
Introduction
Medicinal plants have long been integral to global healthcare systems due to their capacity to synthesize a diverse array of specific organic compounds known as secondary metabolites1. These compounds form the basis of traditional medicine systems such as Ayurveda, Unani, and others. Even in modern contexts, bioactive constituents derived from medicinal plants remain crucial in pharmaceuticals and various industrial applications2. From a plant biology perspective, secondary metabolites play a vital role in defending plants against both biotic and abiotic stressors, including pathogens, herbivores, ultraviolet (UV) radiation, and oxidative stress3. Studies have shown that compounds such as flavonoids, alkaloids, and terpenoids contribute significantly to plant adaptation and survival. These same compounds are also largely responsible for the therapeutic properties of medicinal plants4.
One prominent example of such a plant is Mentha piperita L., commonly known as peppermint. It is a naturally occurring hybrid between Mentha spicata (2n = 48) and Mentha aquatica (2n = 96), and is classified within the Lamiaceae family as a hexaploid species5,6. This economically and medicinally valuable perennial plant is widely cultivated in temperate regions, including Europe, Asia, the United States, India, and the Mediterranean7.
Peppermint essential oil holds high commercial value in the food, cosmetic, and pharmaceutical industries, largely due to its wide range of bioactive compounds. The oil is primarily composed of terpenes, with monoterpenes making up about 52% and sesquiterpenes around 9% 8. Key monoterpenes such as menthol, menthone, limonene, isomenthone, and menthofuran are responsible for its characteristic aroma and a variety of biological effects9. Among these, menthol stands out as the most abundant and biologically active compound. It exhibits antimicrobial, analgesic, and insect-repellent properties and is widely used in medicinal and hygienic products10. Clinically, menthol plays a role in relieving symptoms of irritable bowel syndrome (IBS) by modulating nociceptive and inflammatory pathways11. Additionally, research suggests that menthol may contribute to cholesterol regulation and the inhibition of liver fibrosis progression12. The global industrial relevance of menthol is highlighted by an annual demand of approximately 30,000–32,000 metric tons, ranking it among the most in-demand natural flavoring agents alongside vanilla and citrus13.
The essential oil of peppermint is synthesized and stored specifically in glandular trichomes found on the aerial parts of the plant14. The biosynthesis of menthol begins in coordination with leaf development, as enzymatic activity initiates the accumulation of essential oils within these trichomes. As the plant matures, menthol content increases progressively, reaching its maximum concentration at the flowering stage15.
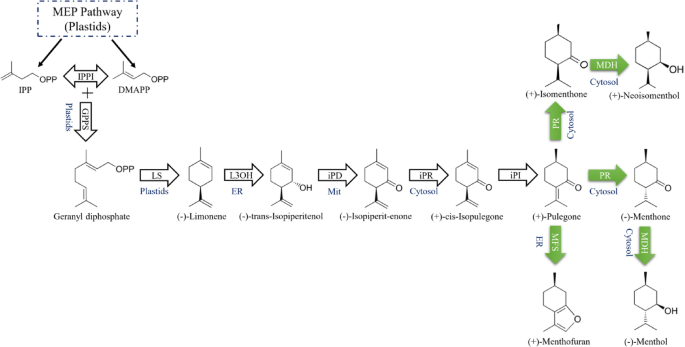
The biosynthetic pathway of menthol in peppermint consists of eight steps, originating from primary metabolism and utilizing the plastidial methylerythritol phosphate (MEP) pathway to supply the C5 isoprenoid precursors, isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP). This pathway begins with the condensation of these precursors to form geranyl diphosphate, which undergoes cyclization to yield (-)-limonene. Subsequent hydroxylation, isomerization, and multiple redox transformations establish the chiral structure of menthol15 (Fig. 1). Although the monoterpene biosynthetic pathway in the Mentha genus has been fully elucidated, the regulatory mechanisms underlying this pathway and the molecular factors that control gene expression remain poorly understood. Recent studies, however, have identified two MYB transcription factors, MhMYB1 and MhMYB2, which play a critical regulatory role by modulating the expression of key enzymatic genes, particularly those encoding menthol dehydrogenase (MDH) and pulegone reductase (PR). These factors significantly influence the biosynthesis of menthol16.

The menthol biosynthetic pathway in the secretory gland cells of peppermint. The arrows indicate the enzymes involved in menthol biosynthesis, while the green arrows highlight the genes analyzed in this study. The verified subcellular locations are identified, with ER representing the endoplasmic reticulum and Mit denoting the mitochondria. The enzymatic reactions are catalyzed by geranyl diphosphate synthase (GPPS), (−)-limonene synthase (LS), (−)-limonene-3-hydroxylase (L3OH), (−)-trans-isopiperitenol dehydrogenase (iPD), (−)-isopiperitenone reductase (iPR), (+)-cis-isopulegone isomerase (iPI), (+)-pulegone reductase (PR), (−)-menthol dehydrogenase (MDH), and menthofuran synthase (MFS). The C5 precursors, IPP and DMAPP, originate from the MEP pathway and serve as initial substrates in the menthol biosynthetic pathway15,17.
The optimization of plant metabolite production for pharmaceutical and industrial applications, through genetic engineering or elicitor-mediated production under tissue culture conditions, fundamentally depends on identifying metabolic pathways and the regulatory factors that control these processes. Consequently, the use of a wide range of biotic and abiotic elicitors in both in vivo and in vitro settings has become a key focus in plant biotechnology and breeding. These strategies aim to uncover the molecular mechanisms, identify effective elicitors, and develop efficient protocols that enhance both the yield and quality of bioactive compounds18.
Click or tap here to enter text.Click or tap here to enter text.Click or tap here to enter text.Click or tap here to enter text.Click or tap here to enter text.Click or tap here to enter text.Click or tap here to enter text.Click or tap here to enter text.Phytohormones are integral to the regulation of plant growth, development, and stress adaptation, with increasing evidence highlighting their central role in secondary metabolism19,20. Key phytohormones such as jasmonic acid (JA), salicylic acid (SA), and abscisic acid (ABA) modulate biosynthetic pathways by regulating gene expression and facilitating post-translational modifications. These hormones promote the production and accumulation of bioactive secondary metabolites by activating complex transcriptional networks responsive to environmental cues23,24. Through these mechanisms, phytohormones enhance plant metabolic plasticity and strengthen defense capabilities under both biotic and abiotic stress conditions19.
Beyond their classical roles, phytohormones have emerged as dynamic regulators of stress-induced secondary metabolism through multilayered mechanisms. Recent findings indicate that hormones such as methyl jasmonate (MJ) and SA not only directly regulate the expression of biosynthetic genes but also modulate broader stress signaling networks19,29. One such network involves the controlled induction of oxidative stress, in which reactive oxygen species (ROS) function as signaling intermediates to activate antioxidant defenses and trigger the accumulation of secondary metabolites28 (Fig. 2).

A schematic illustration depicts the application of phytohormonal elicitor treatments via foliar spraying, along with the complexity of signal transduction cascades associated with the biosynthesis of secondary metabolites in response to phytohormone exposure within the glandular trichomes (orange glands) of peppermint. Upon phytohormone treatment, the interaction between plant hormones and cellular components leads to the extensive generation of reactive oxygen species (ROS)31. These ROS, induced by hormonal signaling, intricately mediate multifaceted responses, including activation of mitogen-activated protein kinase (MAPK) pathways, Ca²⁺ bursts, modulation of signaling molecules, and enhancement of antioxidant defense mechanisms32. Collectively, these processes may finely regulate transcriptional dynamics associated with secondary metabolism. In response to ROS accumulation, the plant cellular machinery rapidly initiates a well-coordinated defensive response aimed at mitigating ROS-induced damage31. Additionally, the direct regulation of gene expression by phytohormones also plays a significant role in orchestrating these biosynthetic pathways19. The peppermint leaf surface image was obtained from Science Photo Library (www.sciencephoto.com), and the overall diagram was created using BioRender (www.biorender.com).
Although significant progress has been made in understanding the role of phytohormones in the regulation of secondary metabolite biosynthesis, several critical questions remain unresolved. For instance, how do different phytohormone signaling pathways interact to influence the activation of biosynthetic pathways? What is the nature of the cross-talk between phytohormones and other cellular signaling networks? Moreover, what factors underlie the differential effectiveness of specific phytohormones in promoting the biosynthesis of particular metabolites?
In this study, we aimed to investigate the effects of four key phytohormones, MJ, ABA, SA, and GA, on oxidative responses, the expression of key transcription factors, and genes involved in the biosynthetic pathway of monoterpenes, specifically menthol, menthone, pulegone, and menthofuran, as well as the biosynthesis and accumulation of these metabolites in peppermint. By comparing the effects of these phytohormones, we aimed to explore their influence on molecular and metabolic regulation at both the transcriptional and biosynthetic levels. This research may provide valuable insights into the role of phytohormones in modulating metabolic processes and the synthesis of bioactive compounds, which could contribute to optimizing the production of plant-derived compounds in various industries.
Materials and methods
Plant material
Rhizomes of peppermint were obtained from the Institute of Medicinal Plants (Karaj, Iran). Healthy rhizomes were cut into approximately 10 cm segments and planted in pots filled with loam soil. The pots were maintained in a controlled growth chamber under a 16/8 h light/dark photoperiod at 25 ± 2 °C. Plants were irrigated at three-day intervals.
Phytohormone treatments
The phytohormones used in this study were obtained from Sigma-Aldrich, Germany. Solutions of four phytohormones MJ, SA, ABA, and GA were prepared by dissolving them in a 0.3% (v/v) ethanol solution. Three molar concentrations (0, 0.2, and 0.4 mM) of each phytohormone were prepared for treatment application. The first foliar spray was applied 60 days after rhizome planting, during the early stages of flowering, ensuring that all leaves were thoroughly coated with the solution. The spraying was performed using a hand pump sprayer to achieve uniform coverage, with a spray volume of approximately 15 mL per pot. The second foliar spray was conducted 7 days after the first, coinciding with 40% flowering of the plants33,34. The control plants received distilled water mixed with 0.3% (v/v) ethanol at the same time as the treatments. For each treatment, three replicates were evaluated. Following the final foliar application, after 48 h, the leaves of the treated seedlings were harvested from approximately 5 cm above the soil surface. The collected samples from each replicate were divided into three portions, rapidly frozen in liquid nitrogen, and stored at -80 °C. These samples were subsequently used for RNA extraction, metabolite quantification, and biochemical analyses.
RNA extraction and quantitative realtime PCR (qRT-PCR) analysis
Treated leaf tissue (100 mg) was finely ground using a mortar and pestle in liquid nitrogen for RNA extraction. Total RNA of the samples was extracted using p-BIOZOL (BioEquip) according to the manufacturer’s instructions. The quality of RNAs was measured using by NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific Inc, USA) and 1% agarose gel electrophoresis. Subsequently, 2 µg of high-quality RNA was treated with DNase I (Fermentas, USA), and the Easy cDNA Ultra-TM Synthesis Kit (Parstous, Iran) synthesized the first strand of cDNA. The coding sequences of the target genes (MhMYB1, MhMYB2, PR, MDH, MFS, and ACT) were retrieved from the National Center for Biotechnology Information (NCBI). Primers were designed using Primer3 and PrimerQuest™ online tools, and their quality parameters were subsequently evaluated using OligoAnalyzer™ and Primer-BLAST tools. The primer sequences and details are listed in Table 1. The actin gene employed as an internal standard. To assess the relative expression of target genes, qRT-PCR was performed using a 2X SYBR Green master mix (Parstous, Iran) in a final reaction volume of 20 µL. The reaction mixture consisted of 4 µL diluted cDNA, 0.5 µL of each primer, and 10 µL of 2X SYBR Green master mix. The qRT-PCR protocol included an initial denaturation step at 94 °C for 5 min (1 cycle), followed by 40 amplification cycles comprising denaturation at 94 °C for 15 s, annealing at primer-specific temperatures for 30 s, and extension at 72 °C for 30 s. qRT-PCR was performed using the Rotor-Gene Q system (Qiagen, Germany) with three technical replicates to determine PCR efficiency, cycle threshold (Ct) values, and melting curve profiles. Relative gene expression was analyzed using the REST 2009 software based on the Pfaffl method for accurate normalization and quantification which uses35
$$:text{R}text{a}text{t}text{i}text{o}=:frac{{left(text{E}text{f}text{f}text{i}text{c}text{i}text{e}text{n}text{c}text{y}:text{o}text{f}:text{t}text{a}text{r}text{g}text{e}text{t}:text{g}text{e}text{n}text{e}right)}^{varDelta:text{c}text{p}:text{o}text{f}:text{t}text{a}text{r}text{g}text{e}text{t}:text{g}text{e}text{n}text{e}}}{{left(text{E}text{f}text{f}text{i}text{c}text{i}text{e}text{c}text{y}:text{o}text{f}:text{r}text{e}text{f}text{e}text{r}text{e}text{n}text{c}text{e}:text{g}text{e}text{n}text{e}right)}^{varDelta:text{c}text{p}:text{r}text{e}text{f}text{e}text{r}text{e}text{n}text{c}text{e}:text{g}text{e}text{n}text{e}}}.$$
Essential oil isolation and gas chromatography analysis
The treated and shade-dried leaf tissue (5 g) was finely powdered to enhance surface area and subsequently subjected to essential oil extraction via the hydro-distillation method. The distillation process was conducted for 3 h using a Clevenger-type apparatus to ensure optimal separation and recovery of volatile compounds17. The collected essential oils were dried by anhydrous sodium sulfate (Na2SO4) to eliminate all traces of water, then stored in a sealed dark vial at 4 °C until analyzed. The identification of essential oil components was performed using a gas chromatography–mass spectrometry (GC-MS) system consisting of an Agilent 7890 gas chromatograph coupled with an Agilent 5975 mass selective detector (Agilent Technologies, USA). Separation was carried out on an HP-5 capillary column (30 m × 0.25 mm i.d., 0.25 μm film thickness). The oven temperature was programmed as follows: the initial temperature was set at 40 °C, then increased at a rate of 5 °C/min to 200 °C, held for 1 min, and then further increased at 10 °C/min to 250 °C and held for 5 min. Helium was used as the carrier gas at a constant flow rate of 1 mL/min and the split ratio was 30:1. The injection volume was 1 µL, and the injector temperature was maintained at 250 °C. The mass spectrometer operated in electron ionization mode with an energy of 70 eV, and the mass scan range was from 40 to 550 m/z. The identification of essential oil components was carried out using the retention indices and mass spectral analysis. The linear retention indices were calculated using the Van Den Dool and Kratz Eq. 36. Simultaneously, mass spectra were matched with reference libraries National Institute of Standards and Technology library (NIST) for compound identification37. The relative percentage of each compound was determined based on the chromatographic peak area in the total ion chromatogram (Fig. 3).

GC-MS chromatogram of the essential oil extracted from M. piperita flowering seedlings treated with 0.4 mM MJ via foliar application. Characteristic peaks corresponding to menthone (a), menthofuran (b), menthol (c), and pulegone (d) are indicated.
Quantification of hydrogen peroxide (H₂O₂) accumulation
To assess H₂O₂ content, 500 mg of treated leaf tissue was homogenized in an ice-cooled bath using 5 mL of trichloroacetic acid (TCA, 0.1% w/v). The homogenate was then centrifuged at 12,000 rpm for 15 min. Subsequently, 0.5 mL of 10 mM potassium phosphate buffer (pH 7.0) and 1 mL of 1 M potassium iodide (KI) were added to 0.5 mL of the supernatant. The absorbance of the samples was measured at 390 nm. To quantify the H₂O₂ content in the samples, a standard calibration curve was established using different concentrations of H₂O₂, and the results were expressed as µmol/g fresh weight (FW)38.
Biochemical analysis of catalase (CAT), ascorbate peroxidase (APX), and Guaiacol peroxidase (GPX) activities
Total soluble protein was quantified following the Bradford method39. For enzymatic extraction, 500 mg of treated tissue was thoroughly ground in liquid nitrogen and subjected to extraction at 4 °C. The homogenization was carried out in an extraction buffer (Tris–HCl, pH 7.8, containing 10% glycerol), ensuring optimal protein stability. The homogenate was then centrifuged at 15,000 rpm for 15 min at 4 °C, and the clarified supernatant was collected for enzymatic assays of CAT, GPX, and APX.
CAT activity was assessed by measuring the decomposition rate of H₂O₂. The reaction mixture consisted of 3 mL of phosphate buffer (pH 7.0), 5 µL of 30% H₂O₂, and 50 µL of crude enzyme extract. The decline in absorbance at 240 nm was recorded over a period of 1 min to quantify CAT activity, which was expressed as µmol of H2O2 decomposed/ (min mg protein)40.
APX activity was evaluated by monitoring the oxidation of ascorbate. The reaction mixture was composed of 100 µL of crude enzyme extract, 600 µL of 0.1 mM EDTA dissolved in 1.5 mL of 0.05 M potassium phosphate buffer (pH 7.0), and 400 µL of 0.5 mM ascorbic acid. The reaction was initiated by adding 400 µL of 30% H₂O₂. The decrease in ascorbate concentration was recorded at 290 nm over a 4 min period. APX activity was defined as the amount of ascorbate oxidized (µmol) / (min mg protein)41.
To measure GPX activity, 50 µL of crude enzyme extract was added to a reaction mixture containing 3 µL of guaiacol, 10 µL of 30% H₂O₂, and 3 mL of sodium phosphate buffer (pH 7.0). Changes in absorbance at 470 nm were recorded over a 1 min period. The activity of GPX was expressed as the amount of guaiacol oxidized (µmol) / (min mg protein)42.
Statistical analysis
Statistical analysis of the experimental data was performed using SAS 9.3.1 Portable. Analysis of variance (ANOVA) was conducted based on a Completely Randomized Design (CRD) with three replications. Mean comparisons were carried out using Duncan’s Multiple Range Test (DMRT) at a significance level of P < 0.05. To determine the optimal treatments for gene expression levels and monoterpene accumulation, hierarchical cluster analysis (HCA) was applied, and the results were visualized through heatmap representation using the ClustVis web-tool43. Additionally, correlations between gene expression levels, monoterpene content, and antioxidant enzymes were assessed utilizing the corrplot function in the RStudio environment (R 4.4.3)44.
Results
H2O2 content and antioxidant enzyme activity
Based on the results, each of the applied phytohormones exerted distinct effects on H2O2 levels. Among them, MJ and ABA exhibited more pronounced alterations, as their lowest applied concentrations (0.2 mM) led to a slight reduction in H2O2 levels, whereas their highest concentrations (0.4 mM) resulted in maximal production and accumulation of H2O2. In contrast, SA and GA demonstrated a divergent pattern compared to the former two phytohormones, wherein increasing their concentrations was associated with a moderate decline in H2O2 levels (Fig. 4A).
The activity of CAT increased in response to treatments with MJ, ABA, SA, and GA at concentrations of 0.2 and 0.4 mM. The highest CAT activity was observed in the ABA treatment at 0.4 mM, which was significantly greater than that of other treatments. MJ also induced a notable increase in CAT activity, albeit to a lesser extent than ABA. In contrast, SA and GA exhibited weaker effects on CAT activity, although a moderate increase was detected at the higher concentration (0.4 mM). Notably, GA at both concentrations and SA at 0.2 mM did not show a significant difference compared to the control group. The lowest CAT activity was recorded in the control group, showing a significant difference from most phytohormones treatments. Overall, an increase in phytohormones concentration was associated with a progressive rise in CAT activity (Fig. 4B).
The activity of APX was assessed in response to treatments with MJ, ABA, SA, and GA at concentrations of 0.2 and 0.4 mM. The highest APX activity was observed in MJ at 0.4 mM, showing a significant increase compared to other treatments. ABA also enhanced APX activity considerably, although to a lesser extent than MJ, with no substantial difference between the two tested concentrations. Notably, SA at 0.2 mM exhibited the lowest APX activity among all treatments, displaying a significant difference from the other groups; however, its activity markedly increased at 0.4 mM. GA treatment led to a moderate elevation in APX activity, with a slight increase at the higher concentration. The lowest APX activity was recorded in the control and SA at 0.2 mM, both significantly lower than other treatments. Overall, increasing phytohormones concentrations, particularly MJ and SA, resulted in a progressive rise in APX activity (Fig. 4C).
The activity of GPX significantly increased in response to most hormonal treatments. The highest GPX activity was observed in the MJ treatment at a concentration of 0.4 mM, which showed a significant difference compared to other treatments. The ABA and SA treatments also led to a notable increase in GPX activity, with their highest concentrations reaching levels comparable to MJ. In contrast, different concentrations of GA did not exhibit a significant difference compared to the control. Overall, increasing the concentration from 0.2 to 0.4 mM resulted in a rise in GPX activity across all treatments, highlighting the regulatory role of these phytohormones in the plant’s antioxidant response (Fig. 4D).

The effects of foliar application of different concentrations of MJ, ABA, SA, and GA phytohormones (0, 0.2, and 0.4 mM) on the H₂O₂ production (a) and the activity of antioxidant enzymes CAT (b), APX (c), and GPX (d) in flowering peppermint plants. Values are means of three replicates. Error bars represent ± SE. Different letters above the columns represent statistically significant differences at P < 0.05 (Duncan’s multiple range test).
Chemical composition of peppermint essential oil
Hormonal treatments exerted diverse effects on the chemical profile of peppermint essential oil. Among the various constituents, the menthol content was particularly responsive, showing significant fluctuations in response to both phytohormone type and concentration.
Treatment with MJ at both 0.2 and 0.4 mM concentrations significantly increased menthol content compared to the control, with the highest accumulation observed in the 0.4 mM MJ treatment. This result highlights the potential of MJ in stimulating menthol biosynthesis and enhancing essential oil quality. SA showed a significant effect only at the 0.4 mM concentration, indicating a concentration-dependent response. In contrast, ABA did not significantly alter menthol content at 0.2 mM, but caused a notable reduction at 0.4 mM, suggesting possible inhibitory effects on menthol-related biosynthetic pathways. GA exhibited no significant influence on menthol levels at either concentration (Fig. 5).

The effects of foliar application of different concentrations of MJ, ABA, SA, and GA phytohormones (0, 0.2, and 0.4 mM) on the percentage composition of menthol, menthone, menthofuran, and pulegone in the essential oil of flowering peppermint plants. Values are means of three replicates. Error bars represent ± SE. Different letters above the columns represent statistically significant differences at P < 0.05 (Duncan’s multiple range test).
Application of MJ at 0.4 mM significantly enhanced menthone accumulation compared to the control, while the lower concentration (0.2 mM) showed no statistically significant effect. ABA at 0.2 mM promoted menthone biosynthesis; however, its effect was diminished at 0.4 mM, with no significant difference from the control group. SA significantly increased menthone levels at both concentrations, whereas GA had no discernible impact. Among the treatments, MJ at 0.4 mM exhibited the most pronounced stimulatory effect on menthone production. Conversely, the reduced effect of ABA at the higher concentration may suggest a redirection of metabolic fluxes away from menthone biosynthesis (Fig. 5).
The results revealed that most treatments did not cause statistically significant changes in menthofuran content. However, MJ and SA at a concentration of 0.4 mM significantly reduced its accumulation, while no notable effects were observed with ABA or GA at either concentration. This pattern suggests that MJ and SA lead to a decrease in menthofuran content at elevated concentration (Fig. 5).
The results indicate that changes in pulegone content were less pronounced compared to the other three metabolites in response to the hormonal treatments. Most treatments had no significant effect on pulegone levels. The only statistically significant reduction was observed with SA at the 0.4 mM concentration, suggesting a possible downregulatory effect at higher doses (Fig. 5).
Differential expression of menthol pathway-related regulatory and biosynthetic genes
The relative expression of MhMYB1 was markedly influenced by hormonal treatments. MJ and SA significantly upregulated the expression of this transcription factor at both concentrations, with the most pronounced induction observed at 0.4 mM. In contrast, ABA exhibited a biphasic behavior; at the lower concentration (0.2 mM), it led to a moderate upregulation, whereas at 0.4 mM, it caused a substantial downregulation of MhMYB1 expression. Furthermore, GA induced gene expression at both concentrations, albeit to a lesser extent compared to MJ and SA. These expression patterns highlight the stimulatory role of MJ, SA, and GA in regulating MhMYB1 expression, while ABA at higher concentrations exerts a repressive effect on its transcription (Fig. 6).

The effects of foliar application of different concentrations of MJ, ABA, SA, and GA phytohormones (0, 0.2, and 0.4 mM) on the relative expression of genes involved in the menthol biosynthesis pathway. Values are means of three replicates. Error bars represent ± SE. Different letters above the columns represent statistically significant differences at P < 0.05 (Duncan’s multiple range test).
The relative expression of the MhMYB2 gene under hormonal treatments exhibited distinct regulatory patterns. MJ and GA followed a similar pattern in increasing gene expression, where an increase in the concentration of these phytohormones led to a greater upregulation of the gene, with the highest induction observed at a concentration of 0.4 mM. In contrast, ABA and SA also induced the expression of the gene, but this increase was concentration-independent; the lower concentration (0.2 mM) resulted in a higher increase in expression compared to the higher concentration (0.4 mM). These findings suggest that MJ and GA act as concentration-dependent positive regulators of MhMYB2 expression, whereas ABA and SA, despite their inducing effect, exhibited reduced effectiveness at higher concentrations. A noteworthy point is that the MJ treatment, at its highest concentration, had a more significant impact on the relative expression of this gene compared to the other treatments (Fig. 6).
Based on the results, the lowest concentrations (0.2 mM) of each of the four phytohormones (MJ, ABA, SA, and GA) led to a slight increase in the expression of the PR gene. However, increasing the concentration of ABA resulted in a reduction in transcript accumulation compared to the control. In contrast, higher concentrations (0.4 mM) of the other three phytohormones consistently enhanced PR gene expression, with a marked increase in transcript accumulation. A comparison of the treatments revealed that the highest concentrations of MJ and SA had the greatest impact on the accumulation of the PR gene transcript (Fig. 6).
The expression of the MDH gene in peppermint, when treated with varying concentrations of MJ and SA, displayed a significantly different and distinctive pattern compared to the effects observed with the other two phytohormones, ABA and GA. Both MJ and SA at their respective concentrations triggered a clear upregulation in gene expression, with the most pronounced induction observed at the highest concentrations of these two phytohormones. This suggests that MJ and SA play a critical role in enhancing the expression of the MDH gene, which could be linked to their known involvement in secondary metabolite biosynthesis pathways. In contrast, ABA and GA appeared to exert an inhibitory effect on the expression of the MDH gene, leading to a significant reduction in its relative expression levels. This suppression indicates that these phytohormones may have a negative regulatory impact on the expression of genes involved in essential oil biosynthesis, particularly in relation to menthol production (Fig. 6).
The expression of the MFS gene, in comparison to other genes of the menthol biosynthetic pathway, exhibited a somewhat different pattern under phytohormonal treatments. Specifically, a decreasing trend in the expression of this gene was observed under most treatments compared to the control, and except for the ABA treatments and the GA treatment at the 0.4 mM concentration, the relative expression in other treatments was negative (Fig. 6).
Cluster analysis of biochemical and molecular traits under different phytohormone treatments
The cluster analysis of hormonal treatments indicates that MJ (at concentrations of 0.2 and 0.4 mM) and SA (at 0.4 mM) exhibit similar effects, grouping into a distinct cluster. This clustering is likely due to the activation of shared oxidative signaling and metabolic pathways. Conversely, ABA (at 0.2 mM) and GA (at 0.4 mM) also clustered together, which may reflect similar regulatory effects of these two phytohormones. Interestingly, GA and SA at 0.2 mM closely resemble the control, indicating that at this concentration, these phytohormones exert no significant impact on the studied biosynthetic pathway at the molecular and biochemical levels. These findings suggest that at higher concentrations, MJ and SA activate oxidative stress-related pathways and signaling cascades leading to the induction of the menthol biosynthetic pathway. Notably, ABA at 0.4 mM did not cluster with any other treatment, implying that at this concentration, it exhibits a distinctly different regulatory behavior in modulating the studied biosynthetic pathway compared to the other hormonal treatments (Fig. 7).

Heatmap clustering of different phytohormone treatments (MJ, ABA, SA, and GA) at three concentrations (0, 0.2, and 0.4 mM), based on the expression patterns of genes involved in menthol biosynthesis, the relative contents of menthol, menthone, menthofuran, and pulegone, as well as oxidative stress markers in peppermint. Both rows and columns were clustered using maximum distance and average linkage methods.
The antioxidant enzymes (CAT, APX, and GPX) clustered together, indicating a coordinated response to oxidative stress induced by hormonal treatments. Since these enzymes play a crucial role in mitigating oxidative stress by scavenging H2O2, their proximity within this cluster suggests that the applied treatments have enhanced the plant’s antioxidant defense system. Given that H2O2 is grouped in a separate cluster, it can be inferred that its levels are regulated by these enzymes, establishing a coordinated balance in response to the hormonal treatments (Fig. 7).
Another distinct cluster was formed by the menthol and menthone metabolites. The clustering of these two metabolites suggests a similar production pattern under hormonal treatments. Given that both menthol and menthone are key components of the monoterpene biosynthetic pathway in aromatic plants and function as precursor and product within this pathway, their co-clustering was predictable (Fig. 7).
The clustering of MhMYB1, with MDH and PR genes suggests a functional association within a regulatory-metabolic pathway. This grouping may indicate that MhMYB1 potentially regulates the expression of MDH and PR under hormonal treatments, thereby coordinating their involvement in the menthol biosynthetic pathway in peppermint (Fig. 7).
Correlations between gene expression, metabolite levels, and oxidative stress factors
To gain deeper insights into menthol biosynthesis, we calculated the correlations between the expression levels of five selected genes (MhMYB1, MhMYB2, PR, MDH, and MFS), the quantities of pulegone, menthofuran, menthone, and menthol, and oxidative stress factors (H2O2, CAT, APX, and GPX) (Fig. 8). Among the 78 correlations obtained from the correlation matrix, 10 positive and prominent correlations were identified. Among these, two strong correlations were observed between antioxidant enzymes APX and CAT (ρ = 0.86, P < 0.01) and GPX and CAT (ρ = 0.82, P < 0.01). Three correlations were found between the studied genes, including MDH and MhMYB1 (ρ = 0.82, P < 0.01), PR and MDH (ρ = 0.83, P < 0.01), and PR and MhMYB1 (ρ = 0.97, P < 0.01). Additionally, five correlations were identified between genes and metabolites in the menthol biosynthesis pathway, including MhMYB1 and menthol (ρ = 0.92, P < 0.01), MhMYB1 and menthone (ρ = 0.81, P < 0.01), PR and menthol (ρ = 0.93, P < 0.01), PR and menthone (ρ = 0.86, P < 0.01), and MFS and menthofuran (ρ = 0.86, P < 0.01). Furthermore, six negative and notable correlations were observed, which included menthofuran and menthol (ρ = -0.88, P < 0.01), MhMYB1 and menthofuran (ρ = -0.94, P < 0.01), PR and menthofuran (ρ = -0.96, P < 0.01), MDH and menthofuran (ρ = -0.91, P < 0.01), MFS and menthol (ρ = -0.84, P < 0.01), and MFS and MDH (ρ = -0.96, P < 0.01) (Fig. 8).

A corrplot-based graph was used to visually interpret the potential pairwise correlations among the relative expression levels of genes involved in the menthol biosynthesis pathway, the contents of menthol, menthone, menthofuran, and pulegone, as well as oxidative stress markers (H₂O₂, CAT, APX, and GPX). The size of each circle and the shading volume of the colors are equivalent to the association degrees between each pairwise comparison.
Discussion
Oxidative stress in plant cells occurs as a result of adverse environmental conditions, including various biotic and abiotic stresses. In response to such conditions, plant cells employ a diverse range of defense mechanisms to mitigate the detrimental effects of oxidative stress and counteract the induced stress conditions. Among the most crucial strategies are the upregulation of antioxidant enzyme activities, stimulation of non-enzymatic antioxidant production, and enhancement of the biosynthesis and accumulation of protective compounds such as secondary metabolites45. Therefore, a precise understanding of the mechanisms underlying stress induction and the cellular processes involved in secondary metabolite production can facilitate the optimization of valuable plant metabolite synthesis in plant cells.
One of the most effective approaches for investigating oxidative stress induced by environmental stress factors is the analysis of ROS production and the assessment of both enzymatic and non-enzymatic antioxidant defense systems in plant cells46. Notably, the generation of ROS, particularly H₂O₂, in plant cells is widely recognized as a hallmark of oxidative stress, which subsequently triggers the activation of antioxidant defense mechanisms, enabling cells to mount an adaptive and regulated response under unfavorable environmental conditions47. The findings of this study indicate that foliar application of phytohormones can induce a diverse and dynamic pattern of oxidative stress manifestation and enzymatic antioxidant responses in M. piperita. This pattern is highly variable and appears to be strongly dependent on both the type of phytohormone applied and its concentration. According to the results, ABA and MJ induced a sustained increase in H₂O₂ production and accumulation in plant cells from the final foliar application until the time of sampling, suggesting a prolonged physiological stress perception. In terms of antioxidant responses, all three phytohormones, except GA, significantly enhanced the activity levels of CAT, APX, and GPX compared to the control. Notably, this enzymatic activity increase was concentration-dependent, with a progressive and measurable rise observed up to a concentration of 0.4 mM, indicating a dose-responsive regulatory mechanism.
Numerous studies have demonstrated that the application of plant hormones is closely associated with increased oxidative stress and enhanced antioxidant activity in plant cells. Notable examples include the upregulation of CAT activity induced by ABA in Orthosiphon stamineus seedlings48as well as the accumulation of H₂O₂ accompanied by elevated CAT and APX activity in Zea mays L. seedlings treated with ABA49. Furthermore, GA treatment has been shown to enhance CAT and APX activities in Spinacia oleracea L. seedlings50reinforcing the link between hormone signaling and antioxidant response. In addition, H₂O₂ accumulation along with increased CAT, APX, and GPX activity has been observed in Ricinus communis leaves following MJ application51. Similarly, foliar application of SA and MJ has been reported to significantly elevate CAT, APX, superoxide dismutase (SOD), and peroxidase (POD) activity in M. piperita seedlings33,52suggesting that such phytohormones can modulate oxidative balance through the activation of enzymatic defense pathways.
Mounting evidence from studies on the influence of elicitors in plant cell cultures suggests that oxidative stress plays a fundamental and indispensable role in the regulation of secondary metabolite biosynthesis. These studies clearly demonstrate that oxidative stress is not merely a byproduct of elicitation, but rather an integral component of the signaling cascade that activates biosynthetic pathways. The findings indicate that effective elicitors exert at least part of their influence on stimulating secondary metabolite production through the deliberate induction of oxidative stress. Specifically, the treatment of Papaver orientale cells with copper oxide nanoparticles resulted in increased biosynthesis and accumulation of alkaloids, accompanied by oxidative stress induction53. Similarly, the exposure of Satureja khuzestanica callus cultures to carbon-based nanoparticles led to enhanced accumulation of phenolic and flavonoid compounds, along with notable signs of oxidative stress54. Furthermore, the application of fungal elicitors to Taxus chinensis cell cultures stimulated taxol biosynthesis while simultaneously triggering oxidative stress, indicating a concurrent activation of both defense and metabolic pathways55. To further support these findings, it is also noteworthy that several studies have demonstrated that oxidative stress elements, even in isolation and in the absence of any specific elicitor, can initiate signal transduction processes leading to the production and accumulation of secondary metabolites. For instance, exogenous application of H₂O₂ has been reported to enhance rosmarinic acid synthesis in Salvia miltiorrhiza cell cultures56increase anthocyanin content in Daucus carota L. hairy root cultures57and improve the quantitative yield of key metabolites in Ginkgo biloba L. leaves following H₂O₂ treatment58.
Therefore, it is evident that phytohormones, in addition to their direct influence on the expression of genes responsible for the biosynthesis of secondary metabolites19also affect their biosynthesis and accumulation indirectly by simulating stress conditions and inducing oxidative stress. In light of these considerations, plant hormones play a critical role in regulating biological pathways, particularly those involved in the biosynthesis of secondary metabolites, through complex and diverse signaling networks. Thus, identifying these pathways, as well as their interactions and cross-talk, is essential for understanding plant cellular responses to various environmental stimuli. Furthermore, elucidating these interactions is crucial for improving the metabolic efficiency of plants, ultimately optimizing the production of valuable secondary metabolites with significant pharmaceutical and industrial applications.
Based on the findings of this study, it is evident that gene expression, as the examined variable, exhibits greater dynamism under phytohormone treatments compared to other assessed factors. The results indicate that the phytohormones MJ and SA, followed by GA, have a more prominent role in inducing the expression of regulatory and biosynthetic genes involved in menthol biosynthesis (MhMYB1, MhMYB2, PR, and MDH) compared to ABA. This suggests that MJ and SA are particularly effective in enhancing the transcription of these genes, while GA also plays a significant role in this process. In contrast, ABA treatment was found to be significantly more effective in enhancing the accumulation of transcripts from the MFS gene, which encodes the key enzyme responsible for menthofuran biosynthesis.
Although research on the simultaneous application of multiple phytohormonal elicitors to induce secondary metabolite biosynthetic pathways, particularly the menthol biosynthetic pathway, is limited, several studies have reported transcriptional changes in key genes involved in menthol biosynthesis following specific phytohormone treatments. Specifically, treatments with SA and MJ have been associated with the upregulation of the PR and MDH genes, while the MFS gene, which encodes the key enzyme for menthofuran biosynthesis, is downregulated33,52,59. In contrast, GA treatment has been linked to an increase in PR expression, while MDH and MFS gene expressions are significantly reduced59.
In this study, the expression of two MYB family transcription factors (MhMYB1 and MhMYB2), which play a regulatory role in the menthol biosynthetic pathway, was examined under various phytohormone treatments. Transcription factors serve as intermediaries, linking signal perception and transduction to the regulatory network that controls gene expression. Thus, analyzing their expression patterns provides valuable insights into the processes of signal reception, transduction, and the downstream effects of signaling, as well as the interactions among key regulatory elements involved in these processes.
MYB transcription factors play a pivotal role in sensing a wide range of upstream signals and relaying them to the gene regulatory network. Previous studies have shown that members of this transcription factor family are involved in the activation of phytohormonal signaling transduction pathways, particularly those associated with SA, ABA, GA, and MJ60,61,62,63. In addition, they contribute to plant responses to oxidative stress induced by reactive oxygen species (ROS), primarily through the reprogramming of stress-responsive gene expression patterns64.
The findings of this study indicate that the expression of the transcription factors MhMYB1 and MhMYB2 is influenced by the applied phytohormones, the oxidative stress induced by these treatments, or a synergistic effect of both phytohormonal signaling and ROS-related stress. This regulatory influence appears to positively modulate downstream genes, as the upregulation of MhMYB1 and MhMYB2 was accompanied by increased expression of MDH and PR, two key genes in the menthol biosynthetic pathway. These observations highlight a strong positive correlation between the expression levels of the MYB transcription factors and the target genes MDH and PR, suggesting a potential transcriptional control mechanism (Fig. 8).
Analysis of essential oil composition in phytohormone-treated samples revealed that MJ elicited the most substantial increase in the accumulation of menthol and menthone, suggesting its strong stimulatory effect on the corresponding biosynthetic pathways. SA also promoted the production of these metabolites, though its impact was comparatively moderate. In contrast, GA treatment did not result in any significant changes in the levels of either compound. Notably, ABA exhibited a divergent effect, as its highest concentration was associated with a marked reduction in both menthol and menthone content.
The enhanced biosynthesis of menthol and menthone appears to be closely associated with the upregulation of transcription factors MhMYB1 and MhMYB2, along with increased expression of the genes encoding PR and MDH enzymes. Correlation analysis reinforces this association, demonstrating a strong positive relationship between the accumulation of these monoterpenes and the transcriptional activity of the aforementioned genes (Fig. 8). Interestingly, treatment with MJ and SA at their highest concentrations resulted in a significant decrease in menthofuran levels. This observation suggests a possible metabolic redirection within the monoterpene biosynthetic pathway, favoring menthol and menthone production. It is likely that the precursor pulegone, which is also a substrate for PR, is preferentially channeled toward menthol and menthone synthesis under these conditions, thereby limiting its conversion into menthofuran.
Among the analyzed monoterpenes, pulegone displayed the least quantitative variation across different phytohormonal treatments. Within the menthol biosynthetic pathway, pulegone serves a dual role as both a biosynthetic intermediate and a product, being synthesized by the iPI enzyme and subsequently converted into downstream metabolites by the PR enzyme. The observed stability in pulegone levels can be attributed to a dynamic equilibrium between its production and consumption, which likely maintains its relatively constant concentration despite external elicitor stimuli.
Changes in both the quantitative and qualitative profiles of essential oils in peppermint have been reported in several previous studies under the influence of phytohormonal elicitors. These studies emphasize that foliar spraying of MJ within the concentration range of 0.1 to 2 mM leads to a substantial increase in menthol and menthone levels, while concurrently decreasing the amount of menthofuran34,65. Additional reports confirm the stimulatory role of MJ in enhancing menthol accumulation66,67and boosting the biosynthesis of menthone68. Likewise, SA has shown comparable effects, where effective concentrations ranging from 0.1 to 2.5 mM have been linked with elevated menthol and menthone levels, along with a noticeable reduction in menthofuran content69. Furthermore, similar trends have been observed in other studies, supporting that SA treatment promotes menthol and menthone biosynthesis while repressing menthofuran production33,70thereby reinforcing the regulatory role of these phytohormones in monoterpene metabolism. In contrast to the well-established effects of MJ and SA, studies examining the role of GA and ABA in regulating the menthol biosynthetic pathway at both the transcriptional and metabolic levels are limited. However, available evidence from a few reports indicates that foliar application of GA at concentrations ranging from 0.02 to 0.15 mM can result in a modest increase in menthol production in peppermint seedlings71,72. These observations imply that GA may exert a regulatory influence, although its effectiveness appears to be relatively low compared to MJ and SA. Interestingly, emerging evidence suggests that the elicitation efficiency of MJ can be further enhanced through synergistic combinations with other elicitors. For instance, a recent investigation demonstrated that the co-application of MJ with titanium dioxide (TiO₂) nanoparticles resulted in significantly higher menthol accumulation in peppermint compared to MJ alone73. This synergistic effect may stem from the combined action of MJ-induced transcriptional activation of biosynthetic genes and TiO₂ nanoparticle-mediated enhancement of oxidative signaling, thereby amplifying the elicitation response. Such findings underline the potential of combinatorial elicitation strategies for optimizing secondary metabolite production in medicinal and aromatic plants.
A key finding of this study is that ABA exhibited a distinct regulatory effect compared to the other three phytohormones by channeling the biosynthetic pathway toward menthofuran. This was evidenced by the upregulation of the MFS gene and the corresponding increase in menthofuran production. These results underscore the fact that phytohormonal elicitors, even when acting within the same metabolic pathway, can selectively modulate the expression of target genes and redirect metabolic flux toward the synthesis of specific secondary metabolites. Although ABA did not significantly enhance menthol biosynthesis in our study, recent evidence suggests that it plays an important regulatory role in the production of other secondary metabolites in plants. For instance, exogenous ABA application has been shown to upregulate key genes involved in flavonoid biosynthesis and promote the accumulation of flavonoid compounds in tea plant74. Similarly, Yang et al.75 reported that ABA treatment markedly increased flavonoid content in Cajanus cajan L. (pigeon pea). In another study, Zhou et al.76 demonstrated that ABA positively influenced the accumulation of phenolic acids in Rhododendron chrysanthum. These findings indicate that while ABA may not directly stimulate the menthol biosynthetic pathway, it can positively influence other branches of secondary metabolism, particularly those involved in flavonoid and phenolic compound accumulation.
The findings of this study demonstrate that both the type and concentration of the applied phytohormone significantly affect the evaluated parameters. Higher concentrations (0.4 mM) had a more notable influence than lower concentrations (0.2 mM) on antioxidant enzyme activity, gene expression at the transcriptional level, and the biosynthesis and accumulation of monoterpenes involved in menthol production. Moreover, the specific effects of each phytohormone were clearly distinguishable. Among them, MJ and SA had the most substantial impact on the upregulation of genes involved in menthol and menthone biosynthesis, which was reflected in the increased levels of these compounds. Cluster analysis further supported this observation by grouping MJ and SA treatments together in a distinct cluster, separate from other treatments.
The results suggest that, at identical concentrations, MJ and SA had relatively similar effects and were more effective than GA and ABA in modulating the studied parameters. At a concentration of 0.4 mM, both MJ and SA showed the highest efficacy in activating the menthol biosynthetic pathway and shifting the essential oil profile toward increased production of menthol and menthone. In contrast, GA had a limited influence on oxidative stress induction, gene expression, and menthol biosynthesis at both tested concentrations. These findings indicate that, within the evaluated concentration range, GA is not a potent elicitor for redirecting the menthol biosynthetic pathway.
Although the phytohormone ABA demonstrated a significant effect on oxidative stress induction, following MJ, it clearly suppressed the expression of genes involved in menthol biosynthesis. However, in contrast, by increasing the expression of the MFS gene, it directed the biosynthesis of monoterpenes in the menthol biosynthesis pathway towards menthofuran.
In addition to phytohormonal elicitors, recent studies have demonstrated that the menthol biosynthetic pathway in Mentha piperita can also be effectively stimulated by non-phytohormonal elicitors. For instance, the application of titanium dioxide (TiO₂) nanoparticles has been shown to enhance the expression of key genes involved in menthol biosynthesis, suggesting a potential role for abiotic elicitors in activating this metabolic route77. Likewise, biotic elicitors such as cyanobacteria have been reported to significantly upregulate gene expression within this pathway and promote the accumulation of menthone78. These findings imply that the regulation of menthol biosynthesis is not limited to phytohormonal signaling but can also be modulated by diverse elicitor types. Focusing future research on such elicitors—particularly in exploring their synergistic interactions with phytohormones—could provide deeper insights into the underlying regulatory mechanisms and offer promising strategies for optimizing menthol production in medicinal and aromatic plants.
Conclusion
This study provides valuable insights into the effects of four phytohormones (MJ, ABA, SA, and GA) at two concentrations (0.2 mM and 0.4 mM) on oxidative stress induction, antioxidant enzyme activity, gene expression, and the biosynthesis and accumulation of menthol and menthone in M. piperita. The results demonstrate that MJ and SA had the most pronounced effects on menthol biosynthesis, primarily by upregulating key genes (MhMYB1, MhMYB2, PR, and MDH) and enhancing menthol and menthone levels while simultaneously reducing menthofuran accumulation. These findings suggest that, particularly at 0.4 mM, MJ and SA effectively redirect metabolic flux toward menthol and menthone production. In contrast, ABA exhibited a distinct regulatory role by promoting menthofuran biosynthesis through upregulation of the MFS gene. Meanwhile, GA showed a comparatively lower influence on oxidative stress responses, gene expression, and monoterpene biosynthesis, indicating a limited role in modulating the menthol biosynthetic pathway under the tested conditions. Overall, this study highlights the complex and diverse regulatory roles of phytohormones in secondary metabolite biosynthesis. The differential effects of these elicitors underscore their potential utility in optimizing the production of valuable phytochemicals in M. piperita. Further research is warranted to explore the synergistic effects of combined phytohormone treatments and to assess their long-term influence on metabolic pathways. Additionally, investigating a broader range of concentrations may offer deeper insights into the regulation of biosynthetic processes. These findings lay a foundation for future biotechnological interventions aimed at enhancing the yield of medicinally important.
Data availability
The datasets generated and analyzed during the current study are available upon reasonable request from the corresponding author.
References
-
Velu, G., Palanichamy, V. & Rajan, A. P. Phytochemical and Pharmacological importance of plant secondary metabolites in modern medicine. Bioorg. Phase Nat. Food: Overv. 135–156. https://doi.org/10.1007/978-3-319-74210-6_8 (2018). Springer International Publishing.
-
Sarwar, S., Zia, M., Fatima, Z. & Ahmad Sial, R. Fayyaz chaudhary, M. In vitro direct regeneration in mint from different explants on half strength MS medium. Afr. J. Biotechnol. 8, 4667–4671 (2009).
-
Kim, Y. S., Choi, Y. E. & Sano, H. Plant vaccination: stimulation of defense system by caffeine production in planta. Plant. Signal. Behav. 5, 489–493 (2010).
-
Selmar, D. & Kleinwächter, M. Stress enhances the synthesis of secondary plant products: the impact of Stress-Related Over-Reduction on the accumulation of natural products. Plant. Cell. Physiol. 54, 817–826 (2013).
-
Talbot, S. C., Pandelova, I., Lange, B. M. & Vining, K. J. A first look at the genome structure of hexaploid Mitcham peppermint (Mentha × Piperita L). G3: Genes Genomes Genet. https://doi.org/10.1093/g3journal/jkae195 (2024).
-
Gobert, V., Moja, S., Colson, M. & Taberlet, P. Hybridization in the section Mentha (Lamiaceae) inferred from AFLP markers. Am. J. Bot. 89, 2017–2023 (2002).
-
Marcum, D. B. & Hanson, B. R. Effect of irrigation and harvest timing on peppermint oil yield in California. Agric. Water Manag. 82, 118–128 (2006).
-
Sahib, N. G. et al. Coriander (Coriandrum sativum L.): A potential source of High-Value components for functional foods and Nutraceuticals‐ A review. Phytother. Res. 27, 1439–1456 (2013).
-
İşcan, G. et al. Antimicrobial screening of Mentha Piperita essential oils. J. Agric. Food Chem. 50, 3943–3946 (2002).
-
Taherpour, A. A. et al. Chemical composition analysis of the essential oil of mentha Piperita L. from kermanshah, Iran by hydrodistillation and HS/SPME methods. J. Anal. Sci. Technol. 8, 11 (2017).
-
Peiris, M. et al. A putative anti-inflammatory role for TRPM8 in irritable bowel syndrome—An exploratory study. Neurogastroenterology & Motility 33, (2021).
-
Feitosa, K. A. et al. Menthol and menthone associated with acetylsalicylic acid and their relation to the hepatic fibrosis in schistosoma mansoni infected mice. Front Pharmacol 8, (2018).
-
Kamatou, G. P. P., Vermaak, I., Viljoen, A. M. & Lawrence, B. M. Menthol: A simple monoterpene with remarkable biological properties. Phytochemistry 96, 15–25 (2013).
-
Gershenzon, J., Maffei, M. & Croteau, R. Biochemical and histochemical localization of monoterpene biosynthesis in the glandular trichomes of spearmint (Mentha spicata). Plant. Physiol. 89, 1351–1357 (1989).
-
Croteau, R. B., Davis, E. M., Ringer, K. L. & Wildung, M. R. (-)-Menthol biosynthesis and molecular genetics. Naturwissenschaften vol. 92 562–577 Preprint at (2005). https://doi.org/10.1007/s00114-005-0055-0
-
An, X. et al. Effects of MhMYB1 and MhMYB2 transcription factors on the monoterpenoid biosynthesis pathway in l-menthol chemotype of mentha Haplocalyx Briq. Planta 260, 3 (2024).
-
Rahimi, Y., Taleei, A. & Ranjbar, M. Changes in the expression of key genes involved in the biosynthesis of menthol and menthofuran in mentha Piperita L. under drought stress. Acta Physiol. Plant 39, (2017).
-
Thakur, M., Bhattacharya, S., Khosla, P. K. & Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. Journal of Applied Research on Medicinal and Aromatic Plants vol. 12 1–12 Preprint at (2019). https://doi.org/10.1016/j.jarmap.2018.11.004
-
Lv, Z. Y. et al. Phytohormones jasmonic acid, salicylic acid, gibberellins, and abscisic acid are key mediators of plant secondary metabolites. World Journal of Traditional Chinese Medicine vol. 7 307–325 Preprint at (2021). https://doi.org/10.4103/wjtcm.wjtcm_20_21
-
Altaf, M. A. et al. Phytohormones mediated modulation of abiotic stress tolerance and potential crosstalk in horticultural crops. J. Plant. Growth Regul. 42, 4724–4750 (2023).
-
Mearaji, H. S., Ansari, A., Igdelou, N. K. M., Lajayer, B. A. & Pessarakli, M. Phytohormones and Abiotic Stresses. in Handbook of Plant and Crop Physiology 175–213 (CRC Press, 4th edition. | Boca Raton, FL: CRC Press, 2021). (2021). https://doi.org/10.1201/9781003093640-16
-
De Geyter, N., Gholami, A., Goormachtig, S. & Goossens, A. Transcriptional machineries in jasmonate-elicited plant secondary metabolism. Trends Plant. Sci. 17, 349–359 (2012).
-
Khan, M. I. R., Fatma, M., Per, T. S., Anjum, N. A. & Khan, N. A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front Plant. Sci 6, (2015).
-
Murcia, G. et al. ABA and GA3 regulate the synthesis of primary and secondary metabolites related to alleviation from biotic and abiotic stresses in grapevine. Phytochemistry 135, 34–52 (2017).
-
Edreva, A. et al. STRESS-PROTECTIVE ROLE OF SECONDARY METABOLITES: DIVERSITY OF FUNCTIONS AND MECHANISMS. (2008).
-
Paek, K. Y., Murthy, H. N., Hahn, E. J. & Zhong, J. J. Large Scale Culture of Ginseng Adventitious Roots for Production of Ginsenosides. in Biotechnology in China I 151–176Springer Berlin Heidelberg, Berlin, Heidelberg, (2009). https://doi.org/10.1007/10_2008_31
-
Bryant, J. P., Chapin, F. S. & Klein, D. R. Carbon/Nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 40, 357 (1983).
-
Murthy, H. N., Lee, E. J. & Paek, K. Y. Production of secondary metabolites from cell and organ cultures: strategies and approaches for biomass improvement and metabolite accumulation. Plant. Cell. Tissue Organ. Cult. (PCTOC). 118, 1–16 (2014).
-
Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant. Sci. 7, 405–410 (2002).
-
Hu, W. H. et al. Changes in electron transport, superoxide dismutase and ascorbate peroxidase isoenzymes in chloroplasts and mitochondria of cucumber leaves as influenced by chilling. Photosynthetica 46, (2008).
-
Ho, T. T., Murthy, H. N. & Park, S. Y. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. International Journal of Molecular Sciences vol. 21 Preprint at (2020). https://doi.org/10.3390/ijms21030716
-
Fazal, H. et al. Sustainable production of biomass and industrially important secondary metabolites in cell cultures of selfheal (Prunella vulgaris L.) elicited by silver and gold nanoparticles. Artif. Cells Nanomed. Biotechnol. 47, 2553–2561 (2019).
-
Jariani, P., Sabokdast, M., Moghadam, T. K., Nabati, F. & Dedicova, B. Modulation of Phytochemical Pathways and Antioxidant Activity in Peppermint by Salicylic Acid and GR24: A Molecular Approach. Cells 13, (2024).
-
Kandoudi, W., Tavaszi-Sárosi, S. & Németh-Zámboriné, E. Inducing the Production of Secondary Metabolites by Foliar Application of Methyl Jasmonate in Peppermint. Plants 12, (2023).
-
Pfaffl, M. W., Tichopad, A., Prgomet, C. & Neuvians, T. P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper – Excel-based tool using pair-wise correlations. Biotechnol. Lett. 26, 509–515 (2004).
-
van Den Dool, H. Dec. Kratz, P. A generalization of the retention index system including linear temperature programmed gas—liquid partition chromatography. J. Chromatogr. A. 11, 463–471 (1963).
-
Adams, R. P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometryvol. 456 (Allured Publishing Corporation, 2007).
-
Alexieva, V., Sergiev, I., Mapelli, S. & Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant. Cell. Environ. 24, 1337–1344 (2001).
-
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
-
Scebba, F., Sebastiani, L. & Vitagliano, C. Changes in activity of antioxidative enzymes in wheat (Triticum aestivum) seedlings under cold acclimation. Physiol. Plant. 104, 747–752 (1998).
-
Ranieri, A. Early production and scavenging of hydrogen peroxide in the Apoplast of sunflower plants exposed to Ozone. J. Exp. Bot. 54, 2529–2540 (2003).
-
Dionisio-Sese, M. L. & Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 135, 1–9 (1998).
-
Metsalu, T. & Vilo, J. ClustVis: a web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res. 43, W566–W570 (2015).
-
R Core Team. R: A Language and Environment for Statistical Computing. Preprint at. (2025).
-
Apel, K. & Hirt, H. REACTIVE OXYGEN SPECIES: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 55, 373–399 (2004).
-
Anjum, N. A., Gill, S. S., Duarte, A. C., Pereira, E. & Ahmad, I. Silver nanoparticles in soil–plant systems. J. Nanopart. Res. 15, 1896 (2013).
-
Low, P. S. & Merida, J. R. The oxidative burst in plant defense: function and signal transduction. Physiol. Plant. 96, 533–542 (1996).
-
Ibrahim, M. H. & Jaafar, H. Z. E. Abscisic acid induced changes in production of primary and secondary metabolites, photosynthetic capacity, antioxidant capability, antioxidant enzymes and Lipoxygenase inhibitory activity of orthosiphon stamineus Benth. Molecules 18, 7957–7976 (2013).
-
Jiang, M. & Zhang, J. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell. Physiol 42 (2001).
-
Gong, Q. et al. Gibberellic acid application on biomass, oxidative stress response, and photosynthesis in spinach (Spinacia Oleracea L.) seedlings under copper stress. Environ. Sci. Pollut. Res. 28, 53594–53604 (2021).
-
Soares, A. M., dos, S., de Souza, T. F., Jacinto, T. & Machado, O. L. T. Effect of Methyl jasmonate on antioxidative enzyme activities and on the contents of ROS and H2O2 in ricinus communis leaves. Braz. J. Plant. Physiol. 22, 151–158 (2010).
-
Afkar, S. & Karimzadeh, G. Changes in physiological traits, gene expression and phytochemical profile of mentha Piperita in response to elicitor. Biochem. Genet. 63, 2025–2039 (2025).
-
khaldari, I., Naghavi, M. R., Motamedi, E. & Zargar, M. The effects of green and chemically-synthesized copper oxide nanoparticles on the production and gene expression of morphinan alkaloids in Oriental poppy. Sci Rep 14, (2024).
-
Ghorbanpour, M. & Hadian, J. Multi-walled carbon nanotubes stimulate callus induction, secondary metabolites biosynthesis and antioxidant capacity in medicinal plant satureja Khuzestanica grown in vitro. Carbon N Y. 94, 749–759 (2015).
-
Yu, L. J., Lan, W. Z., Qin, W. M., Jin, W. W. & Xu, H. B. Oxidative stress and taxol production induced by fungal elicitor in cell suspension cultures of taxus chinensis. Biol. Plant. 45, 459–461 (2002).
-
Hao, W. et al. Hydrogen peroxide is involved in salicylic acid-elicited rosmarinic acid production in salvia miltiorrhiza cell cultures. Scientific World Journal (2014). (2014).
-
Barba-Espín, G. et al. H2O2-elicitation of black Carrot hairy roots induces a controlled oxidative burst leading to increasedanthocyanin production. Plants 10, (2021).
-
Fan, J. et al. Mechanism by which exogenous H2O2 improves main secondary metabolites contents in post-harvest fresh Ginkgo leaves through induced physiological responses mimicking stress. Ind Crops Prod 218, (2024).
-
Soleymani, F., Taheri, H. & Shafeinia, A. R. Relative expression of genes of menthol biosynthesis pathway in peppermint (Mentha Piperita L.) after chitosan, gibberellic acid and Methyl jasmonate treatments. Russ. J. Plant Physiol. 64, 59–66 (2017).
-
Raffaele, S., Rivas, S. & Roby, D. An essential role for Salicylic acid in AtMYB30-mediated control of the hypersensitive cell death program in Arabidopsis. FEBS Lett. 580, 3498–3504 (2006).
-
Abe, H. et al. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant. Cell. 15, 63–78 (2003).
-
Murray, F., Kalla, R., Jacobsen, J. & Gubler, F. A role for HvGAMYB in anther development. Plant J. 33, 481–491 (2003).
-
Lee, M. W., Qi, M. & Yang, Y. A. Novel jasmonic Acid-Inducible rice Myb gene associates with fungal infection and host cell death. Mol. Plant-Microbe Interactions®. 14, 527–535 (2001).
-
Liao, C., Zheng, Y. & Guo, Y. MYB30 transcription factor regulates oxidative and heat stress responses through ANNEXIN-mediated cytosolic calcium signaling in Arabidopsis. New Phytol. 216, 163–177 (2017).
-
Afkar, S., Karimzadeh, G., Jalali-Javaran, M., Sharifi, M. & Behmanesh, M. Journal of Medicinal Plants and By-Products () 1: 75–82 Influence of Methyl Jasmonate on Menthol Production and Gene Expression in Peppermint (Mentha x Piperita L.). (2013). http://www.ncbi.nlm.nih.gov/ (2013).
-
Abu El-Leel, O., Mohamed, S., Sukar, N., Abd, E. L., Aziz, M. & – Influence of Jasmine oil and Methyl jasmonate on gene expression and menthol production in mentha. Sci. J. Agricultural Sci. 0, 0–0 (2021).
-
Mehdizadeh, L., Moghaddam, M., Ganjeali, A. & Mahmoodi Sourestani, M. Changes in growth, essential oil composition and biochemical traits of peppermint in response to coapplication of zinc and Methyl jasmonate in soilless culture. J. Plant. Nutr. 47, 2732–2752 (2024).
-
Cappellari, L. D. R., Santoro, M. V., Schmidt, A., Gershenzon, J. & Banchio, E. Improving phenolic total content and monoterpene in mentha x Piperita by using Salicylic acid or Methyl jasmonate combined with rhizobacteria inoculation. Int J. Mol. Sci 21, (2020).
-
Motiee, M. & Abdoli, M. Changes in essential oil composition of peppermint (Mentha x Piperita L.) affected by yeast extract and Salicylic acid foliar application. J. Med. Plants. 20, 47–58 (2021).
-
Saharkhiz, M. J. & Goudarzi, T. Foliar application of Salicylic acid changes essential oil content and chemical compositions of peppermint (Mentha Piperita L). J. Essent. Oil-Bearing Plants. 17, 435–440 (2014).
-
Taheri, H. Transcriptional modulation of structural and regulatory genes involved in isoprene biosynthesis and their relevance to oil yield and menthol content in peppermint (Mentha Piperita L.) upon MeJA and GA3 treatments. Russ. J. Plant Physiol. 66, 503–508 (2019).
-
Ahmad, B., Jaleel, H., Shabbir, A., Khan, M. M. A. & Sadiq, Y. Concomitant application of depolymerized Chitosan and GA3 modulates photosynthesis, essential oil and menthol production in peppermint (Mentha Piperita L). Sci. Hortic. 246, 371–379 (2019).
-
Shenavaie Zare, A., Ganjeali, A., Vaezi Kakhki, M. R., Cheniany, M. & Mashreghi, M. Plant elicitation and TiO2 nanoparticles application as an effective strategy for improving the growth, biochemical properties, and essential oil of peppermint. Physiol. Mol. Biology Plants. 28, 1391–1406 (2022).
-
Gai, Z. et al. Exogenous abscisic acid induces the lipid and flavonoid metabolism of tea plants under drought stress. Sci Rep 10, (2020).
-
Yang, W. et al. Transcriptome analysis reveals abscisic acid enhancing drought resistance by regulating genes related to flavonoid metabolism in pigeon pea. Environ Exp. Bot 191, (2021).
-
Zhou, X. et al. Abscisic acid affects phenolic acid content to increase tolerance to UV-B stress in rhododendron Chrysanthum pall. Int J. Mol. Sci 25, (2024).
-
Veleshkolaii, F. R., Gerami, M., Younesi-Melerdi, E. & Moshaei, M. R. & Ghanbari Hassan kiadeh, S. Studying the impact of titanium dioxide nanoparticles on the expression of pivotal genes related to menthol biosynthesis and certain biochemical parameters in peppermint plants (Mentha Piperita L). BMC Plant. Biol 24, (2024).
-
Ghotbi-Ravandi, A. A. et al. Enhancement of essential oil production and expression of some menthol Biosynthesis-Related genes in mentha Piperita using cyanobacteria. Iran. J. Biotechnol. 21, 96–107 (2023).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Khaldari, I., Afshoon, E. & Nik, S.H. Phytohormonal elicitation triggers oxidative stress and enhances menthol biosynthesis through modulation of key pathway genes in Mentha piperita L.. Sci Rep 15, 30495 (2025). https://doi.org/10.1038/s41598-025-16667-1
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-16667-1