5 progeny from the cross of WE35 and S2199, exhibits susceptibility to Bgt at the two-leaf stage but shows resistance at the adult plant stage. F1 hybrids, F2 segregating population, and F2-derived F3 families from the cross of WR183 with common wheat cultivar Fielder, which is highly susceptible to Bgt at all plant stages, for genetic analysis and molecular mapping. In addition, wheat collections for the haplotype analysis of PmWR183 included 588 tetraploid wheat accessions and 1292 common wheat accessions, as reported in a previous study22. Plants were grown in a greenhouse with a temperature range of 18–24 °C, humidity of 60–70%, and a 16-/8-h light/dark cycle at the Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China. Field trials were carried out to assess adult plant powdery mildew resistance with mixed Bgt isolates at the Gaoyi Experimental Station, Shijiazhuang, Hebei.
Powdery mildew evaluations
The lines WE35, S2199, WR183, Fielder, and genetic populations from WR183 × Fielder, transgenic plants were challenged with Bgt isolate E20 and evaluated for powdery mildew resistance at the seedling or adult stage. Seedling response to Bgt E20 was evaluated under controlled greenhouse conditions13,22. Inoculation was performed at the one-leaf stage, and disease response was assessed at the two-leaf stage. APR was evaluated at the grain-filling stage under both field and greenhouse environments, with inoculations initiated at the jointing stage. Infection types (IT) were scored on a 0–4 infection type (IT) scale 15–20 days post-inoculation when susceptible Fielder control plants were sporulating profusely. Plants with infection types (ITs) of 0 (immune), 0 (hypersensitive reaction), 1 (highly resistant), and 2 (moderately resistant) were categorized as resistant, while those with ITs of 3 (moderately susceptible) and 4 (highly susceptible) were classified as susceptible.
To visualize the fungal structures, detached leaves from WR183 and Fielder inoculated with Bgt isolate E20 with 5 days post-inoculation (dpi) at the seedling stage and 10 dpi at the adult stage were bleached in absolute ethanol and stained with a 0.6% (w/v) Coomassie Brilliant Blue solution for 10 s and then washed with water. Bright-field images were obtained using an Olympus BX53F fluorescence microscope (Olympus, Tokyo, Japan).
BSR-Seq
Leaf tissues of 40 homozygous resistant and 40 homozygous susceptible F3 families derived from the WR183 × Fielder cross were separately pooled for RNA preparation. Total RNA was extracted using the RNAsimple Total RNA Kit (Tiangen Biotech Co., Ltd., Beijing, China). RNA-Seq was performed by the Illumina HiSeq 4000 platform (Beijing Novogene Bioinformatics Technology Co., Ltd., Beijing, China). Clean reads were aligned to the wheat reference genome assembly IWGSC RefSeq v2.145 using STAR v2.5.1b71, with a mismatch rate of less than 6%. Only uniquely mapped read pairs were retained for further analysis. Prior to variant calling, read alignments were adjusted to account for PCR duplications. SNPs and InDels were identified using the small variant caller Strelka v2.9.1072 with default parameters. Variants with a sequencing depth of less than 6 were filtered out, and the remaining high-confidence SNPs and InDels were used for bulked segregant analysis. Trait-associated SNPs and InDels variants were identified based on an allele frequency difference (AFD) > 0.5 and a Fisher’s exact test P value < 1e-06. These variants were subsequently used as templates for marker development.
Fine mapping of PmWR183
Specific PCR primers were designed using the flanking sequences of associated SNPs or InDels identified through BSR-Seq, utilizing Primer3 (https://bioinfo.ut.ee/primer3/). Genetic markers linked to PmWR183, including SSR and STS markers, were developed using the Fielder reference genome for constructing a high-resolution genetic linkage map. Genomic DNA was extracted from leaf tissues using the cetyltrimethylammonium bromide (CTAB) method73. Genetic markers linked to PmWR183 were genotyped on the mapping population from WR183 × Fielder cross to develop a linkage map using Mapmaker 3.074, with a likelihood of odds score threshold of 3.0. The genetic map was constructed with the software Mapdraw V2.175.
Genome sequencing, assembly, and gene annotation
Seeds of line WE35 were sown in a growth chamber under conditions of 16 h light, 20 °C/8 h dark, 18 °C. High-molecular-weight DNA was isolated from seedling leaves of WE35, which carries PmWR183, using large-scale nucleus extraction followed by the Nanobind Plant Nuclei Kit (PacBio). DNA purity was assessed on a NanoDrop NP-1000 spectrophotometer (NanoDrop Technologies). HiFi sequencing libraries were prepared using the SMRTbell enzyme cleanup kit 1.0 (PacBio), followed by size selection electrophoresis with the BluePippin System from SAGE Science. Sequencing was performed on the Pacific Biosciences Sequel II system, following HiFi sequencing protocols. The libraries were sequenced across three PacBio SMART cells (Supplementary Data 3). All HiFi reads were assembled using the default parameters of Hifiasm 0.16.1-r375, and gfatools was used to convert the sequence graphs from GFA to FASTA format76. A 2.5 Mb contig (ctg001568) was captured using the closest flanking markers (WGGBL578 to WGGBL465) of PmWR183, fully covering the physical mapping interval of PmWR183. The sequence of contig ctg001568 was annotated using Softberry FGENESH (http://www.softberry.com/berry.phtml) and NCBI BLASTP against the nr-protein database to identify homologous proteins. Genome sequence similarity comparison was performed using the YASS genomic similarity search tool (https://bioinfo.cristal.univ-lille.fr/yass/yass.php).
RNA-Seq analysis
Total RNA was extracted from grain-filling stage WR183 leaf samples inoculated with Bgt isolate E20 at 48 hpi using the RNAsimple Total RNA Kit (Tiangen Biotech Co., Ltd., Beijing, China). RNA-Seq was carried out using the Illumina HiSeq 4000 platform (Beijing Novogene Bioinformatics Technology Co., Ltd., Beijing, China). High-quality clean reads were generated and aligned to the contig, ctg001568, using HISAT2. Gene expression levels were subsequently quantified with StringTie (https://ccb.jhu.edu/software/stringtie/). Three biological replicates were performed to ensure reliability and reproducibility.
5′ and 3′ rapid amplification of cDNA ends (RACE)
Total RNA was isolated from the grain-filling adult leaves tissue of WR183 inoculated with the Bgt isolate E20 at 24 hpi using the RNAsimple Total RNA Kit (Tiangen Biotech Co., Ltd., Beijing, China). First-strand cDNA was synthesized with a SMARTer® RACE 5′/3′ Kit (TaKaRa, Kyoto, Japan). The full-length cDNA was amplified using specific primers designed based on the genome sequence of contig ctg001568 (Supplementary Data 2), sequenced, and compared with the genomic sequence of PmWR183–NLR1 and PmWR183–NLR2, respectively, to investigate splicing variants. Thirty colonies per PCR reaction were selected for Sanger sequencing. The coding regions and corresponding amino acid sequences were predicted using ORF Finder on NCBI.
Wheat transformation
The dual gRNA CRISPR/Cas9 system was employed to generate PmWR183–NLR1 and PmWR183–NLR2 KO mutants in WR183. The sgRNAs of the target genes were designed using the online tool CRISPRdirect (http://crispr.dbcls.jp/). We selected two gRNA target sites in the exon of the PmWR183–NLR1 (PmWR183–NLR1-Target1: CTGATAGAGGACATGGTGGA; PmWR183–NLR1-Target2: GTCAGAGTTAAAAACCCGCT), and two gRNA target sites in the exon of the PmWR183–NLR2 (PmWR183–NLR2-Target1: CCGCACGAAGCAGCGTGTTT; PmWR183–NLR2-Target2: AGGCGCATTTGCTGGAGATC) as candidate target sequences and cloned them into the CRISPR/Cas9 construct. The recombinant constructs were introduced into WR183 by the Agrobacterium-mediated transformation to generate KO mutants of PmWR183–NLR1 and PmWR183–NLR247. To identify CRISPR KO mutations, specific primers PmWR183–NLR1-2g and PmWR183–NLR2–2g were designed to amplify the target regions of PmWR183–NLR1 and PmWR183–NLR2, respectively, followed by Sanger sequencing.
The full-length coding sequences of the PmWR183–NLR1 and PmWR183–NLR2 genes were amplified from WR183 using the primers PmWR183–NLR1–OE–F/R and PmWR183–NLR2–OE–F/R, respectively. These sequences were cloned into the pLGY-OE3 vector driven by the Ubiquitin promoter of Zea mays, resulting in ProUbi:PmWR183–NLR1 and ProUbi:PmWR183–NLR2 constructs, respectively. Two constructs, ProUbi:PmWR183–NLR1 and ProUbi:PmWR183–NLR2, were transformed into susceptible hexaploid wheat cultivar Fielder, individually and jointly, using Agrobacterium-mediated transformation47. T0 transgenic plants were grown in the greenhouse and screened by PCR analysis and Sanger sequencing using specific primers of PmWR183–NLR1 and PmWR183–NLR2 to obtain transgene-positive plants. Twenty T1 transgenic plants from each transformation event were challenged with Bgt isolate E20 at both the seedling and adult plant stages. Transgenic positive individuals were identified by PCR and confirmed by Sanger sequencing using specific primers of PmWR183–NLR1 and PmWR183–NLR2.
In parallel, a 7427 bp genomic DNA fragment containing the full-length 2742 bp PmWR183–NLR1 coding region, along with 2945 bp upstream of the start codon and 1740 bp downstream of the stop codon, was cloned into the pCAMBIA1300 vector to generate the ProPmWR183–NLR1:PmWR183–NLR1 construct for transformation. Similarly, an 8145 bp genomic DNA fragment comprising the full-length 3399 bp PmWR183–NLR2 coding region, 2790 bp upstream of the start codon, and 1956 bp downstream of the stop codon was cloned into pCAMBIA1300 to produce the ProPmWR183–NLR2:PmWR183–NLR2 construct. The resulting constructs were then individually or simultaneously introduced into the susceptible wheat cultivar Fielder via Agrobacterium-mediated transformation. T0 transgenic lines were identified by PCR amplification and verified by Sanger sequencing. The primer sequences are provided in Supplementary Data 2.
Quantitative real-time RT-PCR
RNA samples were extracted from the two-leaf stage leaves and the jointing stage leaves of WR183 and transgenic positive plants using the RNAsimple Total RNA Kit (Tiangen Biotech Co., Ltd., Beijing, China). First-strand complementary DNA (cDNA) was synthesized from the extracted total RNA using the PrimeScript™ RT Reagent Kit (Takara Co., Ltd., Kyoto, Japan). Quantitative real-time reverse transcription PCR (qRT-PCR) analysis was performed using the SYBR Premix Ex Taq II (Takara Co., Ltd., Kyoto, Japan) on a Roche LightCycler 480 system (Roche, Colorado Springs, CO, USA). The wheat TaActin gene served as the internal reference for normalization, and relative gene expression levels were calculated using the 2−ΔΔCT method77. Each experiment was conducted independently with three biological replicates.
Cell death assay in N. benthamiana leaves
The cell death assay in N. benthamiana leaves was conducted following established protocols78. To enhance expression efficiency in N. benthamiana, the coding sequences of PmWR183–NLR1 and PmWR183–NLR2 were codon-optimized based on tobacco codon usage preferences. The codon-optimized DNA fragments were synthesized by Beijing Tsingke Biotech Co., Ltd. and subsequently cloned into the p1307-C-3×HA or p1307-C-6×MYC vectors to generate tagged fusion constructs for protein expression in N. benthamiana. The resulting plasmids were transformed into Agrobacterium tumefaciens strain GV3101. After overnight culture in LB medium, the bacterial cells were harvested, resuspended in infiltration buffer, and incubated at 30 °C for 2 h before leaf infiltration. Each construct was adjusted to a final concentration of OD₆₀₀ = 1.0. Leaf tissues were photographed 3 days post-infiltration. For western blot analysis, the infiltrated N. benthamiana leaves were harvested for total protein extraction. Protein expression was confirmed by immunoblotting using anti-HA (MBL, M180-7) and anti-MYC (MBL, M192-7) antibodies. Ponceau S staining of Rubisco served as a loading control. Additionally, primer sequences and codon-optimized sequences used in this study are listed in Supplementary Data 2.
Split firefly luciferase complementation (SFLC) assay
The target coding sequences were cloned into SFLC vectors pCAMBIA-cLUC and pCAMBIA-nLUC to generate fusion constructs79. These constructs were introduced into A. tumefaciens strain GV3101 and cultured overnight in LB medium. The bacterial cells were then harvested and resuspended in infiltration buffer (10 mM MgCl₂, 10 mM MES, 150 μM acetosyringone, pH 5.6), followed by incubation at 30 °C for 2 h prior to leaf infiltration. For protein–protein interaction analysis, the constructs were co-infiltrated into the epidermal cells of 5-week-old N. benthamiana leaves. Luciferase substrate (Promega, E1605) was applied to leaves 3 days post-infiltration, followed by incubation in the dark for 10 min prior to imaging. LUC signals were detected using a cooled CCD imaging system (Berthold, LB985). To verify recombinant protein expression, immunoblotting was performed with anti-HA (MBL, M180-7) or anti-MYC (MBL, M192-7) antibodies. Ponceau S staining of Rubisco was used as a loading control. Primer sequences used in this study are provided in Supplementary Data 2.
Co-IP assay
For the co-immunoprecipitation (Co-IP) assay, the coding sequences of the target genes were amplified and cloned into the vectors p1307-C-3×HA and p1307-C-6×MYC to generate constructs expressing fusion proteins with HA and MYC epitope tags in N. benthamiana, respectively. The resulting plasmids were transformed into the A. tumefaciens strain GV3101, which was cultured overnight in LB medium, harvested by centrifugation, and resuspended in infiltration buffer. The bacterial suspensions were incubated at 30 °C for 2 h prior to agroinfiltration into N. benthamiana leaves. After 2 days post-infiltration, N. benthamiana tissues were collected, flash-frozen, and ground in liquid nitrogen. Total proteins were extracted with the IP extraction buffer (150 mM NaCl, 150 mM Tris-HCl, pH 7.5, 1 mM EDTA, 10% glycerol, 10 mM DTT, 0.4% Nonidet P-40, 2% PVPP, 1× protease inhibitor cocktail). The different supernatants were incubated with HA-Trap magnetic beads (MBL, M180-11) at 4 °C for 60–90 min and subsequently washed five times with wash buffer (500 mM NaCl, 20 mM Tris-HCl (pH 8.0), 0.1% Tween 20). The precipitated proteins were analyzed by western blot using anti-HA (MBL, M180-7) or anti-MYC (MBL, M192-7) antibodies. The primer sequences for cloning are listed in Supplementary Data 2.
Evolutionary analysis of PmWR183–NLR1 and PmWR183–NLR2
The protein and genomic sequences of PmWR183–NLR1 and PmWR183–NLR2 were used as queries to identify their orthologs and homologs in the Aegilops species reference genome, including Ae. speltoides, Ae. sharonensis, Ae. longissima, Ae. bicornis and Ae. searsii52,53. Micro-collinearity analysis was conducted through comparison of the annotated genes in the PmWR183 locus.
PmWR183-NLR1 and PmWR183-NLR2 structure predictions
The predicted structures of PmWR183-NLR1 and PmWR183-NLR2 proteins were generated by AlphaFold380. A total of five structural models were predicted, among which the highest confidence model was selected for detailed structural analysis. The 3D structures of the selected models were visualized and refined using the PyMOL software v3.0.3 (https://www.pymol.org/), where specific amino acid features were annotated.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The raw data of the RNA sequencing of PmWR183 are publicly available from the Genome Sequence Archive in the National Genomics Data Center, China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences, under accession CRA029696. The sequences of PmWR183–NLR1 and PmWR183–NLR2 have been submitted to the National Center for Biotechnology Information (NCBI) and are available under the accession PQ807627 and PQ807628. Source data are provided with this paper.
References
-
Lobell, D. B., Schlenker, W. & Costa-Roberts, J. Climate trends and global crop production since 1980. Science 333, 616–620 (2011).
-
Singh, R. P. et al. Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 54, 303–322 (2016).
-
Sotiropoulos, A. G. et al. Global genomic analyses of wheat powdery mildew reveal association of pathogen spread with historical human migration and trade. Nat. Commun. 13, 4315 (2022).
-
Hewitt, T. et al. A highly differentiated region of wheat chromosome 7AL encodes a Pm1a immune receptor that recognizes its corresponding AvrPm1a effector from Blumeria graminis. New Phytol. 229, 2812–2826 (2021).
-
Sanchez-Martin, J. et al. Rapid gene isolation in barley and wheat by mutant chromosome sequencing. Genome Biol. 17, 221 (2016).
-
Yahiaoui, N., Srichumpa, P., Dudler, R. & Keller, B. Genome analysis at different ploidy levels allows cloning of the powdery mildew resistance gene Pm3b from hexaploid wheat. Plant J. 37, 528–538 (2004).
-
Xie, J. Z. et al. A rare single nucleotide variant in Pm5e confers powdery mildew resistance in common wheat. New Phytol. 228, 1011–1026 (2020).
-
Hurni, S. et al. Rye Pm8 and wheat Pm3 are orthologous genes and show evolutionary conservation of resistance function against powdery mildew. Plant J. 76, 957–969 (2013).
-
Zhu, S. Y. et al. Orthologous genes Pm12 and Pm21 from two wild relatives of wheat show evolutionary conservation but divergent powdery mildew resistance. Plant Commun. 4, 100472 (2023).
-
Singh, S. P. et al. Evolutionary divergence of the rye Pm17 and Pm8 resistance genes reveals ancient diversity. Plant Mol. Biol. 98, 249–260 (2018).
-
He, H. G. et al. Pm21, encoding a typical CC-NBS-LRR protein, confers broad-spectrum resistance to wheat powdery mildew disease. Mol. Plant 11, 879–882 (2018).
-
Xing, L. P. et al. Pm21 from Haynaldia villosa encodes a CC-NBS-LRR protein conferring powdery mildew resistance in wheat. Mol. Plant 11, 874–878 (2018).
-
Li, M. M. et al. A CNL protein in wild emmer wheat confers powdery mildew resistance. New Phytol. 228, 1027–1037 (2020).
-
Lu, C. T. et al. Wheat Pm55 alleles exhibit distinct interactions with an inhibitor to cause different powdery mildew resistance. Nat. Commun. 15, 503 (2024).
-
Zou, S. H., Wang, H., Li, Y. W., Kong, Z. S. & Tang, D. Z. The NB-LRR gene Pm60 confers powdery mildew resistance in wheat. New Phytol. 218, 298–309 (2018).
-
Li, Y. H. et al. Dissection of a rapidly evolving wheat resistance gene cluster by long-read genome sequencing accelerated the cloning of Pm69. Plant Commun. 5, 100646 (2024).
-
Ma, C. et al. An Aegilops longissima NLR protein with integrated CC-BED module mediates resistance to wheat powdery mildew. Nat. Commun. 15, 8281 (2024).
-
Sanchez-Martin, J. et al. Wheat Pm4 resistance to powdery mildew is controlled by alternative splice variants encoding chimeric proteins. Nat. Plants 7, 327–341 (2021).
-
Li, H. H. et al. Wheat powdery mildew resistance gene Pm13 encodes a mixed lineage kinase domain-like protein. Nat. Commun. 15, 2449 (2024).
-
Lu, P. et al. A rare gain of function mutation in a wheat tandem kinase confers resistance to powdery mildew. Nat. Commun. 11, 680 (2020).
-
Gaurav, K. et al. Population genomic analysis of Aegilops tauschii identifies targets for bread wheat improvement. Nat. Biotechnol. 40, 422–431 (2022).
-
Li, M. M. et al. A membrane associated tandem kinase from wild emmer wheat confers broad-spectrum resistance to powdery mildew. Nat. Commun. 15, 3124 (2024).
-
Zhao, Y. et al. Pm57 from Aegilops searsii encodes a tandem kinase protein and confers wheat powdery mildew resistance. Nat. Commun. 15, 4796 (2024).
-
Krattinger, S. G. et al. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 323, 1360–1363 (2009).
-
Moore, J. W. et al. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 47, 1494–1498 (2015).
-
Avni, R. et al. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 357, 93–97 (2017).
-
Wu, Q. H. et al. Bulked segregant CGT-Seq-facilitated map-based cloning of a powdery mildew resistance gene originating from wild emmer wheat (Triticum dicoccoides). Plant Biotechnol. J. 19, 1288–1290 (2021).
-
Wu, Q. H. et al. Functional characterization of powdery mildew resistance gene MlIW172, a new Pm60 allele and its allelic variation in wild emmer wheat. J. Genet. Genom. 49, 787–795 (2022).
-
Li, Y. H. et al. TdPm60 identified in wild emmer wheat is an ortholog of Pm60 and constitutes a strong candidate for PmG16 powdery mildew resistance. Theor. Appl. Genet. 134, 2777–2793 (2021).
-
Jones, J. D., Vance, R. E. & Dangl, J. L. Intracellular innate immune surveillance devices in plants and animals. Science 354, aaf6395 (2016).
-
Adachi, H., Derevnina, L. & Kamoun, S. NLR singletons, pairs, and networks: evolution, assembly, and regulation of the intracellular immunoreceptor circuitry of plants. Curr. Opin. Plant Biol. 50, 121–131 (2019).
-
Tamborski, J. & Krasileva, K. V. Evolution of plant NLRs: from natural history to precise modifications. Annu. Rev. Plant Biol. 71, 355–378 (2020).
-
Kourelis, J. & van der Hoorn, R. A. L. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 30, 285–299 (2018).
-
Jubic, L. M., Saile, S., Furzer, O. J., El Kasmi, F. & Dangl, J. L. Help wanted: helper NLRs and plant immune responses. Curr. Opin. Plant Biol. 50, 82–94 (2019).
-
Narusaka, M. et al. RRS1 and RPS4 provide a dual resistance-gene system against fungal and bacterial pathogens. Plant J. 60, 218–226 (2009).
-
Cesari, S. et al. The rice resistance protein pair RGA4/RGA5 recognizes the Magnaporthe oryzae effectors AVR-Pia and AVR1-CO39 by direct binding. Plant Cell 25, 1463–1481 (2013).
-
Xiao, G. et al. The Piks allele of the NLR immune receptor Pik breaks the recognition of AvrPik effectors of rice blast fungus. J. Integr. Plant Biol. 65, 810–824 (2023).
-
Guo, G. H. et al. The wheat NLR pair RXL/Pm5e confers resistance to powdery mildew. Plant Biotechnol. J. 23, 1260–1276 (2025).
-
Zhu, K. Y. et al. An atypical NLR pair TdCNL1/TdCNL5 from wild emmer confers powdery mildew resistance in wheat. Nat. Genet. 57, 1553–1562 (2025).
-
Hu, Y. et al. A head-to-head NLR pair from wild emmer confers resistance to wheat stripe rust. Nat. Genet. 57, 1543–1552 (2025).
-
Klymiuk, V. et al. Coordinated function of paired NLRs confers Yr84-mediated stripe rust resistance in wheat. Nat. Genet. 57, 1535–1542 (2025).
-
Chen, R. J. et al. A wheat tandem kinase activates an NLR to trigger immunity. Science 387, 1402–1408 (2025).
-
Lu, P. et al. A wheat tandem kinase and NLR pair confers resistance to multiple fungal pathogens. Science 387, 1418–1424 (2025).
-
Zhang, D. Y. et al. Wheat powdery mildew resistance gene Pm64 derived from wild emmer (Triticum turgidum var. dicoccoides) is tightly linked in repulsion with stripe rust resistance gene Yr5. Crop J. 7, 761–770 (2019).
-
Zhu, T. T. et al. Optical maps refine the bread wheat Triticum aestivum cv. Chinese Spring genome assembly. Plant J. 107, 303–314 (2021).
-
Sato, K. et al. Chromosome-scale genome assembly of the transformation-amenable common wheat cultivar ‘Fielder’. DNA Res. 28, dsab008 (2021).
-
Ishida, Y., Tsunashima, M., Hiei, Y. & Komari, T. Wheat (Triticum aestivum L.) transformation using immature embryos. Methods Mol. Biol. 1223, 189–198 (2015).
-
Vo, K. T. X. et al. Pi5 and Pii paired NLRs are functionally exchangeable and confer similar disease resistance specificity. Mol. Cells 42, 637–645 (2019).
-
Forderer, A. et al. A wheat resistosome defines common principles of immune receptor channels. Nature 610, 532–539 (2022).
-
Bi, G. et al. The ZAR1 resistosome is a calcium-permeable channel triggering plant immune signaling. Cell 184, 3528–3541 e3512 (2021).
-
Wang, J. et al. Reconstitution and structure of a plant NLR resistosome conferring immunity. Science 364, eaav5870 (2019).
-
Li, L. F. et al. Genome sequences of five Sitopsis species of Aegilops and the origin of polyploid wheat B subgenome. Mol. Plant 15, 488–503 (2022).
-
Avni, R. et al. Genome sequences of three Aegilops species of the section Sitopsis reveal phylogenetic relationships and provide resources for wheat improvement. Plant J. 110, 179–192 (2022).
-
Klymiuk, V. et al. Cloning of the wheat Yr15 resistance gene sheds light on the plant tandem kinase-pseudokinase family. Nat. Commun. 9, 3735 (2018).
-
Fu, D. L. et al. A kinase-START gene confers temperature-dependent resistance to wheat stripe rust. Science 323, 1357–1360 (2009).
-
Ngou, B. P. M., Ding, P. & Jones, J. D. G. Thirty years of resistance: zig-zag through the plant immune system. Plant Cell 34, 1447–1478 (2022).
-
Chen, J. P. et al. Loss of AvrSr50 by somatic exchange in stem rust leads to virulence for Sr50 resistance in wheat. Science 358, 1607–1610 (2017).
-
Allen, R. L. et al. Host-parasite coevolutionary conflict between Arabidopsis and downy mildew. Science 306, 1957–1960 (2004).
-
Yang, Y. et al. Paired plant immune CHS3-CSA1 receptor alleles form distinct hetero-oligomeric complexes. Science 383, 3468 (2024).
-
De la Concepcion, J. C. et al. Polymorphic residues in rice NLRs expand binding and response to effectors of the blast pathogen. Nat. Plants 4, 576–585 (2018).
-
Le Roux, C. et al. A receptor pair with an integrated decoy converts pathogen disabling of transcription factors to immunity. Cell 161, 1074–1088 (2015).
-
Maqbool, A. et al. Structural basis of pathogen recognition by an integrated HMA domain in a plant NLR immune receptor. eLife 4, e08709 (2015).
-
Sarris, P. F. et al. A plant immune receptor detects pathogen effectors that target WRKY transcription factors. Cell 161, 1089–1100 (2015).
-
Kanzaki, H. et al. Arms race co-evolution of Magnaporthe oryzae AVR-Pik and rice Pik genes driven by their physical interactions. Plant J. 72, 894–907 (2012).
-
Deslandes, L. et al. Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proc. Natl. Acad. Sci. USA 100, 8024–8029 (2003).
-
Song, W. Y. et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 270, 1804–1806 (1995).
-
Marla, S. R. et al. Adult plant resistance in maize to northern leaf spot is a feature of partial loss-of-function alleles of Hm1. PLoS Pathog. 14, e1007356 (2018).
-
Cao, Y. et al. The expression pattern of a rice disease resistance gene Xa3/Xa26 is differentially regulated by the genetic backgrounds and developmental stages that influence its function. Genetics 177, 523–533 (2007).
-
Koller, T. et al. Field grown transgenic Pm3e wheat lines show powdery mildew resistance and no fitness costs associated with high transgene expression. Transgenic Res. 28, 9–20 (2019).
-
Bradeen, J. M. et al. Higher copy numbers of the potato RB transgene correspond to enhanced transcript and late blight resistance levels. Mol. Plant Microbe Interact. 22, 437–446 (2009).
-
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
-
Kim, S. et al. Strelka2: fast and accurate calling of germline and somatic variants. Nat. Methods 15, 591–594 (2018).
-
Porebski, S., Bailey, L. G. & Baum, B. R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 15, 8–15 (1997).
-
Lander, E. S. et al. MAPMAKER: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1, 174–181 (1987).
-
Liu, R. H. & Meng, J. L. MapDraw: a microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Yi Chuan 25, 317–321 (2003).
-
Cheng, H. Y., Concepcion, G. T., Feng, X. W., Zhang, H. W. & Li, H. Haplotype-resolved de novo assembly using phased assembly graphs with hifiasm. Nat. Methods 18, 170–175 (2021).
-
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001).
-
Bai, S. et al. Structure-function analysis of barley NLR immune receptor MLA10 reveals its cell compartment specific activity in cell death and disease resistance. PLoS Pathog. 8, e1002752 (2012).
-
Chen, H. M. et al. Firefly luciferase complementation imaging assay for protein-protein interactions in plants. Plant Physiol. 146, 368–376 (2008).
-
Abramson, J. et al. Addendum: accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 636, 493–500 (2024).
Acknowledgements
The authors are grateful to Prof. Qixin Sun and Tsomin Yang of China Agricultural University, Beijing, China, for their advice and support during this research. Many thanks to Drs. Xueyong Zhang, Xianchun Xia, and Lihui Li of the Chinese Academy of Agricultural Sciences, Beijing, China, for providing the Chinese wheat mini-core collection (MCC), the worldwide wheat landrace collection, and Chinese wheat landraces, respectively. This research was financially supported by the National Key Research and Development Program of China (2023YFD1200402 to Z.Y.L., 2023YFF1000402 to G.J.W., 2022YFF1002803 to P.L.), Biological Breeding-National Science and Technology Major Project (2023ZD04025 to H.Z.Z.), National Natural Science Foundation of China (32301834 to H.Z.Z., 30425039, 30771341, 30971780, and 31030056 to Z.Y.L.), and Youth Innovation Promotion Association CAS (2021093 to P.L.).
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Adachi Hiroaki, Javier Sánchez-Martín, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, H., Li, M., Wang, G. et al. Paired NLRs originated from Triticum dicoccoides coordinately confer resistance to powdery mildew in wheat. Nat Commun 16, 9040 (2025). https://doi.org/10.1038/s41467-025-64049-y
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41467-025-64049-y