Introduction
According to the World Health Organization report 2002, probiotics are living organisms that confer health benefits to the host when administered in sufficient amounts1. Probiotic strains impart a wide range of health benefits, including modulation of the gut microbiota2, reduction of obesity, mitigation of inflammatory bowel diseases, inhibition of intestinal pathogens, regulation of immune responses, and reduction of lactose intolerance3. Probiotic strains are commonly isolated from diverse sources, including animals, dairy products, fermented foods, crops, and plants. Lactic acid bacteria (LAB), ubiquitously found in various environmental sources, are prominent contributors to the probiotic category. Commonly identified probiotic LABs include Lactobacillus spp. (e.g., Lactobacillus delbrueckii ssp. bulgaricus, Lactobacillus acidophilus, and Lactobacillus helveticus), Lacticaseibacillus spp. (e.g., Lacticaseibacillus casei, Lacticaseibacillus paracasei, and Lacticaseibacillus rhamnosus), and Bifidobacteria spp. (e.g., Bifidobacteria animalis, Bifidobacteria longum, Bifidobacteria breve, Bifidobacteria bifidum, and Bifidobacteria infantis)4,5. Additionally, other genera, including Leuconostoc, Streptococcus, Propionibacterium, Bacillus, Enterococcus, and Saccharomyces, have been recognized as probiotics4,5. The application of probiotics across food, healthcare, veterinary medicine, and pharmaceutical industries has led to substantial growth in the global market in recent years6,7, creating an increased demand for the isolation of novel probiotic strains8.
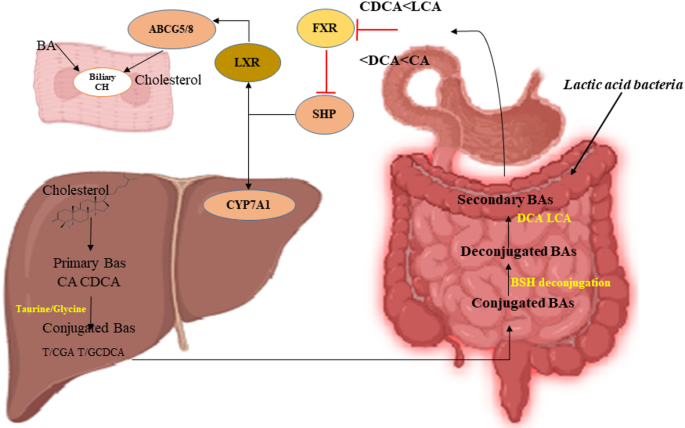
Cardiovascular disease, a global health concern with high mortality, is mainly driven by elevated cholesterol levels9. Previous studies have shown that even a 1% reduction in serum cholesterol levels can reduce the risk of coronary artery disease by 2–3%10. Therefore, pharmaceutical drugs have been widely explored to lower cholesterol levels; however, they often pose challenges such as side effects11,12 and high costs13,14. In contrast, probiotic supplementation offers a promising, cost-effective alternative13, particularly owing to its cholesterol-lowering properties. For instance, LAB contribute to cholesterol reduction through multiple mechanisms, including bile salt hydrolase (BSH) activity15, production of short-chain fatty acids16, conversion of cholesterol to coprostanol17, and direct assimilation of cholesterol18. This study specifically focused on the activity of BSH, which originates from the intestinal gut microbiota and occurs in the small intestine19. A schematic representation of BSH activity, which shows the mechanism of lowering cholesterol levels, is shown in Fig. 1.

A schematic representation of bile salt hydrolase (BSH) activity and its role in cholesterol reduction. Conjugated bile acids (BAs) with taurine or glycine are deconjugated by BSH-active gut lactic acid bacteria. This process affects several key pathways, including the farnesoid X receptor (FXR), small heterodimer partner (SHP), cholesterol 7 alpha-hydroxylase (CYP7A1), liver X receptor (LXR), and ABCG5/8 transporters, which mediate cholesterol excretion into the bile canaliculi.
Ginseng is a widely used medicinal plant cultivated across East Asia, including Korea20. Ginseng sprouts, particularly those cultivated under smart farming systems, offer a controlled and contaminant-limited environment, facilitating the isolation of unique and potentially beneficial LAB strains. Ginseng is well-documented for its bioactive phytochemicals, including ginsenosides, which may support the growth and metabolic adaptation of probiotic bacteria21. Screening for probiotics in this niche not only expands the diversity of functional strains but also leverages the synergistic health benefits of ginseng and probiotics, particularly in the context of cardiovascular and gastrointestinal health.
The goal of this study is to isolate and characterize Lacticaseibacillus rhamnosus strains from ginseng sprouts grown under smart-farming conditions, with a specific focus on evaluating their cholesterol-lowering effects via bile salt hydrolase (BSH) activity, alongside assessment of their probiotic properties and safety profile. Notably, our current study offers several innovative contributions to the field of probiotic-based cholesterol management. In contrast to conventional approaches, it focuses on L. rhamnosus strains isolated from ginseng sprouts grown in a smart-farming system, offering a sustainable and controlled source of novel probiotic candidates.
Results
Isolation of identification of bacteria from ginseng
A total of 485 bacterial colonies were isolated from ginseng sprouts. The BCP test identified 253 BCP-positive bacterial isolates, whereas the catalase assay identified 59 catalase-negative isolates (Table 1). Additionally, Gram staining of the isolates indicated that 35 were gram-positive and rod-shaped, whereas the remaining isolates (n = 450) were gram-negative (Table 1).
Molecular identification of 31 bacterial isolates that were BCP-positive, catalase-negative, and gram-positive showed that 5 isolates belonged to Lacticaseibacillus rhamnosus, 12 to Bacillus megaterium, and 7 to Bacillus thuringiensis. In addition, one isolate each of Bacillus wiedmanni, Krasilnikviella muralis, Staphylococcus hominis, Bacillus stratosphericus, Staphylococcus epidermis, Bacillus pseudomycoides, and Bacillus subtilis was identified. The five Lacticaseibacillus rhamnosus isolates were further confirmed using BLAST (Table 2) and phylogenetic analysis (Fig. 2).

A maximum likelihood phylogenetic tree of the five selected bacterial isolates was constructed based on 16 S rRNA sequences using bootstrap and site coverage tools. The isolates were identified as strains of L. rhamnosus with 99–100% sequence similarity to the reference strains during a BLAST search. Sequences were aligned using MEGA 11 software, and phylogenetic analysis was performed with bootstrap validation. Comparative sequences of Lactobacillus acidophilus and Lactococcus lactis were retrieved from NCBI for confirmation.
Viability of LAB isolates against various gastrointestinal stressors, bile salt, and acidity
Growth curves of the five selected strains under various stressors (temperature, pH, H2O2, and ethanol) are shown in Fig. 3.

Growth curves of five selected L. rhamnosus isolates and LGG under various stress conditions (temperature, pH, H2O2, and ethanol). Optical density at 600 nm (OD₆₀₀) was monitored over 18 h under optimal growth conditions. Data represent the mean of two replicates, with error bars indicating the standard deviation.
The selected bacterial isolates of L. rhamnosus showed promising viability (nearly 7 log10 colony-forming units [CFU]/mL) and a survival rate of 98–99% against 0.3% bile salt and low-pH stressors (pH 3) (Fig. 4A–D). Isolate #3 exhibited significantly higher viability at 0 h than that at 6 h of incubation with bile salt, whereas isolates #2, #4, and #5 showed no significant differences (Fig. 4A). The survival rates (%) of isolates #4 and #5 did not differ significantly at 0.3% bile tolerance compared to the control, whereas that of isolate #3 was significantly lower (Fig. 4B).

Effect of bile and acidic conditions on the viability of five L. rhamnosus strains. (A) Viability (log10 CFU/mL) in 0.3% bile salt for 0–6 h. (B) Survival rate (%) with 0.3% bile salt. (C) Viability (log10 CFU/mL) at pH 3 for 0–3 h. (D) Survival rate (%) under acidic conditions (pH 3). LGG was used as the positive control. Data represent the average of three replicates, with error bars showing the standard error. In A and C; ##, P < 0.01; ###, P < 0.001 indicate significant differences compared to the initial conditions assessed using Student’s t-test. In B and D, *, P < 0.05; **, P < 0.01 indicate significant differences compared to LGG according to Student’s t-test. NS, not significant (P > 0.05).
Acid tolerance of the selected bacterial isolates was also evaluated at pH 3. All the tested isolates maintained a viability of nearly 7 log10 CFU/mL, with no significant differences observed among them (Fig. 4C). Similarly, the survival rates of the selected isolates were comparable to those of control L. rhamnosus GG (LGG) (Fig. 4D). These findings indicate that the selected L. rhamnosus isolates were highly resistant to harsh gastrointestinal conditions, making them promising candidates for safe and functional probiotic applications.
Analysis of antibacterial activity
The selected bacterial isolates demonstrated significant antibacterial activity, surpassing that of the controls (LGG and lactic acid), with inhibition zones ranging from 13.5 to 15.5 mm against Escherichia coli and Staphylococcus aureus. Among the isolates, isolate #4 exhibited the most pronounced antibacterial activity, as evidenced by its largest inhibition zone (Table 3).
Antibiotic resistance assessment
Bacterial isolates displayed varying levels of sensitivity to the antibiotics tested. The isolates were more susceptible to erythromycin and clindamycin than to benzylpenicillin, but exhibited resistance to tetracycline and vancomycin (Fig. 5). The inhibitory effects of erythromycin, clindamycin, and benzylpenicillin were significant, whereas those of tetracycline and vancomycin were ineffective in suppressing the growth of the isolates (Fig. 5).

Antibiotic susceptibility of five L. rhamnosus isolates to erythromycin (EM), clindamycin (CM), benzylpenicillin (PG), tetracycline (TC), and vancomycin (VA). Results are based on inhibition zones on MRS agar plates.
Estimation of antioxidant activities
The DPPH scavenging activity of the selected bacterial isolates ranged from 7.88 to 9.17% (Fig. 6A). Isolate #3 exhibited the highest activity, whereas isolate #4 exhibited the lowest. Comparisons with the control strain (LGG) revealed no significant differences, except for isolate #3, which displayed notably higher activity.

Antioxidant activity of selected bacterial isolates. (A) DPPH free radical-scavenging activity; (B) Oxygen radical absorbance capacity (ORAC); (C) ABTS free radical-scavenging activity. Controls included LGG, L-ascorbic acid (5.68 mM), and Trolox (20 µM). Data represent the mean of three replicates, with error bars indicating the standard error. **P < 0.01; ***, P < 0.001 according to the Student’s t-test. ##, P < 0.01; ###, P < 0.001 showed lower values than LGG. NS, not significant (P > 0.05).
The oxygen radical absorbance capacity (ORAC) assay (Fig. 6B), all selected isolates exhibited lower ORAC values than the control. Isolate #3 showed the highest ORAC value, whereas isolate #5 showed the lowest. Significant differences in ORAC values were observed among the isolates, with isolate #3 showing a notably stronger antioxidant capacity.
The ABTS assay results demonstrated varying degrees of free radical scavenging activity among the tested bacterial strains (Fig. 6C). The positive control (P/C) exhibited the highest antioxidant activity, significantly surpassing the reference strain LGG (P < 0.001), confirming its strong radical-scavenging potential. Among the test isolates, strain #3 showed a statistically significant increase in antioxidant activity compared to LGG (P < 0.01), suggesting its potential as a promising antioxidant-producing probiotic candidate. In contrast, strains #1, #2, #4, and #5 did not display significant differences in activity relative to LGG, indicating comparable but not enhanced antioxidant capacities.
In vitro auto-aggregation assay
The selected bacterial isolates demonstrated auto-aggregation capacities ranging from 20 to 25% after 5 h of incubation at 37 °C. All five isolates exhibited significantly lower auto-aggregation ability than that of the control. Strain #2 displayed the highest auto-aggregation capacity relative to the others (Fig. 7A).

Adhesion properties of the selected bacterial isolates. (A) Auto-aggregation capacity after incubation for 5 h at 37 °C. (B) Cell surface hydrophobicity. LGG was used as the positive control. The data represent the mean of three replicates. #, P < 0.05; ##, P < 0.01; ###, P < 0.001 indicate significant differences compared to LGG, assessed using Student’s t-test.
Assessment of cell-surface hydrophobicity revealed that the isolates exhibited significantly lower hydrophobicity, with values ranging between 20% and 30%, than the LGG strain (Fig. 7B). Strain #4 exhibited the highest hydrophobicity (~ 30%), whereas strain #1 exhibited the lowest hydrophobicity (~ 20%).
Safety assessment of selected bacterial isolates
Estimation of hemolytic activity revealed that none of the selected bacterial isolates exhibited either α-hemolytic or β-hemolytic activity on blood agar plates (Fig. 8). The lack of hemolytic activity suggests that these isolates are safe for potential applications.

Hemolytic activity of five selected bacterial isolates on blood agar plates. S. aureus was used as the positive control.
The BSH enzyme catalyzes the hydrolysis of amide bonds in conjugated bile acids, resulting in the release of free amino acids and the formation of hydrophobic deoxycholic acid or lithocholic acid. This hydrophobic acid causes the observed white precipitation around the colonies. In this study, the qualitative detection of BSH activity showed that all selected isolates demonstrated BSH activity, as evidenced by the formation of white precipitation around colonies on de Man, Rogosa, and Sharpe (MRS) agar plates supplemented with 0.5% (w/v) TDCA and 0.37 g/L CaCl2 (Fig. 9; Table 4). Furthermore, the quantitative analysis of BSH activity revealed that isolate #1 exhibited the highest BSH activity among the tested isolates, whereas isolate #5 showed the lowest activity in terms of free taurine liberation. However, no significant differences in BSH activity were observed between the selected isolates and the positive control (Lactobacillus acidophilus KCTC 3164), indicating comparable enzymatic activity levels (Fig. 10).

Detection of BSH activity through precipitation around colonies of bacterial isolates grown on MRS agar plates supplemented with 0.5% (w/v) TDCA and 0.37 g/L CaCl2. P/C: positive control, L. acidophilus KCTC 3164; N/C: negative control, LGG.

BSH activity of the selected bacterial isolates was expressed as the amount of taurine liberated from TDCA. Data are presented as the mean ± standard error. L. acidophilus KCTC 3164 was used as the positive control. ##, P < 0.01; ###, P < 0.001 indicate significant differences compared to L. acidophilus, as assessed using Student’s t-test.
Assessment of viability of bacterial isolates under oxgall and cholesterol
The selected bacterial isolates exhibited robust growth on MRS agar plates supplemented with oxgall and cholesterol (Table 5). Isolate #1 demonstrated the highest viability (96%), whereas isolates #3 and #5 exhibited slightly lower viabilities (90%). The viability of the control strain L. acidophilus KCTC 3164 was 89%. The superior survivability of the selected isolates under these conditions highlights their potential for gastrointestinal applications.
Discussion
In the current study, 35 isolates were characterized as BCP-positive, catalase-negative, or gram-positive. After 16 S rRNA sequencing of these 35 isolates, five samples were confirmed to be LAB (L. rhamnosus) strains, which correlates with previous studies22,23 in which only BCP-positive, catalase-negative, and gram-positive bacteria were found. Our results strongly suggest that ginseng sprouts grown on smart farms are a promising source of LAB.
For a strain to be used as a probiotic, it must meet several essential criteria, including survivability under extreme conditions (e.g., bile and acidic environments)24, ability to adhere to intestinal cells25, antimicrobial activity against pathogens24, and antioxidant activity24. In this study, the five isolated L. rhamnosus strains demonstrated the ability to survive under varying pH levels, temperatures, and in the presence of H2O2 and ethanol when compared to the positive and negative controls. In the gastrointestinal tract (GIT), LAB are exposed to fluctuating pH values, which vary across different regions of the small and large intestines due to the secretion of digestive juices, food digestion, and bile. Similarly, survival in the presence of ethanol and H2O2 indicates resilience to alcohol and oxidative stress, reflecting the antioxidant capabilities of the strains26. All five L. rhamnosus strains survived in 0.3% bile salt and pH 3 conditions, with no significant differences in viable cell counts compared to the control strain, LGG. This suggests that these strains are capable of tolerating extreme gastrointestinal conditions, aligning with prior studies that have reported high survival rates under similar conditions27,28,29. Our results strongly indicate that all five strains could successfully transit through the gastrointestinal tract and thus hold potential as probiotic strains capable of delivering health benefits to humans.
We also evaluated the antibacterial activity of the five selected isolates against both gram-positive (S. aureus) and gram-negative (E. coli) pathogens. Consistent with the findings of previous studies30,31, all five L. rhamnosus strains inhibited the growth of both S. aureus and E. coli. These findings suggest that all five strains may protect against pathogenic colonization in the GIT, offering potential health benefits.
The five isolated strains of L. rhamnosus showed potential free radical-scavenging activity comparable to that of the control substances, L-ascorbic acid, and Trolox in DPPH, ORAC and ABTS assays. These findings are consistent with those of previous studies32,33,34, suggesting that probiotics exert free radical-scavenging activity through different methods, benefiting host health after colonization of the human GIT. These findings further support the notion that all five strains may benefit human health by reducing oxidative stress through their electron- or hydrogen-donating abilities.
The five strains of L. rhamnosus also exhibited auto-aggregation capacity and cell surface hydrophobicity ranging from 28 to 35%. While these values were slightly lower than those of the control strain, L. rhamnosus GG, several factors, including cultivation time, temperature, pH, media composition, strain specificity, and solvent types, may contribute to the variation35; however, some studies did not show such a correlation between these two properties36. Additionally, previous reports have shown lower levels of aggregation and hydrophobicity in control and other probiotic strains37. In our study, the control strain, LGG, showed slightly lower aggregation and hydrophobicity. Despite these minor differences, all five strains exhibited promising survivability in gastrointestinal environments, with no significant differences in performance compared to the control strain. Together, these findings suggest that all five strains possess the properties necessary to adhere to intestinal epithelial cells38, thus facilitating their potential probiotic effects39,40.
Probiotic bacteria possess intrinsic genetic characteristics that confer susceptibility or resistance to a diverse range of antibiotics41. In this study, all five strains were susceptible to erythromycin, clindamycin, and benzylpenicillin, which is consistent with previous findings42. Furthermore, consistent with previous findings43, the selected strains showed resistance to tetracycline and vancomycin, which may be attributed to the presence of resistance genes or mutations within the strains. Given their ability to survive in the gastrointestinal tract, sensitivity to certain antibiotics, and the presence of resistance genes, these strains can be considered suitable for probiotic applications. Additionally, certain bacteria can produce toxins or enzymes that destroy red blood cells44,45. In this study, all five strains were γ-hemolytic, indicating the absence of hemolysis on blood agar plates. These findings suggest that these five strains are safe for consumption, which is consistent with previous studies46.
BSH activity has been hypothesized to play a key role in cholesterol reduction by probiotics47. Previous studies have demonstrated that different Lactobacillus strains can lower cholesterol levels in vitro through several pathways17,18. Specifically, the cholesterol-lowering activities of probiotics with BSH activity are thought to depend on the biotransformation reaction of bile salt deconjugation48,49. In this study, all five L. rhamnosus strains exhibited BSH activity and hydrolyzed conjugated bile salts on MRS plates supplemented with 0.5% (w/v) TDCA and 0.37 g/L CaCl2. In particular, strains #1 and #2 showed the highest amount of taurine deconjugation compared with the control strain L. acidophilus KCTC 3164, suggesting their elevated potential for cholesterol removal through the deconjugation process50. Hydrophobic deconjugated bile salts are reabsorbed to a lesser extent in the intestine, resulting in higher excretion in the feces. In the absence of probiotic strains, cholesterol precipitates because of the reduced solubility of deconjugated bile salts50,51. Probiotic supplementation accelerates the deconjugation process, resulting in cholesterol removal49. Therefore, the current findings strongly suggest that all five isolated strains have cholesterol-lowering potential and can be selected for in vivo studies to assess their efficacy further. Oxgall is a bile acid that exerts a highly toxic effect on L. rhamnosus growth, causing cell leakage or death52. In this study, the strains demonstrated robust growth under high bile salt concentrations, suggesting their resilience and suitability for probiotic applications.
Conclusion
This study provides foundational evidence supporting the probiotic potential of lactic acid bacteria (LAB) isolated from ginseng sprouts cultivated under smart-farming conditions. The selected strains exhibited essential probiotic traits and safety profiles, aligning with established criteria for functional probiotic candidates. Given their demonstrated bile salt hydrolase activity and ability to deconjugate bile acids, these strains may modulate host lipid metabolism, suggesting a mechanistic basis for their cholesterol-lowering potential. Moreover, their resilience under simulated gastrointestinal conditions reinforces their applicability in oral formulations. These findings collectively position ginseng-derived LAB as promising bioresources for the development of next-generation probiotics aimed at supporting gut and cardiovascular health. Further in vivo validation and clinical studies are recommended to substantiate these health claims.
Materials and methods
General procedure
Seeds of two Korean ginseng varieties, Yunpoong (V1) and Gumpoong (V2), were procured from the Rural Development Authority (RDA) and grown at the Smart Farm of the Korea Institute of Science and Technology (KIST), Gangneung, Korea. The seeds were hydroponically grown under controlled conditions (65–70 µMol light, 16 h/8 h photoperiod, 24 °C/18°C). Polyurethane sponge medium and RDA ginseng nutrient solution (EC 2.5, pH 6.8) were used. After four weeks, the seedlings were transferred to an LED chamber for light treatment. Ginseng sprouts grown under four different LED treatments (blue, infrared red, and white) were collected and stored at 4 °C. The bacterial strains used as controls, including Lacticaseibacillus rhamnosus GG KCTC 5033, Lactobacillus acidophilus KCTC 3164, Staphylococcus aureus KCTC 3881, and Escherichia coli KCTC 2593, were purchased from the Korean Collection for Type Culture (KCTC), Korea Research Institute of Bioscience and Biotechnology (KRIBB), Korea. LAB were grown in MRS broth (Oxoid, Basingstoke, UK).
Isolation of LAB from ginseng sprouts
To isolate LAB, ginseng sprouts were harvested and weighed, and 0.1% peptone solution was added to the samples according to their weight. Ten-fold serial dilutions were performed for both sonicated and blended samples with 0.1% peptone solution and plated on MRS agar plates. The plates were then incubated under aerobic conditions at 37 °C for 48 h. After incubation, bacterial colonies were isolated and separated on fresh MRS agar plates.
Identification and phylogenetic analysis of selected LAB isolates
The selected bacterial isolates were streaked on BCP agar plates for identification of LAB and then incubated aerobically at 37 °C for 18 h. Catalase and Gram staining were carried out53 to identify BCP-positive and gram-positive bacterial isolates. Molecular identification of all gram-positive bacterial isolates, including the five selected LAB isolates, was performed using 16 S rRNA sequencing, followed by phylogenetic analysis using MEGA1154. The Maximum Likelihood method and the Tamura–Nei model55 were used to assess the evolutionary history of the selected isolates.
Viability tests of selected LAB isolates against various gastrointestinal stressors
The effects of various stressors related to the GIT environment, including pH, temperature, H2O2, and ethanol, were analyzed using a UV spectrophotometer (λ = 600 nm) after 18 h of culture. Briefly, isolated bacterial cell cultures were prepared in fresh MRS broth with varying concentrations of ethanol (8%), H2O2 (1 and 10 mM), pH (4 and 5), and temperature (30 and 37 °C). The optical density (OD) at 600 nm (OD600) of each cell culture was measured and recorded after 18 h of incubation. This strain was used as the control. The growth rate was calculated using the following equation:
Growth rate (µ) = (:frac{0.3t}{{{log}_{10:}{N}_{t}:-:log}_{10:}{N}_{0}}) × (:frac{0.63}{{t}_{d}}).
Where t is the time interval, N₀ and Nₜ are the initial and final population sizes, td is the doubling time, and the constants 0.3 and 0.63 account for unit conversions and growth kinetics.
Bile tolerance assay
The bile tolerance of the five selected LAB strains was evaluated following the protocol established by Delgado et al.56, with minor modifications. The OD600 of the culture grown for 18 h in MRS broth at 37 °C was adjusted to < 1 using a UV/VIS Spectrophotometer (OPTIZEN POP, Mecasys Co. Ltd., Korea) with 1X phosphate-buffer saline (PBS) solution. The inoculation volume of each bacterial culture was determined based on the OD. After centrifugation (Gyrozen, Labogene, Korea) at 8,000 rpm for 5 min, cell pellets were collected, suspended in fresh MRS broth, and mixed with 0.3% bile salt MRS broth at a 1:9 ratio. After inoculation (0 h), 10-fold serial dilutions were done using 1X PBS and plated on fresh MRS agar plates with an initial 6 h incubation interval. After 48 h of incubation at 37 °C, bacterial colonies in MRS agar plates were counted and calculated as CFU/mL. The assay was independently performed thrice.
Acid tolerance assay
The acid tolerance capacity of the five selected bacterial isolates was evaluated following a previously described procedure, with some modifications56. Briefly, the cell culture was prepared in fresh MRS broth, and the OD600 of each cell culture was measured after incubation at 37 °C for 18 h. The inoculation volume was calculated based on the OD, and then centrifugation was performed at 8,000 rpm for 5 min at 4 °C. Two milliliters of fresh MRS broth was mixed with the pellet, and 1 mL of the solution was then mixed with MRS broth (pH 3; adjusted with 1 N HCl) separately at a ratio of 1:9. Subsequently, the solution was diluted ten-fold in 1X PBS, plated on fresh MRS agar plates, and incubated for 48 h at 37 °C. Viability was calculated by counting the bacterial colonies grown on MRS agar plates, and the survival rate (%) was calculated. LGG strain was used as a positive control, and the assay was performed three times independently.
Antibacterial activity against pathogenic bacterial strains
The antibacterial assay was performed as described by Yerlikaya et al.57, with some modifications. Briefly, bacterial cultures were grown in fresh MRS broth at 37 °C for 18 h and centrifuged at 8,000 rpm at 4 °C for 5 min. The supernatants were filtered using a 0.45 μm filter, sterilized, and 8 mm paper disks were immersed in the cell-free supernatant at 4 °C for 14 h. Subsequently, 100 µL suspension of each E. coli (1.70 × 108 CFU/mL) and S. aureus (2.63 × 108 CFU/mL) grown on NB agar plates for 18 h at 37 °C were inoculated onto NB agar plates. The paper disks were placed onto the pathogen-inoculated NB agar plates, and the inhibition zone was measured in mm after incubation at 24 h for 37 °C. Lacticaseibacillus rhamnosus GG and MRS broth with lactic acid (pH 4) were used as positive controls, whereas fresh MRS broth was used as negative control.
Antibiotic resistance assay
Antibiotic resistance of the selected LAB isolates was determined according to Georgieva et al.58, with some modifications. Five antibiotic strips (bioMérieux SA, France), including erythromycin, clindamycin, benzylpenicillin, tetracycline, and vancomycin, were placed on the surface of the agar plates using sterile forceps. The concentration of erythromycin, clindamycin, tetracycline, and vancomycin were 0.016–256 µg/mL, while that of benzylpenicillin was 0.002–32 µg/mL The agar plates were incubated at 37 °C for 48 h, and then antibiotic susceptibility or resistance of the bacterial isolates was evaluated based on the inhibition zone around the antibiotic strips. MRS broth was used as the culture medium.
Antioxidant activity
DPPH assay
DPPH radical scavenging activity was analyzed following the protocol established by Chen et al.59 with minor modifications. Briefly, the bacterial cultures were incubated for 16 h at 37 °C in fresh MRS broth and centrifuged at 12,000 rpm at 4 °C for 10 min. The pellets were washed three times using 1X PBS. DPPH (0.4 mM) solution was prepared using 99% methanol (Duksan Pure Chemical Co. Ltd, Korea) in a 4.0 mM DPPH solution (Thermo Fisher Scientific, USA). The bacterial suspension was added to the 0.4 mM DPPH solution at a 1:1 ratio and then incubated for 30 min at 37 °C. After incubation, the solution was centrifuged at 12,000 rpm for 5 min at 4 °C and the supernatant was collected for evaluation. The change in color at OD517 was recorded using a 96-well plate reader (BioTek, Synergy H1, USA). L-Ascorbic acid (5.68 mM; Sigma, USA) was used as a positive control. The LGG strain was used as the positive control. DPPH radical scavenging activity was calculated using the following equation:
$$DPPH;{text{radical}};{text{scavenging}};{text{activity}};(% ) = left( {1 – frac{{{text{Absorbance}}:{text{of}}:{text{sample}}}}{{{text{Absorbance}}:{text{of}}:{text{control}}}}} right)100$$
ORAC assay
The overall antioxidant capacity of the bacterial samples was evaluated following the method described by Li et al.60, with some modifications. Briefly, the five strains were incubated in fresh MRS broth for 16 h at 37 °C, followed by centrifugation at 8,000 rpm for 10 min. The cell pellets were then washed three times with 1X PBS and prepared as suspensions for further analysis. Next, a 78 nM fluorescein working solution was prepared from a 5 mM fluorescein stock solution (Sigma-Aldrich, USA) using 1X PBS. The AAPH reagent (221 mM) was prepared using 1X PBS. Trolox (20 µM) prepared from a 1 mM stock solution was used as a control. In a 96-well plate (black, F-bottom, Greiner Bio-one, Germany), 50 µL sample and 50 µL fluorescein working solution were mixed and incubated at 37 °C for 15 min in a microplate reader. Afterward, 25 µL of 221 mM AAPH reagent was added to the mixture, and fluorescence intensity was measured for 1 h at 10 min intervals using the microplate reader (BioTek) at an excitation of 485 nm and an emission wavelength of 538 nm. The fluorescence intensity of the five strains was expressed as 20 µM Trolox equivalents (TE). Antioxidant activity was calculated using the ORAC assay as TE (µM Trolox) using the following equation:
$${text{Relative}};ORAC;{text{value}} = left[ {(AUC_{{{text{sample}}}} – AUC_{{{text{blank}}}} )/{text{ }}left( {AUC_{{{text{Trolox}}}} – AUC_{{{text{blank}}}} } right)} right]$$
Where AUCsample, AUCblank, and AUCTrolox represent the areas under the curve for the sample, blank, and Trolox standard, respectively, and TE/g sample indicates Trolox equivalents per gram of sample.
ABTS free radical scavenging activity
ABTS (2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid)) scavenging activity was measured using a modified version of the method of Re et al.61. 7 mM ABTS (Thermo fisher scientific, Waltham, USA) solution and 2.45 mM potassium persulfate (Duksan, Seoul, Korea) were mixed in a 1:1 ratio and reacted in a darkroom for more than 16 h. The mixture was diluted with distilled water so that the absorbance at 734 nm was 0.700 ± 0.003. 200 µl of lactic acid protein extract (PE) and 600 µl of ABTS solution were mixed and reacted in a darkroom for 10 min. 150 µl of the supernatant was dispensed into a 96-well plate and the absorbance was measured at 734 nm. The negative control group used the same amount of distilled water as the sample, and the positive control group used L-ascorbic acid, an antioxidant. ABTS free radical scavenging activity was expressed as a percentage according to the following calculation formula.
$$ABTS;{text{radical}};{text{scavenging}};{text{activity}}(% ) = (Ac – As)/Ac times 100%$$
Where Ac is the absorbance of the control at 517 nm, and As is the absorbance of the sample.
In vitro auto-aggregation assay
The auto-aggregation capacity of the five strains was determined following a previously described method25. Briefly, the strains were grown in fresh MRS broth for 16 h, followed by centrifugation at 10,000 rpm for 10 min at 4 °C. The collected pellet was washed three times with 1X PBS, and the suspension was prepared with an adjustment of the OD600 value to 0.5–0.6. The cell suspension (4 mL) was vortexed for 15 s, incubated at 37 °C for 5 h, and 0.1 mL of the upper phase of the suspension was transferred to another tube to measure OD600. Auto-aggregation was calculated using the following equation:
$${text{Auto}} – {text{aggregation}}(% ) = left( {1 – frac{{{text{Absorbance}}:{text{at}}:5{text{}}:{text{hour}}}}{{{text{Absorbance}}:{text{at}}:{text{initial}}:{text{moment}}}}} right) times 100$$
Cell-surface hydrophobicity assay
The cell surface hydrophobicity was determined by following the established protocol62. Briefly, L. rhamnosus cell cultures grown in fresh MRS broth for 16 h at 37 °C were centrifuged at 10,000 rpm for 10 min at 4 °C. The pellets were washed three times using 1X PBS, and 3 mL of the cell suspension was prepared for further experiments. The OD600 was adjusted to 0.6–0.7, and 1 mL ethyl acetate was added to the cell suspension. The mixture was vortexed for 1 min and then kept at room temperature (22 °C) for 10 min to separate the phases. After 10 min, two different phases were observed, and the absorbance of the aqueous phase was measured at 600 nm.
Hemolysis assay
The hemolytic activity of the bacterial isolates was determined following the protocol described by Vasiee et al.63. Briefly, 5% (w/v) sheep blood (Thermo Fisher, USA) was added to the culture plates, incubated at 37 °C for 48 h, and the hemolytic activity of the bacterial strains was evaluated based on the red blood cells in the medium around the colonies. Clear zones around the colony indicated β-hemolysis and no zones around colony indicated γ-hemolysis.
Qualitative determination of BSH activity
The BSH activity of the bacterial isolates was determined as described by du Toit et al.64. Single colonies of selected bacterial isolates were streaked on MRS agar plates supplemented with 0.5% (w/v) TDCA and 0.37 g/L of CaCl2, and then the plates were parafilmed and incubated anaerobically at 37 °C in a GasPak EZ anaerobe container system (Oxoid™; Thermo Fisher). BSH activity was confirmed by the appearance of opaque halos of precipitated, deconjugated bile acids surrounding the colonies on the respective agar plates.
Quantitative determination of BSH activity
The quantitative BSH activity of the selected L. rhamnosus strains was determined by measuring the amino acid (taurine) liberated from conjugated bile salts (TDCA), following a previously described protocol65, with some modifications. L. rhamnosus cells were grown in fresh MRS broth for 18 h at 37 °C and centrifuged at 10,000 rpm at 4 °C for 10 min. The cell pellet was washed twice using 1X PBS, and the concentration was adjusted to an OD600 value of 0.5–0.6. Five milliliters of the cell suspension was again centrifuged at 10,000 rpm at 4 °C for 10 min, and the cell pellet was diluted with 1X PBS. Approximately 0.1 mL of pellet suspension was mixed with 1.8 mL 1X PBS and 0.1 mL conjugated bile salts (10 mM sodium taurocholate). The mixture was then incubated at 37 °C for 30 min, followed by termination of the enzymatic reaction by adding 15% (w/v) trichloroacetic acid (Duksan Pure Chemical Co., Ltd., Seoul, Korea) to the sample (0.5 mL). Subsequently, 0.2 mL distilled water and 0.5 mL ninhydrin reagent (Sigma-Aldrich, USA) were added, and the mixture was vortexed and boiled for 14 min in a thermomixer (Eppendorf Thermo Mixer C; Hamburg, Germany). After cooling, the absorbance was measured at 570 nm.
LAB isolates viability assay
Viability assays were performed according to the protocol of Choi and Chang66, with some modifications. MRS agar plates supplemented with 0.1 g/L cholesterol (Cholesterol-water soluble; Sigma-Aldrich) and 0.5% oxygen (Oxgall bile bovine; Sigma-Aldrich) were prepared. Bacterial cultures were sampled at 24 h intervals, and the cultured samples were serially diluted with 1X PBS and then plated on respective agar plates. After spreading, cultures were incubated anaerobically at 37 °C for 48 h using a GasPak EZ anaerobe container system (Oxoid™; Thermo Fisher).
Statistical analysis
GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA) was used to determine statistical differences between the treatment groups and relevant controls. Mean values were compared using one-way analysis of variance (ANOVA) (Ivanova et al., 2000). Analyses were performed using IBM SPSS Statistics 23 (IBM, Armonk, NY, USA). Data are presented as the mean ± standard deviation of three independent replicates.
Data availability
The accession numbers of our 5 bacterial isolates are given as, B3-0-3 (PQ276975), B5-1-8 (PQ276978), B6-0-1 (PQ276977), S4-0-39 (PQ276979) and S6-0-29 (Q276980). These accession numbers are provided by NCBI GenBank for 16 S rRNA sequences of our bacterial isolates.
References
-
FAO/WHO. Probiotics in food—Health and nutritional properties and guidelines for evaluation. Food Nutr. Pap 85, (2001).
-
Azad, M. A. K., Sarker, M., Li, T. & Yin, J. Probiotic species in the modulation of gut microbiota: An overview. Biomed. Res. Int. 2018, 9478630 (2018).
-
Stavropoulou, E. & Bezirtzoglou, E. Probiotics in medicine: A long debate. Front. Immunol. 11, 2192 (2020).
-
Stanton, C. et al. Market potential for probiotics. Am. J. Clin. Nutr. 73, 476S–483S (2001).
-
Khalid, K. An overview of lactic acid bacteria. Int. J. Biosci. 1, 1–13 (2011).
-
Walter, J. Ecological role of lactobacilli in the Gastrointestinal tract: implications for fundamental and biomedical research. Appl. Environ. Microbiol. 74, 4985–4996 (2008).
-
Hamayun, M. et al. Next-gen nutrition: challenges, innovations and opportunities in 3D food printing with probiotics. Future Foods. 11, 100620 (2025).
-
Niamah, A. K. et al. Phytophagous probiotic foods: exploring the intersection of characteristics, quality implications, health benefits, and market dynamics. Trends Food Sci. Technol. 154, 104795 (2024).
-
Ference, B. A. et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease. Eur. Heart J. 38, 2459–2772 (2017).
-
Jones, M. L., Tomaro-Duchesneau, C., Martoni, C. J. & Prakash, S. Cholesterol Lowering with bile salt hydrolase-active probiotic bacteria. Expert Opin. Biol. Ther. 13, 631–642 (2013).
-
Sridevi, N., Vishwe, P. & Prabhune, A. Hypocholesteremic effect of bile salt hydrolase from Lactobacillus buchneri ATCC 4005. Food Res. Int. 42, 516–520 (2009).
-
Pickin, D. M. et al. Cost effectiveness of Statin treatment related to coronary heart disease risk. Heart 82, 325–332 (1999).
-
Pereira, D. I. & Gibson, G. R. Effects of probiotics and prebiotics on serum lipid levels in humans. Crit. Rev. Biochem. Mol. Biol. 37, 259–281 (2002).
-
Liong, M. T. & Shah, N. P. Acid and bile tolerance and cholesterol removal ability of Lactobacilli strains. J. Dairy. Sci. 88, 55–66 (2005).
-
De Smet, I., De Boever, P. & Verstraete, W. Cholesterol Lowering in pigs through enhanced bacterial bile salt hydrolase activity. Br. J. Nutr. 79, 185–194 (1998).
-
Ichim, T. E., Patel, A. N. & Shafer, K. A. Effects of probiotic/digestive enzyme supplements on cholesterol and Microbiome. J. Transl Med. 14, 184 (2016).
-
Lye, H. S., Rusul, G. & Liong, M. T. Removal of cholesterol by Lactobacilli. J. Dairy. Sci. 93, 1383–1392 (2010).
-
Gilliland, S. E., Nelson, C. R. & Maxwell, C. Assimilation of cholesterol by Lactobacillus acidophilus. Appl. Environ. Microbiol. 49, 377–381 (1985).
-
Oh, J. Y. et al. Investigation of ginsenosides in Panax ginseng. J. Ginseng Res. 38, 270–277 (2014).
-
De Smet, I. et al. Bile salt hydrolase activity of L. plantarum 80 strains and cholesterol Lowering. Microb. Ecol. Health Dis. 7, 315–329 (1994).
-
Sadiq, N. B. et al. Influence of light quality on Lactobacillus in hydroponic ginseng. Chem. Biol. Technol. Agric. 12, 7 (2025).
-
Navarre, W. W. & Schneewind, O. Surface proteins of gram-positive bacteria and their targeting mechanisms. Microbiol. Mol. Biol. Rev. 63, 174–229 (1999).
-
Mokoena, M. P. Lactic acid bacteria and their bacteriocins. Molecules 22, 1255 (2017).
-
Mantzourani, I. et al. Probiotic potential of lactic acid bacteria from Kefir grains. Ann. Microbiol. 69, 1–13 (2019).
-
Polak-Berecka, M. A., Waśko, R. P. & Skrzypek, P. Sroka-Bartnicka, A. Cell surface components and adhesion of Lactobacillus rhamnosus. Antonie Van Leeuwenhoek. 106, 751–762 (2014).
-
Abouelkheir, M. et al. Ethanol-producing bacteria in NAFLD model and Metformin role. Pharmaceuticals 16, 658 (2023).
-
Pringsulaka, O., Rueangyotchanthana, K. & Suwannasai, N. In vitro screening of lactic acid bacteria. Food Microbiol. 23, 74–78 (2006).
-
Huang, Y. & Adams, M. C. Gastrointestinal tolerance of dairy Propionibacteria. Int. J. Food Microbiol. 159, 253–260 (2004).
-
Mahmoudi, I. et al. Adhesion properties of probiotic Lactobacillus from milk. J. Agric. Sci. Technol. 21, 587–600 (2019).
-
Kivanc, M., Yilmaz, M. & Çakir, E. Lactic acid bacteria from Boza and microbial activity. Turk. J. Biol. 35, 313–324 (2011).
-
Monteiro, C. R. A. V. et al. Antimicrobial activity and probiotic potential. Nutrients 11, 448 (2019).
-
Tang, W. et al. Antioxidant effects of L. plantarum MA2. Food Chem. 221, 1642–1649 (2017).
-
Yang, S. J. et al. Probiotic and antioxidant activities in fermented black Gamju. Foods 9, 1154 (2020).
-
Kim, K. T., Yang, S. J. & Paik, H. D. Probiotic potential of L. brevis KU15147 from Kimchi. Food Sci. Biotechnol. 30, 257–265 (2021).
-
Mackenzie, D. A. et al. Mucus-binding proteins and adhesion in L. reuteri. Microbiology 156, 3368–3378 (2010).
-
Chaffanel, F. et al. Surface proteins mediating S. salivarius adhesion. Appl. Microbiol. Biotechnol. 102, 2851–2865 (2018).
-
Krausova, G., Hyrslova, I. & Hynstova, I. Adhesion and hydrophobicity of potential probiotics. Fermentation 5, 100 (2019).
-
Duary, R. K., Rajput, Y. S., Batish, V. K. & Grover, S. Adhesion of probiotic Lactobacilli. Indian J. Med. Res. 134, 664–671 (2011).
-
Kos, B. et al. Adhesion and aggregation of L. acidophilus M92. J. Appl. Microbiol. 94, 981–987 (2003).
-
Boonaert, C. J. & Rouxhet, P. G. Surface chemistry of lactic acid bacteria. Appl. Environ. Microbiol. 66, 2548–2554 (2000).
-
Zheng, M. et al. Risk assessment of probiotic supplements and antibiotic resistance. Front. Microbiol. 8, 908 (2017).
-
Yerlikaya, O., Saygili, D. & Akpinar, A. Antimicrobial and antibiotic profiles of starter strains. Food Sci. Technol. 41, 418–425 (2021).
-
Gueimonde, M. et al. Antibiotic resistance in probiotics. Front. Microbiol. 4, 202 (2013).
-
Ali, S. et al. Recent advances in prebiotics: classification, mechanisms, and health applications. Future Foods. 11, 100680 (2025).
-
González-Rodríguez, N. et al. Hemolytic activity in Plesiomonas shigelloides. J. Food Prot. 70, 885–890 (2007).
-
Sathiyaseelan, A. et al. Probiotics from Korean fermented foods. Int. J. Mol. Sci. 23, 10062 (2022).
-
Lambert, J. M. et al. Bile salt hydrolase genes in L. plantarum WCFS1. Appl. Environ. Microbiol. 74, 4719–4726 (2008).
-
Ishimwe, N. et al. Cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 59, 94–105 (2015).
-
Öner, Ö., Aslim, B. & Aydaş, S. B. BSH gene analysis in probiotic strains. J. Mol. Microbiol. Biotechnol. 24, 12–18 (2014).
-
Begley, M., Hill, C. & Gahan, C. G. Bile salt hydrolase activity in probiotics. Appl. Environ. Microbiol. 72, 1729–1738 (2006).
-
Ahn, Y. T. et al. Deconjugation of bile salts by L. acidophilus. Int. Dairy. J. 13, 303–311 (2003).
-
Charteris, W. P. et al. Bile salts and antibiotic susceptibility in Lactobacillus. J. Food Prot. 63, 1369–1376 (2000).
-
Mohd Adnan, A. F. & Tan, I. K. Isolation of lactic acid bacteria from Malaysian foods. Bioresour Technol. 98, 1380–1385 (2007).
-
Tamura, K. & Nei, M. Estimation of nucleotide substitutions in mitochondrial DNA. Mol. Biol. Evol. 10, 512–526 (1993).
-
Tamura, K., Stecher, G. & Kumar, S. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027 (2021).
-
Delgado, S. et al. Probiotic screening of Lactobacillus species from GI tract. J. Food Sci. 72, M310–M315 (2007).
-
Yerlikaya, O., Saygili, D. & Akpinar, A. Antimicrobial profiles of yogurt starter strains. Food Sci. Technol. 41, 418–425 (2021).
-
Georgieva, R. et al. Antibiotic susceptibility of probiotic strains. Biotechnol. Biotechnol. Equip. 29, 84–89 (2015).
-
Chen, P. et al. Screening for probiotics with α-glucosidase Inhibition. Food Control. 35, 65–72 (2014).
-
Li, C. et al. Bioactivities of polysaccharides from Sargassum pallidum. Carbohydr. Polym. 155, 261–270 (2017).
-
Re, R. et al. Antioxidant activity via improved ABTS assay. Free Radic Biol. Med. 26, 1231–1237 (1999).
-
Abbasiliasi, S. et al. In vitro assessment of Pediococcus acidilactici Kp10. BMC Microbiol. 12, 1–11 (2017).
-
Vasiee, A. et al. Probiotic characterization of Pediococcus strains. J. Biosci. Bioeng. 130, 471–479 (2020).
-
du Toit, M. et al. Probiotic Lactobacilli and serum cholesterol in minipigs. Int. J. Food Microbiol. 40, 93–104 (1998).
-
Tanaka, H. et al. Bile salt hydrolase of B. longum. Appl. Environ. Microbiol. 66, 2502–2512 (2000).
-
Choi, E. & Chang, H. Cholesterol-lowering effects of L. plantarum EM. LWT – Food Sci. Technol. 62, 210–217 (2015).
Acknowledgements
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (Nos. 2020R1C1C1009948 and RS-2023-00213290).
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ahmed, E., Lee, JE., Hamayun, M. et al. Probiotic potential of ginseng derived Lacticaseibacillus rhamnosus strains with hypocholesterolemic, antioxidant and antibacterial activities. Sci Rep 15, 29222 (2025). https://doi.org/10.1038/s41598-025-13364-x
-
Received:
-
Accepted:
-
Published:
-
DOI: https://doi.org/10.1038/s41598-025-13364-x