
Introduction
Weeds significantly impair crop production by competing for nutrients and releasing allelopathic substances that hinder crop growth1. Inadequate weed control is frequently highlighted by farmers as a critical issue leading to considerable economic losses2. Currently, the predominant method for managing weeds involves the use of chemical herbicides. However, their application is associated with numerous risks including adverse effects on public health, soil biology, and environmental pollution, as well as the emergence of herbicide resistance3. Indeed, over 500 cases of resistance have been recorded in 260 weed species against 167 herbicides across 70 countries4. In light of these challenges, there is a pressing need for new, environmentally friendly herbicides derived from natural sources. Bioherbicides, which are advantageous because they are non-toxic to non-target plants and humans, offer target specificity, are effective at low concentrations, and do not contribute to the development of resistance, represent a promising alternative. These agents typically include secondary metabolites produced by phytopathogens5. Fungal metabolites, in particular, have become increasingly recognized in weed control research due to their efficacy in targeting weeds6. For example, bioherbicides derived from fungal sources have shown promising results; a historic case involved the commercialization of Alternaria cuscutacidae spores in the 1950s in Russia to control dodder species4. Fungi are known to be prolific producers of bioactive metabolites, contributing about 70% of all known metabolites used in agriculture and medicine6. Genera such as Alternaria, Botrytis, Colletotrichum, among others, have demonstrated potent herbicidal activities7.
Moreover, the discovery of phytotoxic metabolites like prehelminthosporal and dihydropore from Bipolaris species, and trans-4 aminoproline from Ascochyta caulina, have highlighted the potential of these compounds to inhibit the growth of problematic weeds such as Sorghum halepense and Chenopodium album, respectively8. Despite the urgent need for new herbicides in the face of growing resistance and the high costs associated with chemical herbicides, bioherbicides currently represent only a small fraction of the market9. This indicates a significant gap in the research and development of bioherbicides. Therefore, further investigation into a range of fungal bioherbicides is essential for future advancements in this field10.
Sonchus oleraceus (common sow thistle) and Parthenium hysterophorus (Parthenium weed) are two of the most pervasive and invasive weed species known for their extensive global distribution, affecting more than 50 countries across various continents11,12. These weeds significantly impact agricultural productivity by competing with crops for essential resources and nutrients, ultimately leading to considerable reductions in crop yields13,14. Moreover, the presence of these weeds is not only detrimental to agriculture but also poses a threat to human and animal health. P. hysterophorus, for instance, is notorious for causing allergic reactions, respiratory problems, and dermatitis in humans and animals. Both weeds have a robust adaptability to different environments and can spread rapidly, which makes their management a challenging and critical task for maintaining ecological balance and agricultural health worldwide13,15 .
The present study outlines the purification and NMR-based structural elucidation of a new compound isolated from P. chrysogenum. Following the insights gained from molecular docking and simulation analyses, this compound was evaluated for its bioherbicidal efficacy against two highly invasive weed species, S. oleraceus and P. hysterophorus. The experimental results were encouraging indicating the compound’s potential for possible use as bioherbicide. These initial findings suggest that, with further detailed analysis and comprehensive investigations, the compound could be developed into an effective and environmentally sustainable herbicidal solution for commercial applications.
Methodology
Isolation and purification of fungal strain
Fungal strains for metabolite production were isolated from the soil samples that were collected from various sites within the Botanical garden of the University of Peshawar. These samples were taken from a depth of 4 to 6 inches beneath the soil surface, after the top layer of debris was cleared, and were stored in sterile polyethylene bags. To isolate fungal strains, the serial dilution method was applied, and aliquots were inoculated onto Potato Dextrose Agar (PDA) plates. The inoculated plates were incubated at 28 °C for 10 days to allow fungal growth. Following incubation, distinct colonies were carefully sub-cultured onto fresh PDA plates to ensure purity, and the resulting isolates were maintained for further analysis16.
Identification of fungal strain that showed bioactive potential
The isolated fungal strains underwent preliminary testing for bioactivity against pathogenic bacteria and fungi, as well as for their phytotoxic potential. Only those strains that exhibited bioactivity in these initial tests were selected for further identification and metabolite production. The purified colonies of bioactive fungal strains were cultured on various media to examine their morphology, pigmentation and to observe the color transitions as the colony matures. The structural organization of the conidiophores, conidia and the spore arrangement pattern were observed using the slide culture technique. For this analysis, slides were prepared and stained with lactophenol blue, and subsequently observed under a compound microscope (Nikon Eclipse E200, Japan) at 1000× magnification17.
Fungal metabolites production
For metabolite production, the fungal strain was cultivated in a modified Czapek Yeast Broth (CYB) medium optimized for secondary metabolite synthesis. This medium consists of 3% glucose, 2% peptone, 0.5% KCl, 0.05% MgSO4.7H2O, and 0.001% FeSO4.7H2O, with the pH adjusted to 5.8 (± 0.2). Following inoculation, the culture was incubated in a shaking incubator at 28 °C and 150 rpm for 14 days to promote optimal growth and metabolite accumulation.
Extraction and concentration of crude fungal metabolites
After completion of the incubation period, 250 µL of 40% hydrochloric acid (HCl) was added to each flask to facilitate the separation and settling of media components. An equal volume of ethyl acetate was then introduced, and the mixture was gently shaken intermittently for 20 min to allow efficient partitioning. The fungal mycelia were separated from the broth and mechanically disrupted using an electric blender to release intracellular metabolites. The resulting mixture was further shaken at intervals for 30 min and subsequently filtered through sterile cheesecloth to remove solid debris. The organic layer containing the secondary metabolites was carefully collected using a separating funnel and washed with 2 M brine solution to remove polar impurities. Anhydrous sodium sulfate (Na₂SO₄) was added to the organic phase to eliminate residual moisture. Finally, the organic layer was concentrated under reduced pressure using a rotary evaporator at 45 °C, yielding the crude fungal metabolites for further purification.
Metabolites purification and analysis using HPLC/MS system
In the process of purifying and analyzing fungal compounds, an HPLC/MS system (Waters Corporation UK) was employed, comprising a photodiode array (PDA) detector linked with a Micro-Mass Z-Quattro (ZQ) spectrometer. The solvent system utilized for both analytical and preparative runs consisted of HPLC-grade acetonitrile (CH₃CN), methanol (MeOH), and ultra-pure water, with the addition of formic acid (0.055%).
During the analytical phase, the procedure lasted for 10 min at a flow rate of 1 mL per minute, with an injection volume of 25 µL of crude solution. UV absorption was monitored from 200 to 400 nm. For the preparative phase, a solution with a concentration of 40 mg mL⁻¹ of crude fungal metabolites in HPLC-grade methanol was prepared. A total of 500 µL was injected into the HPLC/MS system, and a methanol-water gradient system ranging from 65% to 90% methanol was used over 102 preparatory HPLC sessions. Each session lasted 30 min, and the flow rate was set at 5 mL/min. UV detection spanned from 200 to 400 nm. The electrospray (ES) mass spectrometry was conducted using a spectrometer with a quadrupole mass analyzer (QM). Quantification of the targeted metabolites was conducted using an Evaporative Light Scattering Detector (ELSD). The purified metabolites were collected in glass tubes via an automatic fraction collector and then evaporated to prepare for NMR analysis. LCMS analysis was conducted with negative ion electrospray ionization (ESI⁻) and positive ion electrospray ionization (ESI⁺), detecting signals at m/z 228 (ESI⁻) and m/z 230 (ESI⁺), confirming the molecular formula C8H8ClN3O3.
Elucidation of the compound structure using nuclear magnetic resonance (NMR) spectroscopy
1D (1H and 13C) and 2D (HSQC, HMBC, COSY) NMR spectra were obtained using a 500 MHz Varian NMR spectrometer. Tetramethylsilane (TMS) served as the internal standard for chemical shifts, reported in parts per million (ppm). For the analysis, 5 mg of the pure compound was dissolved in 0.65 mL of deuterated chloroform. A series of 1D and 2D NMR experiments were conducted on 400 and 500 MHz NMR spectrometers. The 1H NMR spectrum was recorded to identify proton resonances, and the 13C NMR spectrum provided insights into the carbon environments. Heteronuclear Single Quantum Coherence (HSQC), Heteronuclear Multiple Bond Correlation (HMBC), and Correlation Spectroscopy (COSY) techniques were employed to elucidate proton-carbon and proton-proton correlations, facilitating the determination of the compound’s structure. Additionally, the nitrogen (15N) and oxygen (17O) NMR spectra were obtained to analyze heteroatom environments.
Molecular modeling studies
The molecular docking of the compound was performed with 5-enolpyruvyl shikimate-3-phosphate synthase (EPSP synthase) enzyme, which is a broad-spectrum herbicide target18. The enzyme crystal structure was downloaded from Protein Data Bank (PDB) using a three-letter code of 2AAY. The structure has an excellent resolution value of 1.55 Å19. The enzyme structure was then used in UCSF Chimera 1.17, where it was cleaned of co-crystalized structures. The apo-enzyme was then energy minimized via steepest descent and conjugate gradient algorithms for a total of 2000 steps. The enzyme was then imported to PyRx 0.820. The compound 2-((5-chloro-4-oxo-4,5-dihydropyridin-3-yl)amino)-3-oxopropanimidic acid was drawn in ChemDraw 12.0 and converted into .pdb format21. The compound was converted into .pdbqt in PyRx 0.8. The PyRx AutoDock Vina program was utilized to dock the compound at the active pocket of EPSP synthase enzyme. The docking was carried out at active site residues such as Ser169, Ser170, and Ser197. The grid box was set with dimensions of 25 Å along XYZ directions. The number of iterations used for the compound was 100. Only the binding mode with a stable binding energy value in kcal/mol was selected for interaction analysis. The visualization of complex was performed using UCSF Chimera v1.17 and Discovery Studio visualizer v202122.
Molecular dynamics simulations
The strength of the binding modes of the top-ranked poses recruited after molecular docking was assessed by carrying out molecular docking (MD) simulation for a duration of 100 ns. To conduct the MD simulation, the Desmond simulation package was utilized to configure the systems. Using the TIP3P water model, the systems were solvated by water molecules in an orthorhombic box with fixed dimensions of 10 Å and periodic boundaries. The charge was balanced by adding Cl − ions to the water. To prevent Cl − ions from interfering with the interactions, ions located within a 15 Å distance from the ligand atoms were excluded during simulations. OPLS4 force field and the NPT ensemble (temperature (T), pressure (P), and the number of particles (N)) were employed in all the simulation runs. Before carrying out the production simulation, the default Desmond protocol was followed for energy minimization and model relaxation. A cutoff of 9 Å was applied during the simulations to abridge the Lennard–Jones interactions and short-range coulombic interactions. The Particle-Mesh Ewald (PME) summation was used to calculate the long-range electrostatic interactions. Finally, 100 ns MD simulations with a trajectory interval of 200 ps were carried out at a temperature of 300 K and a pressure of 1.01325 bar in the NPT ensemble using a Nose–Hoover chain thermostat and a Martyna–Tobias–Klein barostat. For calculating the long-range electrostatic interactions, Particle-Mesh Ewald (PME) summation method was used. Ultimately, 100 ns MD simulations with a trajectory interval of 200 ps were conducted at a temperature of 300 K and a pressure of 1.01325 bar in the NPT ensemble. The desired conditions were maintained using Nose–Hoover chain thermostat and a Martyna–Tobias–Klein barostat. Further, the trajectories obtained were saved at 2 fs intervals and subjected to further analysis using Simulation Interaction Diagram (SID) and Simulation Event Analysis (SEA) tools applied in the Desmond MD package, allowing deeper insights into the interactions between the compound and the protein. The stability of the complexes was observed by examining the Root Mean Square Deviation (RMSD) and Root Mean Square Fluctuation (RMSF) over time using frame zero as a reference. SEA tool was used to evaluate the distance between the β-lactam electrophilic carbonyl carbon and the Cys-199 nucleophilic thiol sulfur, by adjusting 3.5 Å as a threshold, which accounts for the summation of van der Waals radii of carbon and sulfur atoms. To validate the stability of the poses, MD simulation runs were repeated using a different random seed.
Binding energy calculation through MM-PBSA
g_mmpbsa tool was employed to estimate the binding free energy of the protein-ligand complexes g_mmpbsa tool calculated the binding free energy of complex structure using the Molecular Mechanics Poisson–Boltzmann surface area (MM-PBSA) method. This method calculates the difference between the free energy of the complex and that of the unbound components. The potential energy of solvation (electrostatic + van der Waals interaction) and the free energy of solvation (polar + non-polar solvation energies) were determined for the last 5 ns of molecular dynamics simulation trajectory.
In-vitro herbicidal activity of the pure compound against S. oleraceus and P. hysterophorus
The herbicidal potential of the pure compound derived from P. chrysogenum was assessed against two test weed species, S. oleraceus and P. hysterophorus, which are known to cause significant damage to agricultural lands across Pakistan. The experiment utilized a Completely Randomized Design (CRD) and was conducted in sterile, autoclaved Petri plates, each lined with two layers of sterile tissue paper. Seeds of S. oleraceus and P. hysterophorus were surface-sterilized using a 0.1% solution of mercuric chloride and then sown at equidistant points, with 10 seeds per plate. Various concentrations of the test fungal compound (10, 100, 250, 500, and 1000 µg/mL/mL) were applied to the seeds in the Petri plates using a sterile dropper. The seeds were watered daily as needed using a spray bottle. Each treatment was replicated three times, including an untreated control. In a parallel experiment, the same conditions were replicated using 2, 4-Dichlorophenoxyacetic acid, a selective herbicide, to compare herbicidal effects. Seed germination was monitored daily over 27 days, recording parameters such as germination percentage, shoot and root length (cm), and shoot and root weight (g). Data analysis was performed using SPSS software, with significant differences and means determined by the Least Significant Difference (LSD) test, and results presented as mean ± standard deviation.
Results
Identification of the bioactive fungal strains
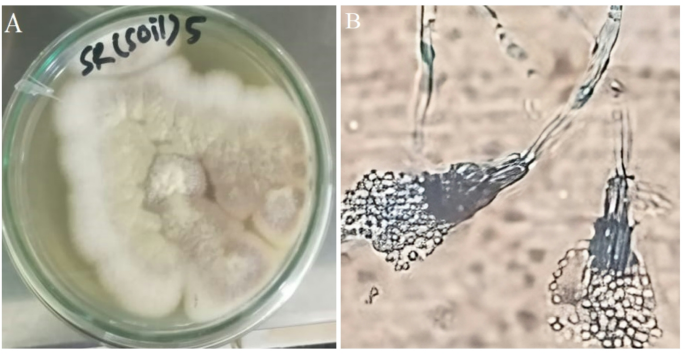
In the identification process, a range of morphological characteristics was systematically examined to identify the bioactive fungal strain. This analysis included detailed observations of conidiophore structures, hyphae, the arrangement of conidia, and the morphologies of colonies. We cultivated the fungal strains on three distinct agar media: Potato Dextrose Agar (PDA), Czapek Dox Agar (CDA), and Malt Extract Agar (MEA), to assess variations in colony pigmentation and the emergence of fruiting bodies. During this observation, notable differences in colony structure and pigmentation were observed based on the growth medium. On PDA, colonies of P. chrysogenum were characterized by a velvety texture with a color palette of dark grey and white. In contrast, colonies on MEA and CDA presented a powdery texture with a greenish-gray hue.
Microscopic investigations further demonstrated that the mycelium was composed of translucent hyphae, occasionally exhibiting subtle green or blue undertones. The conidial formations, primarily cylindrical or oval, were observed at the apices of hyphal branches, frequently forming chain-like structures. These conidial arrangements, which resembled brush strokes, provided distinctive visual markers for the identification of reproductive structures in the strains studied.
The colony morphology, structure of conidiophores and arrangement of conidia are shown in Fig. 1.

(A) Colony morphology of P. chrysogenum. (B) Structure of conidiophores and arrangement of conidia of P. chrysogenum.
Purification and mass determination of the compound
The purification techniques yielded 7.53 mg of the pure compound from an initial 88.6 mg of crude fungal metabolites. LCMS analysis revealed a UV detection profile peaking at 371 nm. The purified compound exhibited a negative ion electrospray ionization (ESI⁻) signal at m/z 228 and a positive ion electrospray ionization (ESI⁺) signal at m/z 230 [M + H]⁺. The compound was identified as having the molecular formula C8H8ClN3O3 with an exact mass of 229.03. The retention time of the compound in the chromatographic system was 11.6 min. The LC-MS chromatogram of the pure compound is presented in Fig. 2.

Liquid Chromatography-Mass Spectrometry (LC-MS) analysis of the compound purified from P. chrysogenum. The figure presents the LC-MS chromatogram of the pure compound. The analysis includes a diode array detection at an ultraviolet maximum absorption (UV λmax) of 371 nm, and electrospray ionization (ESI) profiles in both negative (m/z 228) and positive (m/z 230 with protonation) modes, corresponding to the actual mass of the pure compound (229).
Characterization and description of the purified compound based on NMR spectroscopy
The structural elucidation of the purified compound was supported by detailed NMR analysis in CDCl3.The 1H NMR spectrum revealed well-resolved proton resonances that reflected the unique electronic environments within the molecule (Figure S1). Distinct signals were observed at δ 5.64 (H-2, CH), δ 4.29 (H-10, CH), and δ 9.72 (H-12, CH), each corresponding to protons directly attached to carbons near heteroatoms or conjugated groups. Coupling constants provided additional structural insights, with J = 7.0 Hz between H-4 and H-5 and J = 6.2 Hz between H-10 and H-12, confirming vicinal relationships and spatial proximity. Exchangeable downfield signals at δ 11.97 (NH, H-9), δ 13.23 (OH, H-14), and δ 8.29 (NH, H-15) suggested active hydrogen-bonding sites, consistent with the functional groups indicated in the proposed structure.
The 13C NMR spectrum further supported this interpretation, displaying characteristic resonances at δ 131.5 (C-1), 144.5 (C-2, CH), and 157.9 (C-4, CH), indicative of carbons adjacent to electronegative atoms or conjugated systems (Figure S2). Strong downfield signals at δ 192.8 and 194.5 confirmed the presence of two carbonyl carbons, while δ 52.7 (C-10, CH) indicated a deshielded sp³ carbon in proximity to heteroatoms. Two-dimensional NMR experiments provided key proton–carbon correlations. HSQC confirmed direct attachments, linking H-2 with C-2, H-5 with C-5, and H-10 with C-10. HMBC data revealed long-range couplings between heteroatom-linked protons (NH and OH) and adjacent carbonyl carbons, further validating functional group placement (Figure S3). COSY analysis showed clear spin–spin couplings between H-4/H-5 and H-10/H-12, supporting the molecular connectivity. The 15N and 17O spectrum of the compound is provided in Figure S4 and Figure S5, respectively. Taken together, the 1H, 13C, HSQC, HMBC, and COSY data provided comprehensive confirmation of the compound’s structure with all chemical shifts, multiplicities, and coupling constants fully consistent with the atom numbering shown in Fig. 3. The chemical shifts, multiplicity, and coupling constants (in Hertz, Hz) for both the protons and carbons present in the compound were determined which are given in Table 1. The COSY and HMBC correlations of the pure compound is presented in Fig. 4.

Structure of the pure compound 2-((5-chloro-4-oxo-4, 5-dihydropyridin-3-yl) amino)-3-oxopropanimidic acid.

COSY and HMBC correlations of the pure compound. COSY correlations are denoted by red lines, indicating vicinal coupling between protons, while the HMBC correlations are depicted using blue arrows, illustrating correlations between protons and carbons typically separated by 2–3 bonds.
Molecular Docking studies
Chemically, the compound is 2-((5-chloro-4-oxo-4, 5-dihydropyridin-3-yl) amino)-3-oxopropanimidic acid and is found docked deeply inside the binding pocket of EPSP enzyme. The binding energy of top iteration of the compound with the enzyme is -11.39 kcal/mol. The compound showed a binding conformation that balances itself at the enzyme active pocket. The 2-amino-3-oxopropanimidic acid chemical region of the compound reported robust binding interactions with the enzyme active region. This moiety particularly formed strong hydrogen bonds with Val168, Ser197, Lys340, and Gln171. Similarly, this region produced van der Waals bonds with Asn336, Val196, Ser169, Ser170 and Arg100. The rest 3-chloropyridin-4(3 H)-one chemical region produced a hydrogen bond with Asp313 along with van der Waals interactions with Lys22, Ser23, Arg27, and Thr97. The binding mode and interactions of the compound are given in Fig. 5.

Compound binding to the EPSP synthetase enzyme. The enzyme ribbons are shown by cartoon and is transparent surface. The compound is in red heteroatoms stick. Close view of the docked conformation of the compound is also given. The different binding interactions of the compounds are also provided.
Molecular dynamic simulation
Molecular dynamic simulations present a dynamic relation that concludes the static snapshot determined by docking studies. Simulation studies provide the real-time behavior of docked complexes in sighting the stability of intermolecular interactions over time. This more diligently images the actual phenomenon that occurs within living organisms. For this the simulation complexes were investigated for structural stability in time period of 100 ns. The RMSD graph indicated more pronounced variations between 0 and 8 ns, with a score of 5.9 Å, followed by more variations till 35 ns, with a score of 7.8 Å, further decrease in variations occurs from 35 ns to 67 ns reaching the score of 2.1 Å. 67 ns onwards till 90ns the system achieves a value of 6.2 Å, beyond which the system achieved full stabilization, as depicted in Fig. 6A, with no further observable fluctuations. The N-terminal of compound complex reported high flexibility in the presence of the compound, resulting in higher RMSD value of the region residues. The loop regions being in the loop are naturally more flexible. Another reason may be the compound binding mechanism to gain a more stable binding mode at the binding site and achieve stable interactions. The mean RMSD of system is 7.64 Å reporting the system in good stability level.
Similarly, the residue level fluctuations were investigated using root mean square fluctuation (RMSF) (Fig. 6B). The mean RMSF of the compound complex is 1.51 Å, respectively. In the RMSD, the global structure was noticed in overall global stability, which can also be interpreted from the net RMSF value of complexes. The integrity of the protein is investigated by plotting the radius of gyration (Rg) values against time (Fig. 6C). The radius of gyration describes the overall spread of the molecule and is defined as the root mean square distance of the collection of atoms from their common center of gravity. In the present MD studies, we determined the mean Rg value of compound complex 23.2 angstroms, reporting the stability of the system. The structural fluctuation of different regions of the compound and ligand protein was investigated using β-factors; a mean value of 67.62 angstrom square of the system is reported, showing the stability of the system (Fig. 6D).

RMSD (A), RMSF (B), Rg (C) and β-factor (D) analyses of the simulated docked complex.
Binding free energies
Table 2 illustrates the binding free energies of the complex, resolved by the MM-PBSA method throughout the molecular dynamics simulation. Based on simulation trajectories, estimation of binding free energies is considered to be more reliable and sounder with experimental results. The negative binding values of the complex validate the docking and MD simulation results and point to the compound as a good binder to EPSP synthetase enzyme. The complex was classified as stable due to negative net binding energy of − 55.18 kcal/mol. The major contribution comes from van der Waals energy. For example, van der Waals energy is − 48.64 kcal/mol. The electrostatic energy and gas phase energies are also dominating with net energy contributions of − 16.01 kcal/mol and − 64.65 kcal/mol, respectively.
In-vitro herbicidal activity
To determine the herbicidal potential of the compound, seed germination data were collected for seeds placed in sterile petri plates and observed over 27 days. The compound and the standard herbicide 2, 4-Dichlorophenoxyacetic acid (2, 4-D) were tested at concentrations of 10, 100, 250, 500, and 1000 µg/mL/mL against two weed species, S. oleraceus and P. hysterophorus. The plates were regularly examined every 24 h, and the final results were recorded after 27 days. The seed germination data, including shoot and root lengths (recorded in centimeters) and shoot and root weights (recorded in grams), for both the compound and 2, 4-Dichlorophenoxyacetic acid against S. oleraceus and P. hysterophorus are presented in Tables 3, 4 and 5, and Table 6.
In-vitro herbicidal activity of the pure compound against P. hysterophorus and S. oleraceus
After mass determination and structural elucidation, the isolated compound was tested for herbicidal activity against two common weed species (S. oleraceus and P. hysterophorus) of wheat crops. The pure compound was dissolved in DMSO to prepare stock solutions, from which five different dose concentrations (10, 100, 250, 500, 1000 µg/mL/mL) were prepared and applied to the seeds of both weed species. The results showed that the compound actively inhibited the growth of both weed species. However, compared to P. hysterophorus, the compound exhibited greater herbicidal activity against S. oleraceus.
For P. hysterophorus, control treatment showed 100% seed germination with average shoot and root lengths measured at 8.93 cm and 13.70 cm, respectively, while the average root and shoot biomass in the control treatment was found to be 0.074 g and 0.0546 g, respectively. At a concentration of 10 µg/mL/mL of the compound 100% germination was recorded, where the average shoot length was 8.34 cm and the average root length was 13.20 cm. The biomass of roots and shoots at this concentration was 0.070 g and 0.0546 g, respectively. At higher concentrations, a gradual decrease in germination percentage and growth measurements was observed. A significant reduction in germination to 30% was recorded at a concentration of 500 µg/mL, where both shoot and root lengths diminished substantially to 2.94 cm and 4.25 cm, respectively. Correspondingly, the biomasses were measured at 0.018 g for shoots and 0.029 g for roots. At the maximal concentration tested, 1000 µg/mL, germination fell to 10%, with shoots and roots exhibiting minimal growth, measuring 1.22 cm and 1.25 cm in length, respectively, and their biomasses were recorded at 0.010 g for shoots and 0.013 g for roots. The herbicidal activity of the pure compound against P. hysterophorus and the respective concentrations are presented in Table 3.
In the control treatment for S. oleraceus, complete germination (100%) was observed, with the average shoot and root lengths measuring 12.94 cm and 10.90 cm, respectively. The shoot and root biomasses were recorded at 0.085 g and 0.016 g, respectively. At a concentration of 10 µg/mL/mL of the pure compound, the germination rate decreased slightly to 90%, with resultant average shoot and root lengths of 7.82 cm and 6.17 cm, respectively, and biomasses of 0.0493 g for shoots and 0.0133 g for roots. At 100 µg/mL/mL, the germination further declined to 70%, with corresponding shoot and root lengths of 4.96 cm and 3.88 cm, and biomasses of 0.0294 g for shoots and 0.010 g for roots. A notable reduction in germination to 40% was recorded at 250 µg/mL/mL; seeds began germinating after 18 days, producing significantly shorter shoot and root lengths of 2.57 cm and 2.16 cm, and reduced biomasses of 0.013 g for shoots and 0.002 g for roots. At higher concentrations (500 µg/mL and 1000 µg/mL), seed germination was completely inhibited, demonstrating the potent herbicidal effect of the compound. The detailed herbicidal activity results of the pure compound against S. oleraceus and the respective concentrations are provided in Table 4.
In-vitro herbicidal activity of the standard herbicide (2, 4-Dichlorophenoxyacetic acid)
To compare the weedicidal efficacy of the pure compound, a parallel experiment was conducted using 2, 4-Dichlorophenoxyacetic acid, which was used as a standard herbicide. This standard compound was dissolved in DMSO to prepare a stock solution from which five different concentrations (10, 100, 250, 500, 1000 µg/mL/mL) were prepared and applied using a sterile dropper at regular intervals on the seeds of both test weed species. The results demonstrated that 2, 4-Dichlorophenoxyacetic acid effectively inhibited the growth of both species, showing greater herbicidal activity against S. oleraceus compared to P. hysterophorus.
For P. hysterophorus, control treatments showed 100% seed germination with average shoot and root lengths of 8.93 cm and 13.70 cm, respectively, and biomasses of 0.074 g for roots and 0.0546 g for shoots. At a concentration of 10 µg/mL of 2, 4-Dichlorophenoxyacetic acid, germination remained at 100%, with average shoot and root lengths slightly reduced to 8.28 cm and 13.65 cm, respectively, and biomasses of 0.072 g for roots and 0.0538 g for shoots. At 100 µg/mL, germination decreased to 90%, with shoot and root lengths of 7.85 cm and 13.20 cm, and biomasses of 0.064 g and 0.0520 g, respectively. At 250 µg/mL, 80% of seeds germinated, with shoot and root lengths of 6.12 cm and 12.05 cm, and biomasses of 0.049 g for roots and 0.0404 g for shoots. At 500 µg/mL, 70% germination was observed, with further reductions in shoot and root lengths to 5.48 cm and 10.13 cm, and biomasses of 0.039 g for shoots and 0.033 g for roots. At the highest concentration of 1000 µg/mL/mL, germination dropped to 50%, with shoot and root lengths of 3.68 cm and 4.92 cm, and biomasses of 0.017 g for shoots and 0.022 g for roots. The effects of 2, 4-Dichlorophenoxyacetic acid on P. hysterophorus across these concentrations are detailed in Table 5.
For S. oleraceus, control plots achieved 100% germination, with average shoot and root lengths of 12.94 cm and 10.90 cm, respectively, and biomasses of 0.085 g for shoots and 0.016 g for roots. At 10 µg/mL/mL of 2, 4-Dichlorophenoxyacetic acid, 90% of seeds germinated, producing shoot and root lengths of 12.98 cm and 9.96 cm, and biomasses of 0.0828 g for shoots and 0.0160 g for roots. At 100 µg/mL, germination reduced to 70%, with shoot and root lengths of 9.45 cm and 7.59 cm, and biomasses of 0.0740 g for roots and 0.0157 g for shoots. At 250 µg/mL/mL, only 50% of seeds germinated, with shoot and root lengths of 4.12 cm and 2.05 cm, and biomasses of 0.0305 g for shoots and 0.0110 g for roots. A significant reduction in germination to 20% was noted at 500 µg/mL, with shoot and root lengths further reduced to 1.42 cm and 0.98 cm, and biomasses of 0.015 g for shoots and 0.0004 g for roots. At 1000 µg/mL/mL, no seed germination occurred, indicating complete inhibition of S. oleraceus. The herbicidal activities of 2, 4-Dichlorophenoxyacetic acid against S. oleraceus and the corresponding concentrations are documented in Table 6.
Discussion
Our study extends the understanding of fungal secondary metabolites potential as natural herbicides, specifically identifying a novel compound from P. chrysogenum with significant herbicidal activity. This finding verifies the known capabilities of fungi, such as Fusarium oxysporum and Alternaria alternata, to produce bioactive compounds that inhibit plant growth, positioning fungal metabolites as promising alternatives to synthetic herbicides23,24. The herbicidal potential of fungal secondary metabolites has garnered considerable attention, with species such as Fusarium oxysporum and Fusarium solani recognized for their production of fusaric acid, a compound rigorously studied for its ability to inhibit specific weed species, demonstrating its utility in agricultural settings25. Similarly, Alternaria alternata is known to produce tenuazonic acid, a metabolite effective in suppressing a range of plant species, suggesting its potential for managing unwanted weed flora26. Additionally, certain strains of Penicillium, typically known for antibacterial agents like penicillin, also produce herbicidal compounds such as patulin. The capacity of patulin to inhibit plant growth makes it a promising candidate for further research in weed management strategies27. This study further contributes to the field by identifying a compound derived from P. chrysogenum as a promising candidate for use as a natural herbicide. This compound has been rigorously evaluated through extensive computational modeling, in-vitro testing, and structure-activity relationship (SAR) analysis. Together, these assessments demonstrate their efficacy and potential as bioherbicide, highlighting its preliminary suitability for sustainable agricultural practices.
Following the purification and structural elucidation of the pure compound derived from P. chrysogenum, the compound underwent rigorous molecular docking and simulation studies. This process was essential to assess its potential interactions with target enzymes and to evaluate the stability and efficacy of its binding within biological systems. These computational analyses provided valuable insights into the molecular mechanisms by which the compound might exert its herbicidal effects, laying the groundwork for further experimental validation and potential application in weed management28. In our molecular docking and dynamic simulation studies of the pure compound naming 2-((5-chloro-4-oxo-4, 5-dihydropyridin-3-yl) amino)-3-oxopropanimidic acid demonstrated a high affinity for the EPSP synthase enzyme, docking deeply within its active pocket with a notable binding energy of -11.39 kcal/mol. The compound’s chemical regions formed strong hydrogen and van der Waals interactions with critical residues, stabilizing it within the enzyme. Dynamic simulations further confirmed the stability of these interactions over time, with the system achieving a stable configuration as indicated by the root mean square deviation (RMSD) and radius of gyration metrics. The overall stability and binding efficacy of the compound, underscored by negative binding free energies from MM-PBSA analysis, validate its potential as a potent and stable inhibitor suitable for herbicide development.
The observed in vitro herbicidal effects of the pure compound against S. oleraceus and P. hysterophorus are particularly noteworthy. Our results show a differential sensitivity between these species, with stronger inhibition observed against S. oleraceus. This species-specific efficacy contrasts with the broader activity spectrum typically reported for synthetic herbicides like 2, 4-Dichlorophenoxyacetic acid (2, 4-D). Such findings highlight the potential for developing targeted herbicidal strategies that minimize environmental impact while addressing specific agricultural challenges29.
Comparatively, our compound exhibited superior performance at lower concentrations against P. hysterophorus compared to 2, 4-D, which maintained higher germination rates even at increased concentrations. This suggests a more potent or direct mode of action of the fungal-derived compound, possibly related to its unique structural characteristics24. Structural similarities with established chemical herbicides such as isoxaflutole and atrazine—both known for their specific enzyme inhibition pathways suggest potential mechanisms of action. The presence of ketonic functional groups and nitrogen-containing rings may interfere with critical pathways such as the tyrosine catabolic pathway30, in a manner analogous to isoxaflutole inhibition of 4-hydroxyphenylpyruvate dioxygenase. Moreover, the chlorine atom on an aromatic ring within our compound’s structure likely enhances its lipophilicity, facilitating better penetration and persistence within plant tissues of weeds31. This feature could explain the observed efficacy in inhibiting plant growth, suggesting that similar physicochemical properties may be exploited to enhance the bioactivity of future herbicidal formulations. When compared with previously reported fungal-derived herbicides, our compound showed efficacy that is comparable or in some respects more promising.
For instance, fusaric acid, a phytotoxin from Fusarium species, is known to cause chlorosis and wilting in a range of plants but often requires relatively higher concentrations to achieve consistent inhibition32. Similarly, tenuazonic acid, a metabolite from Alternaria species, demonstrates strong herbicidal activity but has also been associated with cytotoxicity, which can limit its agricultural applicability33. In contrast, our compound achieved substantial inhibition of seed germination and seedling growth at 500–1000 µg/mL mL⁻¹ without evidence of broad, non-specific toxicity. The chlorine-substituted aromatic ring and lipophilic character may further enhance its penetration and persistence within plant tissues31. Together, these comparisons place our compound within the growing class of fungal-derived bioherbicides, with potentially greater selectivity and reduced environmental burden.
Our findings emphasize the need for further studies to confirm the herbicidal potential of this compound beyond laboratory conditions. Future work should address its environmental fate, including degradation and persistence in soil and water, as well as possible effects on non-target organisms such as beneficial microbes and pollinators. These assessments will be critical to ensure that the compound can be developed as a safe and sustainable bioherbicide.
Conclusion
In conclusion, our integrated in-silico and in-vitro studies strongly suggest that the P. chrysogenum derived compound, 2-((5-chloro-4-oxo-4, 5-dihydropyridin-3-yl) amino)-3- oxopropanoic acid, is a potent herbicidal agent. Molecular docking and dynamic simulations highlight its robust binding affinity to EPSP synthase, essential for plant growth, with stable interaction metrics such as RMSD supporting its efficacy. Invitro tests further demonstrate its superior inhibitory effects on seed germination and growth, particularly against S. oleraceus, compared to the standard herbicide 2, 4-Dichlorophenoxyacetic acid. These findings highlights the compound’s potential as a more effective and environmentally friendly alternative for weed control, making it a promising candidate for agricultural applications subject to further investigations.
Study limitation
This study was limited to laboratory-scale assays and computational analyses. Field experiments were not performed, and key parameters such as selective toxicity toward non-target organisms, as well as the compound’s stability and degradability in the environment, remain to be evaluated. These aspects will be essential in future work to fully establish the compound’s potential as a safe and sustainable bioherbicide.
Data availability
The data that support the findings of the present study are available from the corresponding authors upon reasonable request.
References
-
Choudhary, C. S. et al. Mechanisms of allelopathic interactions for sustainable weed management. Rhizosphere 25, 100667 (2023).
-
Mohammed, Y. M. & Badawy, M. E. Potential of phytopathogenic fungal isolates as a biocontrol agent against some weeds. Egypt. J. Biol. Pest Control. 30, 1–9 (2020).
-
Akbar, M., Khalil, T., Andolfi, A. & Javaid, A. Isolation and identification of natural herbicidal compound from a plant pathogenic fungus, Drechslera biseptata. Pak J. Bot. 52, 2245–2249 (2020).
-
Roberts, J., Florentine, S., Fernando, W. D. & Tennakoon, K. U. Achievements, developments and future challenges in the field of bioherbicides for weed control: A global review, Plants, vol. 11, p. 2242, (2022).
-
Namasivayam, S. K. R., Aruna, A. & Choudhury, S. H. Selective isolation and screening of fungi with herbicidal potential and evaluation of herbicidal activity against Vernonia species. African J. Biotechnology, 13, (2014).
-
Cheng, L. et al. Study on herbicidal potential of two fungi in Qinghai region. Open. Access. Libr. J. 9, 1–20 (2022).
-
Javaid, A. Herbicidal potential of allelopathic plants and fungi against parthenium hysterophorus–a review. Allelopathy J. 25, 331–334 (2010).
-
Marrone, P. G. Status of the biopesticide market and prospects for new bioherbicides. Pest Manag. Sci. 80, 81–86 (2024).
-
Assaf, C. E. H. et al. Regulation of secondary metabolism in the penicillium genus. Int. J. Mol. Sci. 21, 9462 (2020).
-
Grijseels, S. et al. Physiological characterization of secondary metabolite producing penicillium cell factories. Fungal Biology Biotechnol. 4, 1–12 (2017).
-
Peerzada, A. M., O’Donnell, C. & Adkins, S. Biology, impact, and management of common Sowthistle (Sonchus oleraceus L). Acta Physiol. Plant. 41, 1–17 (2019).
-
Shabbir, A., Zalucki, M. P., Dhileepan, K., Khan, N. & Adkins, S. W. The Current and Potential Distribution of Parthenium Weed and Its Biological Control Agent in Pakistan, Plants, vol. 12, p. 1381, (2023).
-
Mobli, A., Matloob, A. & Chauhan, B. S. The response of glyphosate-resistant and glyphosate-susceptible biotypes of annual Sowthistle (Sonchus oleraceus) to Mungbean density. Weed Sci. 67, 642–648 (2019).
-
Zareen, S., Khan, N. & Rahman, S. Distributions of invasive weed parthenium (Parthenium hysterophorus L.) in the university campus Peshawar KPK. Acta Ecol. Sin. 41, 10–17 (2021).
-
Ruzmi, R. et al. ,., Growth performance of malaysian Parthenium hysterophorus under various environmental variables, Agriculture, vol. 11, p. 856, (2021).
-
Aziz, N. H. & Zainol, N. Isolation and identification of soil fungi isolates from forest soil for flooded soil recovery, in IOP conference series: materials science and engineering, p. 012028. (2018).
-
RATNAWATI, R., SUDEWI, S., JAYA, K. & SALEH, A. R. Microbial exploration from two different ecosystems in central Sulawesi, Indonesia. Biodiversitas J. Biol. Diversity, 23, (2022).
-
Leino, L. et al. Classification of the glyphosate target enzyme (5-enolpyruvylshikimate-3-phosphate synthase) for assessing sensitivity of organisms to the herbicide. J. Hazard. Mater. 408, 124556 (2021).
-
Priestman, M. A., Healy, M. L., Funke, T., Becker, A. & Schönbrunn, E. Molecular basis for the glyphosate-insensitivity of the reaction of 5-enolpyruvylshikimate 3-phosphate synthase with Shikimate. FEBS Lett. 579, 5773–5780 (2005).
-
Ounthaisong, U. & Tangyuenyongwatana, P. Cross-docking study of flavonoids against tyrosinase enzymes using PyRx 0.8 virtual screening tool, TJPS, vol. 41, (2017).
-
Milne, G. W. Software Review of ChemBioDraw 12.0 Ed (ACS, 2010).
-
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
-
Liu, S. et al. New fusaric acid derivatives from the endophytic fungus fusarium oxysporum and their phytotoxicity to barley leaves. J. Agric. Food Chem. 64, 3127–3132 (2016).
-
Bibi, G. et al. Effect of culture filtrates of alternaria alternata on seed germination and seedling growth of Sesame. Archives Phytopathol. Plant. Prot. 56, 625–635 (2023).
-
Palmieri, D. et al. ,., Alkaline pH, low iron availability, poor nitrogen sources and CWI MAPK signaling are associated with increased fusaric acid production in Fusarium oxysporum, Toxins, vol. 15, p. 50, (2023).
-
Sotelo-Cerón, N. D., Martínez-Alvarez, J. C. & Maldonado-Mendoza, I. E. Recent advances in the bioherbicidal potential of tenuazonic acid, an alternaria spp. Mycotoxin. J. Mex. Chem. Soc. 67, 266–274 (2023).
-
Guo, Y. et al. Action mode of the Mycotoxin patulin as a novel natural photosystem II inhibitor. J. Agric. Food Chem. 69, 7313–7323 (2021).
-
Xu, Y. et al. Computational and experimental investigations on the interactions of aryloxy-phenoxy-propionate herbicides to Estrogen receptor alpha in zebrafish. Ecotoxicol. Environ. Saf. 189, 110003 (2020).
-
Berestetskiy, A. Modern approaches for the development of new herbicides based on natural compounds, Plants, vol. 12, p. 234, (2023).
-
Scheelje, F. C. M., Destaso, F. C., Cramail, H. & Meier, M. A. Nitrogen-containing polymers derived from terpenes: possibilities and limitations. Macromol. Chem. Phys. 224, 2200403 (2023).
-
Magnoli, K., Carranza, C., Aluffi, M., Magnoli, C. & Barberis, C. Fungal biodegradation of chlorinated herbicides: An overview with an emphasis on 2, 4-D in Argentina, Biodegradation, vol. 34, pp. 199–214, (2023).
-
Munkvold, G. P., Proctor, R. H. & Moretti, A. Mycotoxin production in fusarium according to contemporary species concepts. Annu. Rev. Phytopathol. 59, 373–402 (2021).
-
Sotelo-Cerón, N. D., Martínez-Álvarez, J. C. & Maldonado-Mendoza, I. E. Recent advances in the bioherbicidal potential of tenuazonic acid, an alternaria spp. Mycotoxin. J. Mex. Chem. Soc. 67, 266–274 (2023).
Acknowledgements
The Researchers would like to thank the Deanship of Graduate Studies and Scientific Research at Qassim University for financial support (QU-APC-2025).
Funding
The Researchers would like to thank the Deanship of Graduate Studies and Scientific Research at Qassim University for financial support (QU-APC-2025).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Khattak, S.U., Ahmad, S., Saleem, A. et al. Penicillium chrysogenum originated chloro-diydropyridyl-oxopropanimidic acid derivative as a potent EPSP synthase-targeted bioherbicide against invasive weed species. Sci Rep 15, 44480 (2025). https://doi.org/10.1038/s41598-025-28071-w
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
DOI: https://doi.org/10.1038/s41598-025-28071-w